
Multi-Stemmed Habit in Trees Contributes Climate Resilience in Tropical Dry Forest

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
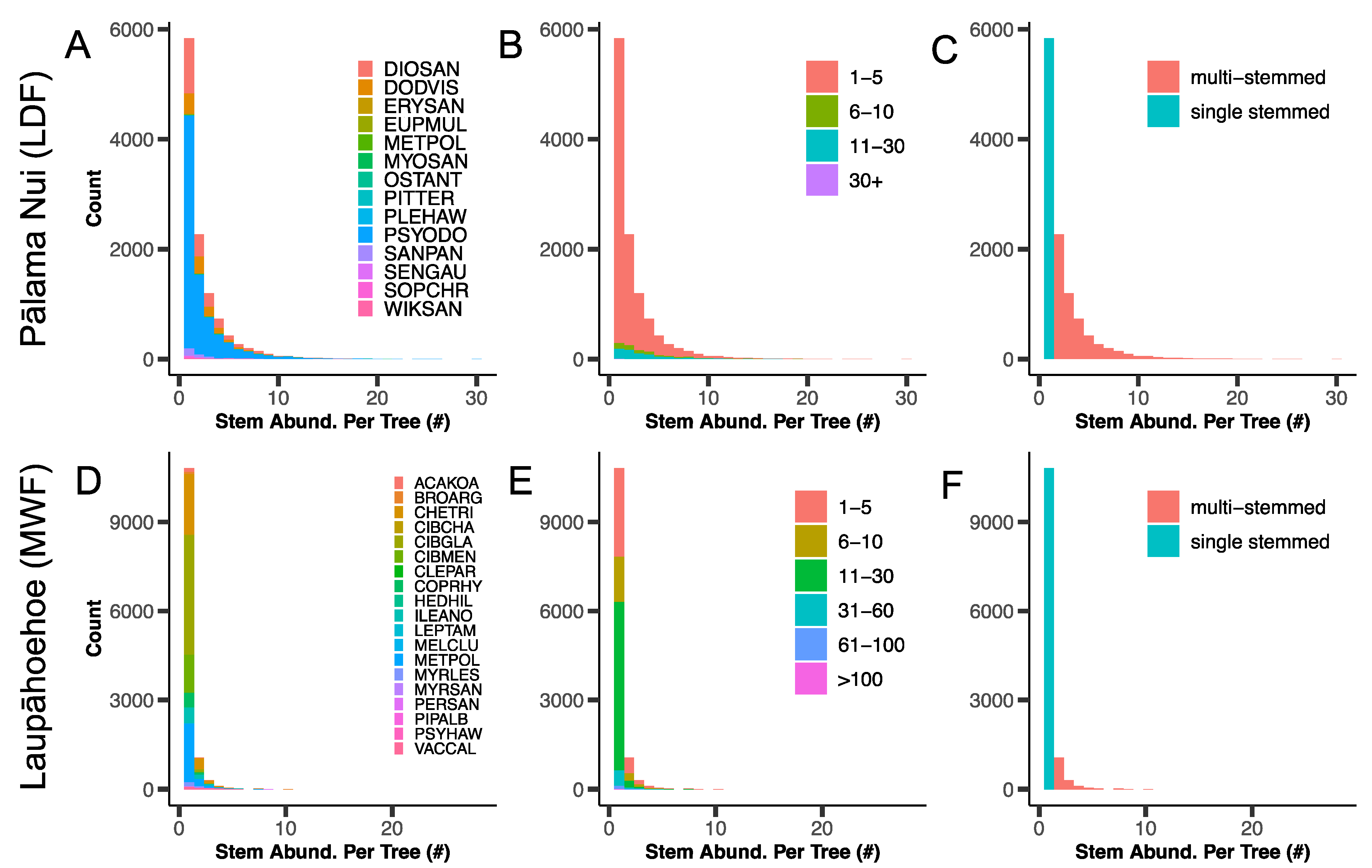
2.1. Forest Dynamics Plot Descriptions
2.2. Structure, Growth, Demography
2.3. Functional Trait Modules
2.4. Statistical Approach
3. Results
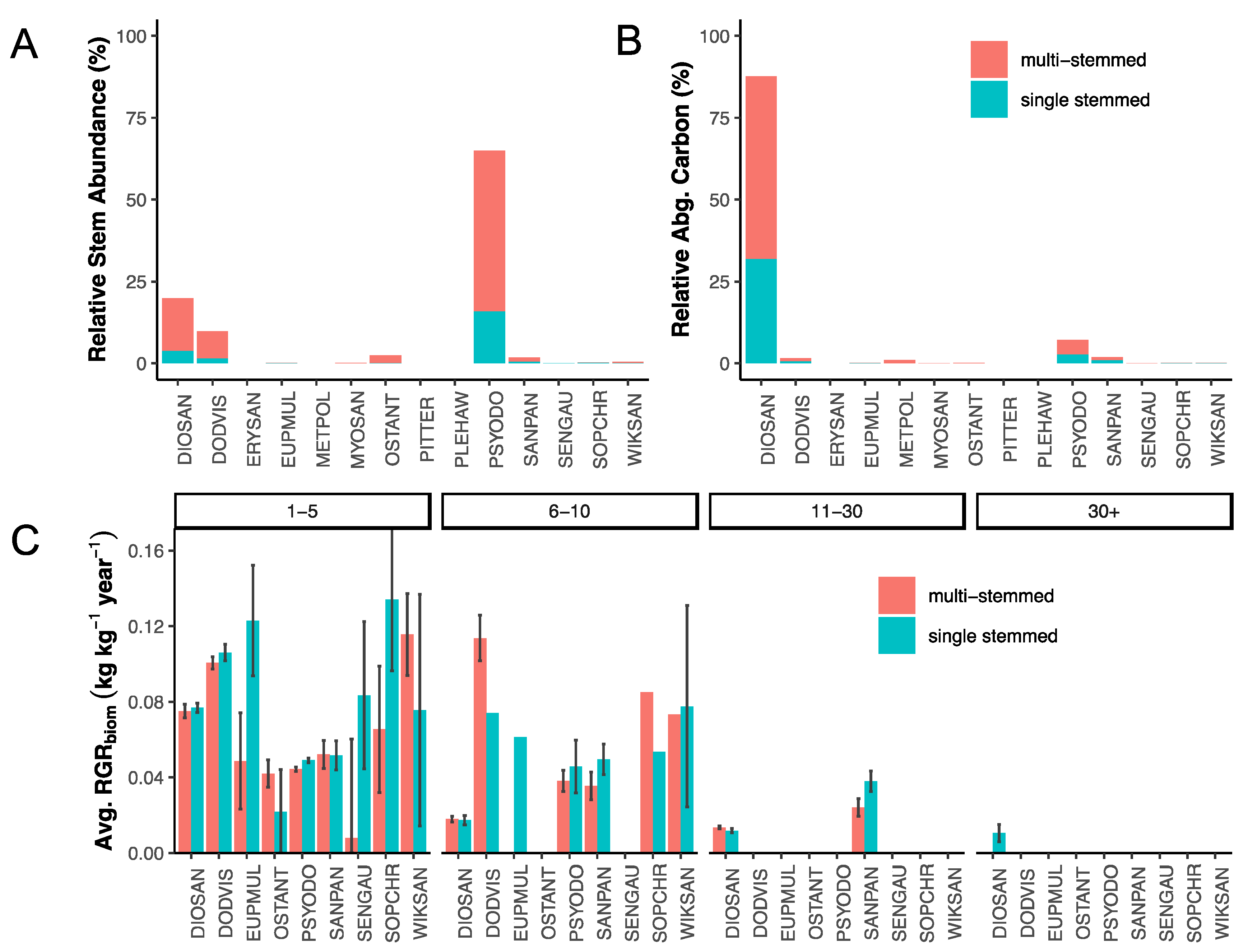
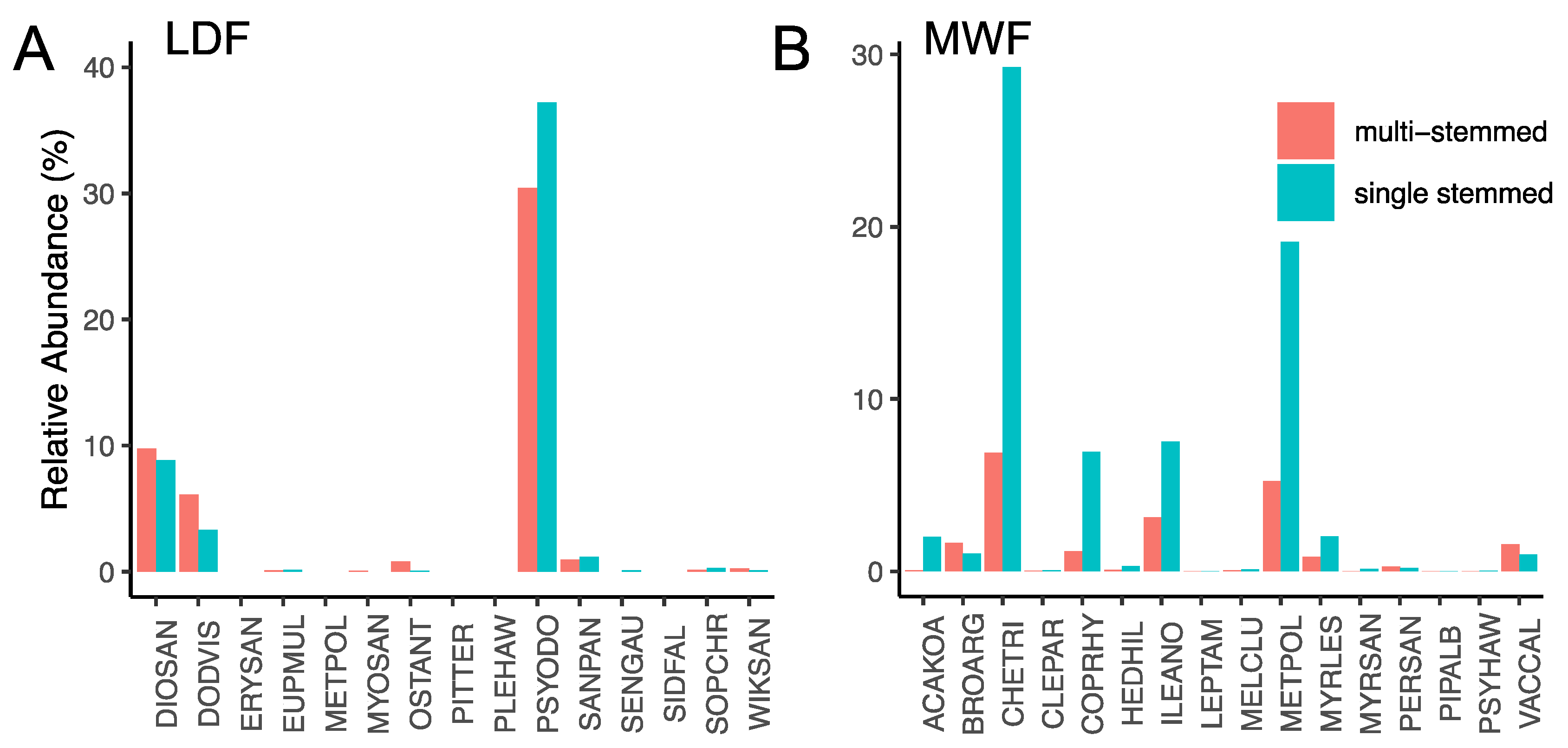
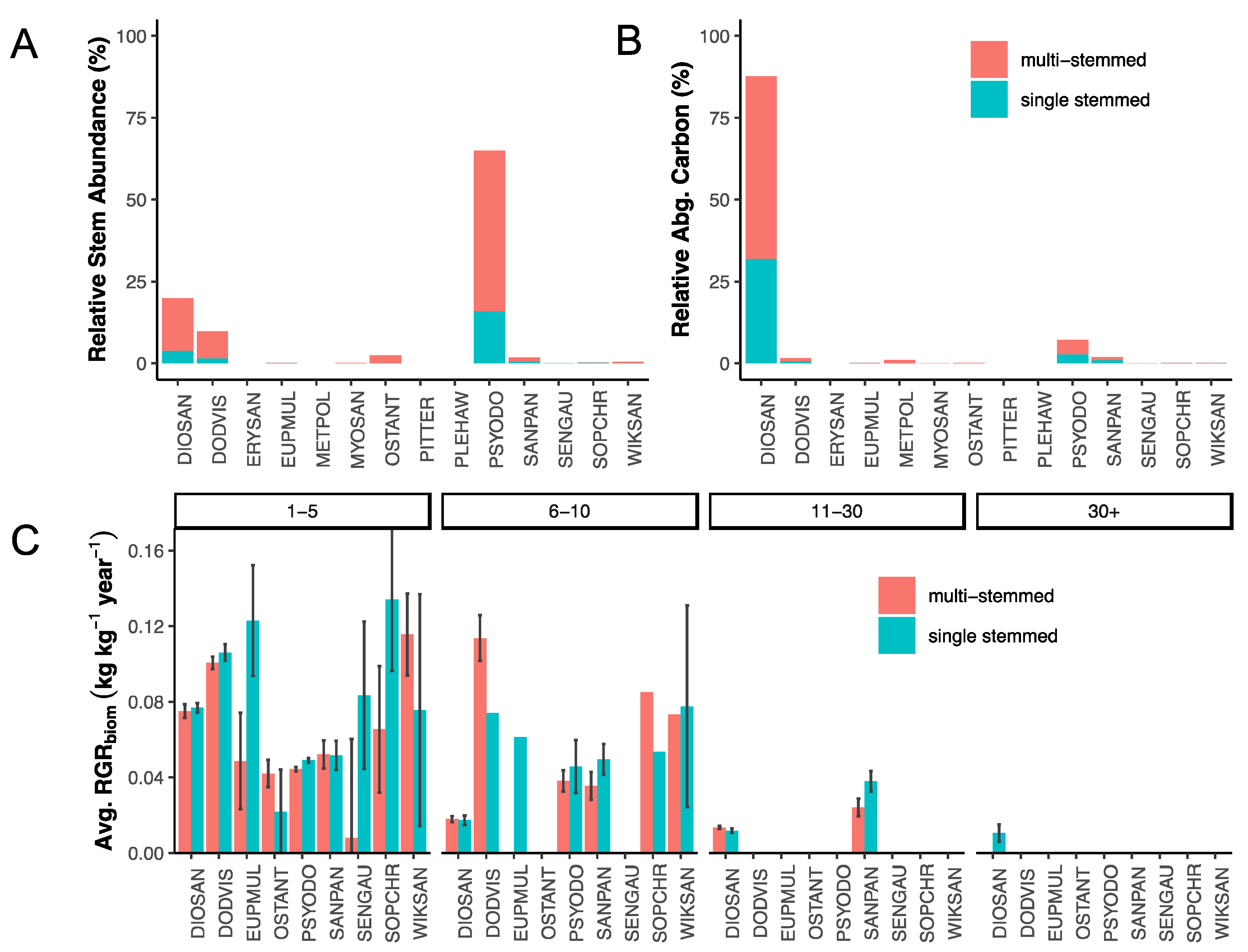
3.1. Stem Abundance and Relative Growth Rate across Plots, Species, Tree Size Class, and Stem Habit
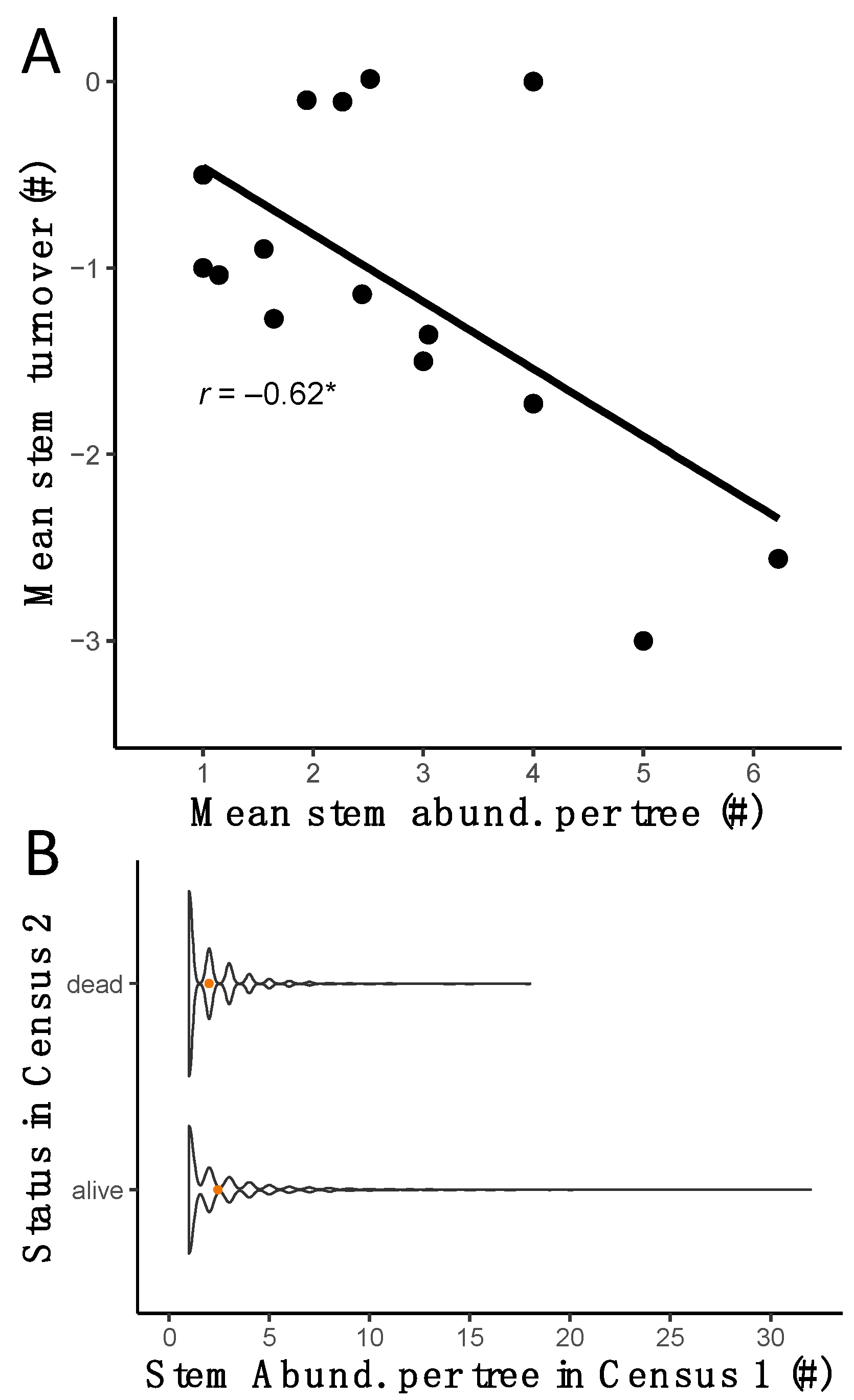
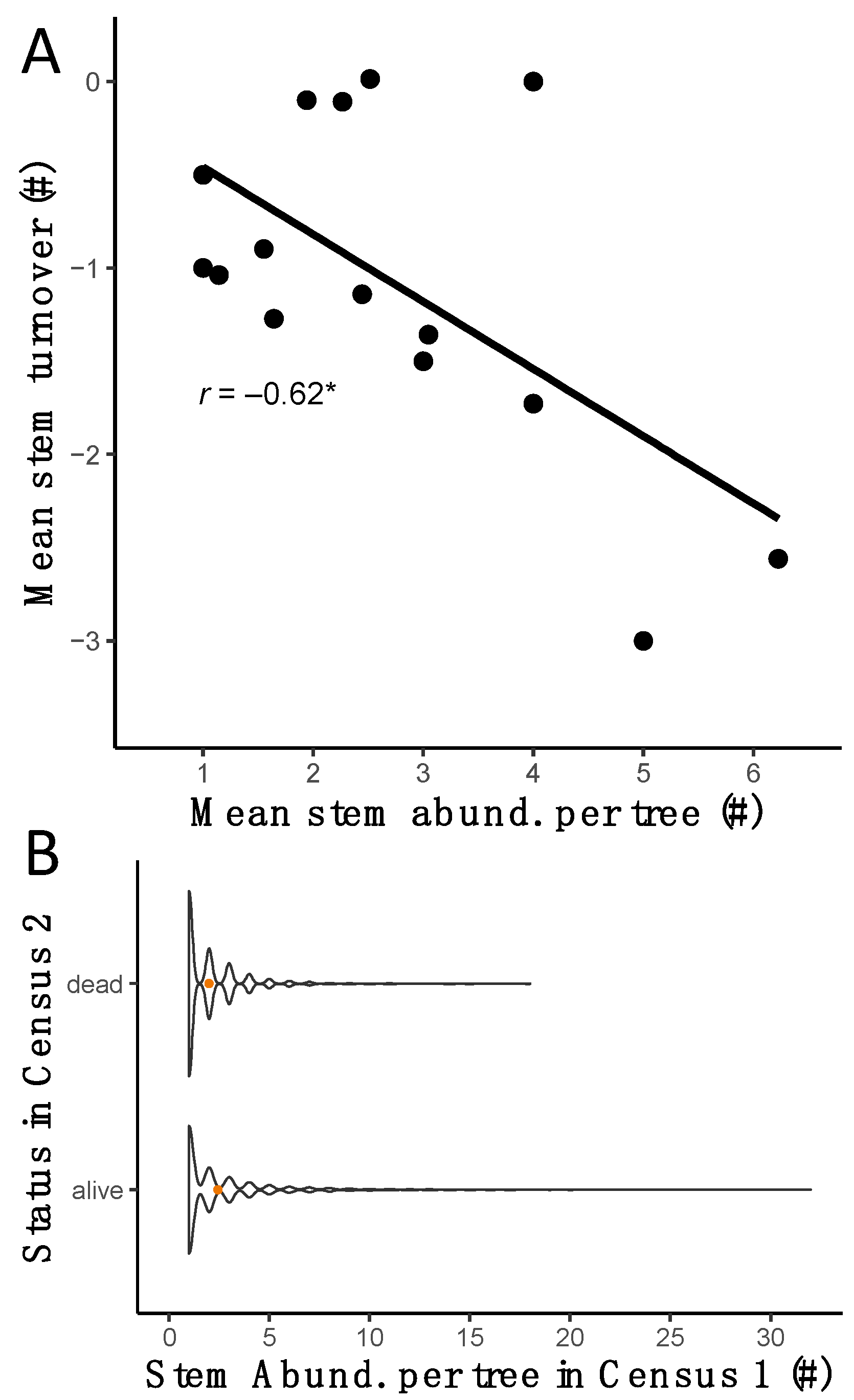
3.2. Stem Abundance and Turnover as Common Components of the Lowland Dry Forest
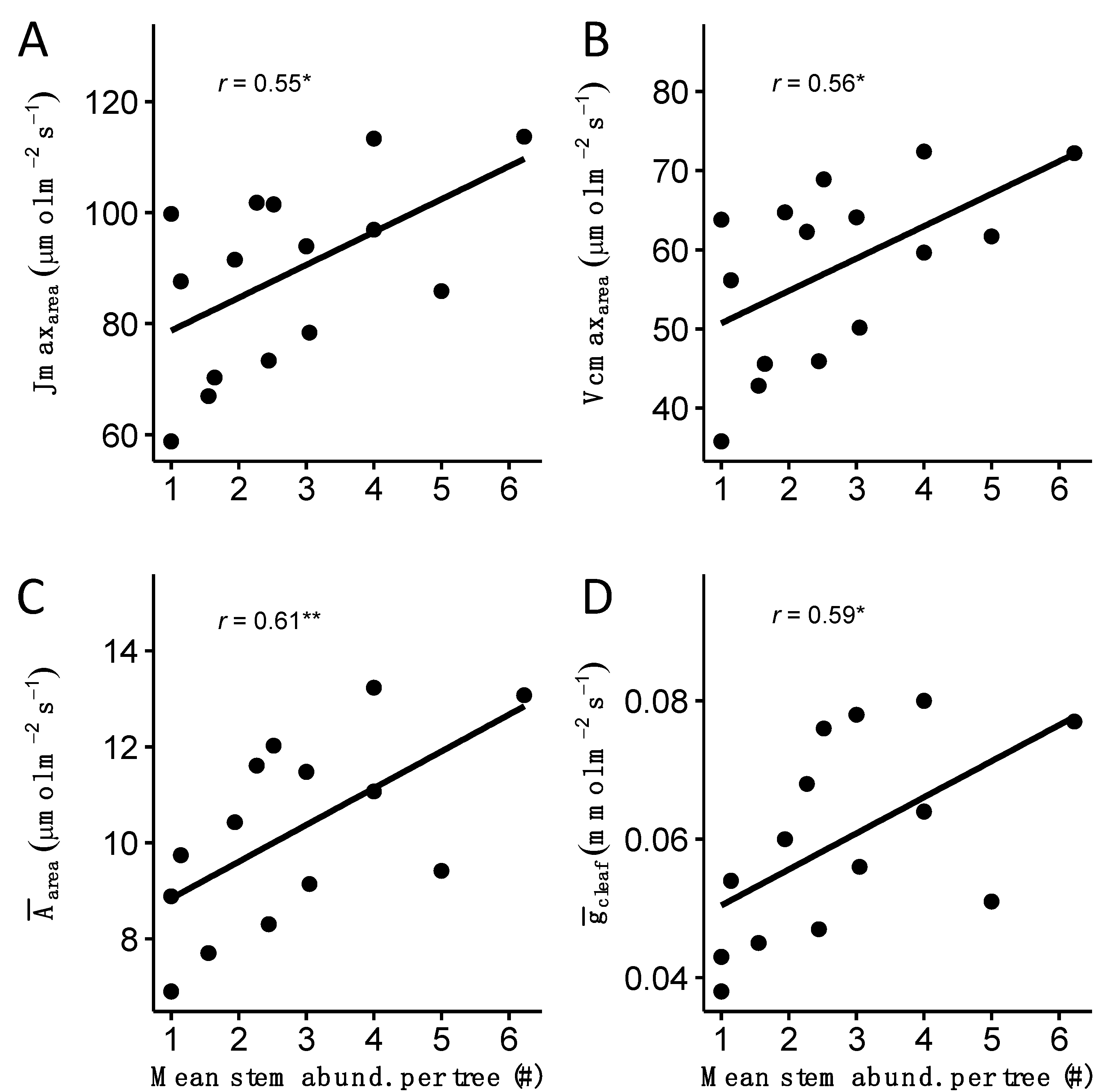
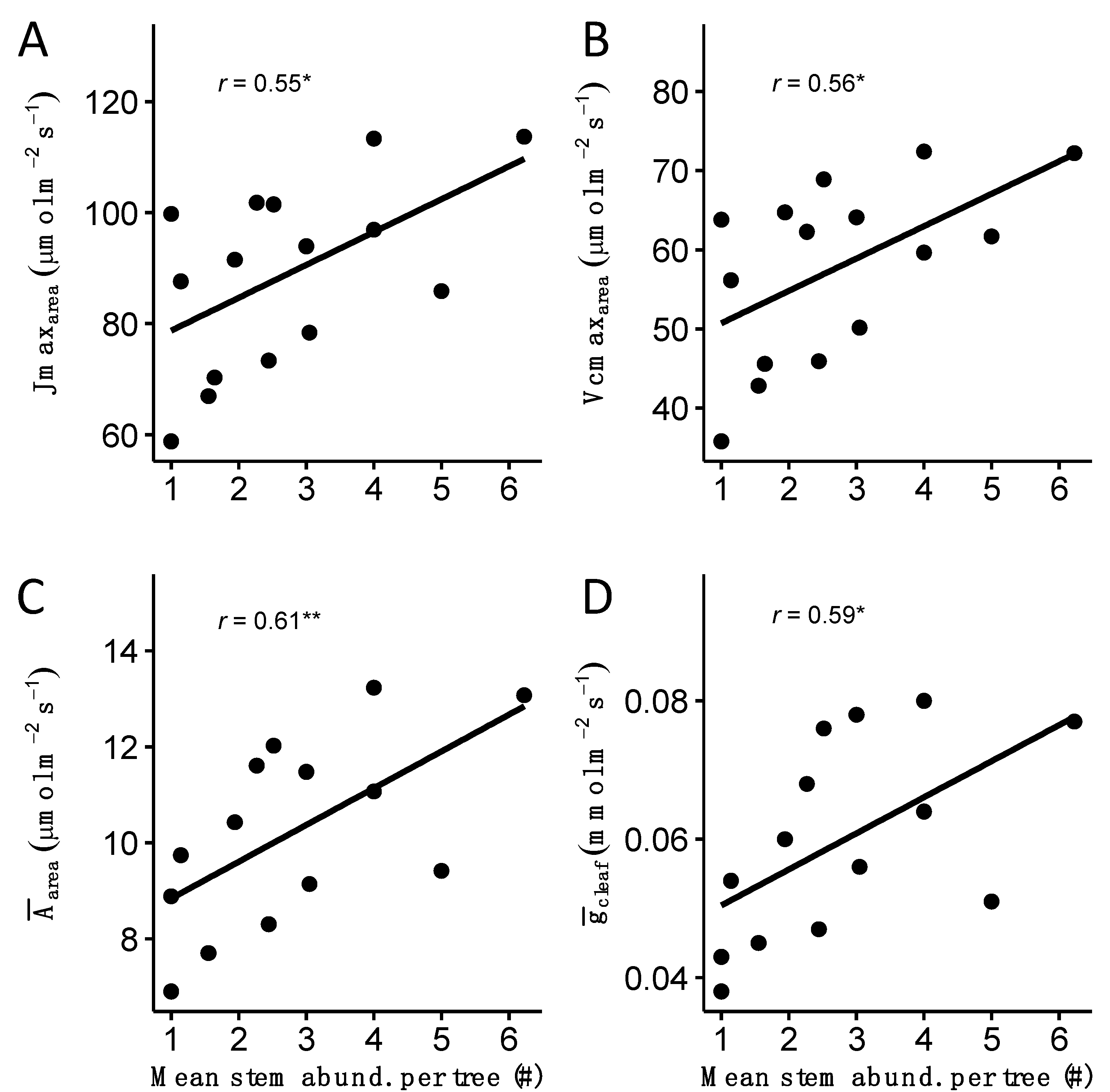
3.3. Trait–Trait Correlations
4. Discussion
4.1. Multi-Stemmed Trees Are an Important Structural and Functional Component of the Hawaiian Lowland Dry Forest
4.2. Multiple-Stemmed Trees Possess Traits That Allow for Persistence in Dry Environments
4.3. Multi-Stemmed Trees and Implications for Restoration Practice in Tropical Dry Forests
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gilman, S.E.; Urban, M.C.; Tewksbury, J.; Gilchrist, G.W.; Holt, R.D. A Framework for Community Interactions under Climate Change. Trends Ecol. Evol. 2010, 25, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Lavergne, S.; Mouquet, N.; Thuiller, W.; Ronce, O. Biodiversity and Climate Change: Integrating Evolutionary and Ecological Responses of Species and Communities. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 321–350. [Google Scholar] [CrossRef] [Green Version]
