
Sustainable Food Production: Innovative Netting Concepts and Their Mode of Action on Fruit Crops

 ,
,  , , and
, , and 
Abstract

1. Introduction
2. Methods
3. Light Manipulation
4. Microclimate Conditions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Country | Source | Protected Plant | Netting | Air Temperature | Relative Humidity | Wind Speed |
|---|---|---|---|---|---|---|
| Australia | [7] | apple | anti-hail | ↓1–3 °C | ↑10–15% | ↓up to 50% |
| [24] | apple | gray quad (10% shading) | ↓0.5 °C | ↑1.8 (afternoon) and 3.8% (during night) | ↓22–24% (i.c.—upper part) | |
| Brasil | [50] | apple | black net (mesh size 4 mm × 7 mm) | m.d. (i.c.) | ic.r. (i.c.) | ↓30% (top of canopy) |
| Chile | [45] | grapevine | red and pearl (20% shading) | m.d. | m.d. | |
| Germany | [51] | apple | white and red-black (mesh size 3 mm × 9 mm and 2.5 mm × 6.5 mm, respectively) | ↓0.5 °C | ↑1–3% | |
| [32] | apple | white, white-red, red-black, and green-black (12, 14, 18, and 20% shading within PAR, respectively) | ↓1.3 °C | ↑2–5% | ||
| Italy | [41] | kiwifruit | blue, red, and gray (26.9, 22.8, and 27.3%, shading within PAR, respectively) | ↓0.1–0.9 °C | ||
| white (20.4% shading within PAR) | m.d. | |||||
| Israel | [49] | pepper | black (25–28% shade), black, dark-green, blue-silver, green-silver (40–45% shade) | ↓3–4 °C | ↑20–35% | |
| [16] | apple | red, blue, gray, and pearl net (30% shade) and white and red-white net (15% shade) | ↓3–6 °C (T-max) (i.c.) | |||
| [6] | apple, grape, etc. | red, yellow, blue, gray, black, and pearl net (30% shading) | ↓1–5 °C (T-max) (i.c.) | ↑3–10% (H-min) (i.c.) | m.d. (a.c.) or ↓ (2 m a.c.) | |
| [52] | mandarin | red, yellow, white, and transparent (25, 24, 18, and 13% shading, respectively) | m.d. or ↓up to 1 °C (T-max) | ↑ | ↓70% (a.c.) | |
| ↓85–90% (i.c.) | ||||||
| Peru | [31] | blueberry | white, red, gray, and black (35 and 50% shading) | m.d. | m.d. | |
| Serbia | [53] | highbush blueberry | gray (mesh size 2.8 mm × 8 mm) | ↓1.5 °C, 2.4 °C (T-max) | ↑1%, 4% (H-min) | |
| Slovenia | [29] | apple | red, gray, blue, green, and black (mesh size 8 mm × 4 mm) | m.d. | m.d. | ↓47–72% |
| South Africa | [39] | avocado | crystal (30% shading) | ↓1–5% | ↓ | |
| Spain | [42] | apple | black and crystal (mesh size 3 × 7.4 mm) | ↓3 °C (T-max) (i.c.) | ↑ (i.c.) | |
| [43,54] | lemon | aluminet (50% transmittance of incident light) | ↓6 °C (T-max) | |||
| [55] | apricot | mosquito (10% interior shading) | ↓20% | |||
| Taiwan | [56] | mandarin | white nylon (20% shading) | ↓3.8–5 °C | ||
| USA | [13] | / | red, blue, and pearl (50% shading) | ↑(T-max) | m.d. | ↓ |
| black (50% shading) | ↓0.1–0.9 °C (T-max) | m.d. | ↓ | |||
| [40] | apple | pearl, blue, and red (20 to 23% shading) | m.d. (a.c.), ↓0.1–0.9 °C (i.c.—pearl net) and m.d. (i.c.—other nets) | ↑0.5–1.1% (a.c.), ↑1–4% (i.c.), | ↓40% (a.c.) | |
| [44] | apple | white (30% shading) | ↓1 °C |
5. Vegetative Growth
6. Photosynthesis Efficiency
7. Production Parameters
8. Fruit Quality
8.1. Fruit Size
8.2. Fruit Color and Pigments
8.3. Fruit Firmness
8.4. Sugar Content in Fruits
8.5. Acid Content in Fruits
8.6. Bioactive Components in Fruits
9. Physiological Disorders—Sunburns
10. Shift in Fruit Ripening Time
11. Conclusions
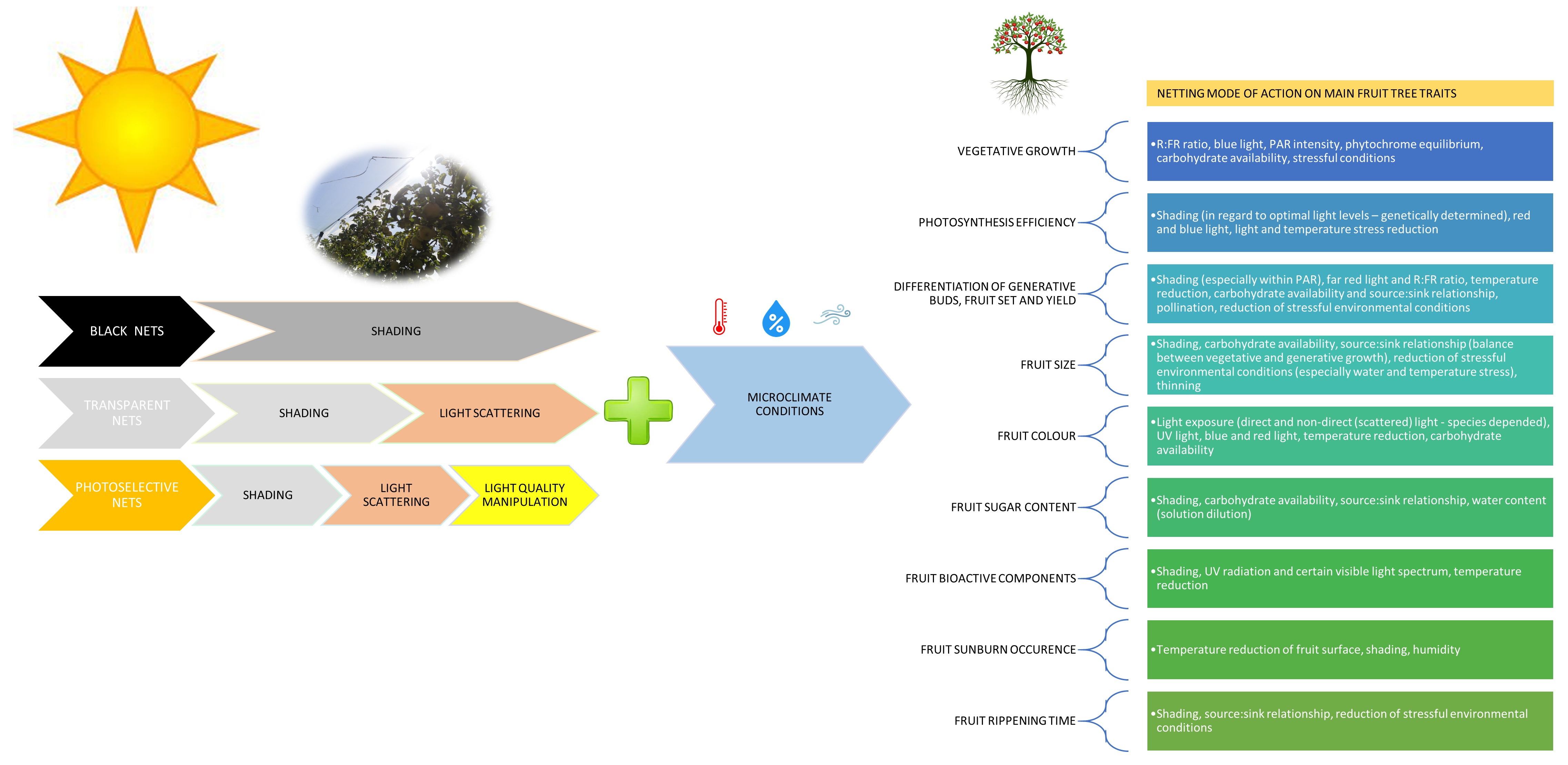
- Vegetative growth—R:FR ratio, blue light, PAR intensity, phytochrome equilibrium, carbohydrate availability, stressful conditions;
- Photosynthesis efficiency—shading (regarding optimal light levels—genetically determined), red and blue light, light and temperature stress reduction;
- Differentiation of generative buds, fruit set, and yield—shading (especially within PAR), far-red light and R:FR ratio, temperature reduction, carbohydrate availability, and source–sink relationship, pollination, reduction in stressful environmental conditions;
- Fruit size—shading, carbohydrate availability, source–sink relationship (balance between vegetative and generative growth), reduction in stressful environmental conditions (especially water and temperature stress), thinning;
- Fruit color—light exposure (direct and non-direct (scattered) light—species-dependent), UV light, blue and red light, temperature reduction, carbohydrate availability;
- Fruit sugar content—shading, carbohydrate availability, source–sink relationship, water content (solution dilution);
- Fruit bioactive components—shading, UV radiation, and certain visible light spectra, temperature reduction;
- Fruit sunburn occurrence—temperature reduction on fruit surface, shading, humidity;
- Fruit ripening time—shading, source–sink relationship, reduction in stressful environmental conditions.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shahak, Y. Photoselective netting: An overview of the concept, R&D and practical implementation in agriculture. Acta Hortic. 2014, 1015, 155–162. [Google Scholar]
- Briassoulis, D.Ã.; Mistriotis, A.; Eleftherakis, D. Mechanical behaviour and properties of agricultural nets. Part II: Analysis of the performance of the main categories of agricultural nets. Polym. Test. 2007, 26, 970–984. [Google Scholar] [CrossRef]
- Castellano, S.; Scarascia Mugnozza, G.; Russo, G.; Briassoulis, D.; Mistriotis, A.; Hemming, S.; Waaijenberg, D. Plastic Nets in Agriculture: A General Review of Types and Applications. Appl. Eng. Agric. 2008, 24, 799–808. [Google Scholar] [CrossRef]
- Bosco, L.C.; Bergamaschi, H.; Cardoso, L.S.; de Paula, V.A.; Marodin, G.A.B.; Nachtigall, G.R. Apple production and quality when cultivated under anti-hail cover in Southern Brazil. Int. J. Biometeorol. 2015, 59, 773–782. [Google Scholar] [CrossRef]
- Lobos, G.A.; Retamales, J.B.; Hancock, J.F.; Flore, J.A.; Romero-bravo, S.; Pozo, A. Productivity and fruit quality of Vaccinium corymbosum cv. Elliott under photo-selective shading nets. Sci. Hortic. 2013, 153, 143–149. [Google Scholar] [CrossRef]
