Abstract
Walker Glacier near the northern coast of Ellesmere Island in the Canadian High Arctic (terrestrial margin of the ‘Last Ice Area’) is undergoing rapid ice attrition in response to climate change. We applied culture and molecular methods to investigate fungal diversity at the terminus of this glacier. Analysis of the mycoflora composition showed that the Walker Glacier isolates separated into two clusters: the surface of the glacier ice and the glacier foreland. The recently exposed sediments of the foreland had a lower fungal diversity and different species from those on the ice, with the exception of five species that occurred in both habitats. This loss of glacial ice in the Arctic is therefore resulting in the loss of habitats for cold-dwelling fungal species. Fungal diversity is a potentially rich biological resource of glacial ecosystems, with unique taxa. The rapid loss of these glacial habitats underscores the urgency for genomic surveys of fungal diversity in the High Arctic, and the need for further isolation of strains as well as cryopreservation of environmental micro-biome samples for future research and conservation.
1. Introduction
Glaciers currently occupy approximately 10% of the Earth’s surface [1], but they are now retreating and shrinking throughout the world [2] as a consequence of global climate change [3,4]. For example, glaciers in the Alps have lost about 50% of their surface area during the past 150 years [5]. Austre Brøggerbreen (79° N, 12° E), located near Ny-Ålesund in the Svalbard archipelago, Norway, lost approximately 1.86 km2 of ice between 1936 and 2010 [6], and the glacier has retreated by 480 m over the last 20 years [7]. Glaciers provide habitats for microbial communities in the cryosphere, and they supply water to streams, rivers, wetlands, and coastal oceans [8]. Glacial ice communities may differ from those found in glacier foreland habitats [9]. As glaciers retreat, rocks and sediments with less available nutrients are released and uncovered from the ice [6,10]. These substrates provide a habitat for bacteria and fungi that can grow under cold oligotrophic soil conditions [11,12]. Fungi act as decomposers and play key biogeochemical roles such as nutrient cycling in ecosystems [13], and losses in their diversity are therefore of broader concern for the functioning of the biosphere.
Walker Glacier (unofficial name; 83°00′ N, 72°12′ W) is located on the far northern coast of Ellesmere Island in the Canadian High Arctic (Figure 1). This region is at the northern limit of Quttinirpaaq National Park, Nunavut, where climate-related effects on the cryosphere have been observed over the past two decades [14,15,16]. This coastal zone is also the terrestrial margin of the ‘Last Ice Area’ (LIA), which contains the thickest ice in the Arctic Ocean and is viewed as the ultimate refuge for certain polar species [17]. However, concerns have been expressed about the rapid warming and ice attrition taking place in the LIA, and the loss of ice-dependent habitats at its coastal margin [18]. At the Walker Glacier site, GPS measurements were taken on 20 July 2013 from a datum pole placed at this location by Paul T. Walker on 10 July 1959 (Figure 1; 83°00′ N, 72°12′ W), showed that the glacier had retreated by 71 m over the preceding 54 years at an average rate of 1.3 m/year. Repeated GPS measurements at this location during the current survey (21 July 2016) showed that it had retreated an additional 10 m at an average rate of 3.3 m/year [19]. These observations suggested that Walker Glacier is in a retreating phase and that the rate of retreat is accelerating.
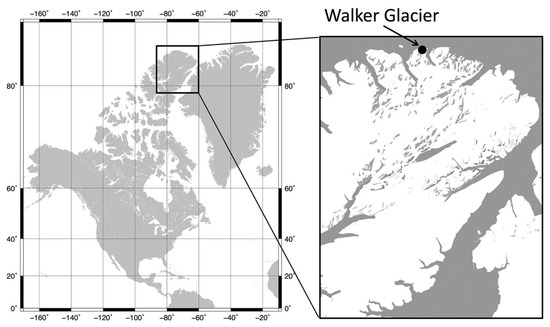
Figure 1.
Location map of Ellesmere Island and Walker Glacier in the Canadian High Arctic.
In our previous study, four new basidiomycete yeasts were described from this coastal LIA region, and two of the four new species were found at the Walker Glacier site [19,20,21,22]. To examine further the fungal taxa inhabiting glacier termini that are likely to be affected by global warming, we extended this work to the full communities by fungal culturing and molecular identification methods, to evaluate the diversity of mycoflora on Walker Glacier and its glacier foreland.
