
Halotolerant Rhizobacteria for Salinity-Stress Mitigation: Diversity, Mechanisms and Molecular Approaches

 ,
,  ,
,  and
and
Abstract
1. Introduction
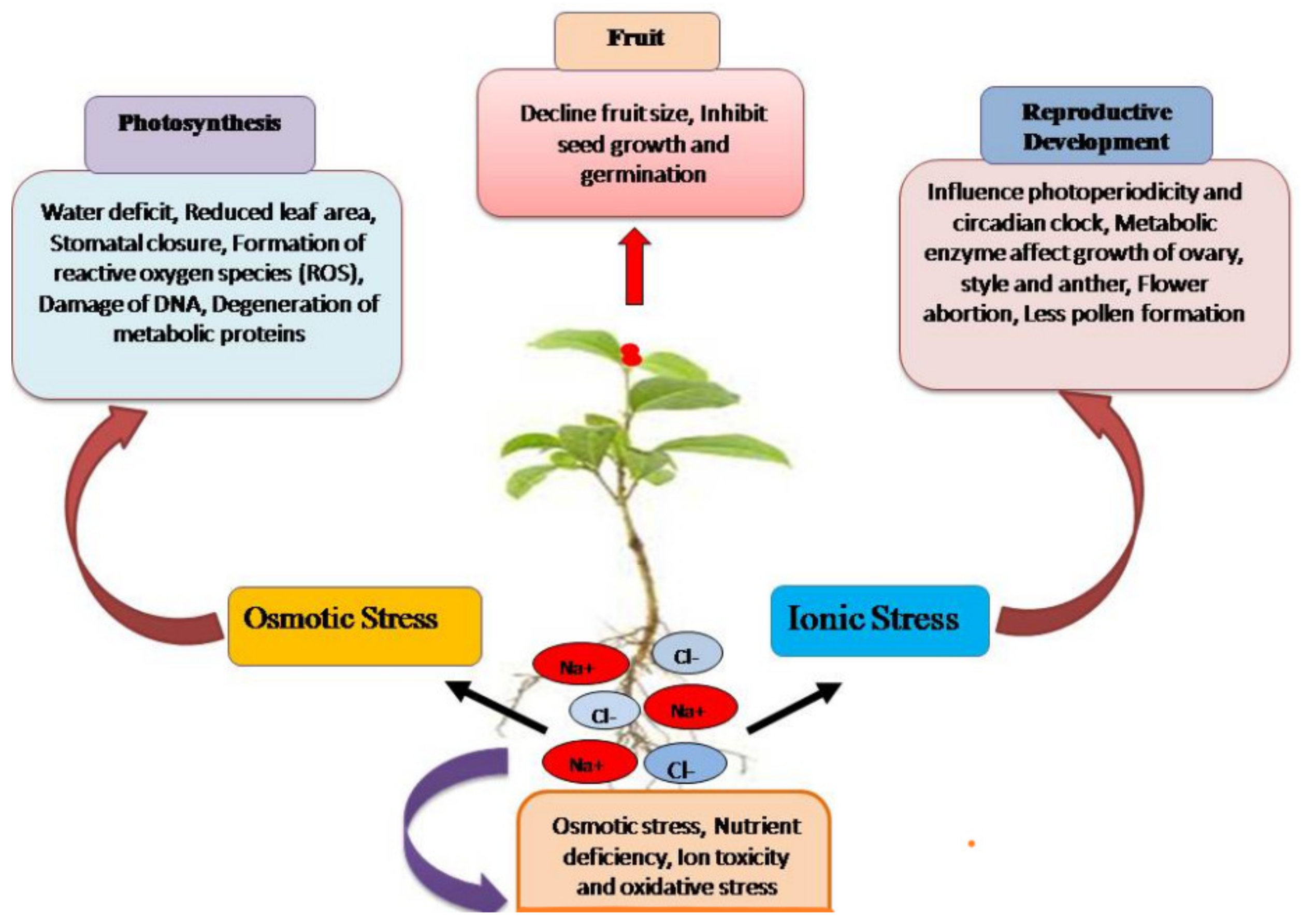
2. Impact of Salinity on Plant Growth and Development
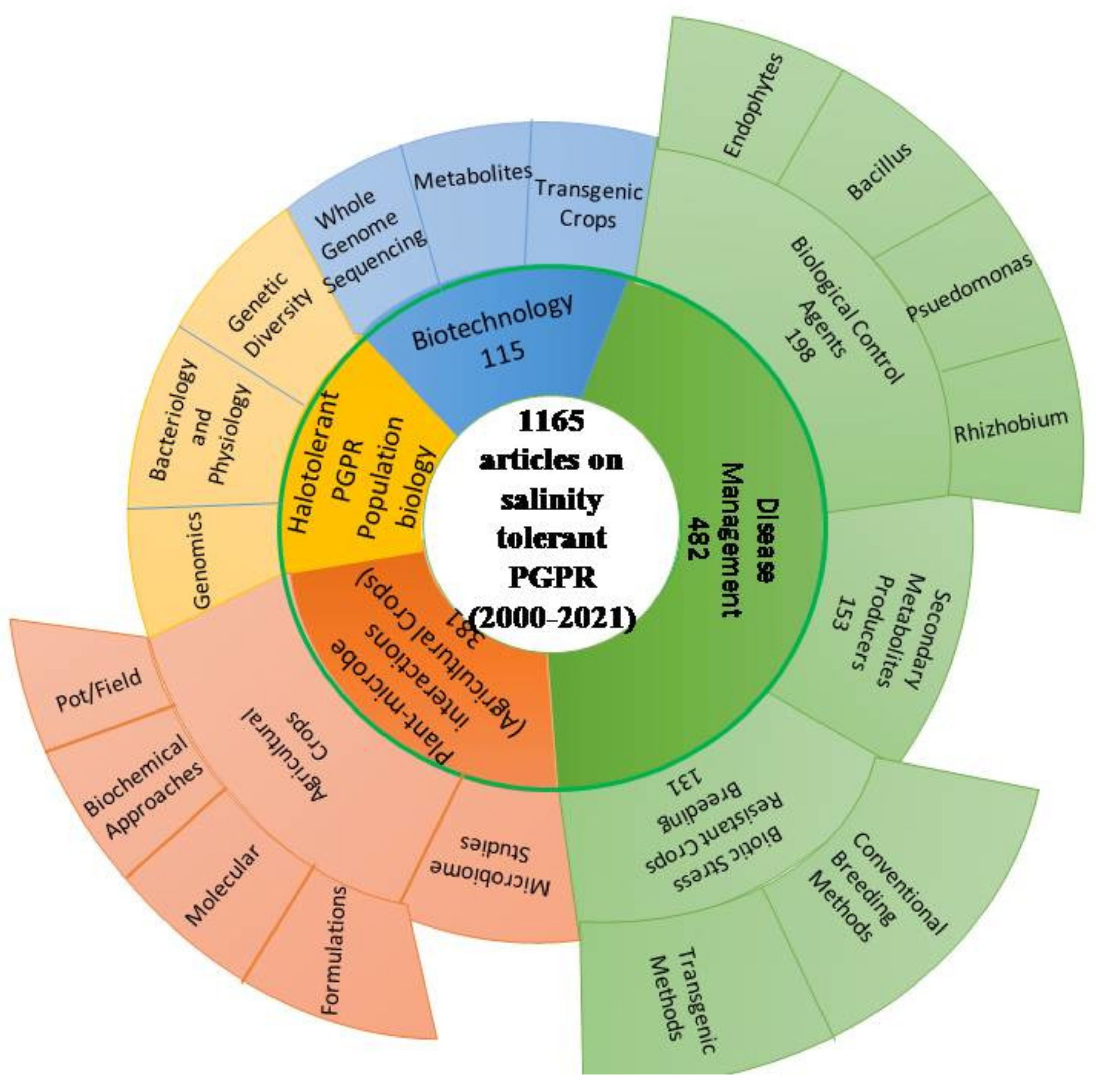
3. Halotolerant/Salinity-Tolerant PGPB
Biodiversity of Salt-Tolerant Plant Growth-Promoting Bacteria [PGPB]
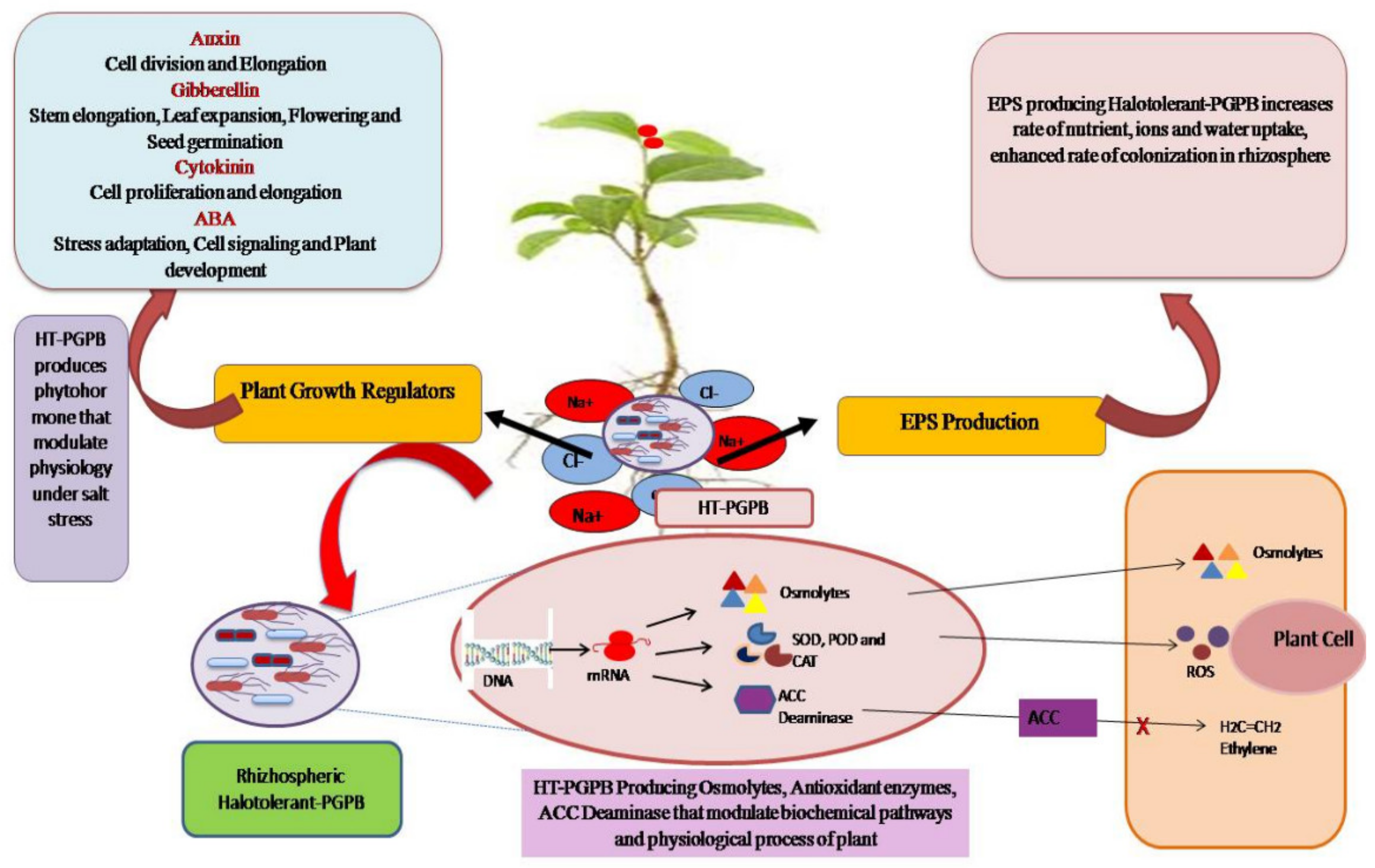
4. Mechanisms of Halotolerant PGPB-Mediated Salinity-Stress Tolerance
4.1. Phytohormone Production
4.1.1. Auxin [Indole Acetic Acid, IAA]
4.1.2. Cytokinin
4.1.3. Antioxidant Activity
4.1.4. Osmoprotectants
4.1.5. Exopolysaccharide (EPS) Production
4.1.6. 1-Aminocyclopropane-1-Carboxylic Deaminase [ACCD]
5. Role of Halotolerant PGPB in the Enhancement of Crop Production under Salinity
6. Molecular Understanding of Halotolerant PGPB-Mediated Salt Tolerance in Plants
7. Conclusions and Future Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shah, F.; Wu, W. Soil and Crop Management Strategies to Ensure Higher Crop Productivity within Sustainable Environments. Sustainability 2019, 11, 1485. [Google Scholar] [CrossRef]
- Wei, B.; Yu, J.; Cao, Z.; Meng, M.; Yang, L.; Chen, Q. The Availability and Accumulation of Heavy Metals in Greenhouse Soils Associated with Intensive Fertilizer Application. Int. J. Environ. Res. Public Health 2020, 17, 5359. [Google Scholar] [CrossRef] [PubMed]
- Isayenkov, S.V.; Maathuis, F.J.M. Plant Salinity Stress: Many Unanswered Questions Remain. Front. Plant Sci. 2019, 10, 80. [Google Scholar] [CrossRef] [PubMed]
- Shahid, S.A.; Zaman, M.; Heng, L. Soil Salinity: Historical Perspectives and a World Overview of the Problem. In Guideline for Salinity Assessment, Mitigation and Adaptation Using Nuclear and Related Techniques; Springer: Cham, Switzerland, 2018; pp. 43–53. [Google Scholar]
- Kumar, P.; Sharma, P.K. Soil Salinity and Food Security in India. Front. Sustain. Food Syst. 2020, 4, 174. [Google Scholar] [CrossRef]
- Sunita, K.; Mishra, I.; Mishra, J.; Prakash, J.; Arora, N.K. Secondary Metabolites From Halotolerant Plant Growth Promoting Rhizobacteria for Ameliorating Salinity Stress in Plants. Front. Microbiol. 2020, 11, 567768. [Google Scholar] [CrossRef]
- Panagos, P.; Borrelli, P.; Robinson, D. FAO calls for actions to reduce global soil erosion. Mitig. Adapt. Strat. Glob. Chang. 2019, 25, 789–790. [Google Scholar] [CrossRef]
- Mandal, S.; Raju, R.; Kumar, A.; Kumar, P.; Sharma, P.C. Current status of research, technology response and policy needs of salt-affected soils in India—A review. Ind. Soc. Coast. Agric. Res. 2018, 36, 40–53. [Google Scholar]
- Szabo, S.; Hossain, M.S.; Adger, W.N.; Matthews, Z.; Ahmed, S.; Lázár, A.N.; Ahmad, S. Soil salinity, household wealth and food insecurity in tropical deltas: Evidence from south-west coast of Bangladesh. Sustain. Sci. 2016, 11, 411–421. [Google Scholar] [CrossRef]
- Zörb, C.; Geilfus, C.-M.; Dietz, K.-J. Salinity and crop yield. Plant Biol. 2019, 21, 31–38. [Google Scholar] [CrossRef]
- Arora, S.; Sharma, V. Reclamation and management of salt-affected soils for safeguarding agricultural productivity. J. Safe Agric. 2017, 1, 1–10. [Google Scholar]
- Schirawski, J.; Perlin, M.H. Plant–Microbe Interaction 2017—The Good, the Bad and the Diverse. Int. J. Mol. Sci. 2018, 19, 1374. [Google Scholar] [CrossRef]
- Rütting, T.; Aronsson, H.; Delin, S. Efficient use of nitrogen in agriculture. Nutr. Cycl. Agroecosyst. 2018, 110, 1–5. [Google Scholar] [CrossRef]
- Arora, N.K.; Fatima, T.; Mishra, I.; Verma, M.; Mishra, J.; Mishra, V. Environmental sustainability: Challenges and viable solutions. Environ. Sustain. 2018, 1, 309–340. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Wirth, S.J.; Bellingrath-Kimura, D.S.D.; Mishra, J.; Arora, N.K. Salt-Tolerant Plant Growth Promoting Rhizobacteria for Enhancing Crop Productivity of Saline Soils. Front. Microbiol. 2019, 18, 2791. [Google Scholar] [CrossRef]
- Vaishnav, A.; Kumari, S.; Jain, S.; Varma, A.; Tuteja, N.; Choudhary, D.K. PGPR-mediated expression of salt tolerance gene in soy-bean through volatiles under sodium nitroprusside. J. Basic Microbiol. 2016, 56, 1274–1288. [Google Scholar] [CrossRef]
- Ayyam, V.; Palanivel, S.; Chandrakasan, S. Approaches in land degradation management for productivity enhance-ment. In Coastal Ecosystems of the Tropics—Adaptive Management; Ayyam, V., Palanivel, S., Chandrakasan, S., Eds.; Springer: Singapore, 2019. [Google Scholar]
- Shilev, S. Plant-Growth-Promoting Bacteria Mitigating Soil Salinity Stress in Plants. Appl. Sci. 2020, 10, 7326. [Google Scholar] [CrossRef]
- Wang, C.; Wang, H.; Li, Y.; Li, Q.; Yan, W.; Zhang, Y.; Wu, Z.; Zhou, Q. Plant growth-promoting rhizobacteria isolation from rhizosphere of submerged macrophytes and their growth-promoting effect on Vallisneria natans under high sediment organic matter load. Microb. Biotechnol. 2021, 14, 726–736. [Google Scholar] [CrossRef]
- Ha-Tran, D.M.; Nguyen, T.T.M.; Hung, S.-H.; Huang, E.; Huang, C.-C. Roles of Plant Growth-Promoting Rhizobacteria (PGPR) in Stimulating Salinity Stress Defense in Plants: A Review. Int. J. Mol. Sci. 2021, 22, 3154. [Google Scholar] [CrossRef]
- Machado, R.; Serralheiro, R. Soil Salinity: Effect on Vegetable Crop Growth. Management Practices to Prevent and Mitigate Soil Salinization. Horticulturae 2017, 3, 30. [Google Scholar] [CrossRef]
- El-Esawi, M.A.; Alaraidh, I.A.; Alsahli, A.A.; Alamri, S.A.; Ali, H.M.; Alayafi, A.A. Bacillus firmus (SW5) augments salt tol-erance in soybean (Glycine max L.) by modulating root system architecture, antioxidant defense systems and stress-responsive genes expressin. Plant Physiol. Biochem. 2018, 132, 375–384. [Google Scholar] [CrossRef]
- Mokrani, S.; Nabti, E.; Cruz, C. Current Advances in Plant Growth Promoting Bacteria Alleviating Salt Stress for Sustainable Agriculture. Appl. Sci. 2020, 10, 7025. [Google Scholar] [CrossRef]
- Muchate, N.S.; Nikalje, G.C.; Rajurkar, N.S.; Suprasanna, P.; Nikam, T.D. Plant Salt Stress: Adaptive Responses, Tolerance Mechanism and Bioengineering for Salt Tolerance. Bot. Rev. 2016, 82, 371–406. [Google Scholar] [CrossRef]
- Bharti, N.; Pandey, S.S.; Barnawal, D.; Patel, V.K.; Kalra, A. Plant growth promoting rhizobacteria Dietzia natronolimnaea modulates the expression of stress responsive genes providing protection of wheat from salinity stress. Sci. Rep. 2016, 6, 34768. [Google Scholar] [CrossRef]
- Ilangumaran, G.; Smith, D.L. Plant Growth Promoting Rhizobacteria in Amelioration of Salinity Stress: A Systems Biology Perspective. Front. Plant Sci. 2017, 8, 1768. [Google Scholar] [CrossRef]
- Vaishnav, A.; Varma, A.; Tuteja, N.; Choudhary, D.K. PGPR-Mediated Amelioration of Crops under Salt Stress. In Plant-Microbe Interaction: An Approach to Sustainable Agriculture; Springer: Singapore, 2016; Volume 10, pp. 205–226. [Google Scholar] [CrossRef]