- Shahak, Y.; Gussakovsky, E.E.; Gal, E.; Ganelevin, R. ColorNets: Crop protection and light-quality manipulation in one technology. Acta Hortic. 2004, 659, 143–151. [Google Scholar] [CrossRef]
- Middleton, S.; McWaters, A. Hail netting of apple orchards: Australian experience. Compact Fruit Tree 2002, 35, 51–55. [Google Scholar]
- Giaccone, M.; Forlani, M.; Basilea, B. Tree vigor, fruit yield and quality of nectarine trees grown under red photoselective anti- hail nets in Southern Italy. Acta Hortic. 2012, 962, 387–394. [Google Scholar] [CrossRef]
- Pajač Živković, I.; Jemrić, T.; Fruk, M.; Buhin, J.; Barić, B. Influence of different netting structures on codling moth and apple fruit damages in Northwest Croatia. Agric. Conspec. Sci. 2016, 81, 99–102. [Google Scholar]
- Pajač Živković, I.; Jemrić, T.; Fruk, M.; Barić, B. Upotreba fotoselektivnih mreža u zaštiti od važnih štetnika breskve. Glas. Biljn. Zašt. 2018, 18, 399–406. [Google Scholar]
- Sauphanor, B.; Severac, G.; Maugin, S.; Toubon, J.F.; Capowiez, Y. Exclusion netting may alter reproduction of the codling moth (Cydia pomonella) and prevent associated fruit damage to apple orchards. Entomol. Exp. Appl. 2012, 145, 134–142. [Google Scholar] [CrossRef]
- Tasin, M.; Demaria, D.; Ryne, C.; Cesano, A.; Galliano, A.; Anfora, G.; Ioriatti, C.; Alma, A. Effect of anti-hail nets on Cydia pomonella behavior in apple orchards. Entomol. Exp. Appl. 2008, 129, 32–36. [Google Scholar] [CrossRef]
- Arthurs, S.P.; Stamps, R.H.; Giglia, F.F. Environmental modification inside photoselective shadehouses. Hortscience 2013, 48, 975–979. [Google Scholar] [CrossRef]
- Ilić, Z.S.; Fallik, E. Light quality manipulation improves vegetable quality at harvest and postharvest: A review. Environ. Exp. Bot. 2017, 139, 79–90. [Google Scholar] [CrossRef]
- Oren-Shamir, M.; Gussakovsky, E.E.; Shpiegel, E.; Nissim-Levi, A.; Ratner, K.; Ovadia, R.; Giller, Y.E.; Shahak, Y. Coloured shade nets can improve the yield and quality of green decorative branches of Pittosporum variegatum. J. Hortic. Sci. Biotechnol. 2001, 76, 353–361. [Google Scholar] [CrossRef]
- Shahak, Y.; Gussakovsky, E.E.; Cohen, Y.; Lurie, S.; Stern, R.; Kfir, S.; Naor, A.; Atzmon, I.; Doron, I.; Greenblat-Avron, Y. ColorNets: A new approach for light manipulation in fruit trees. Acta Hortic. 2004, 636, 609–616. [Google Scholar] [CrossRef]
- Shahak, Y. Photo-selective netting for improved performance of horticultural crops. A review of ornamental and vegetable studies carried out in Israel. Acta Hortic. 2008, 770, 161–168. [Google Scholar] [CrossRef]
- Shahak, Y.; Kong, Y.; Ratner, K. The wonders of yellow netting. Acta Hortic. 2016, 1134, 327–334. [Google Scholar] [CrossRef]
- Bastias, R.M. Morphological and Physiological Responses of Apple Trees under Photoselective Colored Nets. Ph.D. Thesis, Alma Mater Studiorum—Università di Bologna, Bologna, Italy, 2011. [Google Scholar]
- Briassoulis, D.Ã.; Mistriotis, A.; Eleftherakis, D. Mechanical behaviour and properties of agricultural nets—Part I: Testing methods for agricultural nets. Polym. Test. 2007, 26, 822–832. [Google Scholar] [CrossRef]
- Basile, B.; Giaccone, M.; Cirillo, C.; Ritieni, A.; Graziani, G.; Shahak, Y.; Forlani, M. Photo-selective hail nets affect fruit size and quality in Hayward kiwifruit. Sci. Hortic. 2012, 141, 91–97. [Google Scholar] [CrossRef]
- Shahak, Y.; Ratner, K.; Giller, Y.E.; Zur, N.; Or, E.; Gussakovsky, E.E.; Stern, R.; Sarig, P.; Raban, E.; Harcavi, E.; et al. Improving solar energy utilization, productivity and fruit quality in orchards and vineyards by photoselective netting. Acta Hortic. 2008, 772, 65–72. [Google Scholar] [CrossRef]
- Šavikin, K.; Mikulič-Petkovšek, M.; Djordjević, B.; Zdunić, G.; Janković, T.; Djurović, D.; Veberič, R. Influence of shading net on polyphenol profile and radical scavenging activity in different varieties of black currant berries. Sci. Hortic. 2013, 160, 20–28. [Google Scholar] [CrossRef]
- McCaskill, M.R.; McClymont, L.; Goodwin, I.; Green, S.; Partington, D.L. How hail netting reduces apple fruit surface temperature: A microclimate and modelling study. Agric. For. Meteorol. 2016, 226–227, 148–160. [Google Scholar] [CrossRef]
- Zoratti, L.; Jaakola, L.; Häggman, H.; Giongo, L. Modification of sunlight radiation through colored photo-selective nets affects anthocyanin profile in Vaccinium spp. berries. PLoS ONE 2015, 10, e0135935. [Google Scholar] [CrossRef] [PubMed]
- Ordóñez, V.; Molina-Corral, F.J.; Olivas-Dorantes, C.L.; Jacobo-Cuéllar, J.L.; González-Aguilar, G.; Espino, M.; Sepulveda, D.; Olivas, G.I. Comparative study of the effects of black or white hail nets on the fruit quality of ‘Golden Delicious’ apples. Fruits 2016, 71, 229–238. [Google Scholar] [CrossRef]
- Amarante, C.V.T.; Steffens, C.A.; Mota, C.S.; Santos, H.P. Radiation, photosynthesis, yield, and fruit quality of ‘Royal Gala’ apples under hail protection nets. Pesqui. Agropecu. Bras. 2007, 42, 925–931. [Google Scholar] [CrossRef]
- Amarante, C.V.T.; Steffens, C.A.; Zanardi, O.Z.; Miqueloto, A.; Santos, H.P. Light supply to ‘Fuji’ apple trees covered with hail protection nets and its effects on photosynthesis, yield and fruit quality. Rev. Bras. Frutic. 2009, 31, 664–670. [Google Scholar] [CrossRef]
- Brglez Sever, M.; Tojnko, S.; Breznikar, A.; Skendrović Babojelić, M.; Ivančič, A.; Sirk, M.; Unuk, T. The influence of differently coloured anti-hail nets and geomorphologic characteristics on microclimatic and light conditions in apple orchards. J. Cent. Eur. Agric. 2020, 21, 386–397. [Google Scholar] [CrossRef]
- Ilić, Z.S.; Milenković, L.; Stanojević, L.; Cvetković, D.; Fallik, E. Effects of the modification of light intensity by color shade nets on yield and quality of tomato fruits. Sci. Hortic. 2012, 139, 90–95. [Google Scholar] [CrossRef]