2. Materials and Methods
2.1. Fungal Isolation
Sediment samples were aseptically scraped from the surface of the melting ice face and terminal deposits of Walker Glacier (unofficial name; 83°00′ N, 72°12′ W), northern Ellesmere Island, NU, Canada, on 16 July 2016, and transferred to sterile 5-mL tubes; other sites and sampling details are given in Tsuji et al. [19]. Two sites were located on the glacial ice: Site 0 at 0.3 m from the base of the glacier and Site 1 at the base of the glacier. An additional seven sites were located on the exposed ground below the glacier. Sites 2 and 3 were 25 m and 40 m from the glacier terminus, respectively. Site 4 was located 55 m from the terminus of the glacier in an area that had been uncovered by the glacier retreat over the last 50 years. Site 5 was located approximately 71 m from the glacier, which was the position of the glacier terminus in 1959. Sites 6, 7 and 8 were located at 85 m, 100 m and 132 m, respectively, from the glacier terminus (Table 1). Within one hour after sampling, the samples were transferred to a −20 °C freezer and stored at this temperature until analysis.

Table 1.
Sampling site coordinates and distance from the glacier terminus of Walker Glacier.
Preliminary culture tests of Walker Glacier samples with potato dextrose agar (PDA), yeast malt extract agar (YMA), yeast peptone dextrose agar (YpDA), and Czapeck-Dox agar showed that PDA produced the best growth of the largest number of taxa, including those that grew on the other media. We therefore used PDA for all subsequent fungal culture experiments.
Fungal isolates were cultured from the nine samples. The sediment subsamples (0.1 g) were directly plated onto PDA (Difco, Becton Dickinson Japan, Tokyo, Japan) containing 50 μg/mL chloramphenicol and incubated at 10 °C for up to 3 weeks. Fungal colonies of different morphologies were streaked onto new PDA plates until individual colonies were isolated.
2.2. DNA Sequencing, Species Identification, and Data Analysis
A disk of 3 mm in diameter was taken from each fungal colony and placed into 1.5 mL tubes with 100 μL of sterile distilled water. DNA was extracted from the fungal colonies using a NucleoSpin Microbial DNA kit (Takara Bio Inc., Shiga, Japan) following the manufacturer’s protocol. The extracted DNA was then amplified by polymerase chain reaction (PCR) using KOD-plus DNA polymerase (Toyobo, Osaka, Japan) with the following primers: ITS1F (5′-GTAACAAGGTTTCCGT) [23] and NL4 (5′-GGTCCGTGTTTCAAGACGG). These primers are for two DNA sequences that are widely used in molecular fungal taxonomy: the ITS region and the D1/D2 domain of the LSU rDNA gene. The PCR conditions were as follows: primary template melting, 5 min at 94 °C; 35 cycles of 10 s at 98 °C (melting), 30 s at 50–65 °C (primer annealing) and 90 s at 68 °C (elongation); and 10 min at 68 °C (final elongation). PCR was carried out using an Eppendorf Master Cycler Nexus (Eppendorf Japan, Tokyo, Japan). The amplified DNA fragments were purified using Sephacryl S-400HR (Sigma-Aldrich, Tokyo, Japan). The DNA sequences were determined using an ABI Prism 3130xl sequencer (Applied Biosystems, Life Technologies, Tokyo, Japan), and the sequences were deposited in GenBank. The GenBank accession numbers of all the sequences analysed in this study are listed in Supplemental Table S1. The fungal species identifications by BLAST analysis were based on sequence homology.
In order to assess the community similarities among sampling sites based on the ITS region and D1/D2 domain of the LSU rDNA gene, a hierarchical cluster analysis of the fungal communities was carried out using Bray–Curtis similarity and a dendrogram inferred with the average linkage method [24]. These analyses were performed in R using the vegan package.