- Almeida, D.M.; Oliveira, M.M.; Saibo, N.J.M. Regulation of Na+ and K+ homeostasis in plants: Towards improved salt stress tolerance in crop plants. Genet. Mol. Biol. 2017, 40, 326–345. [Google Scholar] [CrossRef]
- Numan, M.; Bashir, S.; Khan, Y.; Mumtaz, R.; Shinwari, Z.K.; Khan, A.L.; Khan, A.; Al-Harrasi, A. Plant growth promoting bacteria as an alternative strategy for salt tolerance in plants: A review. Microbiol. Res. 2018, 209, 21–32. [Google Scholar] [CrossRef]
- Kamran, M.; Parveen, A.; Ahmar, S.; Malik, Z.; Hussain, S.; Chattha, M.S.; Saleem, M.H.; Adil, M.; Heidari, P.; Chen, J.-T. An Overview of Hazardous Impacts of Soil Salinity in Crops, Tolerance Mechanisms, and Amelioration through Selenium Supplementation. Int. J. Mol. Sci. 2019, 21, 148. [Google Scholar] [CrossRef]
- Mirdar Mansuri, R.; Shobbar, Z.-S.; Babaeian Jelodar, N.; Ghaffari, M.; Mohammadi, S.M.; Daryani, P. Salt tolerance involved candidate genes in rice: An integrative meta-analysis approach. BMC Plant Biol. 2020, 20, 452. [Google Scholar] [CrossRef]
- Rai, S.; Solanki, A.K.; Anal, A.K.D. Modern biotechnological tools: An opportunity to discover complex phytobiomes of hor-ticulture crops. In Microbiomes and Plant Health: Panoply and Their Applications; Solanki, M.K., Kashyap, P.L., Ansari, R.A., Kumari, B., Eds.; Elsevier: Berkeley, CA, USA, 2020; pp. 85–124. [Google Scholar]
- Saxena, A.K.; Kumar, M.; Chakdar, H.; Anuroopa, N.; Bagyaraj, D.J. Bacillus species in soil as a natural resource for plant health and nutrition. J. Appl. Microbiol. 2020, 128, 1583–1594. [Google Scholar] [CrossRef]
- Arora, N.K.; Fatima, T.; Mishra, J.; Mishra, I.; Verma, S.; Verma, R.; Verma, M.; Bhattacharya, A.; Verma, P.; Mishra, P.; et al. Halo-tolerant plant growth promoting rhizobacteria for improving productivity and remediation of saline soils. J. Adv. Res. 2020, 26, 69–82. [Google Scholar] [CrossRef]
- Albdaiwi, R.N.; Khyami-Horani, H.; Ayad, J.Y. Plant Growth-Promoting Rhizobacteria: An Emerging Method for the Enhancement of Wheat Tolerance against Salinity Stress-(Review). Jordan J. Biol. Sci. 2019, 12, 525–534. [Google Scholar]
- Phour, M.; Sindhu, S.S. Amelioration of salinity stress and growth stimulation of mustard (Brassica juncea L.) by salt-tolerant Pseudomonas species. Appl. Soil Ecol. 2020, 149, 103518. [Google Scholar] [CrossRef]
- Zhang, S.; Fan, C.; Wang, Y.; Xia, Y.; Xiao, W.; Cui, X. Salt-tolerant and plant-growth-promoting bacteria isolated from high-yield paddy soil. Can. J. Microbiol. 2018, 64, 968–978. [Google Scholar] [CrossRef] [PubMed]
- Aslam, F.; Ali, B. Halotolerant Bacterial Diversity Associated with Suaeda fruticosa (L.) Forssk. Improved Growth of Maize under Salinity Stress. Agronomy 2018, 8, 131. [Google Scholar] [CrossRef]
- Vives-Peris, V.; Gomez-Cadenas, A.; Perez-Clemente, R.M. Salt stress alleviation in citrus plants by plant growth-promoting Pseudomonas putida and Novosphingobium sp. Plant Cell Rep. 2018, 37, 1557–1569. [Google Scholar] [CrossRef]
- Palacio-Rodríguez, R.; Coria-Arellano, J.L.; López-Bucio, J.; Sánchez-Salas, J.; Muro-Pérez, G.; Castañeda-Gaytán, G.; Sáenz-Mata, J. Halophilic rhizobacteria from Distichlis spicata promote growth and improve salt tolerance in heterologous plant hosts. Symbiosis 2017, 73, 179–189. [Google Scholar] [CrossRef]
- Amaresan, N.; Kumar, K.; Madhuri, K.; Usharani, G.K. Isolation and Characterization of Salt Tolerant Plant Growth Promoting Rhizobacteria from Plants Grown in Tsunami Affected Regions of Andaman and Nicobar Islands. Geomicrobiol. J. 2016, 33, 942–947. [Google Scholar] [CrossRef]
- Allaoua, S.; Hafsa, C.S.; Bilal, Y. Growing varieties durum wheat (Triticum durum) in response to the effect of osmolytes and inoculation by Azotobacter chroococcum under salt stress. Afr. J. Microbiol. Res. 2016, 10, 387–399. [Google Scholar] [CrossRef]
- Lastochkina, O.; Pusenkova, L.; Yuldashev, R.; Babaev, M.; Garipova, S.; Blagova, D.; Khairullin, R.; Aliniaeifard, S. Effects of Bacillus subtilis on some physiological and biochemical parameters of Triticum aestivum L. (wheat) under salinity. Plant Physiol. Biochem. 2017, 121, 80–88. [Google Scholar] [CrossRef]
- Singh, R.P.; Jha, P.N. A Halotolerant Bacterium Bacillus licheniformis HSW-16 Augments Induced Systemic Tolerance to Salt Stress in Wheat Plant (Triticum aestivum). Front. Plant Sci. 2016, 7, 1890. [Google Scholar] [CrossRef]
- Singh, R.P.; Runthala, A.; Khan, S.; Jha, P.N. Quantitative proteomics analysis reveals the tolerance of wheat to salt stress in response to Enterobacter cloacae SBP-8. PLoS ONE 2017, 12, e0183513. [Google Scholar] [CrossRef]
- Safari, D.; Jamali, F.; Nooryazdan, H.; Bayat, F. Evaluation of ACC deaminase producing Pseudomonas fluorescens strains for their effects on seed germination and early growth of wheat under salt stress. Aust. J. Crop Sci. 2018, 12, 413–421. [Google Scholar] [CrossRef]
- Vimal, S.R.; Gupta, J.; Singh, J.S. Effect of salt tolerant Bacillus sp. and Pseudomonas sp. on wheat (Triticum aestivum L.) growth under soil salinity: A comparative study. Microbiol. Res. 2018, 9, 26–32. [Google Scholar] [CrossRef]
- Jha, Y.; Subramanian, R.B. Regulation of Plant Physiology and Antioxidant Enzymes for Alleviating Salinity Stress by Potassium-Mobilizing Bacteria. In Potassium Solubilizing Microorganisms for Sustainable Agriculture; Springer: New Delhi, India, 2016; pp. 149–162. [Google Scholar] [CrossRef]
- Shahzad, R.; Khan, A.L.; Bilal, S.; Waqas, M.; Kang, S.-M.; Lee, I.-J. Inoculation of abscisic acid-producing endophytic bacteria enhances salinity stress tolerance in Oryza sativa. Environ. Exp. Bot. 2017, 136, 68–77. [Google Scholar] [CrossRef]
- Kumar, K.; Amaresan, N.; Madhuri, K. Alleviation of the adverse effect of salinity stress by inoculation of plant growth promoting rhizobacteria isolated from hot humid tropical climate. Ecol. Eng. 2017, 102, 361–366. [Google Scholar] [CrossRef]
- Vimal, S.R.; Patel, V.K.; Singh, J.S. Plant growth promoting Curtobacterium albidum strain SRV4: An agriculturally important microbe to alleviate salinity stress in paddy plants. Ecol. Indic. 2019, 105, 553–562. [Google Scholar] [CrossRef]
- Sarkar, A.; Ghosh, P.K.; Pramanik, K.; Mitra, S.; Soren, T.; Pandey, S.; Maiti, T.K. A halotolerant Enterobacter sp. displaying ACC deaminase activity promotes rice seedling growth under salt stress. Res. Microbiol. 2018, 169, 20–32. [Google Scholar] [CrossRef]
- Sagar, A.; Sayyed, R.Z.; Ramteke, P.W.; Sharma, S.; Marraiki, N.; Elgorban, A.M.; Syed, A. ACC deaminase and antioxidant enzymes producing halophilic Enterobacter sp. PR14 promotes the growth of rice and millets under salinity stress. Physiol. Mol. Biol. Plants 2020, 26, 1847–1854. [Google Scholar] [CrossRef]
