
Effect of Organic Potato Farming on Human and Environmental Health and Benefits from New Plant Breeding Techniques. Is It Only a Matter of Public Acceptance?



Abstract
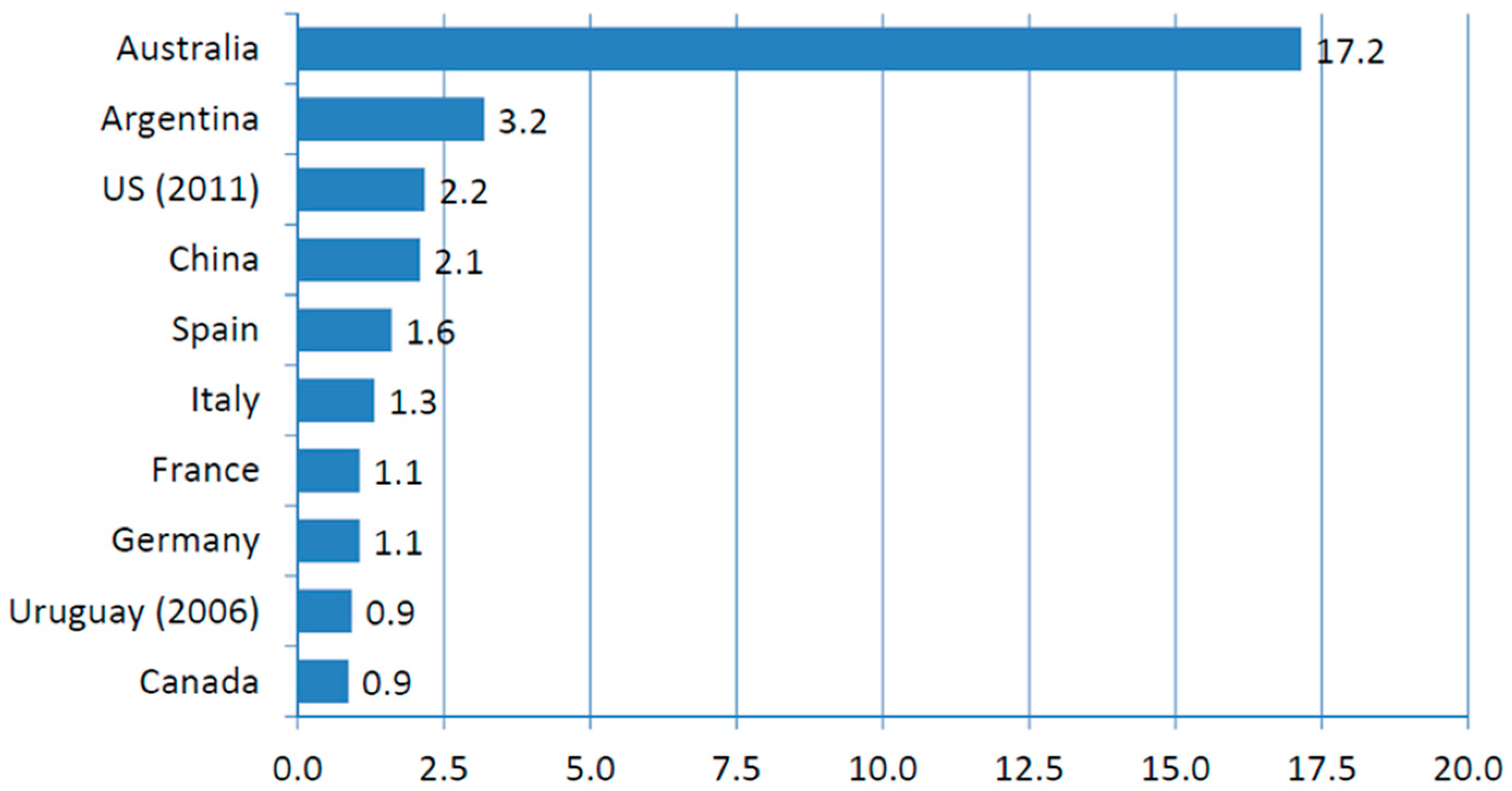
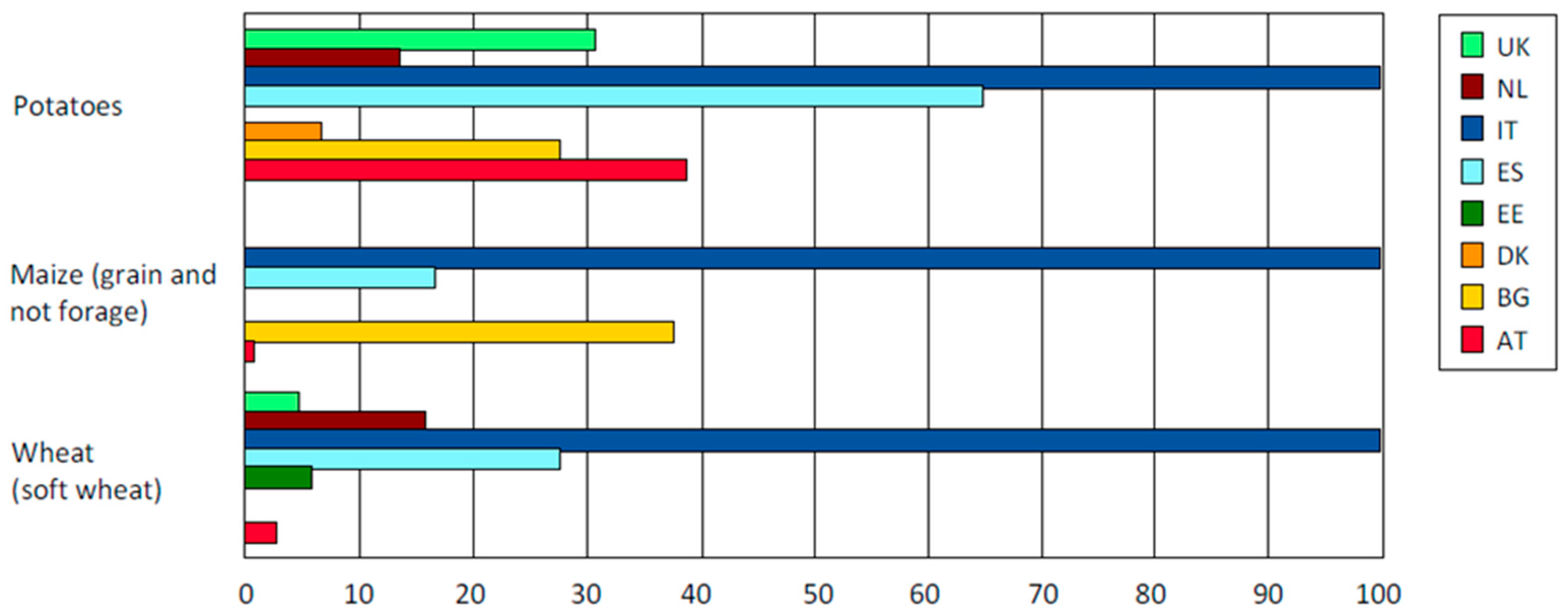
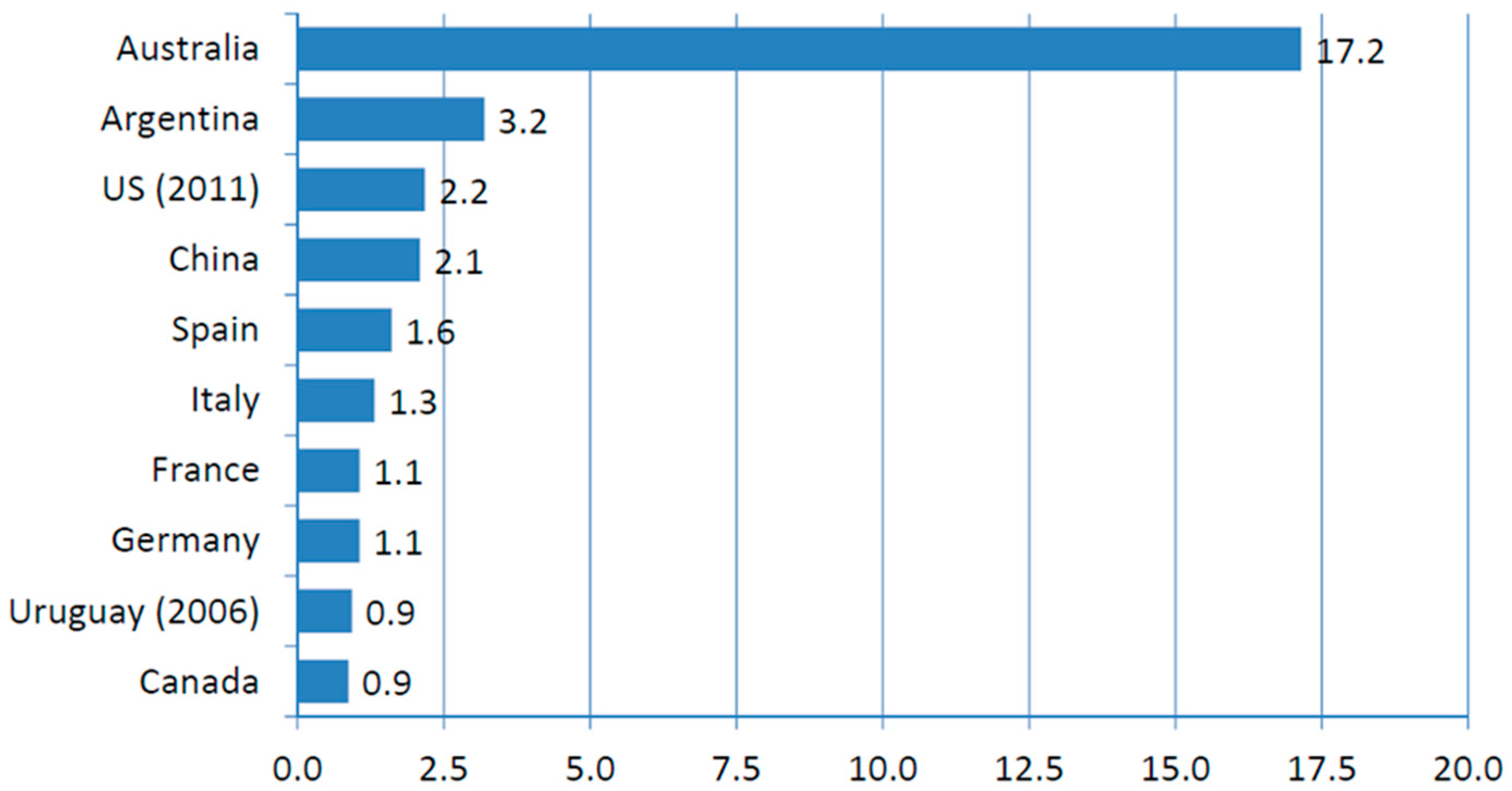
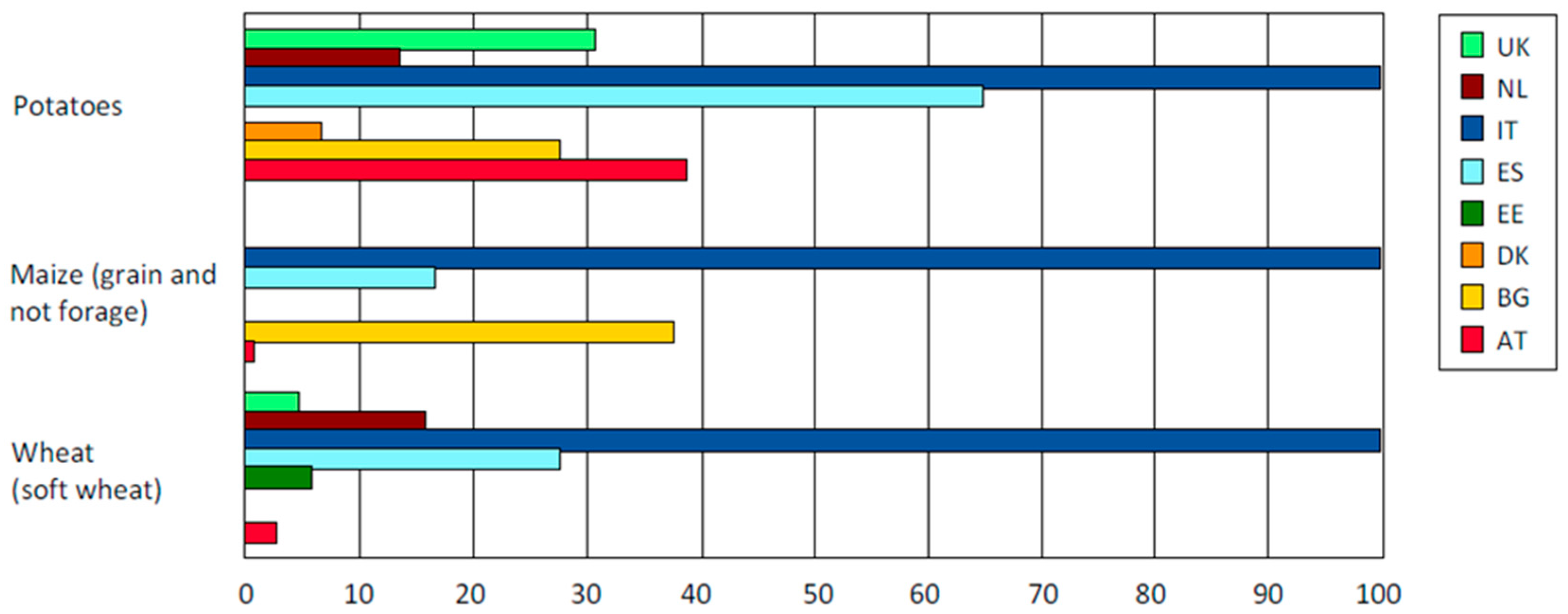
:1. Organic Potatoes Production Requires Breeding
2. Not All GM Potatoes Are Equal
3. Cisgenic Potatoes in Organic Farming: Scientific, Legislative, Ethical, and Social Aspects
4. Discussion and Conclusions
Author Contributions
Conflicts of Interest
References
- International Potato Center. Available online: http://cipotato.org/potato/ (accessed on 5 October 2016).
- FAOSTAT. Available online: http://faostat3.fao.org (accessed on 5 October 2016).
- European Parliament. Directive 2009/128/EC of the European Parliament and of the Council of 21 October 2009 Establishing a Framework for Community Action to Achieve the Sustainable Use of Pesticides. Available online: http://www.uzb.minpolj.gov.rs/attachments/088_Sustainable%20use%20of%20pesticides%202009%20128.pdf (accessed on 10 October 2016).
- Statistics on Organic Agriculture. Available online: http://www.fibl.org/en/themes/organic-farming-statistics.html (accessed on 5 October 2016).
- Consumi: Coldiretti, +20% per Il Biologico nel 2015, Spesa a 2,5 Miliardi. Available online: http://www.coldiretti.it/News/Pagine/636-–-12-Settembre-2015.aspx (accessed on 5 October 2016).
