Canopy Reflectance Modeling of Aquatic Vegetation for Algorithm Development: Global Sensitivity Analysis

 ,
, 
 ,
,
Abstract
:
1. Introduction
2. Methodology
2.1. AVRT Model
2.2. EFAST Method
3. Data
3.1. Input Parameters
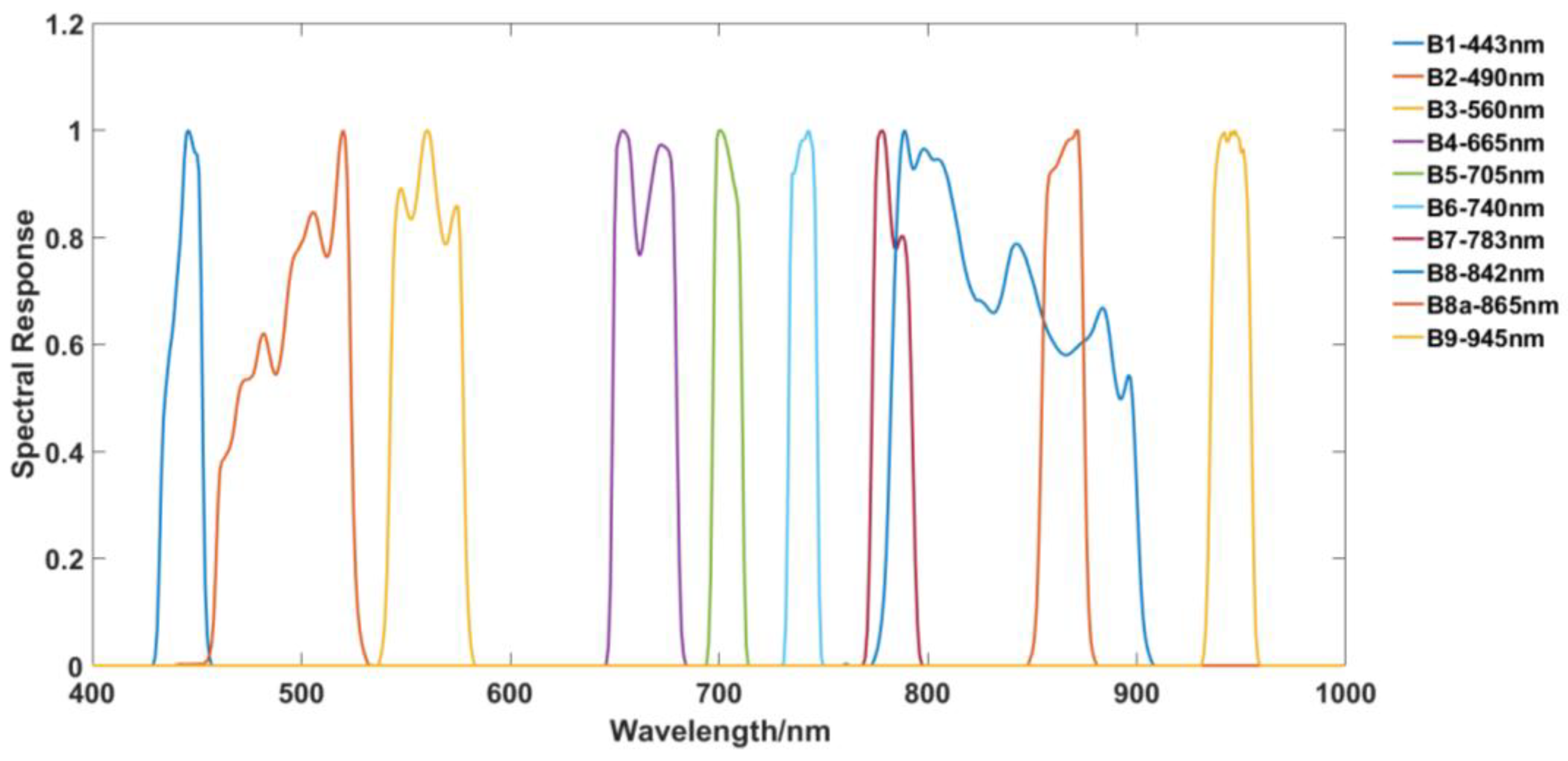
3.2. Sentinel–2A
4. Results
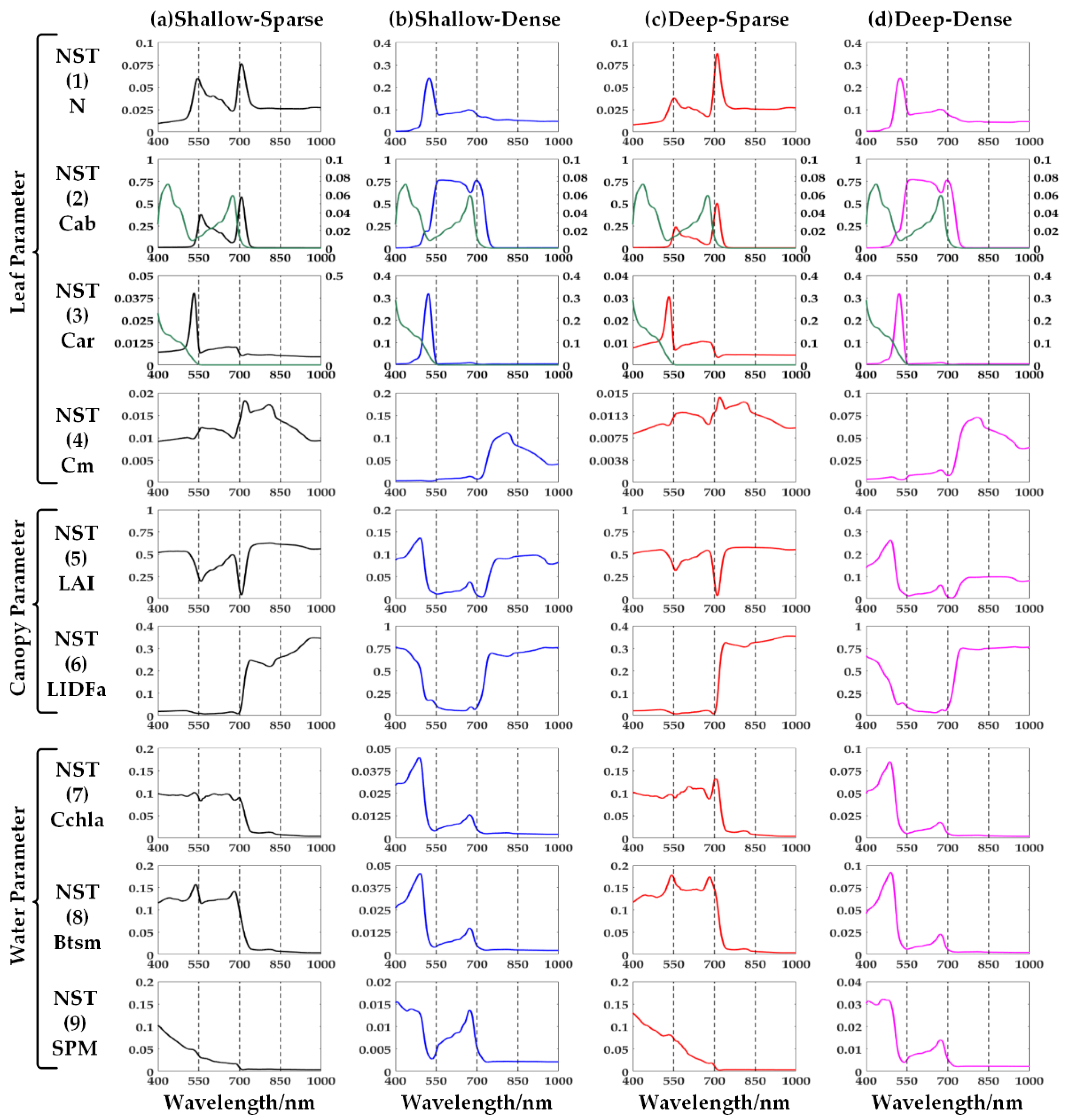
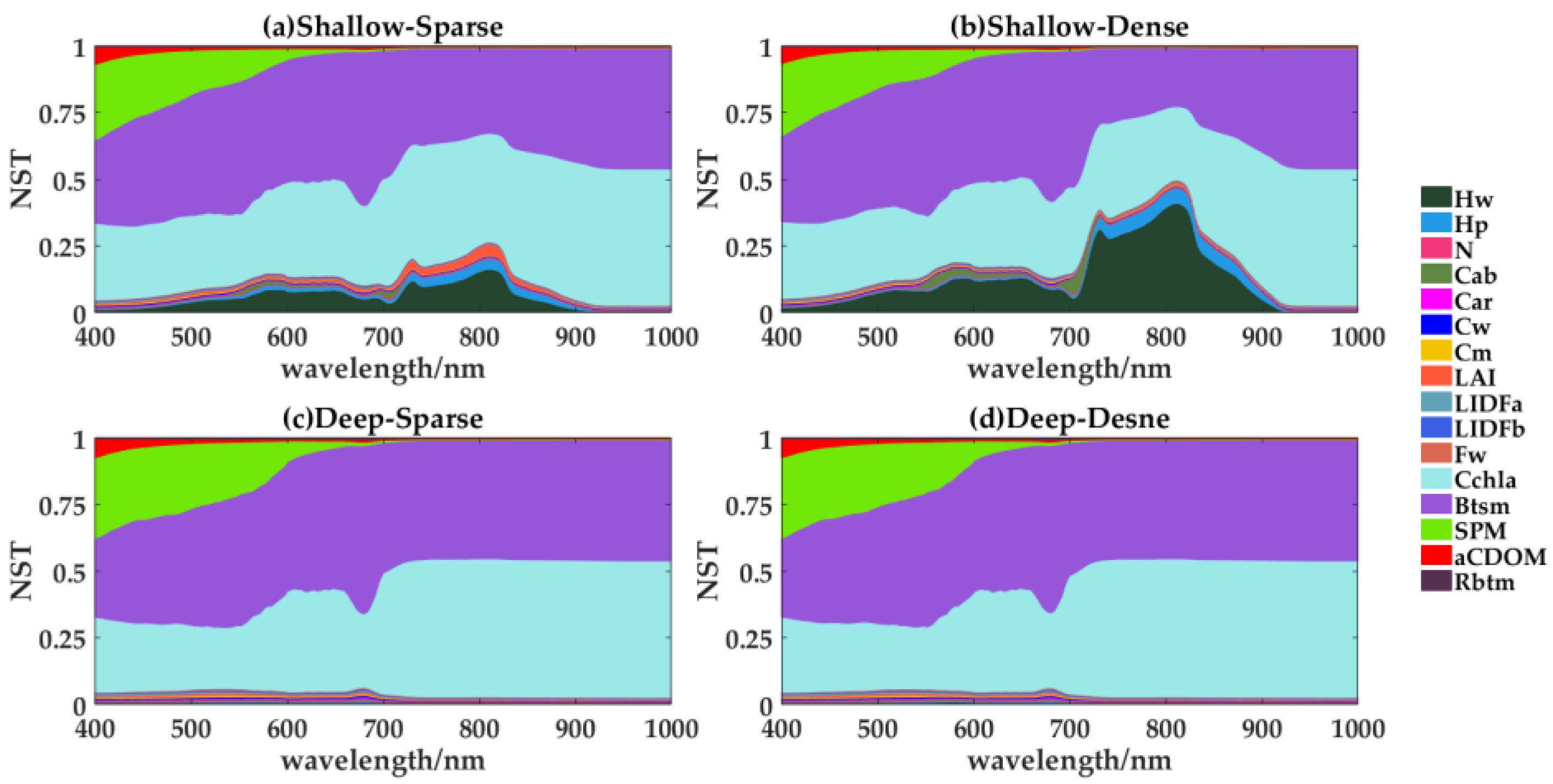
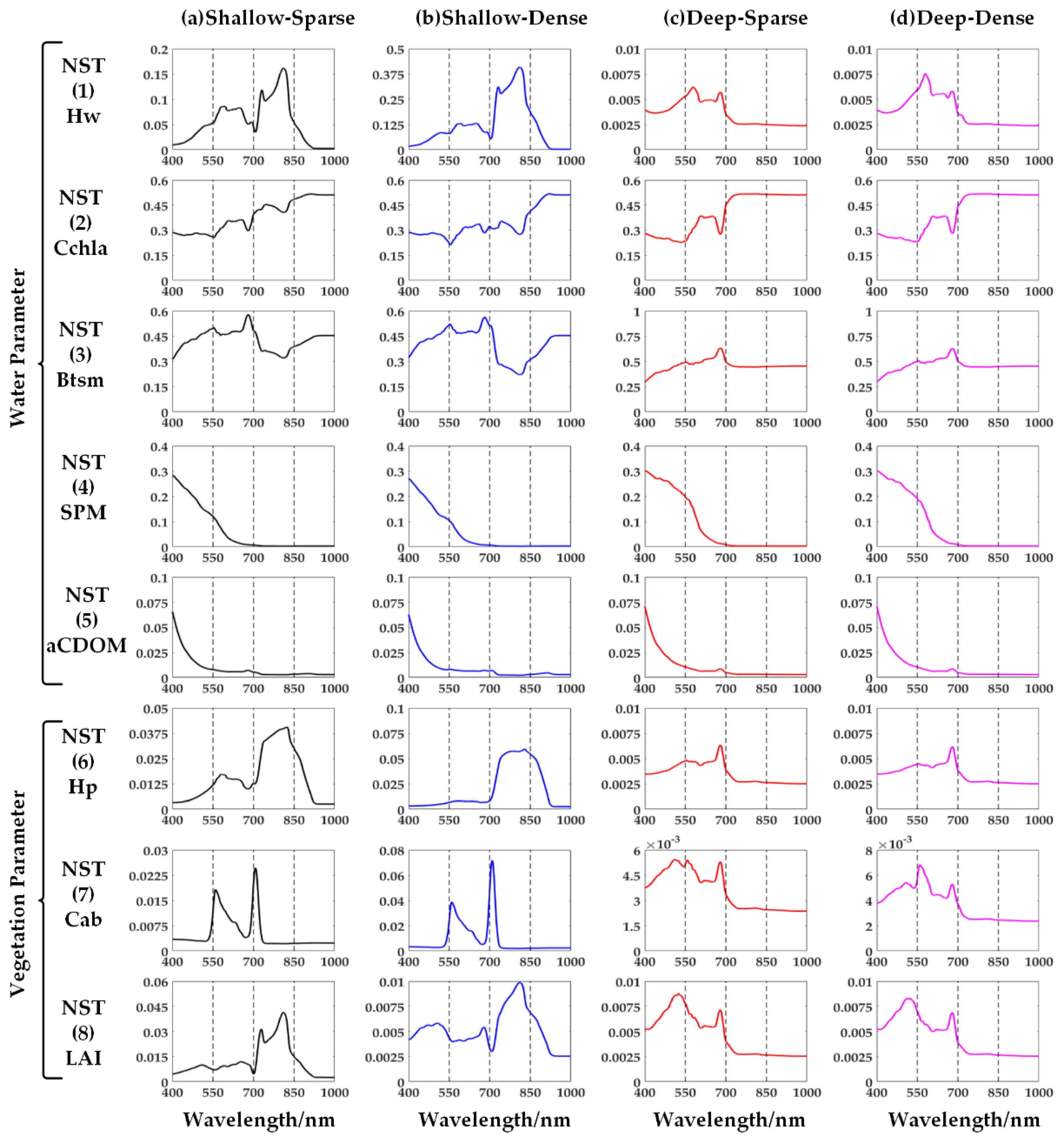
4.1. GSA to Reflectance in Different Cases
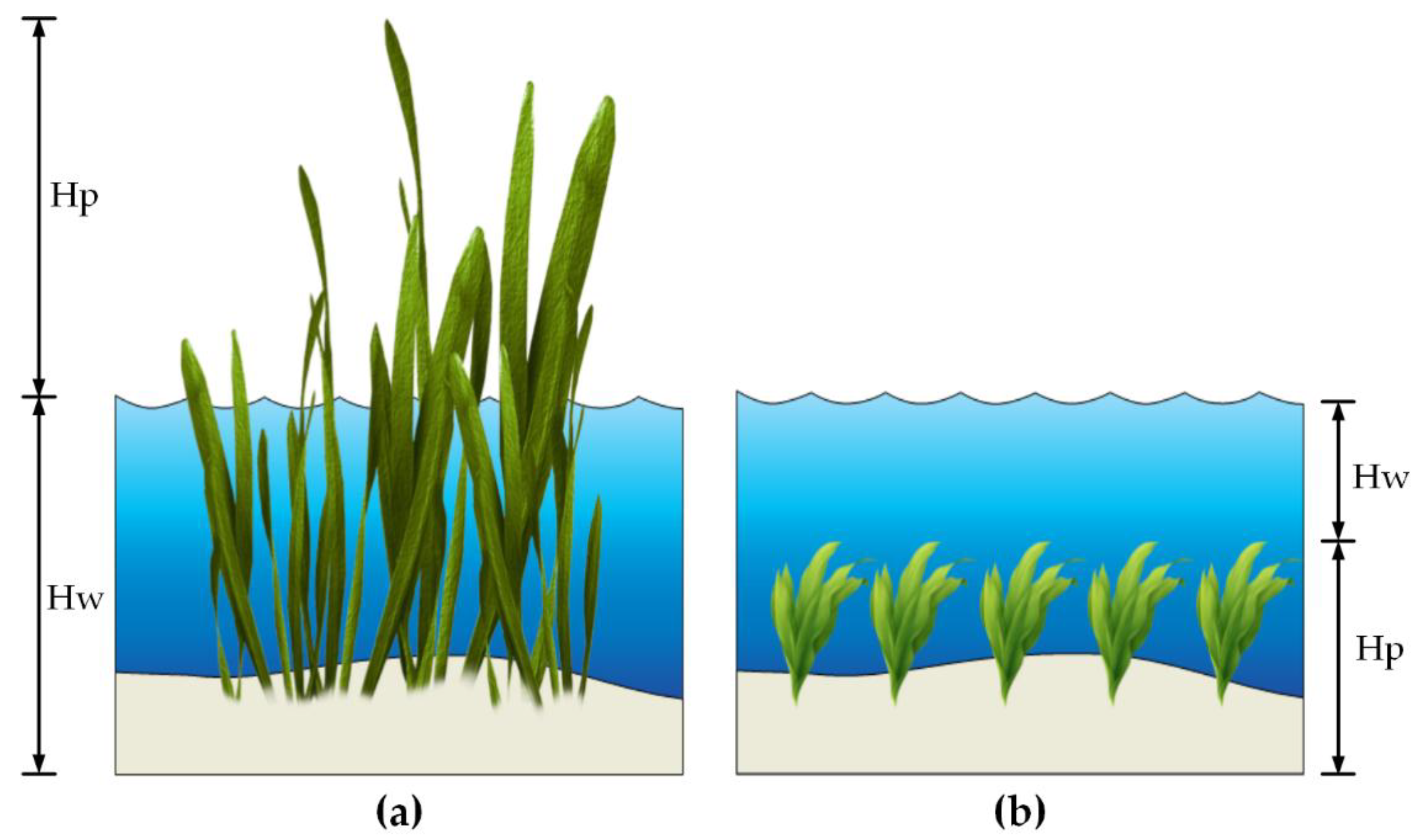
4.1.1. Emergent Vegetation
4.1.2. Submerged Vegetation
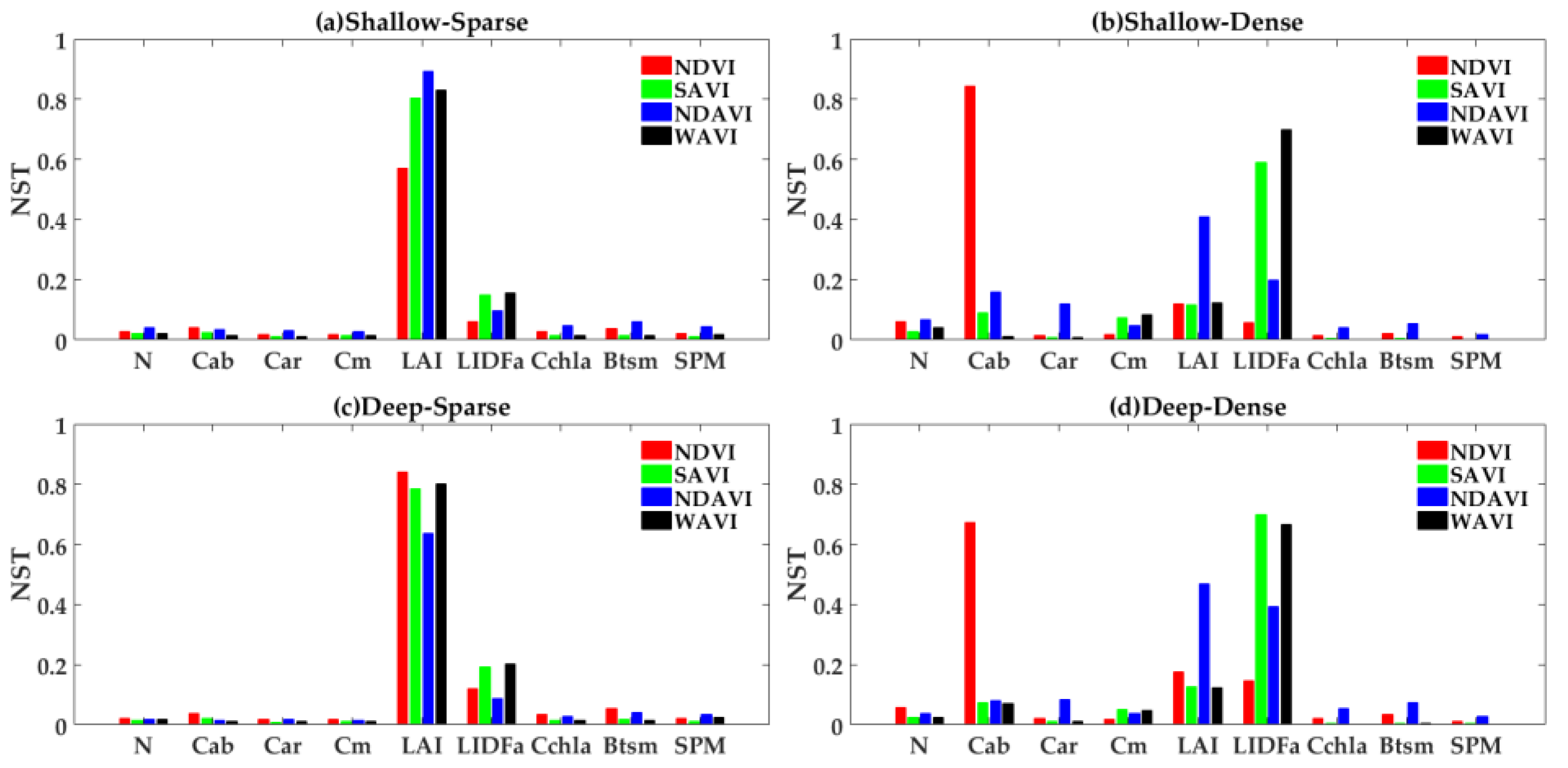
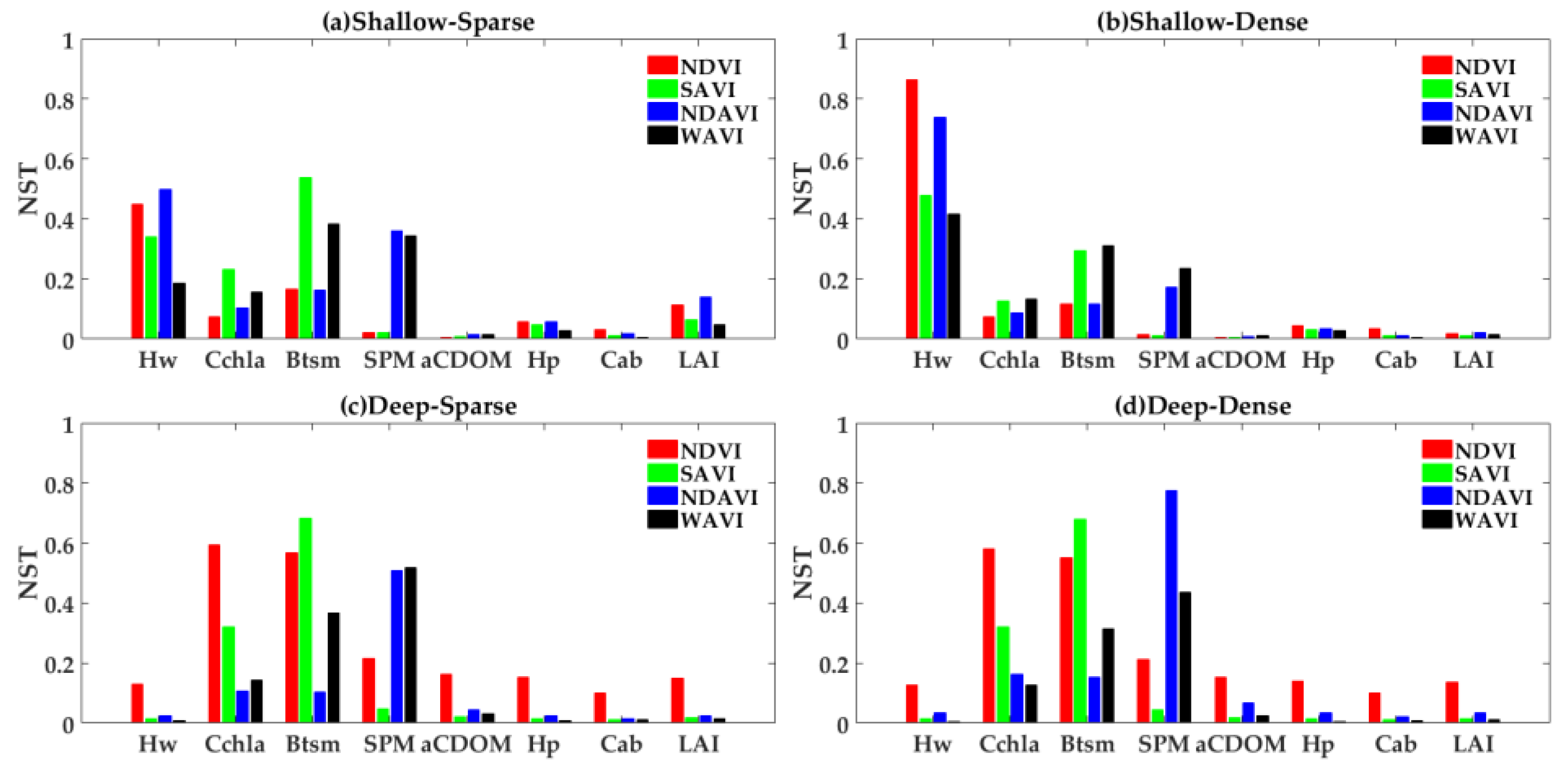
4.2. GSA to VIs in Different Cases
4.2.1. Emergent Vegetation
4.2.2. Submerged Vegetation
5. Discussion
6. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
Appendix A
References
- Ginn, B.K. Distribution and limnological drivers of submerged aquatic plant communities in lake Simcoe (Ontario, Canada): Utility of macrophytes as bioindicators of lake trophic status. J. Great Lakes Res. 2011, 37, 83–89. [Google Scholar] [CrossRef]
- Villa, P.; Pinardi, M.; Tóth, V.R.; Hunter, P.D.; Bolpagni, R.; Bresciani, M. Remote sensing of macrophyte morphological traits: Implications for the management of shallow lakes. J. Limnol. 2017, 76. [Google Scholar] [CrossRef]
- Cook, C.D.K. Aquatic Plant Book; Quarterly Review of Biology; SPB Academic Publishing: Amsterdam, The Netherlands, 1996. [Google Scholar]
- Hunter, P.D.; Gilvear, D.J.; Tyler, A.N.; Willby, N.J.; Kelly, A. Mapping macrophytic vegetation in shallow lakes using the compact airborne spectrographic imager (casi). Aquat. Conserv. Mar. Freshw. Ecosyst. 2010, 20, 717–727. [Google Scholar] [CrossRef]
- Vis, C.; Hudon, C.; Carignan, R. An evaluation of approaches used to determine the distribution and biomass of emergent and submerged aquatic macrophytes over large spatial scales. Aquat. Bot. 2003, 77, 187–201. [Google Scholar] [CrossRef]
- Fritz, C.; Dörnhöfer, K.; Schneider, T.; Geist, J.; Oppelt, N. Mapping submerged aquatic vegetation using rapideye satellite data: The example of Lake Kummerow (Germany). Water 2017, 9, 510. [Google Scholar] [CrossRef]