3. Results and Discussion
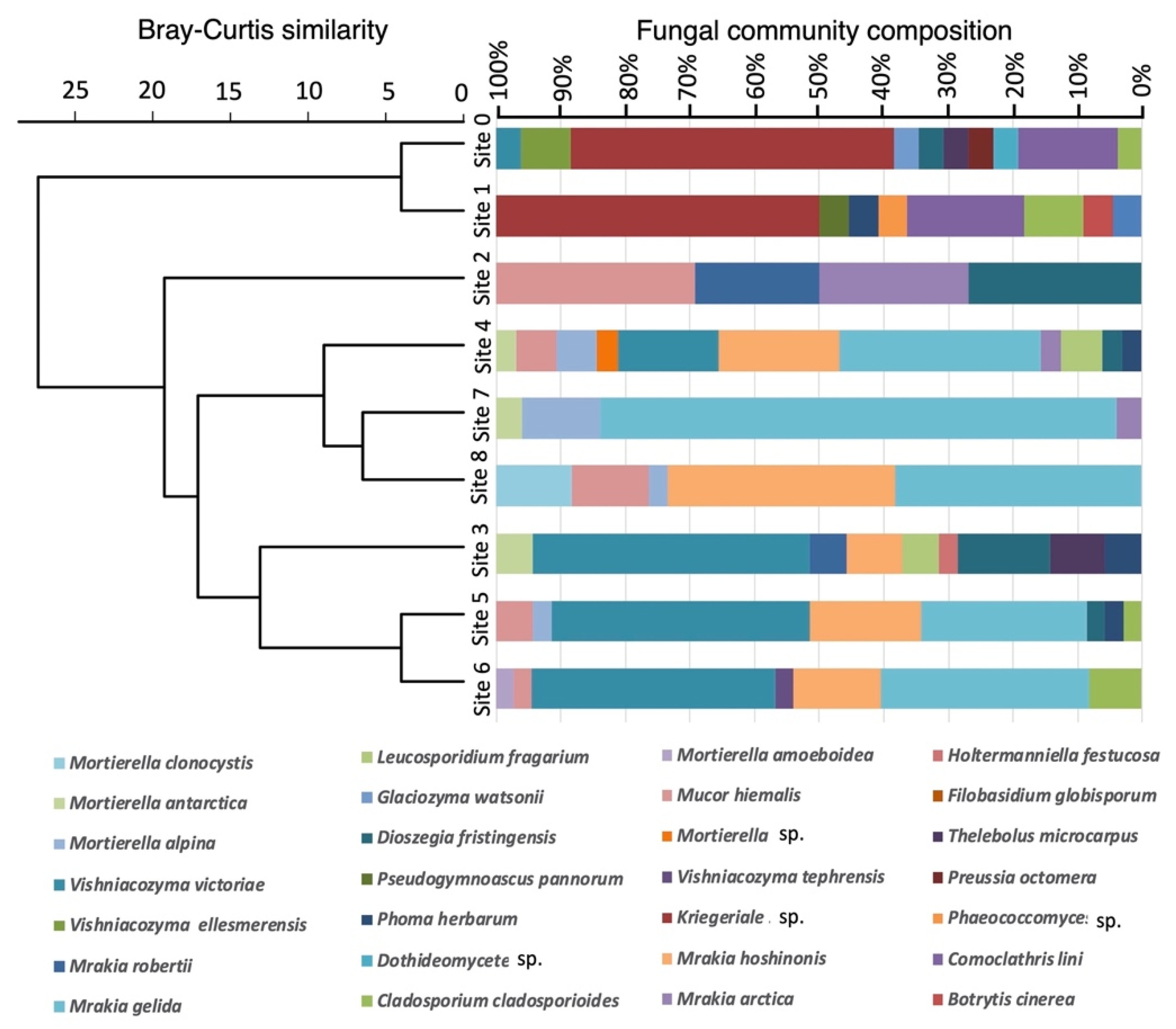
In total, 325 fungal strains were isolated from the nine glacial sediment samples collected from Walker Glacier (Table 1), and of these 273 were successfully extracted and analysed by DNA sequencing. There were 23–37 fungal strains obtained from each sampling site. Based on the ITS region sequences and D1/D2 domain sequences of the LSU rDNA gene, the fungal strains were classified into 10 species of ascomycetes, 12 species of basidiomycetes, and 6 species of zygomycetes, including unclassified species (see below). The abundance and occurrence of the isolated fungal species among the nine sampling sites were statistically compared by hierarchical clustering. The mycoflora composition showed clustering between several pairs of adjacent sites (0 and 1; 7 and 8; 5 and 6), but there was an overall separation into two clusters: isolates from the glacier ice (Site 0 and Site 1), and isolates from sites 2 to 8 in the glacier foreland (Figure 2).
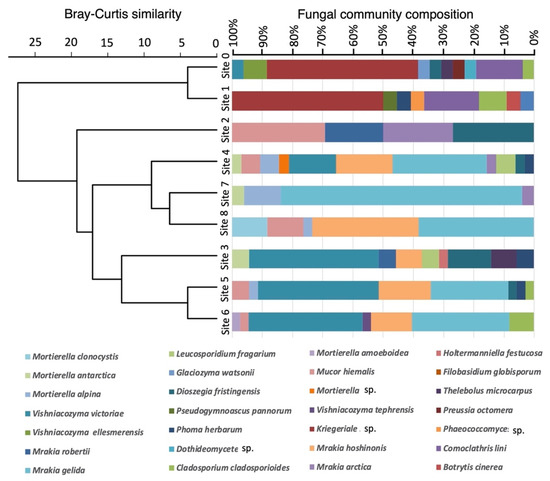
Figure 2.