- Woolbright, S.A.; Whitham, T.G.; Gehring, C.A.; Allan, G.J.; Bailey, J.K. Climate Relicts and Their Associated Communities as Natural Ecology and Evolution Laboratories. Trends Ecol. Evol. 2014, 29, 406–416. [Google Scholar] [CrossRef] [PubMed]
- Pausas, J.G.; Pratt, R.B.; Keeley, J.E.; Jacobsen, A.L.; Ramirez, A.R.; Vilagrosa, A.; Paula, S.; Kaneakua-Pia, I.N.; Davis, S.D. Towards Understanding Resprouting at the Global Scale. New Phytol. 2016, 209, 945–954. [Google Scholar] [CrossRef] [Green Version]
- Bellingham, P.J.; Sparrow, A.D. Resprouting as a Life History Strategy in Woody Plant Communities. Oikos 2000, 89, 409–416. [Google Scholar] [CrossRef]
- Bellingham, P.J.; Sparrow, A.D. Multi-Stemmed Trees in Montane Rain Forests: Their Frequency and Demography in Relation to Elevation, Soil Nutrients and Disturbance. J. Ecol. 2009, 97, 472–483. [Google Scholar] [CrossRef]
- Clarke, P.J.; Lawes, M.J.; Midgley, J.J.; Lamont, B.B.; Ojeda, F.; Burrows, G.E.; Enright, N.J.; Knox, K.J.E. Resprouting as a Key Functional Trait: How Buds, Protection and Resources Drive Persistence after Fire. New Phytol. 2013, 197, 19–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keeley, J.; Bond, W.; Bradstock, R.; Pausas, J.G.; Rundel, P. Fire in Mediterranean Ecosystems; Ecology, Evolution and Management. In Fire in Mediterranean Ecosystems: Ecology, Evolution and Management; Cambridge University Press: Cambridge, UK, 2011; pp. 1–515. [Google Scholar] [CrossRef]