- Husson, E.; Reese, H.; Ecke, F. Combining spectral data and a dsm from uas-images for improved classification of non-submerged aquatic vegetation. Remote Sens. 2017, 9, 247. [Google Scholar] [CrossRef]
- Luo, J.; Duan, H.; Ma, R.; Jin, X.; Li, F.; Hu, W.; Shi, K.; Huang, W. Mapping species of submerged aquatic vegetation with multi-seasonal satellite images and considering life history information. Int. J. Appl. Earth Obs. Geoinf. 2017, 57, 154–165. [Google Scholar] [CrossRef]
- Zhang, X. On the estimation of biomass of submerged vegetation using landsat thematic mapper (tm) imagery: A case study of the Honghu lake, pr China. Int. J. Remote Sens. 1998, 19, 11–20. [Google Scholar] [CrossRef]
- Byrd, K.B.; O’Connell, J.L.; Tommaso, S.D.; Kelly, M. Evaluation of sensor types and environmental controls on mapping biomass of coastal marsh emergent vegetation. Remote Sens. Environ. 2014, 149, 166–180. [Google Scholar] [CrossRef]
- Gao, Y.; Gao, J.; Wang, J.; Wang, S.; Li, Q.; Zhai, S.; Zhou, Y. Estimating the biomass of unevenly distributed aquatic vegetation in a lake using the normalized water-adjusted vegetation index and scale transformation method. Sci. Total Environ. 2017, 601, 998. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.; Yoneda, M.; Susaki, J.; Tamura, M.; Ishikawa, K.; Yamashiki, Y. A satellite-based assessment of the distribution and biomass of submerged aquatic vegetation in the optically shallow basin of Lake Biwa. Remote Sens. 2017, 9, 966. [Google Scholar] [CrossRef]
- Dierssen, H.M.; Zimmerman, R.C.; Leathers, R.A.; Downes, T.V.; Davis, C.O. Ocean color remote sensing of seagrass and bathymetry in the bahamas banks by high-resolution airborne imagery. Limnol. Oceanogr. 2003, 48, 444–455. [Google Scholar] [CrossRef]
- Trilla, G.G.; Pratolongo, P.; Beget, M.E.; Kandus, P.; Marcovecchio, J.; Bella, C.D. Relating biophysical parameters of coastal marshes to hyperspectral reflectance data in the Bahia blanca estuary, Argentina. J. Coast. Res. 2013, 29, 231–238. [Google Scholar] [CrossRef]