- Herencia, J.F.; García-Galavís, P.A.; Dorado, J.A.R.; Maqueda, C. Comparison of nutritional quality of the crops grown in an organic and conventional fertilized soil. Sci. Hortic. 2011, 129, 882–888. [Google Scholar] [CrossRef]
- Hoefkens, C.; Vandekinderen, I.; de Meulenaer, B.; Devlieghere, F.; Baert, K.; Sioen, I.; de Henauw, S.; Verbeke, W.; van Camp, J. A literature-based comparison of nutrient and contaminant contents between organic and conventional vegetables and potatoes. Br. Food J. 2009, 111, 1078–1097. [Google Scholar] [CrossRef]
- Huber, M.; Rembiałkowska, E.; Średnicka, D.; Bügel, S.; van De Vijver, L.P.L. Organic food and impact on human health: Assessing the status quo and prospects of research. NJAS Wagening. J. Life Sci. 2011, 58, 103–109. [Google Scholar] [CrossRef]
- Lombardo, S.; Lo Monaco, A.; Pandino, G.; Parisi, B.; Mauromicale, G. The phenology, yield and tuber composition of “early” crop potatoes: A comparison between organic and conventional cultivation systems. Renew. Agric. Food Syst. 2012, 28, 50–58. [Google Scholar] [CrossRef]
- Pacifico, D.; Casciani, L.; Ritota, M.; Mandolino, G.; Onofri, C.; Moschella, A.; Parisi, B.; Cafiero, C.; Valentini, M. NMR-Based Metabolomics for Organic Farming Traceability of Early Potatoes. J. Agric. Food Chem. 2013, 61, 11201–11211. [Google Scholar] [CrossRef] [PubMed]
- Skrabule, I.; Muceniece, R.; Kirhnere, I. Evaluation of Vitamins and Glycoalkaloids in Potato Genotypes Grown Under Organic and Conventional Farming Systems. Potato Res. 2013, 56, 259–276. [Google Scholar] [CrossRef]
- Grudzińska, M.; Czerko, Z.; Zarzyńska, K.; Borowska-Komenda, M. Bioactive Compounds in Potato Tubers: Effects of Farming System, Cooking Method, and Flesh Color. PLoS ONE 2016, 11, e0153980. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Sattelmacher, B.; Kutzmutz, E.; Mühling, K.H.; Dittert, K. Influence of Nitrogen Nutrition on Tuber Quality of Potato with Special Reference to the Pathway of Nitrate Transport into Tubers. J. Plant Nutr. 2004, 27, 341–350. [Google Scholar] [CrossRef]
- Lehesranta, S.J.; Koistinen, K.M.; Massat, N.; Davies, H.V.; Shepherd, L.V.T.; McNicol, J.W.; Cakmak, I.; Cooper, J.; Lück, L.; Kärenlampi, S.O.; et al. Effects of agricultural production systems and their components on protein profiles of potato tubers. Proteomics 2007, 7, 597–604. [Google Scholar] [CrossRef] [PubMed]
- Brazinskiene, V.; Asakaviciute, R.; Miezeliene, A.; Alencikiene, G.; Ivanauskas, L.; Jakstas, V.; Viskelis, P.; Razukas, A. Effect of farming systems on the yield, quality parameters and sensory properties of conventionally and organically grown potato (Solanum tuberosum L.) tubers. Food Chem. 2014, 145, 903–909. [Google Scholar] [CrossRef] [PubMed]
- Brandt, K.; Leifert, C.; Sanderson, R.; Seal, C.J. Agroecosystem Management and Nutritional Quality of Plant Foods: The Case of Organic Fruits and Vegetables. CRC Crit. Rev. Plant Sci. 2011, 30, 177–197. [Google Scholar] [CrossRef]
- Hajslová, J.; Schulzová, V.; Slanina, P.; Janné, K.; Hellenäs, K.E.; Andersson, C. Quality of organically and conventionally grown potatoes: Four-year study of micronutrients, metals, secondary metabolites, enzymic browning and organoleptic properties. Food Addit. Contam. 2005, 22, 514–534. [Google Scholar] [CrossRef] [PubMed]
- Merkl, R.; Hrádková, I.; Filip, V.; Šmidrkal, J. Antimicrobial and antioxidant properties of phenolic acids alkyl esters. Czech J. Food Sci. 2010, 28, 275–279. [Google Scholar]
- Bejarano, L.; Mignolet, E.; Devaux, A.; Espinola, N.; Carrasco, E.; Larondelle, Y. Glycoalkaloids in potato tubers: The effect of variety and drought stress on the α-solanine and α-chaconine contents of potatoes. J. Sci. Food Agric. 2000, 80, 2096–2100. [Google Scholar] [CrossRef]
- International Federation of Organic Agriculture Movements (IFOAM). The IFOAM Norms Organic Production Processing, version 20; IFOAM: Bonn, Germany, 2014; pp. 1–252. [Google Scholar]
- Bangemann, L.W.; Westphal, A.; Zwerger, P.; Sieling, K.; Kage, H. Copper reducing strategies for late blight (Phytophthora infestans) control in organic potato (Solanum tuberosum) production. J. Plant Dis. Prot. 2014, 121, 105–116. [Google Scholar] [CrossRef]
- Gianessi, L.; Williams, A. Restrictions on Fungicide Use Causing Decline in Organic Potato Production in Europe; Crop Protection Research Institute: Washington, DC, USA, 2011. [Google Scholar]
- Andersen, M.M.; Landes, X.; Xiang, W.; Anyshchenko, A.; Falhof, J.; Østerberg, J.T.; Olsen, L.I.; Edenbrandt, A.K.; Vedel, S.E.; Thorsen, B.J.; et al. Feasibility of new breeding techniques for organic farming. Trends Plant Sci. 2015, 20, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Palmgren, M.G.; Edenbrandt, A.K.; Vedel, S.E.; Andersen, M.M.; Landes, X.; Østerberg, J.T.; Falhof, J.; Olsen, L.I.; Christensen, S.B.; Sandøe, P.; et al. Are we ready for back-to-nature crop breeding? Trends Plant Sci. 2015, 20, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Jurn Sanders. Evaluation of the EU Legislation on Organic Farming: Study Report; Thünen Institute of Farm Economics: Braunschweig, Germany, 2013. [Google Scholar]
- European Commission. Laying Down Detailed Rules for the Implementation of Council Regulation (EC) No 834/2007 on Organic Production and Labelling of Organic Products with Regard to Organic Production, Labelling and Control. Available online: http://eur-lex.europa.eu/legal-content/en/ALL/?uri=CELEX%3A32008R0889 (accessed on 10 October 2016).