- Fazal, A.; Bano, A. Role of Plant Growth-Promoting Rhizobacteria (PGPR), Biochar, and Chemical Fertilizer under Salinity Stress. Commun. Soil Sci. Plant Anal. 2016, 47, 1985–1993. [Google Scholar] [CrossRef]
- Sagar, A.; Dhusiya, K.; Shukla, P.K.; Ramteke, P.W. Salt tolerance plant Growth promoting bacterium Enterobacter cloacae (KP226569) in sustainable maize production under salt stress. In Proceedings of the International Conference on Advancing Frontiersin Biotechnology for Sustainable Agriculture and Health (AFBSAH-2016), Allahabad, India, 25–26 February 2016. [Google Scholar]
- Sagar, A.; Shukla, P.K.; Ramteke, P.W. Salt tolerance plant growth promoting bacterium Azotobacter nigricans (KP966496) in sustainable maize Production under salt stress. In Proceedings of the XXVItalo–Latin American Congress of Ethnomedicine—SILAE, Modena, Italy, 11–15 September 2016. [Google Scholar]
- Ferreira, N.C.; Mazzuchelli, R.D.C.L.; Pacheco, A.C.; De Araujo, F.F.; Antunes, J.E.L.; de Araujo, A.S.F. Bacillus subtilis improves maize tolerance to salinity. Ciência Rural 2018, 48. [Google Scholar] [CrossRef]
- Fukami, J.; De La Osa, C.; Ollero, F.J.; Megías, M.; Hungria, M. Co-inoculation of maize with Azospirillum brasilense and Rhizobium tropici as a strategy to mitigate salinity stress. Funct. Plant Biol. 2018, 45, 328–339. [Google Scholar] [CrossRef]
- Latef, A.; Abu, A.A.H.; Alhmad, M.F.; Kordrostami, M.; Abo–Baker, A.-B.A.-E.; Zakir, A. Inoculation with Azospirillum lipoferum or Azotobacter chroococcum Reinforces Maize Growth by Improving Physiological Activities Under Saline Conditions. J. Plant Growth Regul. 2020, 39, 1293–1306. [Google Scholar] [CrossRef]
- Sagar, A.; Kuddus, M.; Singh, B.P.; Labhane, N.M.; Srivastava, S.P.W. Plant Growth Promotion of millets under Abiotic stress using Enterobacter cloacae PR10 (KP226575b). J. Indian Bot. Soc. 2020, 100, 30–41. [Google Scholar] [CrossRef]
- Omer, A.M.; Emara, H.M.; Zaghloul, R.A.; Abdel-Monem, M.O.; Dawwam, G.E. Potential of Azotobacter Salinestris as Plant Growth Promoting Rhizobacteria under Saline Stress Conditions. RJPBCS Res. J. Pharm. Biol. Chem. Sci. 2016, 7, 2572–2583. [Google Scholar]
- Bhise, K.K.; Bhagwat, P.K.; Dandge, P.B. Plant Growth-Promoting Characteristics of Salt Tolerant Enterobacter cloacae Strain KBPD and Its Efficacy in Amelioration of Salt Stress in Vigna radiata L. J. Plant Growth Regul. 2016, 36, 215–226. [Google Scholar] [CrossRef]
- Viscardi, S.; Ventorino, V.; Duran, P.; Maggio, A.; De Pascale, S.; Mora, M.; Pepe, O. Assessment of plant growth promoting activities and abiotic stress tolerance of Azotobacter chroococcum strains for a potential use in sustainable agriculture. J. Soil Sci. Plant Nutr. 2016, 16, 848–863. [Google Scholar] [CrossRef]
- Van Oosten, M.J.; Di Stasio, E.; Cirillo, V.; Silletti, S.; Ventorino, V.; Pepe, O.; Maggio, A. Root inoculation with Azotobacter chroococcum 76A enhances tomato plants adaptation to salt stress under low N conditions. BMC Plant Biol. 2018, 18, 205. [Google Scholar] [CrossRef] [PubMed]
- Win, K.T.; Tanaka, F.; Okazaki, K.; Ohwaki, Y. The ACC deaminase expressing endophyte Pseudomonas spp. Enhances NaCl stress tolerance by reducing stress-related ethylene production, resulting in improved growth, photosynthetic performance, and ionic balance in tomato plants. Plant Physiol. Biochem. 2018, 127, 599–607. [Google Scholar] [CrossRef]
- Gupta, A.; Rai, S.; Bano, A.; Khanam, A.; Sharma, S.; Pathak, N. Comparative Evaluation of Different Salt-Tolerant Plant Growth-Promoting Bacterial Isolates in Mitigating the Induced Adverse Effect of Salinity in Pisum sativum. Biointerface Res. Appl. Chem. 2021, 11, 13141–13154. [Google Scholar]
- Sapre, S.; Gontia-Mishra, I.; Tiwari, S. Plant Growth-Promoting Rhizobacteria Ameliorates Salinity Stress in Pea (Pisum sativum). J. Plant Growth Regul. 2021, 189, 1–10. [Google Scholar] [CrossRef]
- Abd_Allah, E.F.; Alqarawi, A.A.; Hashem, A.; Radhakrishnan, R.; Al-Huqail, A.A.; Al-Otibi, F.O.N.; Malik, J.A.; Alharbi, R.I.; Egamberdieva, D. Endophytic bacterium Bacillus subtilis (BERA 71) improves salt tolerance in chickpea plants by regulating the plant defense mechanisms. J. Plant Interact. 2018, 13, 37–44. [Google Scholar] [CrossRef]
- Mushtaq, Z.; Faizan, S.; Gulzar, B.; Hakeem, K.R. Inoculation of Rhizobium Alleviates Salinity Stress Through Modulation of Growth Characteristics, Physiological and Biochemical Attributes, Stomatal Activities and Antioxidant Defence in Cicer arietinum L. J. Plant Growth Regul. 2021, 40, 2148–2163. [Google Scholar] [CrossRef]
- Wang, W.; Wu, Z.; He, Y.; Huang, Y.; Li, X.; Ye, B.-C. Plant growth promotion and alleviation of salinity stress in Capsicum annuum L. by Bacillus isolated from saline soil in Xinjiang. Ecotoxicol. Environ. Saf. 2018, 164, 520–529. [Google Scholar] [CrossRef] [PubMed]
- Samaddar, S.; Chatterjee, P.; Choudhury, A.R.; Ahmed, S.; Sa, T. Interactions between Pseudomonas spp. and their role in improving the red pepper plant growth under salinity stress. Microbiol. Res. 2018, 219, 66–73. [Google Scholar] [CrossRef]
- Habib, S.H.; Kausar, H.; Saud, H.M. Plant Growth-Promoting Rhizobacteria Enhance Salinity Stress Tolerance in Okra through ROS-Scavenging Enzymes. BioMed Res. Int. 2016, 2016, 6284547. [Google Scholar] [CrossRef]
- Szymańska, S.; Dąbrowska Grażyna, B.; Jarosław, T.; Katarzyna, N.; Agnieszka, P.; Katarzyna, H. Boosting the Brassica napus L. tolerance to salinity by the halotolerant strain Pseudomonas stutzeri ISE12. Environ. Exp. Bot. 2019, 163, 55–68. [Google Scholar] [CrossRef]
- Yasmin, H.; Naeem, S.; Bakhtawar, M.; Jabeen, Z.; Nosheen, A.; Naz, R.; Keyani, R.; Mumtaz, S.; Hassan, M.N. Halotolerant rhizobacteria Pseudomonas pseudoalcaligenes and Bacillus subtilis mediate systemic tolerance in hydroponically grown soybean (Glycine max L.) against salinity stress. PLoS ONE 2020, 15, e0231348. [Google Scholar] [CrossRef]
- Khan, M.A.; Sahile, A.A.; Jan, R.; Asaf, S.; Hamayun, M.; Imran, M.; Adhikari, A.; Kang, S.; Kim, K.M.; Lee, I. Halo-tolerant bacteria mitigate the effects of salinity stress on soybean growth by regulating secondary metabolites and molecu-lar responses. BMC Plant Biol. 2021, 21, 176. [Google Scholar] [CrossRef]
- Li, H.; Lei, P.; Pang, X.; Li, S.; Xu, H.; Xu, Z.; Feng, X. Enhanced tolerance to salt stress in canola (Brassica napus L.) seedlings inoculated with the halotolerant Enterobacter cloacae HSNJ4. Appl. Soil Ecol. 2017, 119, 26–34. [Google Scholar] [CrossRef]
- Saghafi, D.; Ghorbanpour, M.; Lajayer, B.A. Efficiency of Rhizobium strains as plant growth promoting rhizobacteria on morpho-physiological properties of Brassica napus L. under salinity stress. J. Soil Sci. Plant Nutr. 2018, 18, 253–268. [Google Scholar] [CrossRef]