Dendrogram based on Bray–Curtis dissimilarities among nine sites on Walker Glacier and its foreland.
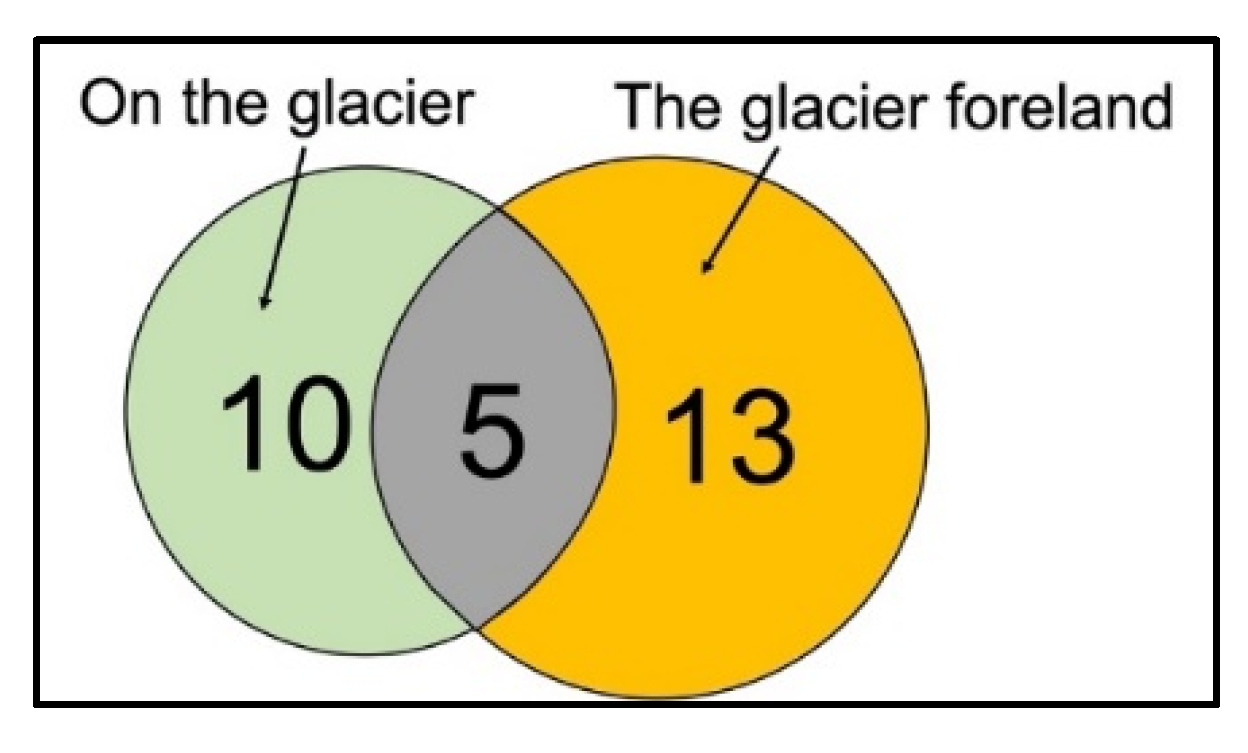
The origin of each isolate was subsequently examined according to the two location clusters. In total, we isolated 15 fungal species from the glacier and 18 from the glacier foreland. Only five of the 15 fungal species isolated from the glacier samples (Cladosporium cladosporioides, Dioszegia fristingensis, Phoma herbarum, Thelebolus microcarpus, and Vishniacozyma victoriae) were also isolated from the glacier foreland samples (Figure 3, Table 2).
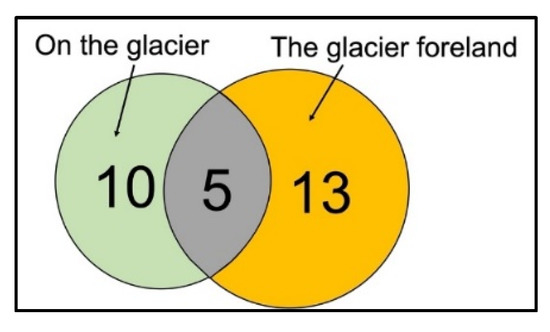
Figure 3.