- Retamales, J.B.; Montecino, J.M.; Lobos, G.A.; Rojas, L.A. Colored shading nets increase yields and profitability of highbush blueberries. Acta Hortic. 2008, 770, 193–197. [Google Scholar] [CrossRef]
- Solomakhin, A.; Blanke, M.M. Can coloured hailnets improve taste (sugar, sugar: Acid ratio), consumer appeal (colouration) and nutritional value (anthocyanin, vitamin C) of apple fruit? LWT-Food Sci. Technol. 2010, 43, 1277–1284. [Google Scholar] [CrossRef]
- Blanke, M.M. The structure of coloured hail nets affects light transmission, light spectrum, phytochrome and apple fruit colouration. Acta Hortic. 2009, 817, 177–184. [Google Scholar] [CrossRef]
- Agrintech. Agrintech Technical Sheet. Available online: https://www.agrintech.it/en/products/iridium (accessed on 19 May 2022).
- Healey, K.D.D.; Rickert, K.G.G.; Hammer, G.L.L.; Bange, M.P.; Bange, M.P. Radiation use efficiency increases when the diffuse component of incident radiation is enhanced under shade. Aust. J. Agric. Res. 1998, 49, 665–672. [Google Scholar] [CrossRef]
- Blanke, M. Coloured hailnets: Their structure, light and UV transmission determine the colouration of apple fruit. Erwerbs-Obstbau 2007, 49, 127–140. [Google Scholar] [CrossRef]
- Bastías, R.M.; Ruíz, K.; Manfrini, L.; Pierpaoli, E.; Zibordi, M.; Morandi, B.; Losciale, P.; Torrigiani, P.; Corelli-Grappadelli, L. Effects of photoselective nets on phenolic composition in apple fruits. Acta Hortic. 2012, 939, 77–83. [Google Scholar] [CrossRef]
- Demotes-Mainard, S.; Péron, T.; Corot, A.; Bertheloot, J.; Le Gourrierec, J.; Travier, S.; Crespel, L.; Morel, P.; Huché-Thélier, L.; Boumaza, R.; et al. Plant responses to red and far-red lights, applications in horticulture. Environ. Exp. Bot. 2016, 121, 4–21. [Google Scholar] [CrossRef]
- Mazhawu, E. The Effect of Shadenetting on ’3-29-5’Avocado Production under Subtropical Conditions. Master’s Thesis, University of KwaZulu-Natal, Durban, South Africa, 2016. [Google Scholar]
- Kalcsits, L.; Musacchi, S.; Layne, D.R.; Schmidt, T.; Mupambi, G.; Serra, S.; Mendoza, M.; Asteggiano, L.; Jarolmasjed, S.; Sankaran, S.; et al. Above and below-ground environmental changes associated with the use of photoselective protective netting to reduce sunburn in apple. Agric. For. Meteorol. 2017, 237–238, 9–17. [Google Scholar] [CrossRef]
- Basile, B.; Giaccone, M.; Shahak, Y.; Forlani, M.; Cirillo, C. Regulation of the vegetative growth of kiwifruit vines by photo-selective anti-hail netting. Sci. Hortic. 2014, 172, 300–307. [Google Scholar] [CrossRef]
- Iglesias, I.; Alegre, S. The effect of anti-hail nets on fruit protection, radiation, temperature, quality and profitability of ‘Mondial Gala’ apples. J. Appl. Hortic. 2006, 8, 91–100. [Google Scholar] [CrossRef]
- García-Sánchez, F.; Simón, I.; Lidón, V.; Manera, F.J.; Simón-Grao, S.; Pérez-Pérez, J.G.; Gimeno, V. Shade screen increases the vegetative growth but not the production in “Fino 49” lemon trees grafted on Citrus macrophylla and Citrus aurantium L. Sci. Hortic. 2015, 194, 175–180. [Google Scholar] [CrossRef]
- Kalcsits, L.; Asteggiano, L.; Schmidt, T.; Musacchi, S.; Serra, S.; Layne, D.R.; Mupambi, G. Shade netting reduces sunburn damage and soil moisture depletion in ‘Granny Smith’ apples. Acta Hortic. 2018, 1228, 85–90. [Google Scholar] [CrossRef]
- Corvalán, N.; Bastías, R.M.; Umanzor, C.; Serra, I. Grapevine root and shoot growth responses to photoselective nets: Preliminary results. Acta Hortic. 2016, 1136, 89–94. [Google Scholar] [CrossRef]
- Stamps, R.H. Use of colored shade netting in horticulture. Hortscience 2009, 44, 239–241. [Google Scholar] [CrossRef]
- Stamps, R.H. Evapotranspiration and Nitrogen Leaching during Leatherleaf Fern Production in Shadehouses; Special Publication; St. Johns River Water Management District: Palatka, FL, USA, 1994. [Google Scholar]
- Rosenberg, N.J.; Blad, B.L.; Verma, S.B. Microclimate: The Biological Environment, 2nd ed.; John Wiley & Sons: New York, NY, USA, 1983. [Google Scholar]
- Elad, Y.; Messika, Y.; Brand, M.; David, D.R.; Sztejnberg, A. Effect of colored shade nets on pepper powdery mildew (Leveillula taurica). Phytoparasitica 2007, 35, 285–299. [Google Scholar] [CrossRef]
- Bosco, L.C.; Bergamaschi, H.; Cardoso, L.S.; de Paula, V.A.; Marodin, G.A.B.; Brauner, P.C. Microclimate alterations caused by agricultural hail net coverage and effects on apple tree yield in subtropical climate of southern Brazil. Bragantia 2018, 77, 181–192. [Google Scholar] [CrossRef]
- Hunsche, M.; Blanke, M.M.; Noga, G. Does the microclimate under hail nets influence micromorphological characteristics of apple leaves and cuticles? J. Plant Physiol. 2010, 167, 974–980. [Google Scholar] [CrossRef] [PubMed]
- Wachsmann, Y.; Zur, N.; Shahak, Y.; Ratner, K.; Giler, Y.; Schlizerman, L.; Sadka, A.; Cohen, S.; Garbinshikof, V.; Giladi, B.; et al. Photoselective anti-hail netting for improved citrus productivity and quality. Acta Hortic. 2014, 1015, 169–176. [Google Scholar] [CrossRef]
- Milivojević, J.; Radivojević, D.; Ruml, M.; Dimitrijević, M.; Maksimović, J.D. Does microclimate under grey hail protection net affect biological and nutritional properties of “Duke” highbush blueberry (Vaccinium corymbosum L.)? Fruits 2016, 71, 161–170. [Google Scholar] [CrossRef]
- Gimeno, V.; Simón, I.; Martínez, V.; Lidón, V.; Shahid, M.A.; Garcia-Sanchez, F. Effect of shade screen on production, fruit quality and growth parameters of “Fino 49” lemon trees grafted on Citrus macrophylla and sour orange. Acta Hortic. 2015, 1065, 1845–1852. [Google Scholar] [CrossRef]
- Melgarejo, P.; Legua, P.; Martínez-Font, R.; Martínez-Nicolás, J.J.; Soriano, J.S.; Carbonell-Barrachina, Á.A.; Hernández, F. Response of apricot fruit quality to protective netting. Agriculture 2021, 11, 260. [Google Scholar] [CrossRef]
- Lee, T.-C.; Zhong, P.-J.; Chang, P.-T. The effects of preharvest shading and postharvest storage temperatures on the quality of ‘Ponkan’ (Citrus reticulata Blanco) mandarin fruits. Sci. Hortic. 2015, 188, 57–65. [Google Scholar] [CrossRef]