- Zeppel, M.J.B.; Harrison, S.P.; Adams, H.D.; Kelley, D.I.; Li, G.; Tissue, D.T.; Dawson, T.E.; Fensham, R.; Medlyn, B.E.; Palmer, A.; et al. Drought and Resprouting Plants. New Phytol. 2015, 206, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Pausas, J.G.; Keeley, J.E. Evolutionary Ecology of Resprouting and Seeding in Fire-prone Ecosystems. New Phytol. 2014, 204, 55–65. [Google Scholar] [CrossRef]
- Bond, W.J.; Wilgen, B.W. Fire and Plants; Springer: New York, NY, USA, 2012; ISBN 978-94-009-1499-5. [Google Scholar]
- Kennard, D.K.; Gould, K.; Putz, F.E.; Fredericksen, T.S.; Morales, F. Effect of Disturbance Intensity on Regeneration Mechanisms in a Tropical Dry Forest. For. Ecol. Manag. 2002, 162, 197–208. [Google Scholar] [CrossRef] [Green Version]
- Van Bloem, S.J.; Murphy, P.G.; Lugo, A.E.; Ostertag, R.; Costa, M.R.; Bernard, I.R.; Colón, S.M.; Mora, M.C. The Influence of Hurricane Winds on Caribbean Dry Forest Structure and Nutrient Pools1. Biotropica 2005, 37, 571–583. [Google Scholar] [CrossRef]
- Fornara, D.A.; Du Toit, J.T. Responses of Woody Saplings Exposed to Chronic Mammalian Herbivory in an African Savanna. Écoscience 2008, 15, 129–135. [Google Scholar] [CrossRef]
- Bond, W.J.; Midgley, J.J. Ecology of Sprouting in Woody Plants: The Persistence Niche. Trends Ecol. Evol. 2001, 16, 45–51. [Google Scholar] [CrossRef]
- Vilagrosa, A.; Hernández, E.I.; Luis, V.C.; Cochard, H.; Pausas, J.G. Physiological Differences Explain the Co-Existence of Different Regeneration Strategies in Mediterranean Ecosystems. New Phytol. 2014, 201, 1277–1288. [Google Scholar] [CrossRef] [PubMed]