- Everitt, J.H.; Yang, C.; Summy, K.R.; Owens, C.S.; Glomski, L.M.; Smart, R.M. Using in situ hyperspectral reflectance data to distinguish nine aquatic plant species. Geocarto Int. 2011, 26, 459–473. [Google Scholar] [CrossRef]
- Pinnel, N.; Heege, T.; Zimmerman, S. Spectral discrimination of submerged macrophytes in lakes using hyperspectral remote sensing data. In Proceedings of the Ocean Optics XVII, Fremantle, Australia, 25–29 October 2004; pp. 1–16. [Google Scholar]
- Han, L.; Rundquist, D.C. The spectral responses of ceratophyllum demersum at varying depths in an experimental tank. Int. J. Remote Sens. 2003, 24, 859–864. [Google Scholar] [CrossRef]
- Ackleson, S.G.; Klemas, V. Remote sensing of submerged aquatic vegetation in lower chesapeake bay: A comparison of landsat mss to tm imagery. Remote Sens. Environ. 1987, 22, 235–248. [Google Scholar] [CrossRef]
- Botha, E.J.; Brando, V.E.; Anstee, J.M.; Dekker, A.G.; Sagar, S. Increased spectral resolution enhances coral detection under varying water conditions. Remote Sens. Environ. 2013, 131, 247–261. [Google Scholar] [CrossRef]
- Hedley, J. A three-dimensional radiative transfer model for shallow water environments. Opt. Express 2008, 16, 21887. [Google Scholar] [CrossRef] [PubMed]
- Hedley, J.; Roelfsema, C.; Koetz, B.; Phinn, S. Capability of the sentinel 2 mission for tropical coral reef mapping and coral bleaching detection. Remote Sens. Environ. 2012, 120, 145–155. [Google Scholar] [CrossRef]
- Sakuno, Y.; Kunii, H. Estimation of growth area of aquatic macrophytes expanding spontaneously in lake Shinji using aster data. Int. J. Geosci. 2013, 4, 1–5. [Google Scholar] [CrossRef]
- Suits, G.H. A versatile directional reflectance model for natural water bodies, submerged objects, and moist beach sands. Remote Sens. Environ. 1984, 16, 143–156. [Google Scholar] [CrossRef]