- Schouten, H.J.; Krens, F.A.; Jacobsen, E. Cisgenic plants are similar to traditionally bred plants: International regulations for genetically modified organisms should be altered to exempt cisgenesis. EMBO Rep. 2006, 7, 750–753. [Google Scholar] [CrossRef] [PubMed]
- Rommens, C.M. All-native DNA transformation: A new approach to plant genetic engineering. Trends Plant Sci. 2004, 9, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Podevin, N.; Davies, H.V.; Hartung, F.; Nogué, F.; Casacuberta, J.M. Site-directed nucleases: A paradigm shift in predictable, knowledge-based plant breeding. Trends Biotechnol. 2013, 31, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Kaniewski, W.K.; Thomas, P.E. The potato story. AgBioForum 2004, 7, 41–46. [Google Scholar]
- Toevs, E.A.; Guenthner, J.F.; Johnson, A.J.; McIntosh, C.S.; Thornton, M.K. An industry perspective of all-native and transgenic potatoes. AgBioForum 2011, 14, 14–19. [Google Scholar]
- Jo, K.-R.; Kim, C.-J.; Kim, S.-J.; Kim, T.-Y.; Bergervoet, M.; Jongsma, M.A.; Visser, R.G.F.; Jacobsen, E.; Vossen, J.H. Development of late blight resistant potatoes by cisgene stacking. BMC Biotechnol. 2014, 14, 50. [Google Scholar] [CrossRef] [PubMed]
- Haesaert, G.; Vossen, J.H.; Custers, R.; De Loose, M.; Haverkort, A.; Heremans, B.; Hutten, R.; Kessel, G.; Landschoot, S.; Van Droogenbroeck, B.; et al. Transformation of the potato variety Desiree with single or multiple resistance genes increases resistance to late blight under field conditions. Crop Prot. 2015, 77, 163–175. [Google Scholar] [CrossRef]
- De Vetten, N.; Wolters, A.M.; Raemakers, K.; van der Meer, I.; ter Stege, R.; Heeres, E.; Heeres, P.; Visser, R. A transformation method for obtaining marker-free plants of a cross-pollinating and vegetatively propagated crop. Nat. Biotechnol. 2003, 21, 439–442. [Google Scholar] [CrossRef] [PubMed]
- Chawla, R.; Shakya, R.; Rommens, C.M. Tuber-specific silencing of asparagine synthetase-1 reduces the acrylamide-forming potential of potatoes grown in the field without affecting tuber shape and yield. Plant Biotechnol. J. 2012, 10, 913–924. [Google Scholar] [CrossRef] [PubMed]
- Rommens, C.M.; Ye, J.; Richael, C.; Swords, K. Improving Potato Storage and Processing Characteristics through All-Native DNA Transformation. J. Agric. Food Chem. 2006, 54, 9882–9887. [Google Scholar] [CrossRef] [PubMed]
- Rommens, C.M.; Yan, H.; Swords, K.; Richael, C.; Ye, J. Low-acrylamide French fries and potato chips. Plant Biotechnol. J. 2008, 6, 843–853. [Google Scholar] [CrossRef] [PubMed]
- Clasen, B.M.; Stoddard, T.J.; Luo, S.; Demorest, Z.L.; Li, J.; Cedrone, F.; Tibebu, R.; Davison, S.; Ray, E.E.; Daulhac, A.; et al. Improving cold storage and processing traits in potato through targeted gene knockout. Plant Biotechnol. J. 2015, 14, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Sawai, S.; Ohyama, K.; Yasumoto, S.; Seki, H.; Sakuma, T.; Yamamoto, T.; Takebayashi, Y.; Kojima, M.; Sakakibara, H.; Aoki, T.; et al. Sterol side chain reductase 2 is a key enzyme in the biosynthesis of cholesterol, the common precursor of toxic steroidal glycoalkaloids in potato. Plant Cell 2014, 26, 3763–3774. [Google Scholar] [CrossRef] [PubMed]
- Visser, R.G.F.; Somhorst, I.; Kuipers, G.J.; Ruys, N.J.; Feenstra, W.J.; Jacobsen, E. Inhibition of the expression of the gene for granule-bound starch synthase in potato by antisense constructs. Mol. Gen. Genet. MGG 1991, 225, 289–296. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (EFSA). Consolidated presentation of the joint Scientific Opinion of the GMO and BIOHAZ Panels on the “Use of Antibiotic Resistance Genes as Marker Genes in Genetically Modified Plants” and the Scientific Opinion of the GMO Panel on “Consequences of the Opinion on the Use of Antibiotic Resistance Genes as Marker Genes in Genetically Modified Plants on Previous EFSA Assessments of Individual GM Plants”. EFSA J. 2009, 1108, 1–8. [Google Scholar]
- Storck, T.; Böhme, T.; Schultheiss, H. Status and perspectives of GM approaches to fight late blight. In Proceedings of the Thirteenth euroblight workshop, St. Petersburg, Russia, 9–12 October 2011; pp. 45–48.