- Benidire, L.; Lahrouni, M.; Khalloufi, F.; Gottfert, M.; Oufdou, K. Effects of Rhizobium leguminosarum inoculation on growth, ni-trogen uptake and mineral assimilation in vicia faba plants under salinity stress. J. Agric. Sci. Technol. (JAST) 2017, 19, 889–901. [Google Scholar]
- Garcia, L.; Dattamudi, S.; Chanda, S.; Jayachandran, K. Effect of Salinity Stress and Microbial Inoculations on Glomalin Production and Plant Growth Parameters of Snap Bean (Phaseolus vulgaris). Agronomy 2019, 9, 545. [Google Scholar] [CrossRef]
- Garg, N.; Pandey, R. High effectiveness of exotic arbuscular mycorrhizal fungi is reflected in improved rhizobial symbiosis and trehalose turnover in Cajanus cajan genotypes grown under salinity stress. Fungal Ecol. 2016, 21, 57–67. [Google Scholar] [CrossRef]
- Yousefi, S.; Kartoolinejad, D.; Bahmani, M.; Naghdi, R. Effect ofAzospirillum lipoferumandAzotobacter chroococcumon germination and early growth of hopbush shrub (Dodonaea viscosa L.) under salinity stress. J. Sustain. For. 2016, 36, 107–120. [Google Scholar] [CrossRef]
- Arora, M.; Saxena, P.; Abdin, M.Z.; Varma, A. Interaction between Piriformospora indica and Azotobacter chroococcum diminish the effect of salt stress in Artemisia annua L. by enhancing enzymatic and non-enzymatic antioxidants. Symbiosis 2019, 80, 61–73. [Google Scholar] [CrossRef]
- Silambarasan, S.; Logeswari, P.; Valentine, A.; Cornejo, P.; Kannan, V.R. Pseudomonas citronellolis strain SLP6 enhances the phytoremediation efficiency of Helianthus annuus in copper contaminated soils under salinity stress. Plant Soil 2020, 457, 241–253. [Google Scholar] [CrossRef]
- Srivastava, S.; Srivastava, S. Prescience of endogenous regulation in Arabidopsis thaliana by Pseudomonas putida MTCC 5279 under phosphate starved salinity stress condition. Sci. Rep. 2020, 10, 5855. [Google Scholar] [CrossRef]
- Noori, F.; Etesami, H.; Najafi Zarini, H.; Khoshkholgh-Sima, N.A.; Hosseini Salekdeh, G.; Alishahi, F. Mining alfalfa (Medicago sativa L.) nodules for salinity tolerant non-rhizobial bacteria to improve growth of alfalfa under salinity stress. Ecotoxicol. Environ. Saf. 2018, 162, 129–138. [Google Scholar] [CrossRef]
- Zhu, Z.; Zhang, H.; Leng, J.; Niu, H.; Chen, X.; Liu, D.; Chen, Y.; Gao, N.; Ying, H. Isolation and characterization of plant growth-promoting rhizobacteria and their effects on the growth of Medicago sativa L. under salinity conditions. Antonie Leeuwenhoek 2020, 113, 1263–1278. [Google Scholar] [CrossRef]
- Tirry, N.; Laghmari, A.K.G.; Lemjereb, M.; Hnadi, H.; Amrani, K.; Bahafid, W.; Ghachtouli, N.E. Improved salinity tolerance of Medicago sativa and soil enzyme activities by PGPR. Biocatal. Agric. Biotechnol. 2021, 31, 101914. [Google Scholar] [CrossRef]
- Rahman, A.; Sultana, V.; Ara, J.; Ehteshamul-Haque, S. Induction of Systemic Resistance in Cotton by the Neem Cake and Pseudomonas aeruginosa under Salinity Stress and Macrophomina phaseolina Infection. Pak. J. Bot. 2016, 48, 1681–1689. [Google Scholar]
- Hashem, A.; Abd-Allah, E.F.; Alqarawi, A.; Al-Huqail, A.A.; Shah, M.A. Induction of Osmoregulation and Modulation of Salt Stress in Acacia gerrardii Benth by Arbuscular Mycorrhizal Fungi andBacillus subtilis(BERA 71). BioMed Res. Int. 2016, 2016, 6294098. [Google Scholar] [CrossRef] [PubMed]
- Egamberdieva, D.; Jabborova, D.; Berg, G. Synergistic interactions between Bradyrhizobium japonicum and the endophyte Stenotrophomonas rhizophila and their effects on growth, and nodulation of soybean under salt stress. Plant Soil 2015, 405, 35–45. [Google Scholar] [CrossRef]
- Raghuwanshi, R.; Prasad, J.K. Perspectives of Rhizobacteria with ACC Deaminase Activity in Plant Growth under Abiotic Stress. In Root Biology; Soil Biology; Giri, B., Prasad, R., Varma, A., Eds.; Springer International Publishing: Cham, Switzerland, 2018; Volume 52, pp. 303–321. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A. Exopolysaccharide producing rhizobacteria and their impact on growth and drought tolerance of wheat grown under rainfed conditions. PLoS ONE 2019, 14, e0222302. [Google Scholar] [CrossRef]
- Shameer, S.; Prasad, T. Plant growth promoting rhizobacteria for sustainable agricultural practices with special reference to biotic and abiotic stresses. Plant Growth Regul. 2018, 84, 603–615. [Google Scholar] [CrossRef]
- Sayyed, R.Z.; Seifi, S.; Patel, P.R.; Shaikh, S.S.; Jadhav, H.P.; El Enshasy, H. Siderophore production in groundnut rhizosphere isolate, Achromobacter sp. RZS2 influenced by physicochemical factors and metal ions. Environ. Sustain. 2019, 2, 117–124. [Google Scholar] [CrossRef]
- Etesami, H.; Emami, S.; Alikhani, H.A. Potassium solubilizing bacteria (KSB): Mechanisms, promotion of plant growth, and future prospects—A review. J. Soil Sci. Plant Nutr. 2017, 17, 897–911. [Google Scholar] [CrossRef]
- Zaidi, A.; Khan, M.S.; Saif, S.; Rizvi, A.; Ahmed, B.; Shahid, M. Role of Nitrogen-Fixing Plant Growth-Promoting Rhizobacteria in Sustainable Production of Vegetables: Current Perspective. In Microbial Strategies for Vegetable Production; Springer: Berlin/Heidelberg, Germany, 2017; pp. 49–79. [Google Scholar]
- Tang, A.; Haruna, A.O.; Majid, N.M.A.; Jalloh, M.B. Potential PGPR Properties of Cellulolytic, Nitrogen-Fixing, Phosphate-Solubilizing Bacteria in Rehabilitated Tropical Forest Soil. Microorganisms 2020, 8, 442. [Google Scholar] [CrossRef]
- Abbas, R.; Rasul, S.; Aslam, K.; Baber, M.; Shahid, M.; Mubeen, F.; Naqqash, T. Halotolerant PGPR: A hope for cultivation of saline soils. J. King Saud Univ.-Sci. 2019, 31, 1195–1201. [Google Scholar] [CrossRef]
- Tsukanova, K.A.; Сhеbotаr, V.К.; Meyer, J.J.M.; Bibikova, T.N. Effect of plant growth-promoting Rhizobacteria on plant hormone homeostasis. S. Afr. J. Bot. 2017, 113, 91–102. [Google Scholar] [CrossRef]
- Grover, M.; Bodhankar, S.; Sharma, A.; Sharma, P.; Singh, J.; Nain, L. PGPR Mediated Alterations in Root Traits: Way toward Sustainable Crop Production. Front. Sustain. Food Syst. 2021, 4, 287. [Google Scholar] [CrossRef]
- Patel, T.; Saraf, M. Biosynthesis of phytohormones from novel rhizobacterial isolates and their in vitro plant growth-promoting efficacy. J. Plant Interact. 2017, 12, 480–487. [Google Scholar] [CrossRef]
- Tewari, S.; Arora, N.K. Fluorescent Pseudomonas sp. PF17 as an efficient plant growth regulator and biocontrol agent for sunflower crop under saline conditions. Symbiosis 2016, 68, 99–108. [Google Scholar] [CrossRef]
- Li, M.; Guo, R.; Yu, F.; Chen, X.; Zhao, H.; Li, H.; Wu, J. Indole-3-Acetic Acid Biosynthesis Pathways in the Plant-Beneficial Bacterium Arthrobacter pascens ZZ21. Int. J. Mol. Sci. 2018, 19, 443. [Google Scholar] [CrossRef]