- Turpie, K.R. Enhancement of a Canopy Reflectance Model for Understanding the Specular and Spectral Effects of an Aquatic Background in an Inundated Tidal Marsh; University of Maryland: College Park, MD, USA, 2012. [Google Scholar]
- Zimmerman, R.C. A biooptical model of irradiance distribution and photosynthesis in seagrass canopies. Limnol. Oceanogr. 2003, 48, 568–585. [Google Scholar] [CrossRef]
- Zhou, G.; Niu, C.; Xu, W.; Yang, W.; Wang, J.; Zhao, H. Canopy modeling of aquatic vegetation: A radiative transfer approach. Remote Sens. Environ. 2015, 163, 186–205. [Google Scholar] [CrossRef]
- Saltelli, A.; Tarantola, S.; Chan, P.S. A quantitative model-independent method for global sensitivity analysis of model output. Technometrics 1999, 41, 39–56. [Google Scholar] [CrossRef]
- Saltelli, A.; Ratto, M.; Andres, T.; Campolongo, F.; Cariboni, J.; Gatelli, D.; Saisana, M.; Tarantola, S. Global Sensitivity Analysis: The Primer; John Wiley & Sons: Hoboken, NJ, USA, 2008; p. 304. [Google Scholar]
- Nossent, J.; Elsen, P.; Bauwens, W. Sobol’ Sensitivity Analysis of a Complex Environmental Model; Elsevier Science Publishers B. V.: Amsterdam, Netherlands, 2011; pp. 1515–1525. [Google Scholar]
- Saltelli, A.; Marivoet, J. Non-parametric statistics in sensitivity analysis for model output: A comparison of selected techniques. Reliab. Eng. Syst. Saf. 1990, 28, 229–253. [Google Scholar] [CrossRef]
- Baroni, G.; Tarantola, S. A general probabilistic framework for uncertainty and global sensitivity analysis of deterministic models: A hydrological case study. Environ. Model. Softw. 2014, 51, 26–34. [Google Scholar] [CrossRef]
- Pianosi, F.; Wagener, T. A simple and efficient method for global sensitivity analysis based oncumulative distribution functions. Environ. Model. Softw. 2015, 67, 1–11. [Google Scholar] [CrossRef]
- Bounceur, N.; Crucifix, M.; Wilkinson, R.D. Global sensitivity analysis of the climate-vegetation system to astronomical forcing: An emulator-based approach. Earth Syst. Dyn. 2015, 5, 901–943. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, K.S. An information entropy-based sensitivity analysis of radar sensing of rough surface. Remote Sens. 2018, 10, 286. [Google Scholar] [CrossRef]