- Knox, K.J.E.; Clarke, P.J. Nutrient Availability Induces Contrasting Allocation and Starch Formation in Resprouting and Obligate Seeding Shrubs. Funct. Ecol. 2005, 19, 690–698. [Google Scholar] [CrossRef]
- Pratt, R.B.; Jacobsen, A.L.; Hernandez, J.; Ewers, F.W.; North, G.B.; Davis, S.D. Allocation Tradeoffs among Chaparral Shrub Seedlings with Different Life History Types (Rhamnaceae). Am. J. Bot. 2012, 99, 1464–1476. [Google Scholar] [CrossRef] [Green Version]
- West, A.G.; Dawson, T.E.; February, E.C.; Midgley, G.F.; Bond, W.J.; Aston, T.L. Diverse Functional Responses to Drought in a Mediterranean-Type Shrubland in South Africa. New Phytol. 2012, 195, 396–407. [Google Scholar] [CrossRef]
- Wilson, E.O.; Peter, F.M. Tropical Dry Forests the Most Endangered Major Tropical Ecosystem; National Academies Press: Washington, DC, USA, 1988. [Google Scholar]
- Gillespie, T.W.; Lipkin, B.; Sullivan, L.; Benowitz, D.R.; Pau, S.; Keppel, G. The Rarest and Least Protected Forests in Biodiversity Hotspots. Biodivers. Conserv. 2012, 21, 3597–3611. [Google Scholar] [CrossRef]
- Blackie, R.; Baldauf, C.; Gautier, D.; Gumbo, D.; Kassa, H.; Parthasarathy, N.; Paumgarten, F.; Sola, P.; Pulla, S.; Waeber, P.; et al. Tropical Dry Forests: The State of Global Knowledge and Recommendations for Future Research; CIFOR: Bogor, Indonesia, 2014. [Google Scholar]