Venn diagram showing overlapping and non-overlapping fungal species isolated from Walker Glacier and its foreland.
Table 2.
Taxonomy and number of strains isolated from each sampling site.
We previously isolated two basidiomycetous yeasts, M. hoshinonis, and V. ellesmerensis, from Walker Glacier and described them as new fungal species [19,22]. Mrakia hoshinonis has also been isolated from the Calderone Glacier in Italy and Iceland [25], however no sequences with high homology (>95%) in the ITS region with V. ellesmerensis have been registered to date in GenBank. These results suggest that M. hoshinonis may be widely distributed in Arctic and alpine regions while V. ellesmerensis may be more locally restricted and perhaps even endemic to the LIA region.
The UNITE database (https://unite.ut.ee/ (accessed on 21 December 2021)) was used to determine the homology between the ITS region sequence of the strains obtained here and sequences in the database [26]. This analysis indicated five potentially new fungal species from Walker Glacier and its glacier foreland. These were classified as Dothideomycetes sp., Ascomycota sp., Herpotrichiellaceae sp., Kriegerialess sp. and Mortierella sp. according to the sequence homology of ITS region and D1/D2 region of 26S RNA (Table S1). The UNITE database also provided information on the sampling location for fungi with high homology in the ITS region to our isolates. One strain of Dothideomycetes sp. (G2-4-2) from Walker Glacier had 100% sequence similarity to three strains of Dothideomycetes sp. in the database (accession numbers MF043960, MF043961, and MF043963). MF043960 and MF043963 were isolated from the Qaanaaq Glacier, in a Greenland cryoconite region, while MF043961 was isolated from the Yala Glacier in a Himalayan cryoconite region. Strain G2-4-2 also showed 100% similarity with three strains of Preussia sp. (accession numbers MK460382, MK460383, and KC333159) isolated from the Greenland Ice Sheet and cryoconite holes in a glacier in Svalbard, Norway [9,27]. A strain of Ascomycota (GR1-1-20-1) isolated from Walker Glacier had 98% ITS region sequence similarity to an uncultured fungal clone (FJ553913) isolated from soil near Skulow Lake, southern Canada [28], and to an uncultured fungal clone (FJ237066) from the snow-covered soil from the foreland of the Rotmoosferner Glacier in Austria [29]. A strain of Herpotrichiellaceae (GR1-3-20-3) from Site 1 at Walker Glacier showed 99% sequence homology to an uncultured fungal clone (KC965554) from Prince Patrick Island, Canada. Our Walker Glacier isolates also included 24 strains of Kriegeriales species, which appear to be a major component of the fungal cryo-community. These Kriegeriales strains had 88% ITS region sequence homology with a basidiomycete strain from an ice core obtained in the Altai Mountains, western Siberia (AB474394). From the glacier foreland, Mortierella sp. (GR4-1-20-5) had 99% ITS region sequence similarity with another uncultured fungus clone (KC965293) from Banks Island in Canada [30].
The five new fungal species from on Walker Glacier and the glacier foreland, and our strains showing high homology with Kriegeriales species were registered in the DNA databank of ITS region sequences. It may be that the Kriegeriales strain is also endemic to the LIA, as suggested for V. ellesmerensis. Most of the potentially new fungal species isolated from Walker Glacier (Sites 0 and 1) were not present among the isolates from the glacier foreland (Sites 2 to 8).
Although the PDA medium used in this study supported the growth of a wide range of species, it likely selected for only a subset of the total fungal community. More dilute media might favour more oligotrophic species, and determination of the true total diversity will require the application of next-generation molecular methods. In particular, metagenomic analysis would allow resolution of the entire glacier microbiome, including algae, bacteria, archaea and viruses, in addition to fungi, and may also reveal their functional diversity. Molecular analyses of other cryo-habitats have shown a rich fungal diversity living on ice. For example, amplicon analyses revealed 184 fungal taxa in samples from glacial ice in maritime Antarctica [31], and 697 fungal taxa from algal associated communities on the surface of the Greenland Ice Sheet [27]. Metagenomic analyses are likely to pick up additional species that elude even amplicon analysis (as found for prokaryotes in High Arctic lakes [32]), and full molecular surveys are urgently required throughout the fast changing cryosphere.