- Dussi, M.C.; Giardina, G.; Sosa, D.; Gonz´alez Junyent, R.; Zecca, A.; Reeb, P. Shade nets effect on canopy light distribution and quality of fruit and spur leaf on apple cv. ‘Fuji.’ Span. J. Agric. Res. 2005, 3, 253–260. [Google Scholar] [CrossRef]
- Amarante, C.V.T.; Steffens, C.A.; Argenta, L.C. Yield and fruit quality of ‘Gala’ and ‘Fuji’ apple trees protected by white anti-hail net. Sci. Hortic. 2011, 129, 79–85. [Google Scholar] [CrossRef]
- Jemrić, T.; Brkljača, M.; Vinceković, M.; Antolković, A.M.; Mikec, D.; Vuković, M. Generative and vegetative traits of the ‘Granny smith’ apple grown under an anti-insect photoselective red net. Poljoprivreda 2021, 27, 34–42. [Google Scholar] [CrossRef]
- Aoun, M.; Manja, K. Effects of a photoselective netting system on Fuji and Jonagold apples in a Mediterranean orchard. Sci. Hortic. 2020, 263, 109104. [Google Scholar] [CrossRef]
- Vuković, M.; Brkljača, M.; Rumora, J.; Fruk, M.; Jatoi, M.A.; Jemrić, T. Vegetative and reproductive traits of young peaches and nectarines grown under red photoselective net. Agric. Conspec. Sci. 2016, 81, 181–185. [Google Scholar]
- Basile, B.; Romano, R.; Giaccone, M.; Barlotti, E.; Colonna, V.; Cirillo, C.; Shahak, Y.; Forlani, M. Use of photo-selective nets for hail protection of kiwifruit vines in southern Italy. Acta Hortic. 2008, 770, 185–192. [Google Scholar] [CrossRef]
- Brar, H.S.; Thakur, A.; Singh, H.; Kaur, N. Photoselective coverings influence plant growth, root development, and buddability of citrus plants in protected nursery. Acta Physiol. Plant. 2020, 42, 18. [Google Scholar] [CrossRef]
- Franklin, K.A. Shade avoidance. New Phytol. 2008, 179, 930–944. [Google Scholar] [CrossRef]
- Smith, H.; Whitelam, G.C. The shade avoidance syndrome: Multiple responses mediated by multiple phytochromes. Plant Cell Environ. 1997, 20, 840–844. [Google Scholar] [CrossRef]
- Casal, J.J. Shade Avoidance. Arab. Book 2012, 2012, e0157. [Google Scholar] [CrossRef]
- Palonen, P.; Karhu, S.; Savelainen, H.; Rantanen, M.; Junttila, O. Growth and cropping of primocane and biennial raspberry cultivars grown under a film absorbing far-red light. J. Hortic. Sci. Biotechnol. 2011, 86, 113–119. [Google Scholar] [CrossRef]
- Mpezamihigo, M. The Use of Photoselective Plastic Films to Control Growth and Cropping of Three Raspberry (Rubus idaeus) Cultivars, ‘Autumn Bliss’, ‘Glen Ample’, and ‘Joan Squire’. Ph.D Thesis, University of Reading, Reading, UK, 2004. [Google Scholar]
- Morgan, D.C.; Stanley, C.J.; Warrington, I.J. The effects of simulated daylight and shade-light on vegetative and reproductive growth in kiwifruit and grapevine. J. Hortic. Sci. 1985, 60, 473–484. [Google Scholar] [CrossRef]
- Pevalek-Kozlina, B. Fiziologija Bilja; Profil International: Zagreb, Croatia, 2003; ISBN 953-200-775-x. [Google Scholar]
- Bastías, R.M.; Corelli-Grappadelli, L. Light quality management in fruit orchards: Physiological and technological aspects. Chil. J. Agric. Res. 2012, 72, 574–581. [Google Scholar] [CrossRef]
- Sager, J.; Smith, W.; Edwards, J.; Cyr, K.L. Photosynthetic efficiency and phytochrome photoequilibria determination using spectral data. Trans. ASAE 1988, 31, 1882–1889. [Google Scholar] [CrossRef]
- Rajapakse, N.C.; Kelly, J.W. Problems of reporting spectral quality and interpreting phytochrome-mediated responses. HortScience 1994, 29, 1404–1407. [Google Scholar] [CrossRef]
- Kasperbahuer, M.J. Phytochrome in crop production. In Plant-Environment Interactions; Wilkinson, R.E., Ed.; Marcel Dekker: New York, NY, USA, 2000; pp. 407–434. [Google Scholar]
- Smith, H. Phytochromes and light signal perception by plants—An emerging synthesis. Nature 2000, 407, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Holmes, M.G.; Smith, H. Function of phytochrome in natural environment. 1. Characterization of daylight for studies in photomorphogenesis and photoperiodism. Photochem. Photobiol. 1977, 25, 533–538. [Google Scholar] [CrossRef]
- Baraldi, R.; Rossi, F.; Facini, O.; Fasolo, F.; Rotondi, A.; Magli, M.; Nerozzi, F. Light environment, growth and morphogenesis in a peach tree canopy. Physiol. Plant. 1994, 91, 339–345. [Google Scholar] [CrossRef]
- Gregoriou, K.; Pontikis, K.; Vemmos, S. Effects of reduced irradiance on leaf morphology, photosynthetic capacity, and fruit yield in olive (Olea europaea L.). Photosynthetica 2007, 45, 172–181. [Google Scholar] [CrossRef]
- Ajmi, A.; Vázquez, S.; Morales, F.; Chaari, A.; El-jendoubi, H.; Abadía, A.; Larbi, A. Prolonged artificial shade affects morphological, anatomical, biochemical and ecophysiological behavior of young olive trees (cv. Arbosana). Sci. Hortic. 2018, 241, 275–284. [Google Scholar] [CrossRef]
- Combes, D.; Sinoquet, H.; Varlet-Grancher, C. Preliminary measurement and simulation of the spatial distribution of the morphogenetically active radiation (MAR) within an isolated tree canopy. Ann. For. Sci. 2000, 57, 497–511. [Google Scholar] [CrossRef]
- Butorac, A. Opća Agronomija; Školska Knjiga: Zagreb, Croatia, 1999; ISBN 953-0-31118-4. [Google Scholar]
- Rapparini, F.; Rotondi, A.; Baraldi, R. Blue light regulation of the growth of Prunus persica plants in a long term experiment: Morphological and histological observations. Trees-Struct. Funct. 1999, 14, 169–176. [Google Scholar] [CrossRef]
- Baraldi, R.; Rapparini, F.; Rotondi, A.; Bertazza, G. Effects of simulated light environments on growth and leaf morphology of peach plants. J. Hortic. Sci. Biotechnol. 1998, 73, 251–258. [Google Scholar] [CrossRef]
- Nanya, K.; Ishigami, Y.; Hikosaka, S.; Goto, E. Effects of blue and red light on stem elongation and flowering of tomato seedlings. Acta Hortic. 2012, 956, 261–266. [Google Scholar] [CrossRef]
