


Terrestrial LiDAR Technology to Evaluate the Vertical Structure of Stands of Bertholletia excelsa Bonpl., a Species Symbol of Conservation Through Sustainable Use in the Brazilian Amazon

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
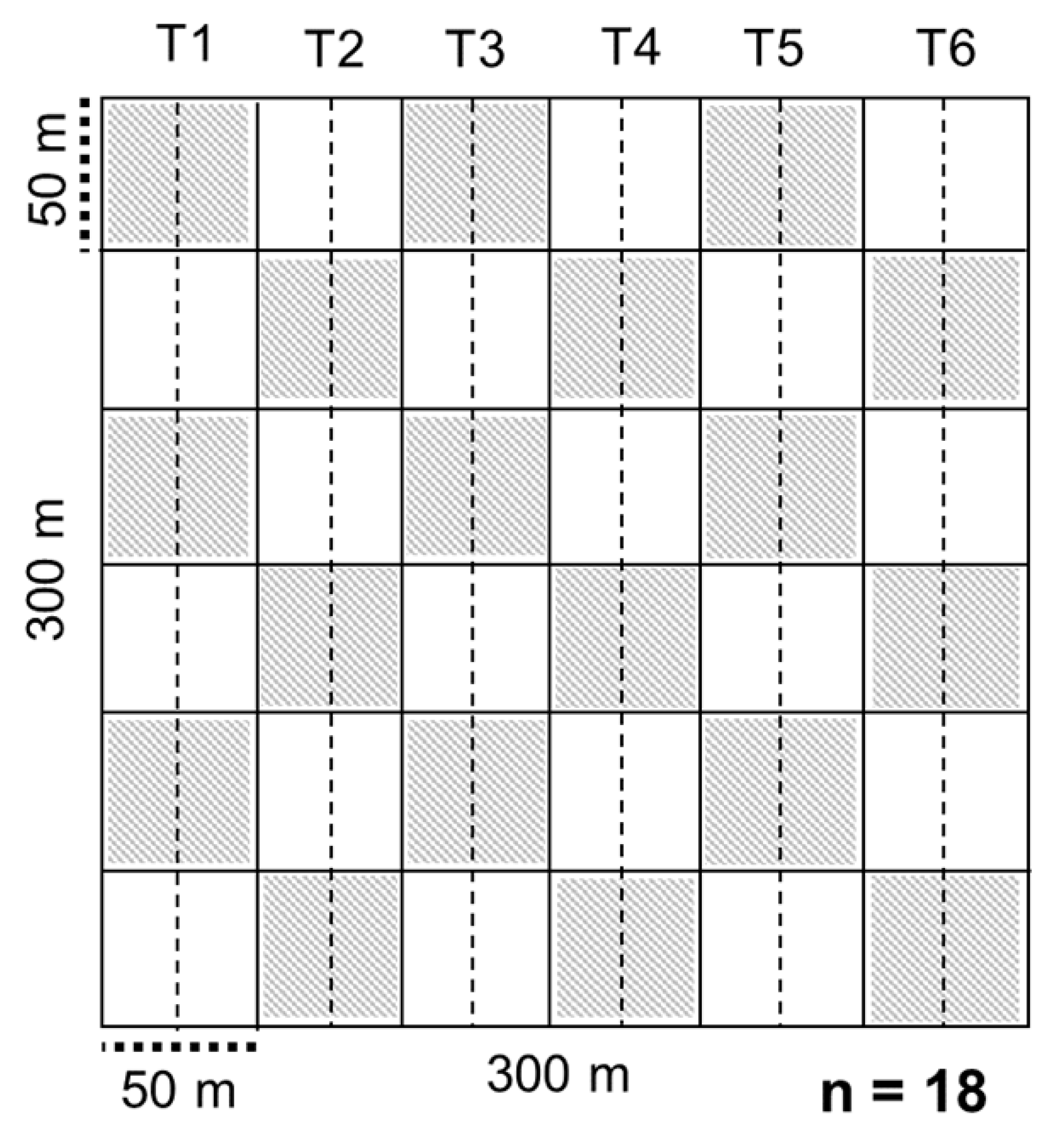
2.1. Study Area and Sampling Design
2.2. Data Collection in the Field
2.3. Lidar Metrics for the Analysis of the Vertical Structure of the Forest
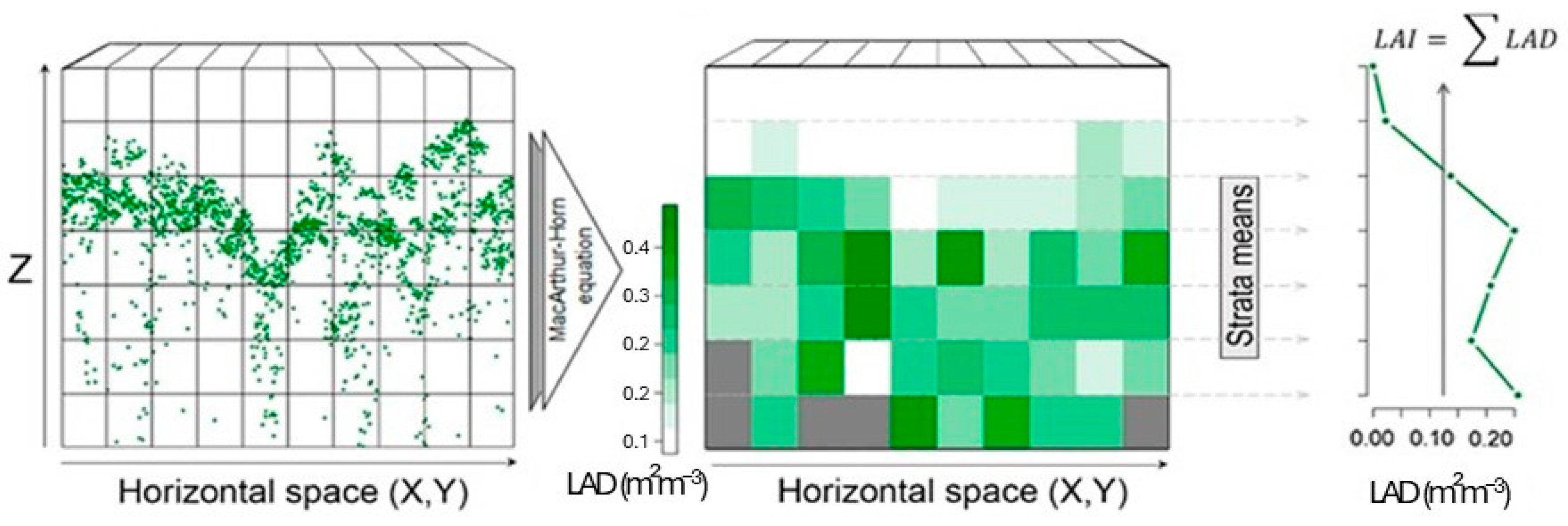
- LAD Brazil nut is the leaf area density of a “voxel” of 1 m in height × 2 m in length;
- i: “voxel” in the vertical plane: 1–2 m, 2–3 m, 3–4 m, etc.;
- j: “voxel” in the horizontal plane, in intervals of 0–2 m, 2–4 m, 4–6 m, etc.;
- pulses.in: number of pulses that entered each voxel;
- pulses.out: number of pulses that passed through that voxel; and
- ∆h: height of the voxel (∆h = 1 m).