- Joshi, G.; Kumar, V.; Brahmachari, S.K. Screening and identification of novel halotolerant bacterial strains and assessment for insoluble phosphate solubilization and IAA production. Bull. Natl. Res. Cent. 2021, 45, 83. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Jabborova, D.; Wirth, S.; Alam, P.; Alyemeni, M.N.; Ahmad, P. Interactive Effects of Nutrients and Bradyrhizobium japonicum on the Growth and Root Architecture of Soybean (Glycine max L.). Front. Microbiol. 2018, 9, 1000. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Wirth, S.; Jabborova, D.; Räsänen, L.A.; Liao, H. Coordination between Bradyrhizobium and Pseudomonas alleviates salt stress in soybean through altering root system architecture. J. Plant Interact. 2017, 12, 100–107. [Google Scholar] [CrossRef]
- Kang, S.-M.; Shahzad, R.; Bilal, S.; Khan, A.L.; Park, Y.-G.; Lee, K.-E.; Asaf, S.; Khan, M.A.; Lee, I.-J. Indole-3-acetic-acid and ACC deaminase producing Leclercia adecarboxylata MO1 improves Solanum lycopersicum L. growth and salinity stress tolerance by endogenous secondary metabolites regulation. BMC Microbiol. 2019, 19, 80. [Google Scholar] [CrossRef]
- Panwar, M.; Tewari, R.; Gulati, A.; Nayyar, H. Indigenous salt-tolerant rhizobacterium Pantoea dispersa (PSB3) reduces sodium uptake and mitigates the effects of salt stress on growth and yield of chickpea. Acta Physiol. Plant. 2016, 38, 278. [Google Scholar] [CrossRef]
- Shultana, R.; Tan Kee Zuan, A.; Yusop, M.R.; Mohd Saud, H.; Ayanda, A.F. Effect of Salt-Tolerant Bacterial Inoculations on Rice Seedlings Differing in Salt-Tolerance under Saline Soil Conditions. Agronomy 2020, 10, 1030. [Google Scholar] [CrossRef]
- Banerjee, A.; Bareh, D.A.; Joshi, S.R. Native microorganisms as potent bio inoculants for plant growth promotion in shifting agriculture (Jhum) systems. J. Soil Sci. Plant Nutr. 2017, 17, 127–140. [Google Scholar]
- Cortleven, A.; Schmülling, T. Regulation of chloroplast development and function by cytokinin. J. Exp. Bot. 2015, 66, 4999–5013. [Google Scholar] [CrossRef]
- TrParray, A.P.; Jan, S.; Kamili, A.N.; Qadri, R.A.; Egamberdieva, D.; Ahmad, P. Current perspectives on plant growth promot-ing rhizobacteria. Plant Growth Regul. 2016, 35, 877–902. [Google Scholar]
- Tahir, H.A.S.; Gu, Q.; Wu, H.; Raza, W.; Hanif, A.; Wu, L.; Colman, M.V.; Gao, X. Plant growth promotion by volatile organ-ic compounds produced by Bacillus subtilis SYST2. Front. Microbiol. 2017, 8, 171. [Google Scholar] [CrossRef]
- Ijaz, M.; Tahir, M.; Shahid, M.; Ul-Allah, S.; Sattar, A.; Sher, A.; Mahmood, K.; Hussain, M. Combined application of biochar and PGPR consortia for sustainable production of wheat under semiarid conditions with a reduced dose of synthetic fertilizer. Braz. J. Microbiol. Publ. Braz. Soc. Microbiol. 2019, 50, 449–458. [Google Scholar] [CrossRef]
- Mukhtar, T.; Rehman, S.U.; Smith, D.; Sultan, T.; Seleiman, M.F.; Alsadon, A.A.; Amna; Ali, S.; Chaudhary, H.J.; Solieman, T.H.I.; et al. Mitigation of Heat Stress in Solanum lycopersicum L. by ACC-deaminase and Exopolysaccharide Producing Bacillus cereus: Effects on Biochemical Profiling. Sustainability 2020, 12, 2159. [Google Scholar] [CrossRef]
- Akhtar, S.S.; Mekureyaw, M.F.; Pandey, C.; Roitsch, T. Role of Cytokinins for Interactions of Plants with Microbial Pathogens and Pest Insects. Front. Plant Sci. 2020, 10, 1777. [Google Scholar] [CrossRef]
- Vaishnav, A.; Singh, J.; Singh, P.; Rajput, R.S.; Singh, H.B.; Sarma, B.K. Sphingo bacterium sp. BHU-AV3 Induces Salt Tolerance in Tomato by Enhancing Antioxidant Activities and Energy Metabolism. Front. Microbiol. 2020, 11, 443. [Google Scholar] [CrossRef]
- Supriya, P.; Kusale, S.; Attar, Y.; Sayyed, R.; Malek, R.; Ilyas, N.; Suriani, N.; Khan, N.; El Enshasy, H. Production of Plant Beneficial and Antioxidants Metabolites by Klebsiellavariicola under Salinity Stress. Molecules 2021, 26, 1894. [Google Scholar] [CrossRef]
- Arora, D.; Bhatla, S.C. Melatonin and nitric oxide regulate sunflower seedling growth under salt stress accompanying differential expression of Cu/Zn SOD and Mn SOD. Free Radic. Biol. Med. 2017, 106, 315–328. [Google Scholar] [CrossRef]
- Sukweenadhi, J.; Balusamy, S.R.; Kim, Y.J.; Lee, C.H.; Kim, Y.J.; Koh, S.C.; Yang, D.C. A growth-promoting bacteria, Paenibacillus yonginensis DCY84T enhanced salt stress tolerance by activating defense-related systems in Panax ginseng. Front. Plant Sci. 2018, 9, 813. [Google Scholar] [CrossRef] [PubMed]
- Ansari, F.A.; Ahmad, I.; Pichtel, J. Growth stimulation and alleviation of salinity stress to wheat by the biofilm forming Bacillus pumilus strain FAB10. Appl. Soil Ecol. 2019, 143, 45–54. [Google Scholar] [CrossRef]
- Khan, M.A.; Asaf, S.; Khan, A.L.; Adhikari, A.; Jan, R.; Ali, S.; Imran, M.; Kim, K.-M.; Lee, I.-J. Halotolerant Rhizobacterial Strains Mitigate the Adverse Effects of NaCl Stress in Soybean Seedlings. BioMed Res. Int. 2019, 2019, 9530963. [Google Scholar] [CrossRef] [PubMed]
- Khanna, K.; Jamwal, V.L.; Kohli, S.K.; Gandhi, S.G.; Ohri, P.; Bhardwaj, R.; Abd-Allah, E.; Hashem, A.; Ahmad, P. Plant growth promoting rhizobacteria induced Cd tolerance in Lycopersicon esculentum through altered antioxidative defense expression. Chemosphere 2018, 217, 463–474. [Google Scholar] [CrossRef]
- Rajput, R.S.; Ram, R.M.; Vaishnav, A.; Singh, H.B. Microbe-based novel biostimulants for sustainable crop production. In Microbial Diversity in Ecosystem Sustainability and Biotechnological Applications; Satynarayan, T., Ed.; Springer: Singapore, 2019; pp. 109–144. [Google Scholar] [CrossRef]
- El-Esawi, M.A.; Al-Ghamdi, A.A.; Ali, H.M.; Alayafi, A.A. Azospirillum lipoferum FK1 confers improved salt tolerance in chickpea (Cicer arietinum L.) by modulating osmolytes, antioxidant machinery and stress-related genes expression. Environ. Exp. Bot. 2018, 159, 55–65. [Google Scholar] [CrossRef]
- Chen, D.; Shao, Q.; Yin, L.; Younis, A.; Zheng, B. Polyamine Function in Plants: Metabolism, Regulation on Development, and Roles in Abiotic Stress Responses. Front. Plant Sci. 2019, 9, 1945. [Google Scholar] [CrossRef]
- Ma, Y.; Dias, M.C.; Freitas, H. Drought and Salinity Stress Responses and Microbe-Induced Tolerance in Plants. Front. Plant Sci. 2020, 11, 1750. [Google Scholar] [CrossRef]
- Cappellari, L.; Banchio, E. Microbial Volatile Organic Compounds Produced by Bacillus amyloliquefaciens GB Ameliorate the Effects of Salt Stress in Mentha piperita Principally Through Acetoin Emission. J. Plant Growth Regul. 2019, 39, 764–775. [Google Scholar] [CrossRef]
- Abdelaal, K.A.; El-Maghraby, L.M.; Elansary, H.; Hafez, Y.M.; Ibrahim, E.; El-Banna, M.; Elkelish, A. Treatment of Sweet Pepper with Stress Tolerance-Inducing Compounds Alleviates Salinity Stress Oxidative Damage by Mediating the Physio-Biochemical Activities and Antioxidant Systems. Agronomy 2019, 10, 26. [Google Scholar] [CrossRef]