- Xiao, Y.; Zhou, D.; Gong, H.; Zhao, W. Sensitivity of canopy reflectance to biochemical and biophysical variables. J. Remote Sens. 2015, 19, 368–374. [Google Scholar]
- Mousivand, A.; Menenti, M.; Gorte, B.; Verhoef, W. Global sensitivity analysis of the spectral radiance of a soil–vegetation system. Remote Sens. Environ. 2014, 145, 131–144. [Google Scholar] [CrossRef]
- Verrelst, J.; Sabater, N.; Rivera, J.; Muñozmarí, J.; Vicent, J.; Campsvalls, G.; Moreno, J. Emulation of leaf, canopy and atmosphere radiative transfer models for fast global sensitivity analysis. Remote Sens. 2016, 8, 673. [Google Scholar] [CrossRef]
- Villa, P.; Mousivand, A.; Bresciani, M. Aquatic vegetation indices assessment through radiative transfer modeling and linear mixture simulation. Int. J. Appl. Earth Obs. Geoinf. 2014, 30, 113–127. [Google Scholar] [CrossRef]
- Verrelst, J.; Rivera, J.P.; Tol, C.V.D.; Magnani, F.; Mohammed, G.; Moreno, J. Global sensitivity analysis of the scope model: What drives simulated canopy-leaving sun-induced fluorescence? Remote Sens. Environ. 2015, 166, 8–21. [Google Scholar] [CrossRef]
- Jacquemoud, S.; Baret, F. Prospect: A model of leaf optical properties spectra. Remote Sens. Environ. 1990, 34, 75–91. [Google Scholar] [CrossRef]
- Bricaud, A.; Babin, M.; Morel, A.E.; Claustre, H.E. Variability in the chlorophyll-specific absorption coefficients of natural phytoplankton: Analysis and parameterization. J. Geophys. Res. 1995, 100, 13321–13332. [Google Scholar] [CrossRef]
- Buiteveld, A.H.; Hakvoort, J.H.M.; Donze, M. Optical properties of pure water. In Proceedings of the SPIE—The International Society for Optical Engineering, Bergen, Norway, 13–15 June 1994; pp. 368–373. [Google Scholar]
- Hr, P.A.S.; Markager, S. Parameterization of the chlorophyll a-specific in vivo light absorption coefficient covering estuarine, coastal and oceanic waters. Int. J. Remote Sens. 2004, 25, 5117–5130. [Google Scholar]