Fungi inhabiting the High Arctic must survive the extreme polar conditions of persistent cold temperatures, continuous bright light exposure, and winter-freeze-up, and are likely to have many unusual biochemical properties. Studies elsewhere in the cryosphere have drawn attention to the value of low-temperature fungi in the search for cold-active enzymes, materials for new drugs, bio-surfactants and other biotechnological applications (as described for Antarctic isolates [33]). These fungi represent a new bioresource that could contribute to sustainable development goals. The present study took place in the Inuit homeland of Nunavut, Canada, and any such commercial development would require close attention to Indigenous rights and benefit sharing.
The results of this study suggest a unique ice-dependent mycoflora on Walker Glacier, with strains that were not able to compete or survive in the glacier foreland environment. Fungi living on glaciers likely play key biogeochemical roles on these cryo-ecosystems in carbon and nutrient cycling [13] and may have unique biochemical and physiological characteristics. The ongoing retreat of High Arctic glaciers due to global warming, which is amplified at these high northern latitudes [34,35], is resulting in accelerated habitat loss and may lead to the eventual extinction of these extreme cold-adapted microbes. Elsewhere along the LIA margin of Canada and Greenland, the highest latitude coastal region of the Arctic, there is recent evidence of complete loss of specific ecosystem types, and widespread ice attrition [18]. In our study region on northern Ellesmere Island, glaciers on average retreated by 5% over the period 1999–2015, with the complete disappearance of three small ice caps [16]. The fungal taxa described here are therefore prone to ongoing climate warming and habitat loss, which underscores the urgency of greenhouse gas mitigation at a global scale as well as the importance of local conservation initiatives and biodiversity surveys. Part of that conservation effort should include genomic surveys and cryopreservation of environmental microbiome samples from High Arctic glaciers, as well as further isolation of fungal strains from these habitats for culture and analysis.
Supplementary Materials
The following are available online at www.mdpi.com/article/10.3390/su14031617/s1, Table S1: species names, strain names, sources of isolates, GenBank accession number of the ITS region, and D1/D2 domain sequences of isolated fungi.
Author Contributions
Conceived and designed the experiments: M.T., Y.T., W.F.V. and M.U.; performed the field sampling: Y.T. and W.F.V.; performed the laboratory experiments and analyses: M.T.; analysed the data: M.T.; wrote the manuscript: M.T. (with input from all authors). All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the NIPR Research Project (KP-309), a JSPS Grant-in-Aid for Young Scientists (A) to M. Tsuji (no. 16H06211), Institution for Fermentation, Osaka, by Young Scientists to M.T. (no. Y-2018-004), and the ArCS (Arctic Challenge for Sustainability) II project (Program Grant Number JPMXD1420318865) provided by the Ministry of Education, Culture, Sports, Science and Technology, Japan. Additional support for the fieldwork was provided through the project NEIGE (Northern Ellesmere Island in the Global Environment) funded through the Natural Sciences and Engineering Research Council of Canada (NSERC), the NCE program ArcticNet, and the CFREF program Sentinel North.
Acknowledgments
We thank M. Kumagai, Ritsumeikan University; A. I. Culley, Université Laval, and D. Sarrazin, CEN, for their assistance in the field. We are grateful to Parks Canada and CEN for the use of their facilities at Ward Hunt Island in Quttinirpaaq National Park, Nunavut, and the Polar Continental Shelf Program (PCSP) for aircraft support. The authors also thank K. Watanabe and M. Mori for providing technical assistance. This work is also a contribution to the International Arctic Science Committee (IASC) project T-MOSAiC (Terrestrial Multidisciplinary distributed Observatories for the Study of Arctic Connections).
Conflicts of Interest
The authors declare no competing interests.
References
- Cauvy-Fraunié, S.; Dangles, O. A global synthesis of biodiversity responses to glacier retreat. Nat. Ecol. Evol. 2019, 3, 1675–1685. [Google Scholar] [CrossRef]
- Zemp, M.; Huss, M.; Thibert, E.; EcKert, N.; McNabb, R.; Huber, J.; Barandun, M.; Machguth, H.; Nussbaumer, S.U.; Gärtner-Roser, I.; et al. Global glacier mass changes and their contributions to sea-level rise from 1961 to 2016. Nature 2019, 568, 382–386. [Google Scholar] [CrossRef]
- Qin, D.; Ding, Y. Key issues on cryospheric changes, trends and their impacts. Adv. Clim. Chang. Res. 2010, 1, 1–10. [Google Scholar] [CrossRef]
- IPCC. Special Report on the Ocean and Cryosphere in a Changing Climate. Available online: https://www.ipcc.ch/srocc/ (accessed on 21 December 2021).