- Christophe, A.; Moulia, B.; Varlet-Grancher, C. Quantitative contributions of blue light and PAR to the photocontrol of plant morphogenesis in Trifolium repens (L.). J. Exp. Bot. 2006, 57, 2379–2390. [Google Scholar] [CrossRef]
- Kozlowski, T.T.; Keller, T. Food relations of woody plants. Bot. Rev. 1966, 32, 293–382. [Google Scholar] [CrossRef]
- Fan, X.X.; Xu, Z.G.; Liu, X.Y.; Tang, C.M.; Wang, L.W.; Han, X.L. Effects of light intensity on the growth and leaf development of young tomato plants grown under a combination of red and blue light. Sci. Hortic. 2013, 153, 50–55. [Google Scholar] [CrossRef]
- Smith, H.M.; Samach, A. Constraints to obtaining consistent annual yields in perennial tree crops. I: Heavy fruit load dominates over vegetative growth. Plant Sci. 2013, 207, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Barradas, V.L.; Nicolás, E.; Torrecillas, A.; Alarcón, J.J. Transpiration and canopy conductance in young apricot (Prunus armenica L.) trees subjected to different PAR levels and water stress. Agric. Water Manag. 2005, 77, 323–333. [Google Scholar] [CrossRef]
- Stampar, F.; Veberic, R.; Zadravec, P.; Hudina, M.; Usenik, V.; Solar, A.; Osterc, G. Yield and fruit quality of apples cv. “Jonagold” under hail protection nets. Gartenbauwissenschaft 2002, 67, 205–210. [Google Scholar]
- Gindaba, J.; Wand, S.J.E. Sunburn in apples and effectiveness of control measures. In Proceedings of the Combined Congress, Potchefstroom, South Africa, 10–13 January 2005. [Google Scholar]
- Gindaba, J.; Wand, S.J.E. Comparative effects of evaporative cooling, kaolin particle film, and shade net on sunburn and fruit quality in apples. HortScience 2005, 40, 592–596. [Google Scholar] [CrossRef]
- Terashima, I.; Fujita, T.; Inoue, T.; Chow, W.S.; Oguchi, R. Green light drives leaf photosynthesis more efficiently than red light in strong white light: Revisiting the enigmatic question of why leaves are green. Plant Cell Physiol. 2009, 50, 684–697. [Google Scholar] [CrossRef] [PubMed]
- Hogewoning, S.W.; Trouwborst, G.; Maljaars, H.; Poorter, H.; van Ieperen, W.; Harbinson, J. Blue light dose-responses of leaf photosynthesis, morphology, and chemical composition of Cucumis sativus grown under different combinations of red and blue light. J. Exp. Bot. 2010, 61, 3107–3117. [Google Scholar] [CrossRef] [PubMed]
- Snelgar, W.P.; Manson, P.J.; Hopkirk, G. Effect of overhead shading on fruit size and yield potential of kiwifruit (Actinidia deliciosa). J. Hort. Sci. 1991, 66, 261–273. [Google Scholar] [CrossRef]
- Leite, G.B.; Petri, J.L.; Mondardo, M. Effects of net shield against hailstorm on feature of apples production and fruit quality. Rev. Bras. Frutic. 2002, 24, 714–716. [Google Scholar] [CrossRef]
- Westwood, M.N. Temperate-Zone Pomology: Physiology and Culture, 3rd ed.; Timber Press: Portland, OR, USA, 1993; ISBN 0881922536, 9780881922530. [Google Scholar]
- Koutinas, N.; Pepelyankov, G.; Lichev, V. Flower induction and flower bud development in apple and sweet cherry. Biotechnol. Biotechnol. Equip. 2010, 24, 1549–1558. [Google Scholar] [CrossRef]
- Yanez, P.; Retamales, J.B.; Lobos, G.A.; del Pozo, A. Light environment within mature rabbiteye blueberry canopies influences flower bud formation. Acta Hortic. 2009, 810, 417–473. [Google Scholar]
- Williams, M.W. Use of bioregulators to control vegetative growth of fruit trees and improve fruiting efficiency. Acta Hortic. 1984, 146, 97–104. [Google Scholar] [CrossRef]
- Rossi, R.; Facini, O.; Holmes, M.G. Light quality effects on bud differentiation in apple. In Proceedings of the XXIII International Horticultural Congress, Firenze, Italy, 27 August–1 September 1990. [Google Scholar]
- Yanagi, T.; Yachi, T.; Okuda, N.; Okamoto, K. Light quality of continuous illuminating at night to induce floral initiation of Fragaria chiloensis L. CHI-24-1. Sci. Hortic. 2006, 109, 309–314. [Google Scholar] [CrossRef]
- Tromp, J. Flower-bud formation and shoot growth in apple as affected by temperature. Sci. Hortic. 1976, 5, 331–338. [Google Scholar] [CrossRef]
- Ramírez, F.; Davenport, T.L. Mango (Mangifera indica L.) flowering physiology. Sci. Hortic. 2010, 126, 65–72. [Google Scholar] [CrossRef]
- Abbott, D.L.; Bull, V.; Bishop, S.N. Effect of Summer Temperature on Flower Initiation; Report of the Long Ashton Research Station for 1973; Long Ashton Research Station: Long Ashton, UK, 1974. [Google Scholar]
- Chen, K.; Hu, G.Q.; Lenz, F. Training and shading effects on vegetative and reproductive growth and fruit quality of apple. Gartenbauwiss 1997, 62, 207–213. [Google Scholar]
- Wünsche, J.N.; Palmer, J.W.; Greer, D.H. Effects of crop load on fruiting and gas-exchange characteristic of ’Braeburn’/M.26 apple trees at full canopy. J. Am. Soc. Hortic. Sci. 2000, 125, 93–99. [Google Scholar] [CrossRef]
- Wünsche, J.N.; Ferguson, I.B. Crop Load Interactions in Apple. Hortic. Rev. (Am. Soc. Hortic. Sci). 2005, 31, 233–245. [Google Scholar]
- Smit, A. Apple Tree and Fruit Responses to Shade Netting. Master’s Thesis, University of Stellenbosch, Stellenbosch, South Africa, 2007. [Google Scholar]
- Lloyd, A.; Hamacek, E.; George, A.; Nissen, R.; Waite, G. Evaluation of exclusion netting for insect pest control and fruit quality enhancement in tree crops. Acta Hortic. 2005, 694, 253–258. [Google Scholar] [CrossRef]
- Byers, R.E.; Barden, J.A.; Carbaugh, D.H. Thinning of spur `Delicious’ apples by shade, terbacil, carbaryl, and ethephon. J. Am. Soc. Hortic. Sci. 1990, 115, 9–13. [Google Scholar] [CrossRef]
- Grauslund, J.; Hansen, P. Fruit Trees and Climate. III. The Effect of Temperature on Fruit Set in Apple Trees. State Research Organisation for Plant Culture, Report No. 1235. 1975. Available online: https://www.google.com/url?sa=t&rct=j&q=&esrc=s&source=web&cd=&cad=rja&uact=8&ved=2ahUKEwj2-KiK9Jj5AhUNgf0HHSg8APsQFnoECAkQAQ&url=https%3A%2F%2Fdcapub.au.dk%2Fpub%2Fplanteavl_79_481.pdf&usg=AOvVaw0FC-atp1wxkCKPtVD5bBLv (accessed on 19 May 2022).