- Leaf Area Index (LAI): the calculation of LAI was conducted using an equation that sums all the LADs from a pile of voxels in a specific height interval (Equation (2) and Figure A2).
- LAIj: leaf area index of the linear stretch j of the transect;
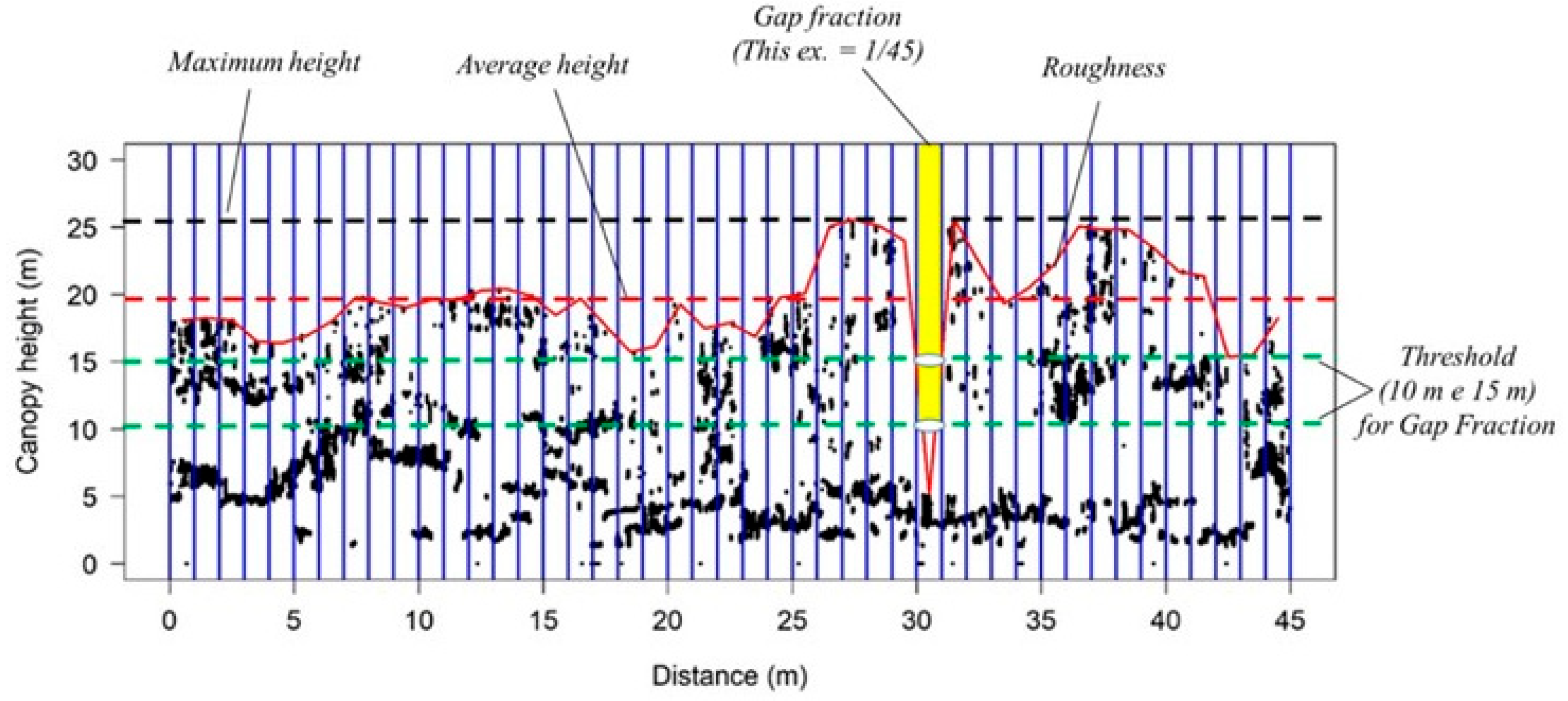
- Canopy Rugosity (R): standard deviation of maximum heights measured at 1 m horizontal resolution;
- Maximum height (Hmax): highest value found in the interval of 50 m, considering variations at each meter along the axis of the LiDAR path;
- Average height (Havg): average value of the highest points, considering variations at each meter in the interval of 50 m;
- Fraction of clearings (F): opening of the canopy at a specific height (10 m or 15 m), which is calculated using the proportion of columns of 1 m in width along the x-axis, without information about the vegetation starting from the threshold of height, as defined by the total number of columns in the sampled interval (Appendix C, Table A3).