- Ye, J.; Shakya, R.; Shrestha, P.; Rommens, C.M. Tuber-Specific Silencing of the Acid Invertase Gene Substantially Lowers the Acrylamide-Forming Potential of Potato. J. Agric. Food Chem. 2010, 58, 12162–12167. [Google Scholar] [CrossRef] [PubMed]
- Rommens, C.M.; Conner, A.; Yan, H.; Hanley, Z. Intragenic Vectors and Marker-Free Transformation: Tools for a Greener Biotechnology. Plant Transform. Technol. 2011, 60, 93–107. [Google Scholar]
- Bachem, C.W.B.; Speckmann, G.-J.; van der Linde, P.C.G.; Verheggen, F.T.M.; Hunt, M.D.; Steffens, J.C.; Zabeau, M. Antisense Expression of Polyphenol Oxidase Genes Inhibits Enzymatic Browning in Potato Tubers. Nat. Biotechnol. 1994, 12, 1101–1105. [Google Scholar] [CrossRef]
- Innate® Second Generation Potato Receives Fda Safety Clearance. Available online: http://www.simplot.com/news/innate_second_generation_potato_receives_fda_safety_clearance (accessed on 5 October 2016).
- Haverkort, A.J.; Boonekamp, P.M.; Hutten, R.; Jacobsen, E.; Lotz, L.A.P.; Kessel, G.J.T.; Vossen, J.H.; Visser, R.G.F. Durable late blight resistance in potato through dynamic varieties obtained by cisgenesis: Scientific and societal advances in the DuRPh project. Potato Res. 2016, 59, 35–66. [Google Scholar] [CrossRef]
- Haverkort, A.J.; Struik, P.C.; Visser, R.G.F.; Jacobsen, E. Applied Biotechnology to Combat Late Blight in Potato Caused by Phytophthora Infestans. Potato Res. 2009, 52, 249–264. [Google Scholar] [CrossRef]
- Christian, M.; Cermak, T.; Doyle, E.L.; Schmidt, C.; Zhang, F.; Hummel, A.; Bogdanove, A.J.; Voytas, D.F. Targeting DNA double-strand breaks with TAL effector nucleases. Genetics 2010, 186, 756–761. [Google Scholar] [CrossRef] [PubMed]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A Programmable Dual-RNA-Guided DNA Endonuclease in Adaptive Bacterial Immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Nicolia, A.; Proux-Wéra, E.; Ahman, I.; Onkokesung, N.; Andersson, M.; Andreasson, E.; Zhu, L.H. Targeted gene mutation in tetraploid potato through transient TALEN expression in protoplasts. J. Biotechnol. 2015, 204, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Butler, N.M.; Atkins, P.A.; Voytas, D.F.; Douches, D.S. Generation and Inheritance of Targeted Mutations in Potato (Solanum tuberosum L.) Using the CRISPR/Cas System. PLoS ONE 2015, 10, e0144591. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, S.; Wang, W.; Xiong, X.; Meng, F.; Cui, X. Efficient targeted mutagenesis in potato by the CRISPR/Cas9 system. Plant Cell Rep. 2015, 34, 1473–1476. [Google Scholar] [CrossRef] [PubMed]
- Ryffel, G.U. Orgenic plants: Gene-manipulated plants compatible with organic farming. Biotechnol. J. 2012, 7, 1328–1331. [Google Scholar] [CrossRef] [PubMed]
- Van Bueren, E.T.L.; Verhoog, H.; Tiemens-Hulscher, M.; Struik, P.C.; Haring, M.A. Organic agriculture requires process rather than product evaluation of novel breeding techniques. NJAS Wagening. J. Life Sci. 2007, 54, 401–412. [Google Scholar] [CrossRef]
- International Federation of Organic Agriculture Movements (IFOAM). Position on Genetic Engineering and Genetically Modified Organism; IFOAM: Bonn, Germany, 2002. [Google Scholar]
- International Federation of Organic Agriculture Movements (IFOAM). Public Consultation on the Position of Ifoam—Organics International on Genetic Engineering and Genetically Modified Organisms. Available online: http://www.ifoam.bio/en/news/2016/02/26/public-consultation-position-ifoam-organics-international-genetic-engineering-and (accessed on 5 October 2016).