- Libby, R.; Sato, A.Y.; Alapai, L.; Brawner, W.P.; Carter, Y.Y.; Carter, K.A.; Tomich, K.; Ticktin, T. A Hawaiian Tropical Dry Forest Regenerates: Natural Regeneration of Endangered Species under Biocultural Restoration. Sustainability 2022, 14, 1159. [Google Scholar] [CrossRef]
- Reich, P.B.; Borchert, R. Water Stress and Tree Phenology in a Tropical Dry Forest in the Lowlands of Costa Rica. J. Ecol. 1984, 72, 61–74. [Google Scholar] [CrossRef]
- Murphy, P.G.; Lugo, A.E. Ecology of Tropical Dry Forest. Annu. Rev. Ecol. Syst. 1986, 17, 67–88. [Google Scholar] [CrossRef]
- Ostertag, R.; Inman-Narahari, F.; Cordell, S.; Giardina, C.P.; Sack, L. Forest Structure in Low-Diversity Tropical Forests: A Study of Hawaiian Wet and Dry Forests. PLoS ONE 2014, 9, e103268. [Google Scholar] [CrossRef] [Green Version]
- Sandquist, D.R.; Cordell, S. Functional Diversity of Carbon-Gain, Water-Use, and Leaf-Allocation Traits in Trees of a Threatened Lowland Dry Forest in Hawaii. Am. J. Bot. 2007, 94, 1459–1469. [Google Scholar] [CrossRef] [PubMed]
- Busby, P.E.; Vitousek, P.; Dirzo, R. Prevalence of Tree Regeneration by Sprouting and Seeding Along a Rainfall Gradient in Hawai’i. Biotropica 2010, 42, 80–86. [Google Scholar] [CrossRef]
- Frazier, A.G.; Giambelluca, T.W. Spatial Trend Analysis of Hawaiian Rainfall from 1920 to 2012. Int. J. Climatol. 2017, 37, 2522–2531. [Google Scholar] [CrossRef]
- Mesa-Sierra, N.; de la Peña-Domene, M.; Campo, J.; Giardina, C.P. Restoring Mexican Tropical Dry Forests: A National Review. Sustainability 2022, 14, 3937. [Google Scholar] [CrossRef]