2.4. Data Analysis and Processing
3. Results
3.1. Vertical Structure of Amazonian Forests with Brazil Nut Trees (Regional Scale)
3.2. The Structure of the Canopy in Different Forest Typologies in Sites in the State of Amapá (Local Scale)
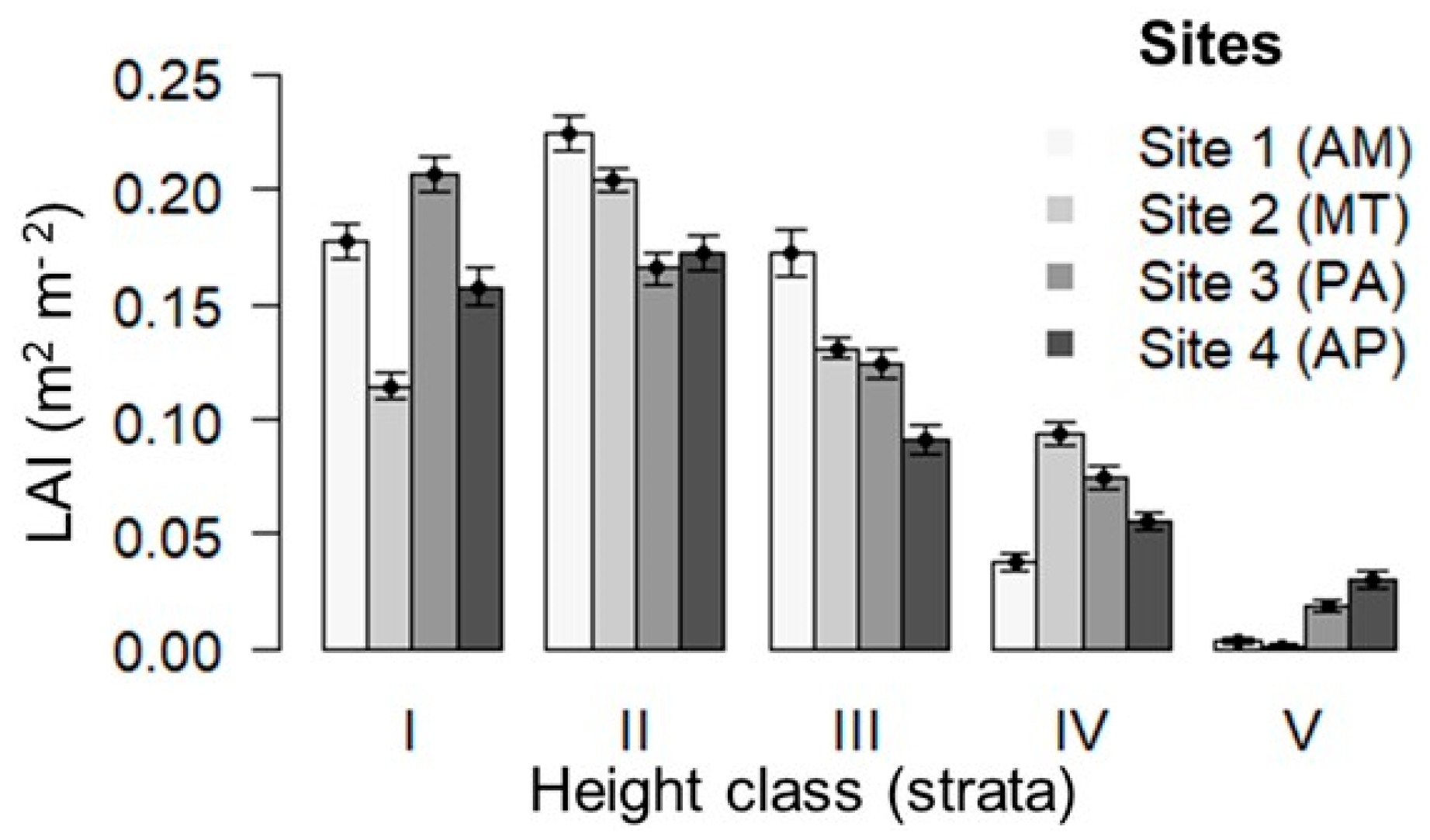
3.3. Relationship Between Abundance of Brazil Nut Trees and Leaf Area Index in Different Height Strata
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest Sites | Climate | Soil | Vegetation | Latitude (S) | Longitude (W) |
|---|---|---|---|---|---|
| AM | Af | PVAd | Dbe | 3°38′3.6″ | 64°18′37.8″ |
| MT | Am | LVd | Fse | 11°5′55.1″ | 55°2′12.5″ |
| PA | Af | LAd | Dbe | 3°3′18,6″ | 54°55′41.9″ |
| AP | Am | PVAd | Dse | 0°33′50.1″ | 52°18′23.8″ |
| AP* | Am | LAd | Spf | 0°25′15.5″ | 51°57′50.1″ |

Appendix B
| Metrics | AM-AP | AM-MT | AP-MT | AM-PA | AP-PA | MT-PA | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| p | p.ad | p | p.ad | p | p.ad | p | p.ad | p | p.ad | p | p.ad | |
| Hmax | 0.000 | 0.000 | 0.154 | 0.924 | 0.009 | 0.055 | 0.000 | 0.000 | 0.905 | 1.00 | 0.012 | 0.077 |
| R | 0.000 | 0.000 | 0.235 | 1.00 | 0.003 | 0.019 | 0.000 | 0.001 | 0.673 | 1.00 | 0.011 | 0.068 |
| S (%) | 0.315 | 1.00 | 0.949 | 1.00 | 0.347 | 1.00 | 0.006 | 0.033 | 0.000 | 0.000 | 0.005 | 0.027 |
| LAI I | 0.308 | 1.00 | 0.005 | 0.033 | 0.078 | 0.470 | 0.141 | 0.844 | 0.013 | 0.076 | 0.000 | 0.000 |
| LAI III | 0.003 | 0.015 | 0.255 | 1.00 | 0.060 | 0.361 | 0.130 | 0.781 | 0.132 | 0.794 | 0.708 | 1.00 |
| LAI IV | 0.170 | 1.00 | 0.001 | 0.005 | 0.048 | 0.288 | 0.005 | 0.027 | 0.142 | 0.855 | 0.610 | 1.00 |
| LAI V | 0.001 | 0.006 | 0.847 | 1.00 | 0.001 | 0.003 | 0.013 | 0.075 | 0.426 | 1.00 | 0.007 | 0.043 |
| LAI IV + V | 0.002 | 0.012 | 0.003 | 0.020 | 0.886 | 1.00 | 0.001 | 0.006 | 0.833 | 1.00 | 0.722 | 1.00 |
| LAI General | 0.051 | 0.306 | 0.454 | 1.00 | 0.229 | 1.00 | 0.205 | 1.00 | 0.001 | 0.008 | 0.044 | 0.263 |
Appendix C
| Metrics | Forest Sites (M ± SD) | Test Wilcoxon–Mann–Whitney | ||
|---|---|---|---|---|
| Forest | Savannah/Forest | W | p-Value | |
| Hmax | 38.1 ± 8.77 | 37.4 ± 4.55 | 200 | 0.239 |
| Hmed | 28.9 ± 9.03 | 31.2 ± 4.96 | 148 | 0.673 |
| R | 7.54 ± 2.54 | 5.08 ± 1.66 | 252 | 0.004 |
| S (%) | 3.71 ± 5.39 | 3.15 ± 1.91 | 120 | 0.192 |
| F (10m) | 0.05 ± 0.08 | 0.00 ± 0.01 | 227 | 0.007 |
| F (15m) | 0.15 ± 0.20 | 0.01 ± 0.03 | 232 | 0.012 |
| LAI I | 1.10 ± 0.57 | 0.69 ± 0.28 | 239 | 0.014 |
| LAI II | 1.90 ± 0.57 | 1.51 ± 0.29 | 231 | 0.029 |
| LAI III | 0.73 ± 0.61 | 1.29 ± 0.69 | 85 | 0.014 |
| LAI IV | 0.49 ± 0.39 | 0.65 ± 0.40 | 120 | 0.189 |
| LAI V | 0.44 ± 0.50 | 0.31 ± 0.34 | 173 | 0.725 |
| LAI IV + V | 0.94 ± 0.69 | 0.92 ± 0.63 | 199 | 0.248 |
| LAI General | 4.65 ± 0.68 | 4.45 ± 0.41 | 211 | 0.126 |
Appendix D
Appendix E
Appendix F
References
- Thomas, E.; Caicedo, C.A.; Loo, J.; Kindt, R. The distribution of the Brazil nut (Bertholletia excelsa) through time: From range contraction in glacial refugia, over human-mediated expansion, to anthropogenic climate change. Bol. Mus. Para. Emílio Goeldi 2014, 9, 267–291. [Google Scholar]
- Mori, S.A.; Prance, G.T. Taxonomy, ecology, and economy botany of Brazil nut (Bertholletia excelsa Humb. & Bonpl.: Lecythidaceae). Adv. Econ. Bot. 1990, 8, 130–150. [Google Scholar]
- Haugaasen, J.M.T.; Haugaasen, T.; Peres, C.A.; Gribel, R.; Wegge, P. Fruit Removal and Natural Seed Dispersal of the Brazil Nut Tree (Bertholletia excelsa) in Central Amazonia, Brazil. Biotropica 2011, 44, 205–210. [Google Scholar] [CrossRef]
- Duchelle, A.E.; Guariguata, M.R.; Less, G.; Albornoz, M.A.; Chavez, A.; Melo, T. Evaluating the opportunities and limitations to multiple use of Brazil nuts and timber in Western Amazonia. For. Ecol. Manag. 2012, 268, 39–48. [Google Scholar] [CrossRef]
- Scoles, R.; Gribel, R. The regeneration of Brazil nut trees in relation to nut harvest intensity in the Trombetas River valley of Northern Amazonia, Brazil. For. Ecol. Manag. 2012, 265, 71–81. [Google Scholar] [CrossRef]
- Zuidema, P.A.; Boot, R.G.A. Demography of the Brazil nut tree (Bertholletia excelsa) in the Bolivian Amazon: Impact of seed extraction on recruitment and population dynamics. J. Trop. Ecol. 2002, 18, 1–31. [Google Scholar] [CrossRef]