- Zemp, M.; Haeberli, W.; Hoelzle, M.; Paul, F. Alpine glaciers to disappear within decades? Geophys. Res. Lett. 2006, 33, L13504. [Google Scholar] [CrossRef] [Green Version]
- Nowak, A.; Hodson, A. Changes in meltwater chemistry over a 20-year period following a thermal regime switch from polythermal to cold-based glaciation at Austre Brøggerbreen, Svalbard. Polar Res. 2014, 33, 22779. [Google Scholar] [CrossRef] [Green Version]
- Kume, A.; Nakatsubo, T.; Bekku, Y.; Masuzawa, T. Ecological significance of different growth forms of purple Saxifrage, Saxifraga oppositifolia L., in the High Arctic, Ny-Ålesund, Svalbard. Arct. Antarct. Alp. Res. 1999, 31, 27–33. [Google Scholar] [CrossRef]
- Sharp, M.; Parkes, J.; Cragg, B.; Fairchild, I.J.; Lamb, H.; Tranter, M. Widespread bacterial populations at glacier beds and their relationship to rock weathering and carbon cycling. Geology 1999, 27, 107–110. [Google Scholar] [CrossRef]
- Edwards, A.; Douglas, B.; Anesio, A.M.; Rassner, S.M.; Irvine-Fynn, T.D.L.; Sattler, B.; Griffith, G.W. A distinctive fungal community inhabiting cryoconite holes on glaciers in Svalbard. Fungal Ecol. 2013, 6, 168–176. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Richter, A.; Bol, R.; Garnett, M.H.; Bäumler, R.; Xu, X.; Lopez-Capel, E.; Manning, D.A.C.; Hobbs, P.J.; Hartley, I.R.; et al. Heterotrophic microbial communities use ancient carbon following glacial retreat. Biol. Lett. 2007, 3, 487–490. [Google Scholar] [CrossRef]
- Bergero, R.; Girlanda, M.; Varese, G.C.; Intili, D.; Luppi, A.M. Psychrooligotrophic fungi from arctic soils of Franz Joseph Land. Polar Biol. 1999, 21, 361–368. [Google Scholar] [CrossRef]
- Sattin, S.R.; Cleveland, C.C.; Hood, E.; Reed, S.C.; King, A.J.; Schmidt, S.K.; Robeson, M.S.; Ascarrunz, N.; Neergut, D.R. Functional shifts in unvegetated, perhumid, recently-deglaciated soils do not correlate with shifts in soil bacterial community composition. J. Microbiol. 2009, 47, 673–681. [Google Scholar] [CrossRef]
- Welander, U. Microbial degradation of organic pollutants in soil in a cold climate. Soil Sediment Contam. 2005, 14, 281–291. [Google Scholar] [CrossRef]
- Copland, L.; Mueller, D. (Eds.) Arctic Ice Shelves and Ice Islands; Springer: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Vincent, W.F.; Fortier, D.; Lévsque, E.; Boulanger-Lapointe, N.; Tremblay, B.; Sarrazin, D.; Antoniades, D.; Mueller, D.R. Extreme ecosystems and geosystems in the Canadian High Arctic: Ward Hunt Island and vicinity. Ecoscience 2011, 18, 236–261. [Google Scholar] [CrossRef]
- White, A.; Copland, L. Area change of glaciers across northern Ellesmere Island, Nunavut, between ~1999 and ~2015. J. Glaciol. 2018, 64, 609–623. [Google Scholar] [CrossRef] [Green Version]
- Newton, R.; Pfirman, S.; Tremblay, L.B.; De Repentigny, P. Defining the “ice shed” of the Arctic Ocean’s Last Ice Area and its future evolution. Earth’s Future 2021, 9, e2021EF001988. [Google Scholar] [CrossRef]
- Vincent, W.F.; Mueller, D. Witnessing ice habitat collapse in the Arctic. Science 2020, 370, 1031–1032. [Google Scholar] [CrossRef]
- Tsuji, M.; Tanabe, Y.; Vincent, W.F.; Uchida, M. Vishniacozyma ellesmerensis sp. nov., a new psychrophilic yeast isolated from a retreating glacier in the Canadian High Arctic. Int. J. Syst. Evol. Microbiol. 2019, 69, 696–700. [Google Scholar] [CrossRef]
- Tsuji, M.; Tanabe, Y.; Vincent, W.F.; Uchida, M. Mrakia arctica sp. nov., a new psychrophilic yeast isolated from an ice island in the Canadian High Arctic. Mycoscience 2018, 59, 54–58. [Google Scholar] [CrossRef]