- Palmer, K.F.; Williams, D. Optical properties of water in the near infrared. J. Opt. Soc. Am. 1974, 64, 1107–1110. [Google Scholar] [CrossRef]
- Smith, R.C.; Baker, K.S. Optical properties of the clearest natural waters (200–800 nm). Appl. Opt. 1981, 20, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Volpe, V.; Silvestri, S.; Marani, M. Remote sensing retrieval of suspended sediment concentration in shallow waters. Remote Sens. Environ. 2011, 115, 44–54. [Google Scholar] [CrossRef]
- Cox, C.; Munk, W. Measurement of the roughness of the sea surface from photographs of the sun’s glitter. J. Opt. Soc. Am. 1954, 44, 838–850. [Google Scholar] [CrossRef]
- Cooper, K.; Smith, J.A.; Pitts, D. Reflectance of a vegetation canopy using the adding method. Appl. Opt. 1982, 21, 4112–4118. [Google Scholar] [CrossRef] [PubMed]
- Verhoef, W. Light scattering by leaf layers with application to canopy reflectance modeling: The sail model. Remote Sens. Environ. 1984, 16, 125–141. [Google Scholar] [CrossRef]
- Babin, M.; Stramski, D.; Ferrari, G.M.; Claustre, H.; Bricaud, A.; Obolensky, G.; Hoepffner, N. Variations in the light absorption coefficients of phytoplankton, nonalgal particles, and dissolved organic matter in coastal waters around europe. J. Geophys. Res. Oceans 2003, 108. [Google Scholar] [CrossRef]
- Lee, Z.; Carder, K.L.; Mobley, C.D.; Steward, R.G.; Patch, J.S. Hyperspectral remote sensing for shallow waters. I. A semianalytical model. Appl. Opt. 1998, 37, 6329. [Google Scholar] [CrossRef] [PubMed]
- Dimitris, S.; Heiko, B.; Olga, S.; András, Z.; Tóth, V.R. Evaluating sentinel-2 for lakeshore habitat mapping based on airborne hyperspectral data. Sensors 2015, 15, 22956–22969. [Google Scholar]
- Dall’Olmo, G.; Gitelson, A.A. Effect of bio-optical parameter variability on the remote estimation of chlorophyll-a concentration in turbid productive waters: Experimental results. Appl. Opt. 2005, 44, 412–422. [Google Scholar] [CrossRef] [PubMed]
- Le, C.; Li, Y.; Zha, Y.; Sun, D.; Huang, C.; Lu, H. A four-band semi-analytical model for estimating chlorophyll a in highly turbid lakes: The case of Taihu Lake, china. Remote Sens. Environ. 2009, 113, 1175–1182. [Google Scholar] [CrossRef]