- Myers, G.P.; Newton, A.C.; Melgarejo, O. The influence of canopy gap size on natural regeneration of Brazil nut (Bertholletia excelsa) in Bolivia. For. Ecol. Manag. 2000, 127, 119–128. [Google Scholar] [CrossRef]
- Peres, C.A.; Baider, C. Seed dispersal, spatial distribution and population structure of Brazil nut trees (Bertholletia excelsa) in southeastern Amazonia. J. Trop. Ecol. 1997, 13, 595–616. [Google Scholar] [CrossRef]
- Conab. Proposta de Preços Mínimos (safra 2015/16); Conab: Brasília, Distrito Federal, Brazil, 2015; Volume 2, pp. 1–159. [Google Scholar]
- Angelo, H.; De Almeida, A.N.; Calderon, R.A.; Pompermayer, R.S.; De Souza, A.N. Determinantes do preço da castanha-do-brasil (Bertholletia excelsa) no mercado interno brasileiro. Sci. For. 2013, 41, 195–203. [Google Scholar]
- Engelbrecht, B.M.J.; Herz, H.M. Evaluation of different methods to estimate understory light conditions in Tropical Forests. J. Trop. Ecol. 2001, 17, 207–224. [Google Scholar] [CrossRef]
- Jardim, F.C.S.; Serrão, D.R.; Nemer, T.C. Efeito de diferentes tamanhos de clareiras sobre o crescimento e a mortalidade de espécies arbóreas em Moju-PA. Acta Amaz. 2007, 37, 36–48. [Google Scholar] [CrossRef]
- Scoles, R.; Gribel, R. Population structure of Brazil nut (Bertholletia excelsa, Lecythidaceae) stands in two areas with different occupation histories in the Brazilian Amazon. Hum. Ecol. 2011, 39, 455–464. [Google Scholar] [CrossRef]
- Scoles, R.; Klein, G.N.; Gribel, R. Crescimento e sobrevivência de castanheira (Bertholletia excelsa Bonpl., Lecythidaceae) plantada em diferentes condições de luminosidade após seis anos de plantio na região do rio Trombetas, Oriximiná, Pará. Bol. Do Mus. Para. Emílio Goeldi. Ciências Nat. 2014, 9, 321–336. [Google Scholar] [CrossRef]
- Kainer, K.A.; Duryea, M.L.; Costa de Macedo, N.; Williams, K. Brazil nut seedling establishment and autoecology in an extractive reserve in Acre, Brazil. Ecol. Appl. 1998, 8, 397–410. [Google Scholar] [CrossRef]
- Salomão, R.P.; Rosa, N.A.; Castilho, A.; Morais, K.A.C. Castanheira-do-brasil recuperando áreas degradadas e provendo alimento e renda para comunidades da Amazônia Setentrional. Bol. Do Mus. Para. Emílio Goeldi 2006, 1, 65–78. [Google Scholar] [CrossRef]
- Salomão, R.P. Densidade, estrutura e distribuição espacial de castanheira-do-brasil (Bertholletia excelsa H. & B.) em dois platôs de floresta ombrófila densa na Amazônia setentrional brasileira. Bol. Do Mus. Para. Emílio Goeldi 2009, 4, 11–25. [Google Scholar]
- Wilson, J.W. Analysis of the spatial distribution of foliage by two-dimensional point quadrats. New Phytol. 1958, 59, 92–101. [Google Scholar] [CrossRef]
- Aber, J.D. A method for estimating foliage-height profiles in broad-leaved forests. J. Ecol. 1979, 67, 35–40. [Google Scholar] [CrossRef]
- Lefsky, M.A.; Cohen, W.B.; Parker, G.G.; Harding, D.K. Lidar Remote Sensing for Ecosystem Studies. BioScience 2002, 52, 19–30. [Google Scholar] [CrossRef]
- Almeida, D.R.A.; Nelson, B.W.; Schietti, J.; Gorgens, E.B.; Resende, A.F.; Stark, S.C.; Valbuena, R. Contrasting fire damage and fire susceptibility between seasonally flooded forest and upland forest in the Central Amazon using portable profiling LiDAR. Remote Sens. Environ. 2016, 184, 153–160. [Google Scholar] [CrossRef]
- Stark, S.C.; Leitold, V.; Wu, J.; Hunter, M.O.; De Castilho, C.V.; Carolina, V.; Costa, F.R.C.; Mcmahon, S.M.; Parker, G.G.; ShimaBukuro, M.T.; et al. Amazon forest carbon dynamics predicted by profiles of canopy leaf area and light environment. Ecol. Lett. 2012, 15, 1406–1414. [Google Scholar] [CrossRef] [PubMed]
- Stark, S.C.; Enquist, B.J.; Saleska, S.R.; Leitold, V.; Schietti, J.; Longo, M.; Alves, L.F.; Camargo, P.B.; De Oliveira, R.C. Linking canopy leaf area and light environments with tree size distributions to explain Amazon forest demography. Ecol. Lett. 2015, 18, 636–645. [Google Scholar] [CrossRef] [PubMed]
- Parker, G.G.; Harding, D.J.; Berger, M.L. A portable LIDAR system for rapid determination of forest canopy structure. J. Appl. Ecol. 2004, 41, 755–767. [Google Scholar] [CrossRef]