- Zahir, Z.A.; Nadeem, S.M.; Khan, M.Y.; Binyamin, R.; Waqas, M.R. Role of Halotolerant Microbes in Plant Growth Promotion Under Salt Stress Conditions. In Saline Soil-Based Agriculture by Halotolerant Microorganisms; Kumar, M., Ed.; Springer: Singapore, 2019; pp. 209–253. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Kumar, V.; Kohli, S.K.; Sidhu, G.P.S.; Bali, A.S.; Zheng, B. Phytohormones Regulate Accumulation of Osmolytes under Abiotic Stress. Biomolecules 2019, 9, 285. [Google Scholar] [CrossRef]
- Mishra, J.; Fatima, T.; Arora, N.K. Role of Secondary Metabolites from Plant Growth-Promoting Rhizobacteria in Combating Salinity Stress. In Plant Microbiome: Stress Response; Ahmad, P., Egamberdieva, D., Eds.; Springer: Singapore, 2018; pp. 127–163. [Google Scholar]
- Bremer, E.; Krämer, R. Responses of Microorganisms to Osmotic Stress. Annu. Rev. Microbiol. 2019, 73, 313–334. [Google Scholar] [CrossRef]
- Kushwaha, B.; Jadhav, I.; Verma, H.N.; Geethadevi, A.; Parashar, D.; Jadhav, K. Betaine accumulation suppresses the de-novo synthesis of ectoine at a low osmotic concentration in Halomonas sp SBS 10, a bacterium with broad salinity tolerance. Mol. Biol. Rep. 2019, 46, 4779–4786. [Google Scholar] [CrossRef]
- Qin, S.; Feng, W.-W.; Zhang, Y.-J.; Wang, T.-T.; Xiong, Y.-W.; Xing, K. Diversity of Bacterial Microbiota of Coastal Halophyte Limonium sinense and Amelioration of Salinity Stress Damage by Symbiotic Plant Growth-Promoting Actinobacterium Glutamicibacter halophytocola KLBMP 5180. Appl. Environ. Microbiol. 2018, 84, e01533-18. [Google Scholar] [CrossRef]
- Forni, C.; Duca, D.; Glick, B.R. Mechanisms of plant response to salt and drought stress and their alteration by rhizobacteria. Plant Soil 2017, 410, 335–356. [Google Scholar] [CrossRef]
- Shim, J.S.; Seo, J.-S.; Seo, J.S.; Kim, Y.; Koo, Y.; Choi, Y.D.; Jung, C. Heterologous expression of bacterial trehalose biosynthetic genes enhances trehalose accumulation in potato plants without adverse growth effects. Plant Biotechnol. Rep. 2019, 13, 409–418. [Google Scholar] [CrossRef]
- Atouei, M.T.; Pourbabaee, A.A.; Shorafa, M. Alleviation of Salinity Stress on Some Growth Parameters of Wheat by Exopolysaccharide-Producing Bacteria. Iran. J. Sci. Technol. Trans. A Sci. 2019, 43, 2725–2733. [Google Scholar] [CrossRef]
- Chu, T.N.; Tran, B.T.H.; Van Bui, L.; Hoang, M.T.T. Plant growth-promoting rhizobacterium Pseudomonas PS01 induces salt tolerance in Arabidopsis thaliana. BMC Res. Notes 2019, 12, 11. [Google Scholar] [CrossRef]
- Ali, S.; Kim, W.-C. Plant Growth Promotion under Water: Decrease of Waterlogging-Induced ACC and Ethylene Levels by ACC Deaminase-Producing Bacteria. Front. Microbiol. 2018, 9, 1096. [Google Scholar] [CrossRef]
- Saleem, A.R.; Brunetti, C.; Khalid, A.; Della Rocca, G.; Raio, A.; Emiliani, G.; DE Carlo, A.; Mahmood, T.; Centritto, M. Drought response of Mucuna pruriens (L.) DC. inoculated with ACC deaminase and IAA producing rhizobacteria. PLoS ONE 2018, 13, e0191218. [Google Scholar] [CrossRef]
- Del Carmen Orozco-Mosqueda, M.; Glick, B.R.; Santoyo, G. ACC deaminase in plant growth-promoting bacteria (PGPB): An efficient mechanism to counter salt stress in crops. Microbiol. Res. 2020, 235, 126439. [Google Scholar] [CrossRef]
- Acuña, J.J.; Camposa, M.; Morab, M.D.L.; Jaisic, D.P.; Jorquera, M.A. ACCD-producing rhizobacteria from an Andean Altiplano native plant (Parastrephia quadrangularis) to alleviate salt stress in wheat seedlings. Appl. Soil Ecol. 2019, 36, 184–190. [Google Scholar] [CrossRef]
- Gupta, S.; Pandey, S. ACC Deaminase Producing Bacteria with Multifarious Plant Growth Promoting Traits Alleviates Salinity Stress in French Bean (Phaseolus vulgaris) Plants. Front. Microbiol. 2019, 10, 1506. [Google Scholar] [CrossRef]
- Maxton, A.; Singh, P.; Masih, S.A. ACC deaminase-producing bacteria mediated drought and salt tolerance in Capsicum annuum. J. Plant Nutr. 2017, 41, 574–583. [Google Scholar] [CrossRef]
- Yoo, S.; Weon, H.Y.; Song, J.; Sang, M.K. Induced Tolerance to Salinity Stress by Halotolerant Bacteria Bacillus ary-abhattai H19-1 and B. mesonae H20-5 in Tomato Plants. J. Microbiol. Biotechnol. 2019, 29, 1124–1136. [Google Scholar] [CrossRef]
- Santoyo, G.; Moreno-Hagelsieb, G.; del Carmen Orozco-Mosqueda, M.; Glick, B.R. Plant growth-promoting bacterial endophytes. Microbiol. Res. 2016, 183, 92–99. [Google Scholar] [CrossRef]
- Shah, G.; Jan, M.; Afreen, M.; Anees, M.; Rehman, S.; Daud, M.; Malook, I.; Jamil, M. Halophilic bacteria mediated phytoremediation of salt-affected soils cultivated with rice. J. Geochem. Explor. 2017, 174, 59–65. [Google Scholar] [CrossRef]
- Aw, X.; Li, Z.; Wc, L.; Zh, Y. The effect of plant growth-promoting rhizobacteria (PGPR) on arsenic accumulation and the growth of rice plants (Oryza sativa L.). Chemosphere 2019, 242, 125136. [Google Scholar] [CrossRef]
- Habibi, S.; Djedidi, S.; Ohkama-Ohtsu, N.; Sarhadi, W.A.; Kojima, K.; Rallos, R.V.; Ramirez, M.D.A.; Yamaya, H.; Sekimoto, H.; Yokoyama, T. Isolation and Screening of Indigenous Plant Growth-promoting Rhizobacteria from Different Rice Cultivars in Afghanistan Soils. Microbes Environ. 2019, 34, 347–355. [Google Scholar] [CrossRef]
- Sagar, A.; Shukla, P.K.; Ramteke, P.W. Stimulation of Seed Germination and Growth Parameters of Rice var. Sahbhagi by Enterobacter cloacae (KP226569) in Presence of Ammonia Sulphate as Substitute of 1-Aminocyclopropane-1-Carboxylate. In Plant Growth Promoting Rhizobacteria (PGPR): Prospects in Sustainable Agriculture; Springer-Nature: Singapore, 2019; pp. 117–124. [Google Scholar]
- Javed, H.; Riaz, A.; Qureshi, A.; Javed, K.; Mujeeb, F.; Ijaz, F.; Akhtar, M.S.; Ali, M.A.; Gul, R.; Aftab, M. Isolation, characterization and screening of PGPR capable of providing relief in salinity stress. Eurasian J. Soil Sci. 2020, 9, 85–91. [Google Scholar] [CrossRef][Green Version]
- Fatima, T.; Mishra, I.; Verma, R.; Arora, N.K. Mechanisms of halotolerant plant growth promoting Alcaligenes sp. involved in salt tolerance and enhancement of the growth of rice under salinity stress. 3 Biotech 2020, 10, 361. [Google Scholar] [CrossRef]
- Prittesh, P.; Avnika, P.; Kinjal, P.; Jinal, H.N.; Sakthivel, K.; Amaresan, N. Amelioration effect of salt-tolerant plant growth-promoting bacteria on growth and physiological properties of rice (Oryza sativa) under salt-stressed conditions. Arch. Microbiol. 2020, 202, 2419–2428. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.; Yuan, D.; Jin, C.; Wang, G.; Li, X.; Guan, C. Enhancement of growth and salt tolerance of rice seedlings (Oryza sativa L.) by regulating ethylene production with a novel halotolerant PGPR strain Glutamicibacter sp. YD01 containing ACC deaminase activity. Acta Physiol. Plant. 2020, 42, 42. [Google Scholar] [CrossRef]