- Kondo, S.; Takahashi, Y. Effects of high temperature in the nighttime and shading in the daytime on the early drop of apple fruit ‘Starking Delicious’. J. Jpn. Soc. Hort. Sci. 1987, 56, 142–150. [Google Scholar] [CrossRef]
- Lakso, A.N. Early fruit growth and drop—The role of carbon balance in the apple tree. Acta Hortic. 2011, 903, 733–742. [Google Scholar] [CrossRef]
- Anila, R.; Radha, T. Studies on Fruit Drop in Mango Varieties. J. Trop. Agric. 2003, 41, 30–32. [Google Scholar]
- Vuković, M.; Buhin, J.; Brkljača, M.; Jatoi, M.A.; Jemrić, T. Postharvest quality of ´Granny Smith´ apple grown under photo-selective red net. J. Cent. Eur. Agric. 2020, 21, 124–128. [Google Scholar] [CrossRef]
- Iglesias, I.; Echeverría, G. Differential effect of cultivar and harvest date on nectarine colour, quality and consumer acceptance. Sci. Hortic. 2009, 120, 41–50. [Google Scholar] [CrossRef]
- Corollaro, M.L.; Manfrini, L.; Endrizzi, I.; Aprea, E.; Demattè, M.L.; Charles, M.; Bergamaschi, M.; Biasioli, F.; Zibordi, M.; Corelli Grappadelli, L.; et al. The effect of two orchard light management practices on the sensory quality of apple: Fruit thinning by shading or photo-selective nets. J. Hortic. Sci. Biotechnol. 2015, 90, 99–107. [Google Scholar] [CrossRef]
- Jackson, J.E.; Palmer, J.W.; Perring, M.A.; Sharples, R.O. Effects of shade III, the growth and cropping of apple trees. Effects on fruit growth, chemical composition and quality at harvest and after storage. J. Hort. Sci. 1977, 52, 267–282. [Google Scholar] [CrossRef]
- Corelli-Grappadelli, L.; Lakso, A.N.; Flore, J.A. Early season pattern of carbohydrate partitioning in exposed and shaded apple branches. J. Am. Soc. Hortic. Sci. 1994, 119, 596–603. [Google Scholar] [CrossRef]
- Brglez Sever, M. Influence of Different Colours of Anti-Hailnets on the Natural Thinning, Microclimate Andlight Conditions in an Orchard and Qualityparameters of Apple Fruits (Malus domestica Borkh.). Ph.D. Thesis, University of Maribor—Faculty of Agriculture and Life Sciences, Maribor, Slovenia, 2021. [Google Scholar]
- Wünsche, J.N.; Greer, D.H.; Laing, W.A.; Palmer, J.W. Physiological and biochemical leaf and tree responses to crop load in apple. Tree Physiol. 2005, 25, 1253–1263. [Google Scholar] [CrossRef] [PubMed]
- Little, A.C. A Research note: Off on a Tangent. J. Food Sci. 1975, 40, 410–411. [Google Scholar] [CrossRef]
- Jakopic, J.; Veberic, R.; Stampar, F. The effect of reflective foil and hail nets on the lighting, color and anthocyanins of ‘Fuji’ apple. Sci. Hortic. 2007, 115, 40–46. [Google Scholar] [CrossRef]
- Vuković, M.; Jurić, S.; Vinceković, M.; Levaj, B.; Fruk, G.; Jemrić, T. Effect of yellow and Stop Drosophila Normal anti-insect photoselective nets on vegetative, generative and bioactive traits of peach (cv. Suncrest). J. Agric. Sci.-Tarım Bilim. Derg. 2022, in press. [Google Scholar] [CrossRef]
- Jaakola, L. New insights into the regulation of anthocyanin biosynthesis in fruits. Trends Plant Sci. 2013, 18, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Garcia, S.N.; Guevara-Gonzalez, R.G.; Miranda-Lopez, R.; Feregrino-Perez, A.A.; Torres-Pacheco, I.; Vazquez-Cruz, M.A. Functional properties and quality characteristics of bioactive compounds in berries: Biochemistry, biotechnology, and genomics. Food Res. Int. 2013, 54, 1195–1207. [Google Scholar] [CrossRef]
- Awad, M.A.; Wagenmakers, P.S.; Jager, A.D. Effects of light on flavonoid and chlorogenic acid levels in the skin of ‘Jonagold’ apples. Sci. Hortic. 2001, 88, 289–298. [Google Scholar] [CrossRef]
- González-Talice, J.; Yuri, J.A.; del Pozo, A. Relations among pigments, color and phenolic concentrations in the peel of two Gala apple strains according to canopy position and light environment. Sci. Hortic. 2013, 151, 83–89. [Google Scholar] [CrossRef]
- Hamadziripi, E.T. The Effect of Canopy Position on the Fruit Quality and Consumer Preference of Apples. Master’s Thesis, Stellenbosch University, Stellenbosch, South Africa, 2012. [Google Scholar]
- Wagenmakers, P.S.; Callesen, O. Light distribution in apple orchard systems in relation to production and fruit quality. J. Hortic. Sci. 1995, 70, 935–948. [Google Scholar] [CrossRef]
- Zoratti, L.; Karppinen, K.; Escobar, A.L.; Häggman, H.; Jaakola, L. Light-controlled flavonoid biosynthesis in fruits. Front. Plant Sci. 2014, 5, 534. [Google Scholar] [CrossRef]
- Anttonen, M.J.; Hoppula, K.I.; Nestby, R.; Verheul, M.J.; Karjalainen, R.O. Influence of fertilization, mulch color, early forcing, fruit order, planting date, shading, growing environment, and genotype on the contents of selected phenolics in strawberry (Fragaria × ananassa Duch.) fruits. J. Agric. Food Chem. 2006, 54, 2614–2620. [Google Scholar] [CrossRef]
- Azuma, A.; Yakushiji, H.; Koshita, Y.; Kobayashi, S. Flavonoid biosynthesis-related genesin grape skin are differentially regulated by temperature and light conditions. Planta 2012, 236, 1067–1080. [Google Scholar] [CrossRef] [PubMed]
- Cortell, J.M.; Kennedy, J.A. Effect of shading on accumulation of flavonoidcompounds in (Vitis vinifera L.) pinot noir fruit and extraction in a modelsystem. J. Agric. Food Chem. 2006, 54, 8510–8520. [Google Scholar] [CrossRef] [PubMed]
- Spayd, S.E.; Tarara, J.M.; Mee, D.L.; Ferguson, J.C. Separation of sunlight and temperature effects on the composition of Vitis vinifera cv. Merlot berries. Am. J. Enol. Vitic. 2002, 53, 171–182. [Google Scholar]
- Takos, A.M.; Jaffe, F.W.; Jacobs, S.R.; Bogs, J.; Robinson, S.P.; Walker, A.R. Light induced expression of MYB gene regulates anthocyanin biosynthesis in red apples. Plant Physiol. 2006, 142, 1216–1232. [Google Scholar] [CrossRef]