- Hardiman, B.S.; Bohrer, G.; Gough, C.M.; Vogel, C.S.; Curtis, P.S. The role of canopy structural complexity in wood net primary production of a maturing northern deciduous forest. Ecology 2011, 92, 1818–1827. [Google Scholar] [CrossRef]
- Hardiman, B.S.; Gough, C.M.; Halperin, A.; Hofmeister, K.L.; Nave, L.E.; Bohrer, G.; Curtis, P.S. Maintaining high rates of carbon storage in old forests: A mechanism linking canopy structure to forest function. For. Ecol. Manag. 2013, 298, 111–119. [Google Scholar] [CrossRef]
- Sawada, Y.; Rempei, S.; Jindo, K.; Endo, T.; Oki, K.; Sawada, H.; Arai, E.; Shimabukuro, Y.E.; Celes, C.H.S.; Campos, M.A.A.; et al. A new 500-m resolution map of canopy height for Amazon forest using spaceborne LiDAR and cloud-free MODIS imagery. Int. J. Appl. Earth Obs. Geoinf. 2015, 43, 92–101. [Google Scholar] [CrossRef]
- Simard, M.; Pinto, N.; Fisher, J.B.; Baccini, A. Mapping forest canopy height globally with spaceborne lidar. J. Geophys. Res. Biogeosci. 2011, 116, G04021. [Google Scholar] [CrossRef]
- Ter Steege, H.; Pitman, N.; Sabatier, D.; Castellanos, H.; Hout, P.V.D.; Daly, D.C.; Silveira, M.; Phillips, O.; Vasquez, R.; Andel, T.V.; et al. A spatial model of tree α-diversity and -density for the Amazon. Biodivers. Conserv. 2003, 12, 2255–2277. [Google Scholar] [CrossRef]
- Vieira, S.; De Camargo, P.B.; Selhorst, D.; Da Silva, R.; Hutyra, L.; Chambers, J.Q.; Brown, I.F.; Higuchi, N.; Dos Santos, J.; Wofsy, S.C.; et al. Forest structure and carbon dynamics in Amazonian tropical rain forest. Oecologia 2004, 140, 468–479. [Google Scholar] [CrossRef]
- Malhi, Y.; Wood, D.; Baker, T.R.; Wright, J.; Phillips, O.L.; Cochrane, T.; Meir, P.; Chave, J.; Almeida, S.; Arroyo, L.; et al. The regional variation of aboveground live biomass in old-growth Amazonian forests. Glob. Change Biol. 2006, 12, 1107–1138. [Google Scholar] [CrossRef]
- Saatchi, S.S.; Houghton, R.A.; Dos Santos Alvala, R.C.; Soares, J.V.; Yu, Y. Distribution of aboveground live biomass in the Amazon basin. Glob. Change Biol. 2007, 13, 816–837. [Google Scholar] [CrossRef]
- Hunter, M.O.; Keller, M.; Morton, D.C.; Cook, B.D.; Lefsky, M.A.; De Oliveira Junior, R.C. Structural dynamics of tropical moist forest gaps. PLoS ONE 2015, 10, e0132144. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; Hedblom, M.; Emilsson, T.; Nielsen, A.B. The role of forest stand structure as biodiversity indicator. For. Ecol. Manag. 2014, 330, 82–93. [Google Scholar] [CrossRef]
- Skovsgaard, J.P.; Vanclay, J.K. Forest site productivity: A review of the evolution of dendrometric concepts for even-aged stands. Forestry 2008, 81, 13–31. [Google Scholar] [CrossRef]
- Tabarelli, M.; Montovani, W. Clareiras naturais e a riqueza de espécies pioneiras em uma floresta Atlântica montana. Rev. Bras. De Biol. 1999, 59, 251–256. [Google Scholar] [CrossRef]
- Kohyama, T. Simulating stationary size distribution of trees in rain forests. Ann. Bot. 1991, 68, 173–180. [Google Scholar] [CrossRef]
- Souza, D.R.; Souza, A.L. Estratificação vertical em floresta ombrófila densa de terra firme não explorada, Amazônia oriental. Rev. Árvore 2004, 28, 691–698. [Google Scholar] [CrossRef]
- Ibge. Manual Técnico da Vegetação Brasileira, 2nd ed.; Série Manuais Técnicos em Geociências; IBGE: Rio de Janeiro, Brazil, 2012; Volume 1. [Google Scholar]
- Ibge. Manual Técnico de Pedologia, 3rd ed.; Série Manuais Técnicos em Geociências; IBGE: Rio de Janeiro, Brazil, 2015; Volume 1. [Google Scholar]
- Alvares, C.A.; Stape, J.L.; Sentelhas, P.C.; Gonçalves, J.D.; Sparovek, G. Köppen’s climate classification map for Brazil. Meteorol. Z. 2014, 22, 711–728. [Google Scholar] [CrossRef]
- Guedes, M.C.; Tonini, H.; Wadt, L.H.O.; Silva, K.E. Instalação e medição de parcelas permanentes para estudos com produtos florestais não madeireiros. In Produtos Florestais não Madeireiros—Guia Metodológico da Rede Kamukaia; Wadt, L.H.O., Santos, L.M.H., Bentes, M.P.M., Oliveira, V.B.V., Eds.; Embrapa: Brasília, Brazil, 2017; pp. 13–32. [Google Scholar]
- Macarthur, R.H.; Horn, J.W. Foliage profiles by vertical measurements. Ecology 1969, 50, 802–804. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: https://www.R-project.org/ (accessed on 5 June 2020).