- Sultana, S.; Paul, S.C.; Parveen, S.; Alam, S.; Rahman, S.; Bushra Jannat, B.; Hoque, S.; Rahman, M.T.; Karim, M.M. Iso-lation and identification of salt-tolerant plant-growth-promoting rhizobacteria and their application for rice cultivation. Can. J. Microbiol. 2020, 66, 144–160. [Google Scholar]
- Nawaz, A.; Shahbaz, M.; Imran, A.; Marghoob, M.U.; Imtiaz, M.; Mubeen, F. Potential of salt tolerant PGPR in growth and yield augmentation of wheat (Triticum aestivum L.) under saline conditions. Front. Microbiol. 2020, 11, 2019. [Google Scholar] [CrossRef]
- Singh, R.P.; Jha, P.N. Analysis of fatty acid composition of PGPR Klebsiella sp. SBP-8 and its role in ameliorating salt stress in wheat. Symbiosis 2017, 73, 213–222. [Google Scholar] [CrossRef]
- Nguyen, M.L.; Spaepen, S.; Du Jardin, P.; Delaplace, P. Biostimulant effects of rhizobacteria on wheat growth and nutrient uptake depend on nitrogen application and plant development. Arch. Agron. Soil Sci. 2018, 65, 58–73. [Google Scholar] [CrossRef]
- Sagar, A.; Thomas, G.; Rai, S.; Mishra, R.K.; Ramteke, P.W. Enhancement of Growth and Yield parameters of Wheat variety AAI- W6 by an organic farm isolate of plant growth promoting Erwinia species KP226572. Int. J. Agric. Environ. Biotechnol. 2018, 11, 159–171. [Google Scholar]
- Akbar, M.; Aslam, N.; Khalil, T.; Akhtar, S.; Siddiqi, E.H.; Iqbal, M.S. Effects of seed priming with plant growth-promoting rhizobacteria on wheat yield and soil properties under contrasting soils. J. Plant Nutr. 2019, 42, 2080–2091. [Google Scholar] [CrossRef]
- Cortivo, C.D.; Ferrari, M.; Visioli, G.; Lauro, M.; Fornasier, F.; Barion, G.; Panozzo, A.; Vamerali, T. Effects of Seed-Applied Biofertilizers on Rhizosphere Biodiversity and Growth of Common Wheat (Triticum aestivum L.) in the Field. Front. Plant Sci. 2020, 11, 72. [Google Scholar] [CrossRef]
- El-Nahrawy, S.; Yassin, M. Response of Different Cultivars of Wheat Plants (Triticum aestivum L.) to Inoculation by Azotobacter sp. under Salinity Stress Conditions. J. Adv. Microbiol. 2020, 20, 59–79. [Google Scholar] [CrossRef]
- Akram, M.S.; Shahid, M.; Tariq, M.; Azeem, M.; Javed, M.T.; Saleem, S.; Riaz, S. Deciphering Staphylococcus sciuri SAT-17 Mediated Anti-oxidative Defense Mechanisms and Growth Modulations in Salt Stressed Maize (Zea mays L.). Front. Microbiol. 2016, 7, 867. [Google Scholar] [CrossRef]
- Peng, J.; Ma, J.; Wei, X.; Zhang, C.; Jia, N.; Wang, X.; Wang, E.T.; Hu, D.; Wang, Z. Accumulation of beneficial bacteria in the rhizosphere of maize (Zea mays L.) grown in a saline soil in responding to a consortium of plant growth promoting rhizobacteria. Ann. Microbiol. 2021, 71, 40. [Google Scholar] [CrossRef]
- Li, H.; Jiang, X. Inoculation with plant growth-promoting bacteria (PGPB) improves salt tolerance of maize seedling. Russ. J. Plant Physiol. 2017, 64, 235–241. [Google Scholar] [CrossRef]
- Rafiq, K.; Akram, M.S.; Shahid, M.; Qaisar, U.; Rashid, N. Enhancement of salt tolerance in maize (Zea mays L.) using locally isolated Bacillus sp. SR-2-1/1. Biologia 2020, 75, 1425–1436. [Google Scholar] [CrossRef]
- El-Esawi, M.A.; Alaraidh, I.A.; Alsahli, A.A.; Alzahrani, S.A.; Ali, H.M.; Alayafi, A.A.; Ahmad, M. Serratia liquefaciens KM4 improves salt stresstolerance in maize by regulating redox potential, ion homeostasis, leaf gas exchange and stress-related gene expression. Int. J. Mol. Sci. 2018, 19, 3310. [Google Scholar] [CrossRef]
- Mukhtar, S.; Zareen, M.; Khaliq, Z.; Mehnaz, S.; Malik, K.A. Phylogenetic analysis of halophyte-associated rhizobacteria and effect of halotolerant and halophilic phosphate-solubilizing biofertilizers on maize growth under salinity stress conditions. J. Appl. Microbiol. 2019, 128, 556–573. [Google Scholar] [CrossRef]
- Misra, S.; Chauhan, P.S. ACC deaminase-producing rhizosphere competent Bacillus spp. mitigate salt stress and promote Zea mays growth by modulating ethylene metabolism. 3 Biotech 2020, 10, 119. [Google Scholar] [CrossRef]
- Dixit, V.K.; Misra, S.; Mishra, S.K.; Tewari, S.K.; Joshi, N.; Chauhan, P.S. Characterization of plant growth-promoting alkalotolerant Alcaligenes and Bacillus strains for mitigating the alkaline stress in Zea mays. Antonie Leeuwenhoek 2020, 113, 889–905. [Google Scholar] [CrossRef]
- Sapre, S.; Gontia-Mishra, I.; Tiwari, S. Klebsiella sp. confers enhanced tolerance to salinity and plant growth promotion in oat seedlings (Avena sativa). Microbiol. Res. 2018, 206, 25–32. [Google Scholar] [CrossRef]
- Chibeba, A.M.; Kyei-Boahen, S.; Guimarães, M.D.F.; Nogueira, M.A.; Hungria, M. Feasibility of transference of inoculation-related technologies: A case study of evaluation of soybean rhizobial strains under the agro-climatic conditions of Brazil and Mozambique. Agric. Ecosyst. Environ. 2017, 261, 230–240. [Google Scholar] [CrossRef]
- Jiménez, J.A.; Novinscak, A.; Filion, M. Pseudomonas fluorescens LBUM677 differentially increases plant biomass, total oil content and lipid composition in three oilseed crops. J. Appl. Microbiol. 2020, 128, 1119–1127. [Google Scholar] [CrossRef]
- Kumawat, K.C.; Sharma, P.; Singh, I.; Sirari, A.; Gill, B.S. Co-existence of Leclercia adecarboxylata (LSE-1) and Bradyrhizobium sp. (LSBR-3) in nodule niche for multifaceted effects and profitability in soybean production. World J. Microbiol. Biotechnol. 2019, 35, 172. [Google Scholar] [CrossRef]
- Sibponkrung, S.; Kondo, T.; Tanaka, K.; Tittabutr, P.; Boonkerd, N.; Yoshida, K.-I.; Teaumroong, N. Co-Inoculation of Bacillus velezensis Strain S141 and Bradyrhizobium Strains Promotes Nodule Growth and Nitrogen Fixation. Microorganisms 2020, 8, 678. [Google Scholar] [CrossRef]
- Zeffa, D.M.; Fantin, L.H.; Koltun, A.; De Oliveira, A.L.; Nunes, M.P.; Canteri, M.G.; Gonçalves, L.S. Effects of plant growth-promoting rhizobacteria on co-inoculation with Bradyrhizobium in soybean crop: A meta-analysis of studies from 1987 to 2018. PeerJ 2020, 8, e7905. [Google Scholar] [CrossRef]
- Delamuta, J.R.M.; Menna, P.; Ribeiro, R.A.; Hungria, M. Phylogenies of symbiotic genes of Bradyrhizobium symbionts of legumes of economic and environmental importance in Brazil support the definition of the new symbiovars pachyrhizi and sojae. Syst. Appl. Microbiol. 2017, 40, 254–265. [Google Scholar] [CrossRef]
- Sharma, S.; Kulkarni, J.; Jha, B. Halotolerant Rhizobacteria Promote Growth and Enhance Salinity Tolerance in Peanut. Front. Microbiol. 2016, 7, 1600. [Google Scholar] [CrossRef]
- Irizarry, I.; White, J.F. Application of bacteria from non-cultivated plants to promote growth, alter root architecture and alleviate salt stress of cotton. J. Appl. Microbiol. 2017, 122, 1110–1120. [Google Scholar] [CrossRef]
- Santos, A.A.; Da Silveira, J.A.G.; Bonifacio, A.; Rodrigues, A.C.; Figueiredo, M.D.V.B. Antioxidant response of cowpea co-inoculated with plant growth-promoting bacteria under salt stress. Braz. J. Microbiol. 2018, 49, 513–521. [Google Scholar] [CrossRef]