- He, F.; Mu, L.; Yan, G.L.; Liang, N.N.; Pan, Q.H.; Wang, J.; Reeves, M.J.; Duan, C.Q. Biosynthesis of anthocyanins and their regulation in colored grapes. Molecules 2010, 15, 9057–9091. [Google Scholar] [CrossRef] [PubMed]
- Jaakola, L.; Määttä-Riihinen, K.; Kärenlampi, S.; Hohtola, A. Activation of flavonoid biosynthesis by solar radiation in bilberry (Vaccinium myrtillus L.) leaves. Planta 2004, 218, 721–728. [Google Scholar] [PubMed]
- Zhang, X.; Allan, A.C.; Yi, Q.; Chen, L.; Li, K.; Shu, Q. Differential gene expression analysis of Yunnan red pear, Pyrus pyrifolia, during fruit skin coloration. Plant Mol. Biol. Rep. 2011, 29, 305–314. [Google Scholar] [CrossRef]
- Arakawa, O.; Hori, Y.; Ogata, R. Relative effectiveness and interaction of ultraviolet-B, red and blue light in anthocyanin synthesis of apple fruit. Physiol. Plant. 1985, 64, 323–327. [Google Scholar] [CrossRef]
- Reay, P.F.; Lancaster, J.E. Accumulation of anthocyanins and quercetin glycosides in ‘Gala’ and ‘Royal Gala’ apple fruit skinwith UVB- Visible irradiation: Modifying effects of fruit maturity, fruit side, and temperature. Sci. Hortic. 2001, 90, 57–68. [Google Scholar]
- Kondo, S.; Tomiyama, H.; Rodyoung, A.; Okawa, K.; Ohara, H.; Sugaya, S.; Terahara, N.; Hirai, N. Abscisic acid metabolism and anthocyanin synthesis in grape skin are affected by light emitting diode (LED) irradiation at night. J. Plant Physiol. 2014, 171, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Arakawa, O. Effect of temperature on anthocyanin accumulation in apple fruit as affected by cultivar, stage of fruit ripening and bagging. J. Hortic. Sci. 1991, 66, 763–768. [Google Scholar] [CrossRef]
- Faragher, J.D. Temperature regulation of anthocyanin accumulation in apple skin. J. Expt. Bot. 1983, 34, 1291–1298. [Google Scholar] [CrossRef]
- Arakawa, O.; Shinoda, M.; Hiraga, M.; Wang, H. Comparison of anthocyanin synthesis of true-totype ‘Tsugaru’ apple and its red sport strains. J. Hortic. Sci. 1999, 74, 738–742. [Google Scholar]
- Mori, K.; Goto-Yamamoto, N.; Kitayama, M.; Hashizume, K.; Poremećaji, F. Loss of anthocyanins in red-wine grape under high temperature. J. Exp. Bot. 2007, 58, 1935–1945. [Google Scholar] [PubMed]
- Lin-Wang, K.; Micheletti, D.; Palmer, J.; Volz, R.; Lozano, L.; Espley, R.; Hellens, R.P.; Chagnè, D.; Rowan, D.D.; Troggio, M.; et al. High temperature reduces apple fruit colour via modulation of the anthocyanin regulatory complex. Plant Cell Environ. 2011, 34, 1176–1190. [Google Scholar] [CrossRef] [PubMed]
- Arena, M.E.; Postemsky, P.D.; Curvetto, N.R. Changes in the phenolic compounds and antioxidant capacity of Berberis microphylla G. Forst. berries in relation to light intensity and fertilization. Sci. Hortic. 2017, 218, 63–71. [Google Scholar] [CrossRef]
- DeEll, J.R.; Khanizadeh, S.; Saad, F.; Ferree, D.C. Factors affecting apple fruit firmness-a review. J. Am. Pomol. Soc. 2001, 55, 8–26. [Google Scholar]
- Skendrović Babojelić, M.; Fruk, G. Priručnik iz Voćarstva: Građa, Svojstva i Analize Voćnih Plodova; Hrvatska sveučilišna naklada; Sveučilište u Zagrebu Agronomski Fakultet: Zagreb, Croatia, 2016; ISBN 978-953-169-318-9. [Google Scholar]
- Campbell, R.J.; Marini, R.P. Light environment and time of harvest affect ‘Delicious’ apple fruit quality characteristics. J. Am. Soc. Hortic. Sci. 1992, 117, 551–557. [Google Scholar] [CrossRef]
- Loreti, F.; Morini, S.; Muleo, R.; Masetti, C.; Vitagliano, C. Effect of solar radiation deprival on selected parameters of peach fruits. Adv. Hortic. Sci. 1993, 7, 105–108. [Google Scholar]
- Brkljača, M.; Rumora, J.; Vuković, M.; Jemrić, T. The effect of photoselective nets on fruit quality of apple cv. ´Cripps Pink´. Agric. Conspec. Sci. 2016, 81, 87–90. [Google Scholar]
- Musacchi, S.; Serra, S. Apple fruit quality: Overview on pre-harvest factors. Sci. Hortic. 2018, 234, 409–430. [Google Scholar] [CrossRef]
- Cronje, A. Effect of Canopy Position on Fruit Quality and Consumer Preference for the Appearance and Taste of Pears. Master’s Thesis, Faculty of AgriSciences, Stellenbosch University, Stellenbosch, South Africa, 2014. [Google Scholar]
- Lambers, H.; Chapin, F.S.; Pons, T.L. Plant Physiological Ecology; Springer: New York, NY, USA, 1998; ISBN 978-1-4757-2857-6. [Google Scholar]
- Lewallen, K. Effects of Light Availability and Canopy Position on Peach Fruit Quality. Master’s Thesis, Faculty of the Virginia Polytechnic Institute and State University, Blacksburg, VA, USA, 2000. [Google Scholar]
- Corelli Grappadelli, L.; Marini, R.P. Orchard planting system. In The Peach: Botany, Production and Uses; Layne, D.R., Bassi, D., Eds.; Cabi: London, UK, 2008; pp. 264–288. [Google Scholar]
- Lakso, A.N. Water Relations of Apples. In Apples: Botany, Production and Uses; Ferree, D.C., Warrington, I.J., Eds.; CAB International: Wallingford, UK, 2003; pp. 195–216. [Google Scholar]
- Kader, A.A. Flavor quality of fruits and vegetables. J. Sci. Food Agric. 2008, 88, 1863–1868. [Google Scholar] [CrossRef]
- Kader, A.A.A. Fruit maturity, ripening, and quality relationships. Acta Hortic. 1999, 485, 203–208. [Google Scholar] [CrossRef]
- Quilot-Turion, B.; Causse, M. Natural diversity and genetic control of fruit sensory quality. In Fruit Ripening: Physiology, Signalling and Genomics; Nath, P., Bouzayen, M., Pech, J.C., Mattoo, A.K., Eds.; CABI: Wallingford, UK, 2014; ISBN 978-1845939625. [Google Scholar]
- Ramos, E.; Weinbaum, A.; Shackel, K.A.; Schwankl, L.J.; Mitcham, E.J.; Mitchell, F.G.; Snyder, R.G.; Mayer, G.; McGourty, G. Influence of tree water status and canopy position on fruit size and quality of ‘Bartlett’ pears. Acta Hortic. 1994, 367, 192–200. [Google Scholar] [CrossRef]