- Freifelder, R.R.; Vitousek, P.M.; D’Antonio, C.M. Microclimate Change and Effect on Fire Following Forest-Grass Conversion in Seasonally Dry Tropical Woodland1. Biotropica 1998, 30, 286–297. [Google Scholar] [CrossRef]
- Litton, C.M.; Sandquist, D.R.; Cordell, S. Effects of Non-Native Grass Invasion on Aboveground Carbon Pools and Tree Population Structure in a Tropical Dry Forest of Hawaii. For. Ecol. Manag. 2006, 231, 105–113. [Google Scholar] [CrossRef]
- Kellner, J.R.; Asner, G.P. Convergent Structural Responses of Tropical Forests to Diverse Disturbance Regimes. Ecol. Lett. 2009, 12, 887–897. [Google Scholar] [CrossRef]
- Mueller-Dombois, D. Rain Forest Establishment and Succession in the Hawaiian Islands. Landsc. Urban Plan. 2000, 51, 147–157. [Google Scholar] [CrossRef]
- Friday, J.B.; Scowcroft, P.G.; Ares, A. Responses of Native and Invasive Plant Species to Selective Logging in an Acacia Koa-Metrosideros Polymorpha Forest in Hawai’i. Appl. Veg. Sci. 2008, 11, 471–482. [Google Scholar] [CrossRef]
- Murphy, M.J.; Inman-Narahari, F.; Ostertag, R.; Litton, C.M. Invasive Feral Pigs Impact Native Tree Ferns and Woody Seedlings in Hawaiian Forest. Biol. Invasions 2014, 16, 63–71. [Google Scholar] [CrossRef]
- Réjou-Méchain, M.; Tanguy, A.; Piponiot, C.; Chave, J.; Hérault, B. Biomass: An r Package for Estimating above-Ground Biomass and Its Uncertainty in Tropical Forests. Methods Ecol. Evol. 2017, 8, 1163–1167. [Google Scholar] [CrossRef]
- Shen, Y.; Yu, S.; Lian, J.; Shen, H.; Cao, H.; Lu, H.; Ye, W. Tree Aboveground Carbon Storage Correlates with Environmental Gradients and Functional Diversity in a Tropical Forest. Sci. Rep. 2016, 6, 25304. [Google Scholar] [CrossRef] [Green Version]