- Smith, M.N.; Stark, S.C.; Taylor, T.C.; Ferreira, M.L.; De Oliveira, E.; Restrepo-Coupe, N.; Chen, S.; Woodcock, T.; Bentes dos Santos, D.; Alves, L.F.; et al. Seasonal and drought-related changes in leaf area profiles depend on height and light environment in an Amazon forest. New Phytol. 2019, 222, 741–757. [Google Scholar] [CrossRef]
- Wirth, R.; Weber, B.; Ryel, J.R. Spatial and temporal variability of canopy structure in a tropical moist forest. Acta Oecologica 2001, 22, 235–244. [Google Scholar] [CrossRef]
- Fayolle, A.; Doucet, J.L.; Gillet, J.F.; Bourland, N.; Lejeune, P. Tree allometry in Central Africa: Testing the validity of pantropical multi-species allometric equations for estimating biomass and carbon stocks. For. Ecol. Manag. 2012, 274, 119–131. [Google Scholar] [CrossRef]
- Slik, J.W.F.; Aiba, S.-I.; Brearley, F.Q.; Cannon, C.H.; Forshed, O.; Kitayama, K.; Nagamasu, H.; Nilus, R.; Payne, J.; Paoli, G.; et al. Environmental correlates of tree biomass, basal area, wood density, and stem density gradients in Bornean tropical forests. Glob. Ecol. Biogeogr. 2010, 19, 50–60. [Google Scholar] [CrossRef]
- Ryan, C.M.; Williams, M.; Hill, T.C.; Grace, J. Forests and carbon storage in tropical savannas: A win–win scenario? Carbon Balance Manag. 2011, 6, 6. [Google Scholar] [CrossRef]
- Neves, E.S.; Guedes, M.C.; Rodrigues, E.G. Relação da produção de frutos de castanha-da-amazônia (Bertholletia excelsa Bonpl.) com variáveis das próprias castanheiras, em capoeira e floresta da RESEX Cajari. Biota Amaz. 2015, 5, 31–37. [Google Scholar] [CrossRef]
- Shepard, G.H., Jr.; Ramirez, H.; Shepard, G.H. Ecological and biogeographical factors shaping Bertholletia excelsa populations in Amazonia. J. Trop. Ecol. 2019, 35, 149–161. [Google Scholar]
- Reis, C.R.; Aragão, L.E.O.C.; Silva, C.A.; Coe, M.T. Regional environmental gradients and their role in shaping Amazonian forest structure. Glob. Ecol. Biogeogr. 2018, 27, 1122–1132. [Google Scholar]
- Jucker, T.; Bongalov, B.; Burslem, D.F.R.P.; Nilus, R.; Dalponte, M.; Lewis, S.L.; Coomes, D.A. Topography shapes the structure, composition and function of tropical forest landscapes. Ecol. Lett. 2018, 21, 989–1000. [Google Scholar] [CrossRef]