- Nilsson, T.; Gustavsson, K.E. Postharvest physiology of ‘Aroma’ apples in relation to position on the tree. Postharvest Biol. Technol. 2007, 43, 36–46. [Google Scholar] [CrossRef]
- Bakhshi, D.; Arakawa, O. Induction of phenolic compounds biosynthesis with light irradiation in the flesh of red and yellow apples. J. Appl. Hortic. 2006, 8, 101–104. [Google Scholar] [CrossRef]
- Gullo, G.; Motisi, A.; Zappia, R.; Dattola, A.; Diamanti, J.; Mezzetti, B. Rootstock and fruit canopy position affect peach [Prunus persica (L.) Batsch] (cv. Rich May) plant productivity and fruit sensorial and nutritional quality. Food Chem. 2014, 153, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.Y.; Zheng, W. Effect of plant temperature on antioxidant capacity in strawberry. J. Agric. Food Chem. 2001, 49, 4977–4982. [Google Scholar] [CrossRef]
- Do Nascimento, N.C.; Menguer, P.K.; Sperotto, R.A.; De Almeida, M.R.; Fett-Neto, A.G. Early changes in gene expression induced by acute UV exposure in leaves of psychotria brachyceras, a bioactive alkaloid accumulating plant. Mol. Biotechnol. 2013, 54, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Frohnmeyer, H.; Staiger, D. Ultraviolet-Bradiation-mediated responses in plants.Balancing damage and protection. Plant Physiol. 2003, 133, 1420–1428. [Google Scholar] [CrossRef] [PubMed]
- Giliberto, L.; Perrotta, G.; Pallara, P.; Weller, J.L.; Fraser, P.D.; Bramley, P.M.; Fiore, A.; Tavazza, M.; Giuliano, G. Manipulation of the blue light photoreceptor cryptochrome 2 in tomato affects vegetative development, flowering time, and fruit antioxidant content. Plant Physiol. 2005, 137, 199–208. [Google Scholar] [CrossRef]
- Kokalj, D.; Hribar, J.; Blaž, C.; Zlatić, E.; Demšar, L.; Sinkovič, L.; Šircelj, H.; Bizjak, G.; Vidrih, R. Influence of yellow light-emitting diodes at 590 nm on storage of apple, tomato and bell pepper fruit. Food Technol. Biotechnol. 2016, 54, 228–235. [Google Scholar]
- Josuttis, M.; Carlen, C.; Crespo, P.; Nestby, R.; Toldam-Andersen, T.B.; Dietrich, H.; Krüger, E. A comparison of bioactive compounds of strawberry fruit from Europe affected by genotype and latitude. J. Berry Res. 2012, 2, 73–95. [Google Scholar] [CrossRef]
- Josuttis, M.; Dietrich, H.; Patz, C.D.; Krüger, E. Effects of air and soil temperatures on the chemical composition of fruit and agronomic performance in strawberry (Fragaria × ananassa Duch.). J. Hortic. Sci. Biotechnol. 2011, 86, 415–421. [Google Scholar] [CrossRef]
- Remberg, S.F.; Sønsteby, A.; Aaby, K.; Heide, O.M. Influence of postflowering temperature on fruit size and chemical composition of glen ample raspberry (Rubus idaeus L.). J. Agric. Food Chem. 2010, 58, 9120–9128. [Google Scholar] [CrossRef] [PubMed]
- Racsko, J.; Schrader, L.E. Sunburn of apple fruit: Historical background, recent advances and future perspectives. CRC. Crit. Rev. Plant Sci. 2012, 31, 455–504. [Google Scholar] [CrossRef]
- Wand, S.J.E.; Theron, K.I.; Ackerman, J.; Marais, S.J.S. Harvest and post-harvest apple fruit quality following applications of kaolin particle film in South African orchards. Sci. Hortic. 2006, 107, 271–276. [Google Scholar] [CrossRef]
- Felicetti, D.A.; Schrader, L.E. Photooxidative sunburn of apples:characterization of a third type of apple sunburn. Int. J. Fruit Sci. 2008, 8, 160–172. [Google Scholar] [CrossRef]
- Schrader, L.E.; Zhang, J.; Sun, J. Environmental stresses that cause sunburn of apple. Acta Hortic. 2003, 618, 397–405. [Google Scholar] [CrossRef]
- Yuri, J.A.; Lepe, V.; Moggia, C.; Bastias, R.; Bertschinger, L. Sunburn on apple. Schweiz. Z. Obs. Weinbau 2004, 8, 7–10. [Google Scholar]
- Makeredza, B.; Schmeisser, M.; Lötze, E.; Steyn, W.J. Water stress increases sunburn in ´Cripps Pink´ apple. HortScience 2013, 48, 444–447. [Google Scholar] [CrossRef]
- Hicklenton, P.; Forney, C.F.; Domytrak, C. Row covers to delay or advance maturity in highbush blueberry. Small Fruits Rev. 2004, 3, 169–181. [Google Scholar] [CrossRef]
- Smart, R.E.; Smith, S.M.; Winchester, R.V. Effects, Light quality and quantity Sauvignon, on fruit ripening for Cabernet. Am. J. Enol. Viticult. 1988, 39, 250–258. [Google Scholar]








| Netting | Source | R:FR Ratio | |
|---|---|---|---|
| Diffuse Light | Total Light | ||
| Black | [6] | ↓ * | m.d. |
| Blue | [6,21] | ↓ | |
| [16] | small ↑ | ||
| [6,15,16,21,25] | m.d. | ||
| Green | [15] | ↓ | |
| Gray | [6,16,21] | ↓ | |
| [6,15,16,21] | m.d. | ||
| Pearl | [6,16] | ↓ (mostly highest) | ↓ (mostly highest) |
| [25] | m.d. | ||
| Red | [6,16,21] | ↓ | |
| [6,15,16,21] | ↓ | ||
| [25] | m.d. | ||
| White | [21] | ↓ | m.d. ** |
| Yellow | [6,18] | ↓ | ↓ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vuković, M.; Jurić, S.; Maslov Bandić, L.; Levaj, B.; Fu, D.-Q.; Jemrić, T. Sustainable Food Production: Innovative Netting Concepts and Their Mode of Action on Fruit Crops. Sustainability 2022, 14, 9264. https://doi.org/10.3390/su14159264
Vuković M, Jurić S, Maslov Bandić L, Levaj B, Fu D-Q, Jemrić T. Sustainable Food Production: Innovative Netting Concepts and Their Mode of Action on Fruit Crops. Sustainability. 2022; 14(15):9264. https://doi.org/10.3390/su14159264
Chicago/Turabian StyleVuković, Marko, Slaven Jurić, Luna Maslov Bandić, Branka Levaj, Da-Qi Fu, and Tomislav Jemrić. 2022. "Sustainable Food Production: Innovative Netting Concepts and Their Mode of Action on Fruit Crops" Sustainability 14, no. 15: 9264. https://doi.org/10.3390/su14159264
APA StyleVuković, M., Jurić, S., Maslov Bandić, L., Levaj, B., Fu, D.-Q., & Jemrić, T. (2022). Sustainable Food Production: Innovative Netting Concepts and Their Mode of Action on Fruit Crops. Sustainability, 14(15), 9264. https://doi.org/10.3390/su14159264