- Huete, A.; Justice, C.; Liu, H. Development of vegetation and soil indices for modis-eos. Remote Sens. Environ. 1994, 49, 224–234. [Google Scholar] [CrossRef]
- Bannari, A.; Morin, D.; Bonn, F.; Huete, A.R. A review of vegetation indices. Remote Sens. Rev. 1995, 13, 95–120. [Google Scholar] [CrossRef]
- Huete, A.R. A soil-adjusted vegetation index (savi). Remote Sens. Environ. 1988, 25, 295–309. [Google Scholar] [CrossRef]
- Huete, A.R.; Liu, H.Q.; Batchily, K.; Leeuwen, W.V. A comparison of vegetation indices over a global set of tm images for eos-modis. Remote Sens. Environ. 1997, 59, 440–451. [Google Scholar] [CrossRef]
- Silva, T.S.; Costa, M.P.; Melack, J.M.; Novo, E.M. Remote sensing of aquatic vegetation: Theory and applications. Environ. Monit. Assess. 2008, 140, 131–145. [Google Scholar] [CrossRef] [PubMed]
- Duarte-Carvajalino, J.M. Sensitivity Analysis of the Water Column Effect on the Remote Sensing Reflectance, the Shallow Waters Case. Master’s Thesis, University of Puerto Rico, Mayaguez, Puerto Rico, 2003. [Google Scholar]
- Saltelli, A.; Annoni, P. Sensitivity Analysis; Wiley: Hoboken, NJ, USA, 2000; pp. 1298–1301. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Description | Unit | Range |
|---|---|---|---|
| Hw 1 | Water depth for emergent vegetation The height of the upper water layer for submerged vegetation | m | 0.1–1.0 (Shallow) |
| 1.0–2.0 (Deep) | |||
| Hp | The height of the upper vegetation layer for emergent vegetation Plant height for submerged vegetation | m | 0.1–1.0 |
| N | Leaf structure parameter | — | 1–2.5 |
| Cab | Concentration of chlorophyll a + b, in leaves | µg/cm2 | 10–80 |
| Car | Concentration of carotenoid, in leaves | µg/cm2 | 10–30 |
| Cw | Concentration of equivalent water thickness, in leaves | µg/cm2 | 0.004–0.04 |
| Cm | Concentration of dry matter, in leaves | µg/cm2 | 0.002–0.01 |
| LAI | Leaf area index | — | 0–2 (Sparse) |
| 2–4 (Dense) | |||