- Food and Agriculture Organization of the United Nations. Council Regulation (EEC) No 2092/91 on Organic Production of Agricultural Products and Indications Referring There to on Agricultural Products and Foodstuffs. Counc. Regul.; (EEC) No 2092/91; Food and Agriculture Organization of the United Nations: Rome, Italy, 1991; pp. 1–95. [Google Scholar]
- Kuiper, H.A.; Kleter, G.A.; Noteborn, H.P.J.M.; Kok, E.J. Assessment of the food safety issues related to genetically modified foods. Plant J. 2001, 27, 503–528. [Google Scholar] [CrossRef] [PubMed]
- Cellini, F.; Chesson, A.; Colquhoun, I.; Constable, A.; Davies, H.V.; Engel, K.H.; Gatehouse, A.M.R.; Kärenlampi, S.; Kok, E.J.; Leguay, J.J.; et al. Unintended effects and their detection in genetically modified crops. Food Chem. Toxicol. 2004, 42, 1089–1125. [Google Scholar] [CrossRef] [PubMed]
- Organisation for Economic Co-Operation and Development (OECD). Safety Evaluation of Foods Derived by Modern Biotechnology: Concepts and Principles; OECD: Paris, France, 1993. [Google Scholar]
- World Health Organization. Safety Aspects of Genetically Modified Foods of Plant Origin. Available online: http://apps.who.int/iris/handle/10665/66575 (accessed on 18 October 2016).
- Shepherd, L.V.T.; McNicol, J.W.; Razzo, R.; Taylor, M.A.; Davies, H.V. Assessing the potential for unintended effects in genetically modified potatoes perturbed in metabolic and developmental processes. Targeted analysis of key nutrients and anti-nutrients. Transgenic Res. 2006, 15, 409–425. [Google Scholar] [CrossRef] [PubMed]
- Catchpole, G.S.; Beckmann, M.; Enot, D.P.; Mondhe, M.; Zywicki, B.; Taylor, J.; Hardy, N.; Smith, A.; King, R.D.; Kell, D.B.; et al. Hierarchical metabolomics demonstrates substantial compositional similarity between genetically modified and conventional potato crops. Proc. Natl. Acad. Sci. USA 2005, 102, 14458–14462. [Google Scholar] [CrossRef] [PubMed]
- Lehesranta, S.J.; Davies, H.V.; Shepherd, L.V.T.; Nunan, N.; McNicol, J.W.; Auriola, S.; Koistinen, K.M.; Suomalainen, S.; Kokko, H.I.; Kärenlampi, S.O. Comparison of Tuber Proteomes of Potato Varieties, Landraces, and Genetically Modified Lines. Plant Physiol. 2005, 138, 1690–1699. [Google Scholar] [CrossRef] [PubMed]
- Kortstee, A.J.; Khan, S.A.; Helderman, C.; Trindade, L.M.; Wu, Y.; Visser, R.G.F.; Brendolise, C.; Allan, A.; Schouten, H.J.; Jacobsen, E. Anthocyanin production as a potential visual selection marker during plant transformation. Transgenic Res. 2011, 20, 1253–1264. [Google Scholar] [CrossRef] [PubMed]
- Krens, F.A.; Schaart, J.G.; van der Burgh, A.M.; Tinnenbroek-Capel, I.E.M.; Groenwold, R.; Kodde, L.P.; Broggini, G.A.L.; Gessler, C.; Schouten, H.J. Cisgenic Apple Trees; Development, Characterization, and Performance. Available online: http://journal.frontiersin.org/article/10.3389/fpls.2015.00286/full (accessed on 10 October 2016).
- Arber, W. Genetic engineering compared to natural genetic variations. New Biotechnol. 2010, 27, 517–521. [Google Scholar] [CrossRef] [PubMed]
- Kyndt, T.; Quispe, D.; Zhai, H.; Jarret, R.; Ghislain, M.; Liu, Q.; Gheysen, G.; Kreuze, J.F. The genome of cultivated sweet potato contains Agrobacterium T-DNAs with expressed genes: An example of a naturally transgenic food crop. Proc. Natl. Acad. Sci. USA 2015, 112, 5844–5849. [Google Scholar] [CrossRef] [PubMed]
- Haverkort, A.J.; Boonekamp, P.M.; Hutten, R.; Jacobsen, E.; Lotz, L.A.P.; Kessel, G.J.T.; Visser, R.G.F.; Van Der Vossen, E.A.G. Societal costs of late blight in potato and prospects of durable resistance through cisgenic modification. Potato Res. 2008, 51, 47–57. [Google Scholar] [CrossRef]
- International Potato Center (CIP). Available online: http://cipotato.org (accessed on 7 October 2016).