- Chave, J.; Réjou-Méchain, M.; Búrquez, A.; Chidumayo, E.; Colgan, M.S.; Delitti, W.B.C.; Duque, A.; Eid, T.; Fearnside, P.M.; Goodman, R.C.; et al. Improved Allometric Models to Estimate the Aboveground Biomass of Tropical Trees. Glob. Chang. Biol. 2014, 20, 3177–3190. [Google Scholar] [CrossRef]
- Cavanaugh, K.C.; Gosnell, J.S.; Davis, S.L.; Ahumada, J.; Boundja, P.; Clark, D.B.; Mugerwa, B.; Jansen, P.A.; O’Brien, T.G.; Rovero, F.; et al. Carbon Storage in Tropical Forests Correlates with Taxonomic Diversity and Functional Dominance on a Global Scale. Glob. Ecol. Biogeogr. 2014, 23, 563–573. [Google Scholar] [CrossRef]
- Fiske, I.J.; Bruna, E.M.; Bolker, B.M. Effects of Sample Size on Estimates of Population Growth Rates Calculated with Matrix Models. PLoS ONE 2008, 3, e3080. [Google Scholar] [CrossRef]
- Medeiros, C.D.; Scoffoni, C.; John, G.P.; Bartlett, M.K.; Inman-Narahari, F.; Ostertag, R.; Cordell, S.; Giardina, C.; Sack, L. An Extensive Suite of Functional Traits Distinguishes Hawaiian Wet and Dry Forests and Enables Prediction of Species Vital Rates. Funct. Ecol. 2019, 33, 712–734. [Google Scholar] [CrossRef]
- Armbruster, W.; Pélabon, C.; Bolstad, G.; Hansen, T. Integrated Phenotypes: Understanding Trait Covariation in Plants and Animals. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 245. [Google Scholar] [CrossRef] [Green Version]
- R: The R Project for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 1 April 2022).