| LIDFa | Leaf inclination distribution function parameter a (which represents the average leaf slope) | — | −1–1 |
| LIDFb | Leaf inclination distribution function parameter b (which represents the distribution’s bimodality) | — | −1–1 |
| Fw | volume fraction of water in a layer | — | 0.7–1 |
| Cchla | Concentration of chlorophyll a, in water | mg/m3 | 0–80 |
| Btsm | Coefficient to calculate scattering of total suspended matter | — | 1–5 |
| SPM | Concentration of suspended matter, in water | g/m3 | 0–80 |
| aCDOM | Absorption coefficient of CDOM at 375 nm | /m | 0.5–3 |
| Rbtm | Reflectance of bottom | — | 0–1 |
| SZA | Sun zenith angle in air | degree | 30 |
| VZA | Viewing zenith angle in air | degree | 0 |
| RA | Relative azimuth angle in air | degree | 0 |
| VI | Description | Formula |
|---|---|---|
| NDVI | Normalized Difference Vegetation Index | |
| SAVI | Soil Adjusted Vegetation Index | |
| NDAVI | Normalized Difference Aquatic Vegetation Index | |
| WAVI | Water Adjusted Vegetation Index |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, G.; Ma, Z.; Sathyendranath, S.; Platt, T.; Jiang, C.; Sun, K. Canopy Reflectance Modeling of Aquatic Vegetation for Algorithm Development: Global Sensitivity Analysis. Remote Sens. 2018, 10, 837. https://doi.org/10.3390/rs10060837
Zhou G, Ma Z, Sathyendranath S, Platt T, Jiang C, Sun K. Canopy Reflectance Modeling of Aquatic Vegetation for Algorithm Development: Global Sensitivity Analysis. Remote Sensing. 2018; 10(6):837. https://doi.org/10.3390/rs10060837
Chicago/Turabian StyleZhou, Guanhua, Zhongqi Ma, Shubha Sathyendranath, Trevor Platt, Cheng Jiang, and Kang Sun. 2018. "Canopy Reflectance Modeling of Aquatic Vegetation for Algorithm Development: Global Sensitivity Analysis" Remote Sensing 10, no. 6: 837. https://doi.org/10.3390/rs10060837
APA StyleZhou, G., Ma, Z., Sathyendranath, S., Platt, T., Jiang, C., & Sun, K. (2018). Canopy Reflectance Modeling of Aquatic Vegetation for Algorithm Development: Global Sensitivity Analysis. Remote Sensing, 10(6), 837. https://doi.org/10.3390/rs10060837