- Tourne, D.C.M.; Ballester, M.V.R.; James, P.M.A.; Martorano, L.G.; Guedes, M.C.; Thomas, E. Strategies to optimize modeling habitat suitability of Bertholletia excelsa in the Pan-Amazonia. Ecol. Evol. 2019, 9, 12623–12638. [Google Scholar] [CrossRef]
- Laurance, W.F.; Ferreira, L.V.; Rankin-de Merona, J.M.; Laurance, S.G. Biomass collapse in Amazonian forest fragments. Science 1997, 278, 1117–1118. [Google Scholar] [CrossRef]
- Reis, C.R.; Jackson, T.D.; Gorgens, E.B.; Dalagnol, R.; Jucker, T.; Nunes, M.H.; Ometto, J.P.; Aragão, L.E.O.C.; Rodriguez, L.C.E.; Coomes, D.A. Forest disturbance and growth processes are reflected in the geographical distribution of large canopy gaps across the Brazilian Amazon. J. Ecol. 2022, 110, 2971–2983. [Google Scholar] [CrossRef]

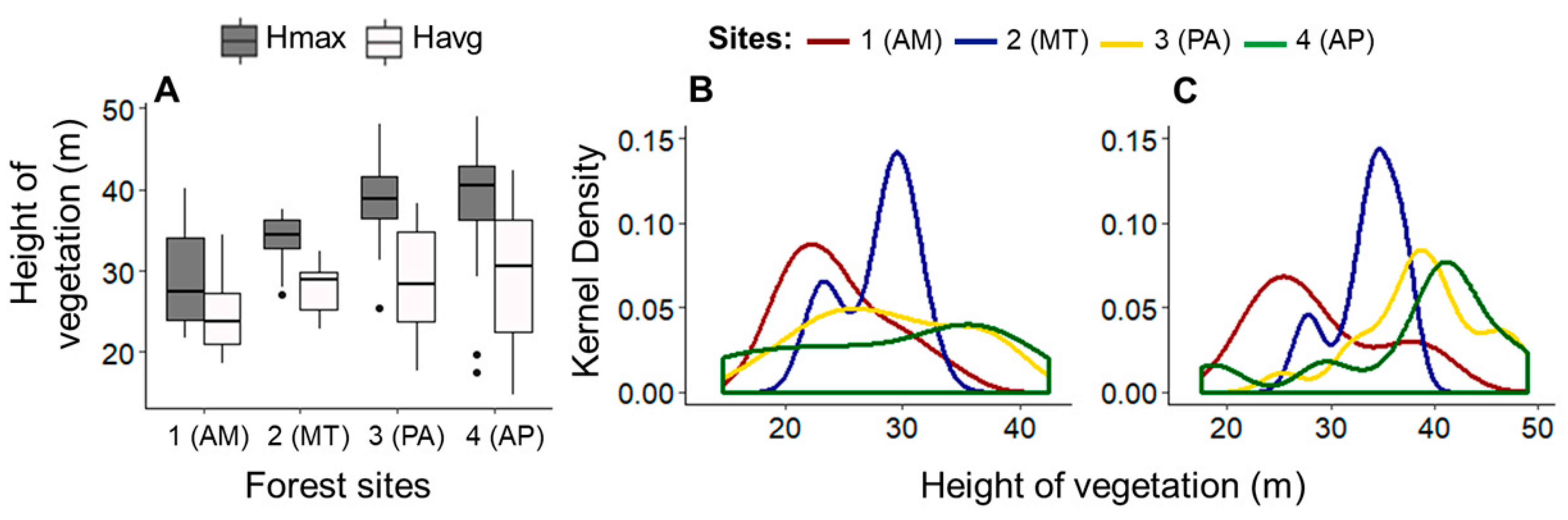

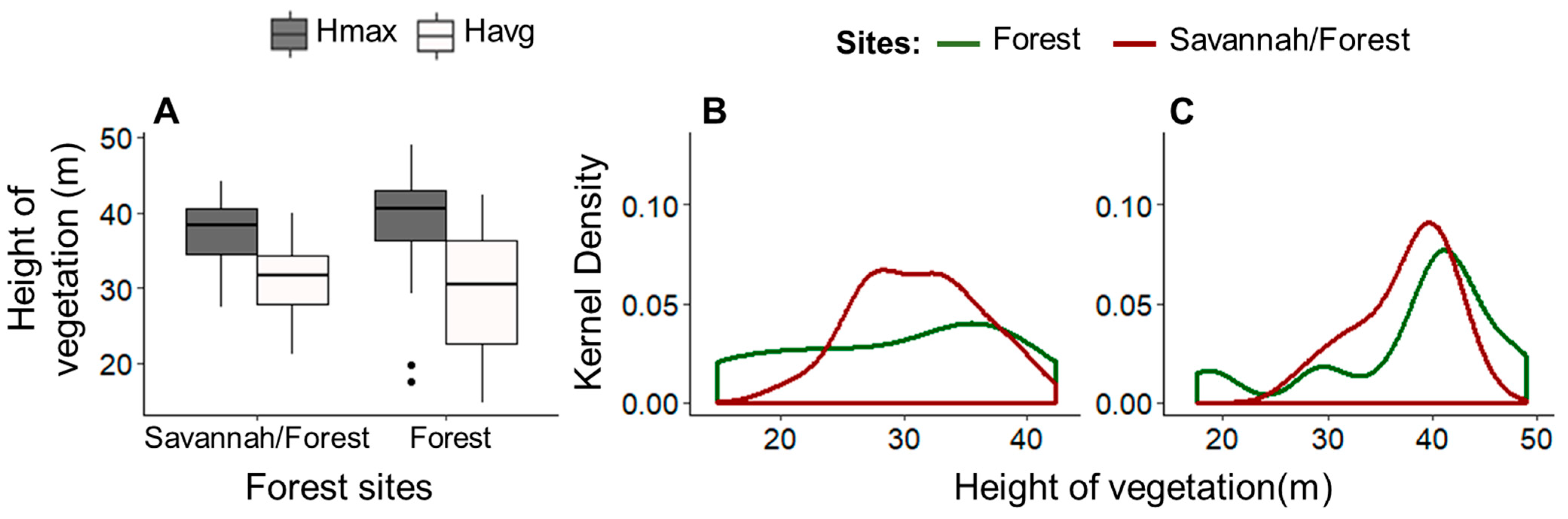
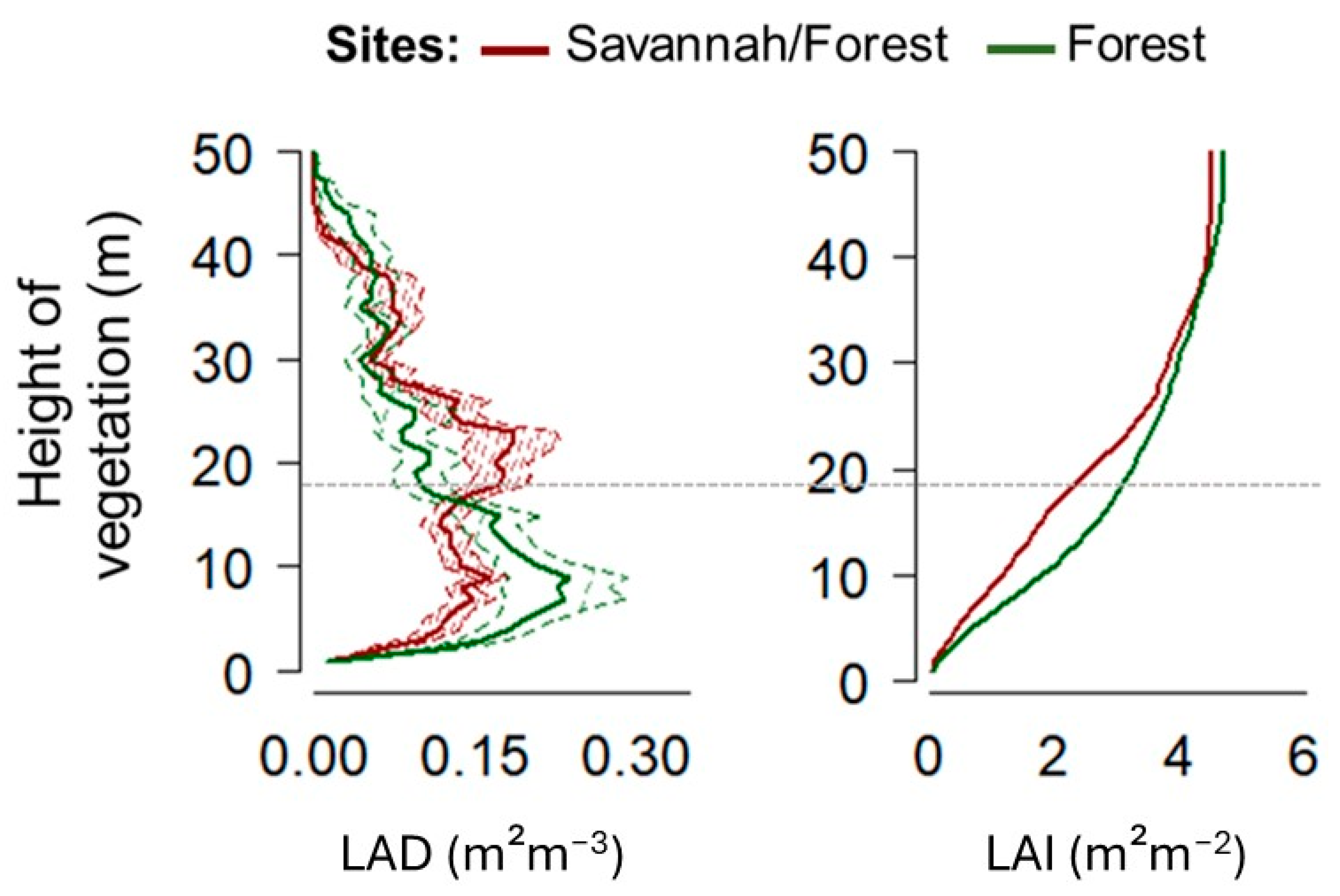
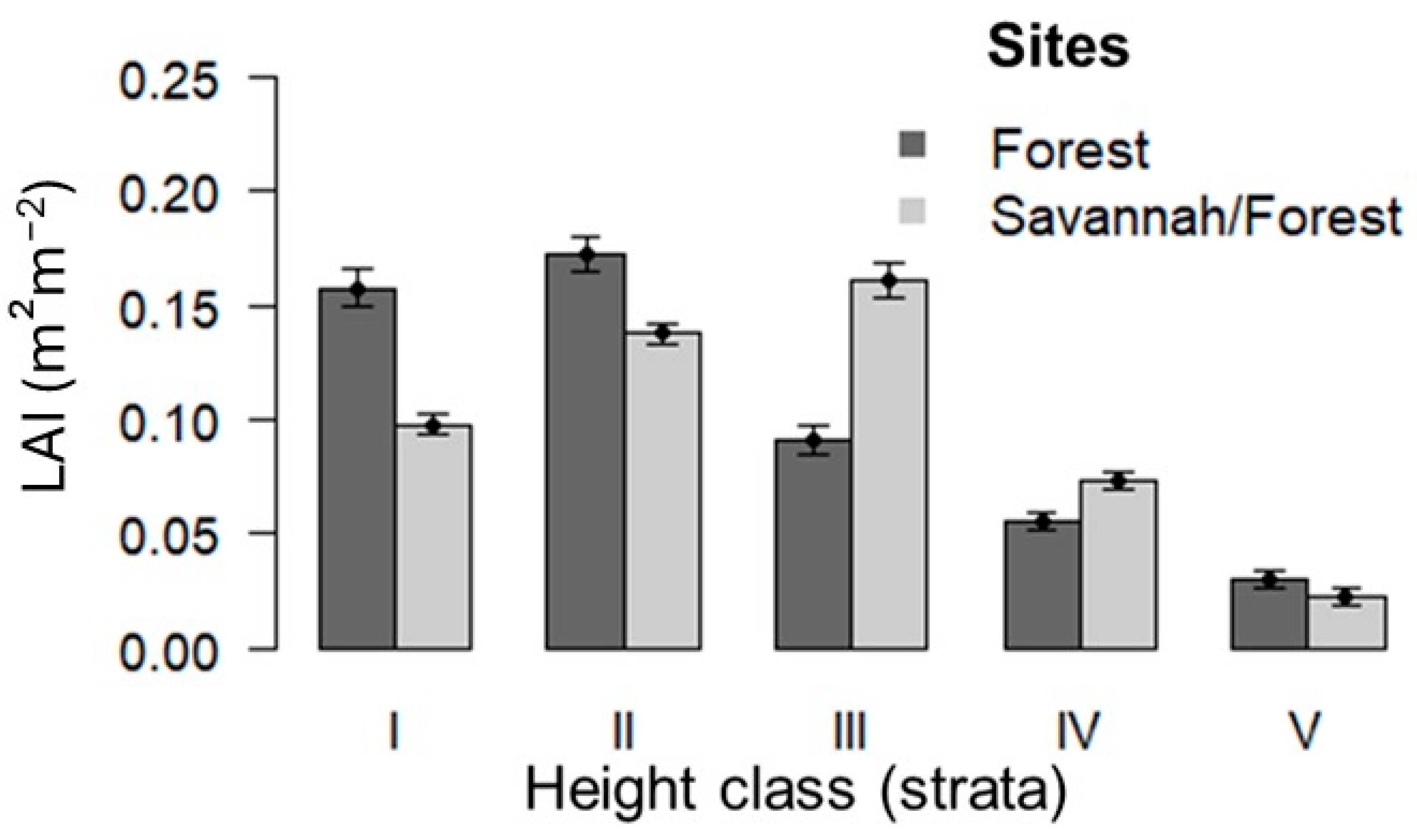


| Forest Sites | Brazil Nut | Matrix Vegetation | |||||
|---|---|---|---|---|---|---|---|
| D (ind. ha−1) | Havg | Hmax | HmaxA | Havg | Hmax | Hmax A | |
| AM | 5 | 33 | 36 | 49 | 25 | 29 | 40 |
| MT | 11 | 31 | 34 | 41 | 28 | 34 | 38 |
| PA | 10 | - | - | - | 29 | 39 | 48 |
| AP | 12 | 38 | 42 | 51 | 29 | 38 | 49 |
| Metrics | Forest Sites (M ± SD) | Test Kruskal–Wallis | ||||
|---|---|---|---|---|---|---|
| 1 (AM) | 2 (MT) | 3 (PA) | 4 (AP) | W | p-Value | |
| Hmax | 29.1 ± 6.20 | 33.7 ± 3.18 | 38.8 ± 5.80 | 38.1 ± 8.77 | 23.3 | <0.01 |
| Havg | 24.6 ± 4.46 | 27.9 ± 3.14 | 28.8 ± 6.55 | 28.9 ± 9.03 | 5.85 | 0.120 |
| R | 3.98 ± 1.79 | 4.90 ± 1.39 | 7.51 ± 3.45 | 7.54 ± 2.54 | 23.9 | <0.01 |
| S (%) | 1.82 ± 1.79 | 2.51 ± 4.91 | 0.64 ± 0.97 | 3.71 ± 5.39 | 16.0 | 0.001 |
| F (10m) | 0.01 ± 0.02 | 0.00 ± 0.01 | 0.06 ± 0.13 | 0.05 ± 0.08 | 6.56 | 0.087 |
| F (15m) | 0.03 ± 0.05 | 0.03 ± 0.04 | 0.12 ± 0.18 | 0.15 ± 0.20 | 3.72 | 0.292 |
| LAI I | 1.16 ± 0.35 | 0.80 ± 0.26 | 1.42 ± 0.44 | 1.10 ± 0.57 | 19.2 | <0.01 |
| LAI II | 2.29 ± 0.60 | 2.25 ± 0.39 | 2.04 ± 0.77 | 1.90 ± 0.57 | 5.12 | 0.163 |
| LAI III | 1.31 ± 0.57 | 1.05 ± 0.31 | 1.00 ± 0.44 | 0.73 ± 0.61 | 9.32 | 0.025 |
| LAI IV | 0.30 ± 0.36 | 0.82 ± 0.45 | 0.83 ± 0.71 | 0.49 ± 0.39 | 13.7 | 0.003 |