- Xu, X.; Pan, S.; Cheng, S.; Zhang, B.; Mu, D.; Ni, P.; Zhang, G.; Yang, S.; Li, R.; Wang, J.; et al. Genome sequence and analysis of the tuber crop potato. Nature 2011, 475, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Aversano, R.; Contaldi, F.; Ercolano, M.R.; Grosso, V.; Iorizzo, M.; Tatino, F.; Xumerle, L.; Dal Molin, A.; Avanzato, C.; Ferrarini, A.; et al. The Solanum commersonii Genome Sequence Provides Insights into Adaptation to Stress Conditions and Genome Evolution of Wild Potato Relatives. Plant Cell 2015, 27, 954–968. [Google Scholar] [CrossRef] [PubMed]
- Cardi, T. Cisgenesis and genome editing: Combining concepts and efforts for a smarter use of genetic resources in crop breeding. Plant Breed. 2016. [Google Scholar] [CrossRef]
- The Pew Charitable Trusts. Guide to U.S. Regulation of Genetically Modified Food and Agricultural Biotechnology Products; Pew Initiative on Food and Biotechnology; The Pew Charitable Trusts: Washington, DC, USA, 1986. [Google Scholar]
- European Parliament. Directive 2001/18/EC of the European Parliament and of the Council of 12 March 2001 on the Deliberate Release into the Environment of Genetically Modified Organisms and Repealing Council Directive; 90/220/EEC; Official Journal of the European Union: Brussels, Belgum, 2001; Volume 2000, pp. 1–38. [Google Scholar]
- European Parliament. Council of the European Union Regulation (EC) No 1829/2003 of the European Parliament and of the Council of 22 September 2003 on Genetically Modified Food and Feed; Official Journal of the European Union: Brussels, Belgum, 2003. [Google Scholar]
- Schouten, H.J.; Krens, F.A.; Jacobsen, E. Do cisgenic plants warrant less stringent oversight? Nat. Biotechnol. 2006, 24, 753. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (EFSA). Panel on Genetically Modified Organisms Scientific opinion addressing the safety assessment of plants developed using Zinc Finger Nuclease 3 and other Site-Directed Nucleases with similar function. EFSA J. 2012, 10, 2943. [Google Scholar]
- DeFrancesco, L. How safe does transgenic food need to be? Nat. Biotechnol. 2013, 31, 794–802. [Google Scholar] [CrossRef] [PubMed]
- Delwaide, A.-C.; Nalley, L.L.; Dixon, B.L.; Danforth, D.M.; Nayga, R.M., Jr.; Van Loo, E.J.; Verbeke, W. Revisiting GMOs: Are There Differences in European Consumers’ Acceptance and Valuation for Cisgenically vs Transgenically Bred Rice? PLoS ONE 2015, 10, e0126060. [Google Scholar] [CrossRef] [PubMed]
- Holme, I.B.; Wendt, T.; Holm, P.B. Intragenesis and cisgenesis as alternatives to transgenic crop development. Plant Biotechnol. J. 2013, 11, 395–407. [Google Scholar] [CrossRef] [PubMed]
- Lammerts Van Bueren, E.T.; Tiemens-Hulscher, M.; Struik, P.C. Cisgenesis does not solve the late blight problem of organic potato production: Alternative breeding strategies. Potato Res. 2008, 51, 89–99. [Google Scholar] [CrossRef]
- Connor, D.J. Organically grown crops do not a cropping system make and nor can organic agriculture nearly feed the world. Field Crop. Res. 2013, 144, 145–147. [Google Scholar] [CrossRef]
- Seufert, V.; Ramankutty, N.; Foley, J.A. Comparing the yields of organic and conventional agriculture. Nature 2012, 485, 229–232. [Google Scholar] [CrossRef] [PubMed]
- Crespo-Herrera, L.A.; Ortiz, R. Plant breeding for organic agriculture: Something new? Agric. Food Secur. 2015. [Google Scholar] [CrossRef]
- Van Bueren, E.T.L.; Jones, S.S.; Tamm, L.; Murphy, K.M.; Myers, J.R.; Leifert, C.; Messmer, M.M. The need to breed crop varieties suitable for organic farming, using wheat, tomato and broccoli as examples: A review. NJAS Wagening. J. Life Sci. 2011, 58, 193–205. [Google Scholar] [CrossRef]
- Van Bueren, E.T.L.; Struik, P.C.; Jacobsen, E. Ecological concepts in organic farming and their consequences for an organic crop ideotype. NJAS Wagening. J. Life Sci. 2002, 50, 1–26. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Trait | Gene | Technology | References |
|---|---|---|---|
| Late blight resistance | Rpi-sto1, Rpvnt1.1 | Cisgenesis | [32] |
| Late blight resistance | Rpvnt1.1 | Cisgenesis | [33] |
| High amylopectin | GBSS | Intragenesis | [34] |
| Accumulation of reducing sugars after cold storage and acrylamide after high temperature processing | R1, PhL, StAs1, StAs2 | Intragenesis | [35,36,37] |
| Accumulation of reducing sugars after cold storage and acrylamide after high temperature processing | VInv | TALEN | [38] |
| Accumulation of steroidal glycoalkaloids | StSSR2 | TALEN | [39] 1 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pacifico, D.; Paris, R. Effect of Organic Potato Farming on Human and Environmental Health and Benefits from New Plant Breeding Techniques. Is It Only a Matter of Public Acceptance? Sustainability 2016, 8, 1054. https://doi.org/10.3390/su8101054
Pacifico D, Paris R. Effect of Organic Potato Farming on Human and Environmental Health and Benefits from New Plant Breeding Techniques. Is It Only a Matter of Public Acceptance? Sustainability. 2016; 8(10):1054. https://doi.org/10.3390/su8101054
Chicago/Turabian StylePacifico, Daniela, and Roberta Paris. 2016. "Effect of Organic Potato Farming on Human and Environmental Health and Benefits from New Plant Breeding Techniques. Is It Only a Matter of Public Acceptance?" Sustainability 8, no. 10: 1054. https://doi.org/10.3390/su8101054
APA StylePacifico, D., & Paris, R. (2016). Effect of Organic Potato Farming on Human and Environmental Health and Benefits from New Plant Breeding Techniques. Is It Only a Matter of Public Acceptance? Sustainability, 8(10), 1054. https://doi.org/10.3390/su8101054