- Tsuji, M.; Tanabe, Y.; Vincent, W.F.; Uchida, M. Gelidatrema psychrophila sp. nov., a novel yeast species isolated from an ice island in the Canadian High Arctic. Mycoscience 2018, 59, 67–70. [Google Scholar] [CrossRef]
- Tsuji, M.; Tanabe, Y.; Vincent, W.F.; Uchida, M. Mrakia hoshinonis sp. nov., a novel psychrophilic yeast isolated from a retreating glacier on Ellesmere Island in the Canadian High Arctic. Int. J. Syst. Evol. Microbiol. 2019, 69, 944–948. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest communities of southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Turchetti, B.; Sannino, C.; Mezzasoma, A.; Zucconi, L.; Onofri, S.; Buzzini, P. Mrakia stelviica sp. nov. and Mrakia montana sp. nov., two novel basidiomycetous yeast species isolated from cold environments. Int. J. Syst. Evol. Microbiol. 2020, 70, 704–4713. [Google Scholar] [CrossRef]
- Nilsson, R.H.; Larsson, K.H.; Taylor, A.F.S.; Bengtsson-Palme, J.; Jeppesen, T.S.; Schigel, D.; Kennedy, P.; Picard, K.; Glöckner, F.O.; Tedersoo, L.; et al. The UNITE database for molecular identification of fungi: Handling dark taxa and parallel taxonomic classifications. Nucleic Acids Res. 2018, 47, D259–D264. [Google Scholar] [CrossRef]
- Perini, L.; Gostinčar, C.; Anesio, A.M.; Williamson, C.; Tranter, M.; Gude-Cimerman, N. Darkening of the Greenland Ice Sheet: Fungal abundance and diversity are associated with algal bloom. Front. Microbiol. 2019, 10, 557. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, M.; Lee, S.; Hallam, S.J.; Mohn, W.W. Bacterial, archaeal and eukaryal community structures throughout soil horizons of harvested and naturally disturbed forest stands. Environ. Microbiol. 2009, 11, 3045–3062. [Google Scholar] [CrossRef]
- Kuhnert, R.; Oberkofler, I.; Peintner, U. Fungal growth and biomass development is boosted by plants in snow-covered soil. Microbiol. Ecol. 2012, 64, 79–90. [Google Scholar] [CrossRef]
- Timling, I.; Walker, D.A.; Nusbaum, C.; Lennon, N.J.; Taylor, D.L. Rich and cold: Diversity, distribution and drivers of fungal communities in patterned-ground ecosystems of the North American Arctic. Mol. Ecol. 2014, 23, 3258–3272. [Google Scholar]
- De Menezes, G.C.A.; Câmara, P.E.A.S.; Pinto, O.H.B.; Convey, P.; Carvalho-Silva, M.; Simões, J.C.; Rosa, C.A.; Rosa, L.H. Fungi in the Antarctic cryosphere: Using DNA metabarcoding to reveal fungal diversity in glacial ice from the Antarctic Peninsula Region. Microb. Ecol. 2021; in press. [Google Scholar] [CrossRef]
- Vigneron, A.; Cruaud, P.; Lovejoy, C.; Vincent, W.F. Genomic evidence of functional diversity in DPANN archaea, from oxic species to anoxic vampiristic consortia. ISME Comm. 2022, 2, 4. [Google Scholar] [CrossRef]
- De Menezes, G.C.A.; Porto, B.A.; Amorim, S.S.; Zani, C.L.; Alves, T.M.D.A.; Policarpo, A.S., Jr.; Murta, S.M.F.; Simões, J.C.; Cota, B.B.; Rosa, C.A.; et al. Fungi in glacial ice of Antarctica: Diversity, distribution and bioprospecting of bioactive compounds. Extremophiles 2020, 24, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Vincent, W.F. Arctic climate change: Local impacts, global consequences, and policy implications. In Palgrave Handbook of Arctic Policy and Politics; Coates, K., Holroyd, C., Eds.; Palgrave Macmillan: London, UK, 2020; pp. 507–526. [Google Scholar]
- Bégin, P.N.; Tanabe, Y.; Kumagai, M.; Culley, A.I.; Paquette, M.; Sarrazin, D.; Uchida, M.; Vincent, W.F. Extreme warming and regime shift toward amplified variability in a far northern lake. Limnol. Oceanogr. 2021, 66, S17–S29. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).