| LAI V | 0.04 ± 0.09 | 0.01 ± 0.03 | 0.30 ± 0.48 | 0.44 ± 0.50 | 18.6 | <0.01 |
| LAI IV + V | 0.30 ± 0.38 | 0.82 ± 0.45 | 1.12 ± 1.05 | 0.94 ± 0.69 | 14.6 | 0.002 |
| LAI General | 5.11 ± 0.49 | 4.94 ± 0.51 | 5.58 ± 0.98 | 4.65 ± 0.68 | 10.9 | 0.012 |
| Forest Sites | Brazil Nut | Matrix Vegetation | |||||
|---|---|---|---|---|---|---|---|
| D (ind. ha−1) | Havg | Hmax | Hmax A | Havg | Hmax | Hmax A | |
| Forest | 12 | 38 | 42 | 51 | 29 | 38 | 49 |
| Savannah/Forest | 17 | 36 | 38 | 42 | 31 | 37 | 44 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costa, F.F.; Oliveira Júnior, R.C.d.; de Almeida, D.R.A.; Rosa, D.M.; da Silva, K.E.; Tonini, H.; Beldini, T.P.; Santos, D.B.d.; Guedes, M.C. Terrestrial LiDAR Technology to Evaluate the Vertical Structure of Stands of Bertholletia excelsa Bonpl., a Species Symbol of Conservation Through Sustainable Use in the Brazilian Amazon. Sustainability 2025, 17, 6049. https://doi.org/10.3390/su17136049
Costa FF, Oliveira Júnior RCd, de Almeida DRA, Rosa DM, da Silva KE, Tonini H, Beldini TP, Santos DBd, Guedes MC. Terrestrial LiDAR Technology to Evaluate the Vertical Structure of Stands of Bertholletia excelsa Bonpl., a Species Symbol of Conservation Through Sustainable Use in the Brazilian Amazon. Sustainability. 2025; 17(13):6049. https://doi.org/10.3390/su17136049
Chicago/Turabian StyleCosta, Felipe Felix, Raimundo Cosme de Oliveira Júnior, Danilo Roberti Alves de Almeida, Diogo Martins Rosa, Kátia Emídio da Silva, Hélio Tonini, Troy Patrick Beldini, Darlisson Bentes dos Santos, and Marcelino Carneiro Guedes. 2025. "Terrestrial LiDAR Technology to Evaluate the Vertical Structure of Stands of Bertholletia excelsa Bonpl., a Species Symbol of Conservation Through Sustainable Use in the Brazilian Amazon" Sustainability 17, no. 13: 6049. https://doi.org/10.3390/su17136049
APA StyleCosta, F. F., Oliveira Júnior, R. C. d., de Almeida, D. R. A., Rosa, D. M., da Silva, K. E., Tonini, H., Beldini, T. P., Santos, D. B. d., & Guedes, M. C. (2025). Terrestrial LiDAR Technology to Evaluate the Vertical Structure of Stands of Bertholletia excelsa Bonpl., a Species Symbol of Conservation Through Sustainable Use in the Brazilian Amazon. Sustainability, 17(13), 6049. https://doi.org/10.3390/su17136049