- Frazier, A.G.; Giambelluca, T.W.; Diaz, H.F.; Needham, H.L. Comparison of Geostatistical Approaches to Spatially Interpolate Month-Year Rainfall for the Hawaiian Islands. Int. J. Climatol. 2016, 36, 1459–1470. [Google Scholar] [CrossRef] [Green Version]
- Gharun, M.; Turnbull, T.L.; Adams, M.A. Stand Water Use Status in Relation to Fire in a Mixed Species Eucalypt Forest. For. Ecol. Manag. 2013, 304, 162–170. [Google Scholar] [CrossRef]
- Heath, J.T.; Chafer, C.J.; Bishop, T.F.A.; Van Ogtrop, F.F. Post-Fire Recovery of Eucalypt-Dominated Vegetation Communities in the Sydney Basin, Australia. Fire Ecol. 2016, 12, 53–79. [Google Scholar] [CrossRef]
- Kelley, D.I.; Harrison, S.P.; Prentice, I.C. Improved Simulation of Fire–Vegetation Interactions in the Land Surface Processes and EXchanges Dynamic Global Vegetation Model (LPX-Mv1). Geosci. Model Dev. 2014, 7, 2411–2433. [Google Scholar] [CrossRef] [Green Version]
- Philipson, C.D.; Dent, D.H.; O’Brien, M.J.; Chamagne, J.; Dzulkifli, D.; Nilus, R.; Philips, S.; Reynolds, G.; Saner, P.; Hector, A. A Trait-Based Trade-off between Growth and Mortality: Evidence from 15 Tropical Tree Species Using Size-Specific Relative Growth Rates. Ecol. Evol. 2014, 4, 3675–3688. [Google Scholar] [CrossRef] [Green Version]
- Wright, S.J.; Kitajima, K.; Kraft, N.J.B.; Reich, P.B.; Wright, I.J.; Bunker, D.E.; Condit, R.; Dalling, J.W.; Davies, S.J.; Díaz, S.; et al. Functional Traits and the Growth–Mortality Trade-off in Tropical Trees. Ecology 2010, 91, 3664–3674. [Google Scholar] [CrossRef]
- Nzunda, E.F.; Griffiths, M.E.; Lawes, M.J. Sprouting by Remobilization of Above-Ground Resources Ensures Persistence after Disturbance of Coastal Dune Forest Trees. Funct. Ecol. 2008, 22, 577–582. [Google Scholar] [CrossRef]
- Vieira, D.L.M.; Scariot, A. Principles of Natural Regeneration of Tropical Dry Forests for Restoration. Restor. Ecol. 2006, 14, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Allen, K.; Dupuy, J.M.; Gei, M.G.; Hulshof, C.; Medvigy, D.; Pizano, C.; Salgado-Negret, B.; Smith, C.M.; Trierweiler, A.; Bloem, S.J.V.; et al. Will Seasonally Dry Tropical Forests Be Sensitive or Resistant to Future Changes in Rainfall Regimes? Environ. Res. Lett. 2017, 12, 023001. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Response: stem abundance | DF | χ2 | Pr(>F) |
|---|---|---|---|
| Species | 13 | 417.01 | <0.0001 |
| Tree size class | 3 | 401.57 | <0.0001 |
| Species × tree size class | 8 | 43.55 | <0.0001 |
| Response: stem abundance | |||
| Species | 13 | 206.5 | <0.0001 |
| Stem habit | 1 | 9442.1 | <0.0001 |
| Species × stem habit | 9 | 24.9 | 0.003 |
| Response: relative growth rate (RGRbiom) | |||
| Species | 12 | 411.63 | <0.0001 |
| Stem habit | 2 | 22.73 | <0.0001 |
| Species × stem habit | 9 | 49.63 | <0.0001 |
| Response: stem turnover | |||
| Species | 9 | 332.06 | <0.0001 |
| Tree size class | 2 | 26.80 | <0.0001 |
| Species × tree size class | 7 | 10.72 | 0.15 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ware, I.M.; Ostertag, R.; Cordell, S.; Giardina, C.P.; Sack, L.; Medeiros, C.D.; Inman, F.; Litton, C.M.; Giambelluca, T.; John, G.P.; et al. Multi-Stemmed Habit in Trees Contributes Climate Resilience in Tropical Dry Forest. Sustainability 2022, 14, 6779. https://doi.org/10.3390/su14116779
Ware IM, Ostertag R, Cordell S, Giardina CP, Sack L, Medeiros CD, Inman F, Litton CM, Giambelluca T, John GP, et al. Multi-Stemmed Habit in Trees Contributes Climate Resilience in Tropical Dry Forest. Sustainability. 2022; 14(11):6779. https://doi.org/10.3390/su14116779
Chicago/Turabian StyleWare, Ian M., Rebecca Ostertag, Susan Cordell, Christian P. Giardina, Lawren Sack, Camila D. Medeiros, Faith Inman, Creighton M. Litton, Thomas Giambelluca, Grace P. John, and et al. 2022. "Multi-Stemmed Habit in Trees Contributes Climate Resilience in Tropical Dry Forest" Sustainability 14, no. 11: 6779. https://doi.org/10.3390/su14116779
APA StyleWare, I. M., Ostertag, R., Cordell, S., Giardina, C. P., Sack, L., Medeiros, C. D., Inman, F., Litton, C. M., Giambelluca, T., John, G. P., & Scoffoni, C. (2022). Multi-Stemmed Habit in Trees Contributes Climate Resilience in Tropical Dry Forest. Sustainability, 14(11), 6779. https://doi.org/10.3390/su14116779