- Anzuay, M.S.; Ciancio, M.G.R.; Ludueña, L.M.; Angelini, J.G.; Barros, G.; Pastor, N.; Taurian, T. Growth promotion of peanut (Arachis hypogaea L.) and maize (Zea mays L.) plants by single and mixed cultures of efficient phosphate solubilizing bacteria that are tolerant to abiotic stress and pesticides. Microbiol. Res. 2017, 199, 98–109. [Google Scholar] [CrossRef]
- Shahid, M.; Akram, M.S.; Khan, M.A.; Zubair, M.; Shah, S.M.; Ismail, M.; Shabir, G.; Basheer, S.; Aslam, K.; Tariq, M. A phy-tobeneficial strain Planomicrobium sp. MSSA-10 triggered oxidative stress responsive mechanisms and regulatedthe growth of pea plants under induced saline environment. J. Appl. Microbiol. 2018, 124, 1566–1579. [Google Scholar] [CrossRef]
- Yasin, N.A.; Akram, W.; Khan, W.U.; Ahmad, S.R.; Ahmad, A.; Ali, A. Halotolerant plant-growth promoting rhizobacteria modulate gene expression and osmolyte production to improve salinity tolerance and growth in Capsicum annum L. Environ. Sci. Pollut. Res. 2018, 25, 23236–23250. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, A.F. Effectiveness of exopolysaccharides and biofilm forming plant growth promoting rhizobacteria on salinity tolerance of faba bean (Vicia faba L.). Afr. J. Microbiol. Res. 2018, 12, 399–404. [Google Scholar] [CrossRef][Green Version]
- Khan, A.; Ali, L.; Chaudhary, H.J.; Hussain Munis, M.F.; Bano, A.; Masood, S. Bacillus pumilus alleviates boron toxicity in tomato (Lycopersicum esculentum L.) due to enhanced antioxidant enzymatic activity. Sci. Hortic. 2016, 200, 178–185. [Google Scholar]
- Qi, J.; Aiuchi, D.; Tani, M.; Asano, S.I.; Koike, M. Potential of Entomopathogenic Bacillus thuringiensis as Plant Growth Promoting Rhizobacteria and Biological Control Agents for Tomato Fusarium Wilt. Int. J. Environ. Agric. Res. 2016, 2, 55–63. [Google Scholar]
- Sharma, I.P.; Sharma, A.K. Physiological and biochemical changes in tomato cultivar PT-3 with dual inoculation of my-corrhiza and PGPR against root-knot nematode. Symbiosis 2017, 71, 175–183. [Google Scholar] [CrossRef]
- Sagar, A.; Ramteke, P.W. Isolation, characterization and evaluation of Multi traits Pseudomonas sp. KP966497 (PR21) for their growth promoting effect on Tomato (Lycopersicon esculentum Mill.). In Proceedings of the International Conference Climate Change, Biodiversity and Sustainable Agriculture (ICCBSA-2018), Jorhat, India, 13–16 December 2018. [Google Scholar]
- Orozco-Mosqueda, M.D.C.; Duan, J.; Dibernardo, M.; Zetter, E.; Campos-García, J.; Glick, B.R.; Santoyo, G. The Production of ACC Deaminase and Trehalose by the Plant Growth Promoting Bacterium Pseudomonas sp. UW4 Synergistically Protect Tomato Plants Against Salt Stress. Front. Microbiol. 2019, 10, 1392. [Google Scholar] [CrossRef]
- Hussein, K.A.; Joo, J.H. Plant Growth-Promoting Rhizobacteria Improved Salinity Tolerance of Lactuca sativa and Raphanus sativus. J. Microbiol. Biotechnol. 2018, 28, 938–945. [Google Scholar] [CrossRef]
- Tahir, M.; Ahmad, I.; Shahid, M.; Shah, G.M.; Farooq, A.B.U.; Akram, M.; Tabassum, S.A.; Naeem, M.A.; Khalid, U.; Ahmad, S.; et al. Regulation of antioxidant production, ion uptake and productivity in potato (Solanum tuberosum L.) plant inoculated with growth promoting salt tolerant Bacillus strains. Ecotoxicol. Environ. Saf. 2019, 178, 33–42. [Google Scholar] [CrossRef]
- Yu, Y.; Wang, N.; Hu, R.; Xiang, F. Genome-wide identification of soybean WRKY transcription factors in response to salt stress. Springerplus 2016, 5, 920. [Google Scholar] [CrossRef]
- Wu, M.; Liu, H.; Han, G.; Cai, R.; Pan, F.; Xiang, Y. A moso bamboo WRKY gene PeWRKY83 confers salinity tolerance in transgenic Arabidopsis plants. Sci. Rep. 2017, 7, 11721. [Google Scholar] [CrossRef]
- Barnawal, D.; Bharti, N.; Pandey, S.S.; Pandey, A.; Chanotiya, C.S.; Kalra, A. Plant growth-promoting rhizobacteria enhance wheat salt and drought stress tolerance by altering endogenous phytohormone levels and TaCTR1 /TaDREB2 expression. Physiol. Plant. 2017, 161, 502–514. [Google Scholar] [CrossRef]
- De la Torre-González, A.; Navarro-León, E.; Albacete, A.; Blasco, B.; Ruiz, J.M. Study of phytohormone profile and oxidative metabolism as key process to identification of salinity response in tomato commercial genotypes. J. Plant Physiol. 2017, 216, 164–173. [Google Scholar] [CrossRef]
- Joshi, R.; Sahoo, K.K.; Tripathi, A.K.; Kumar, R.; Gupta, B.K.; Pareek, A.; Singla Pareek, S.L. Knockdown of an inflo-rescence meristem-specific cytokinin oxidase—OsCKX2 in rice reduces yield penalty under salinity stress condition. Plant Cell Environ. 2018, 41, 936–946. [Google Scholar] [CrossRef]
- Asaf, S.; Khan, A.L.; Khan, M.A.; Al-Harrasi, A.; Lee, I.-J. Complete genome sequencing and analysis of endophytic Sphingomonas sp. LK11 and its potential in plant growth. 3 Biotech 2018, 8, 389. [Google Scholar] [CrossRef]
- Chen, L.; Liu, Y.; Wu, G.; Njeri, K.V.; Shen, Q.; Zhang, N.; Zhang, R. Induced maize salt tolerance by rhizosphere inoculation ofBacillus amyloliquefaciensSQR9. Physiol. Plant. 2016, 158, 34–44. [Google Scholar] [CrossRef]
- Safdarian, M.; Askari, H.; Shariati, V.S.; Nematzadeh, G. Transcriptional responses of wheat roots inoculated with Arthrobacter nitroguajacolicus to salt stress. Sci. Rep. 2019, 9, 1792. [Google Scholar] [CrossRef]
- Kazerooni, E.A.; Maharachchikumbura, S.S.N.; Adhikari, A.; Al-Sadi, A.M.; Kang, S.-M.; Kim, L.-R.; Lee, I.-J. Rhizospheric Bacillus amyloliquefaciens Protects Capsicum annuum cv. Geumsu gangsan From Multiple Abiotic Stresses via Multifarious Plant Growth-Promoting Attributes. Front. Plant Sci. 2021, 12, 669693. [Google Scholar] [CrossRef]
- Liu, H.; Wang, J.; Sun, H.; Han, X.; Peng, Y.; Liu, J.; Liu, K.; Ding, Y.; Wang, C.; Du, B. Transcriptome Profiles Reveal the Growth-Promoting Mechanisms of Paenibacillus polymyxa YC0136 on Tobacco (Nicotiana tabacum L.). Front. Microbiol. 2020, 11, 2384. [Google Scholar] [CrossRef]
- Akram, W.; Aslam, H.; Ahmad, S.R.; Anjum, T.; Yasin, N.A.; Khan, W.U.; Ahmad, A.; Guo, J.; Wu, T.; Luo, W.; et al. Bacillus megaterium strain A12 ameliorates salinity stress in tomato plants through multiple mechanisms. J. Plant Interact. 2019, 14, 506–518. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Crop | Strain | Na+Cl− Tolerance(mM) | Condition | Effect | Country | Reference |
|---|---|---|---|---|---|---|
| Wheat (Triticumaestivum) | Azotobacter chroococcumAZ6 | 200 | Pot | Proline and amino acid | Algérie | [42] |
| Bacillus Subtilis | 200 | Field | Improvement in plant growth | Russia | [43] | |
| Enterobacter cloacaeZNP-3 | 150–200 | Pot | Enlargement of agronomic traits and chlorophyll content | India | [44] | |
| Enterobacter cloacae SBP-8 | 200 | Pot | Antioxidant activity | India | [45] | |
| P. fluorescens | 18–36 | Invitro | Enhancement of agronomic traits | Iran | [46] | |
| Pseudomonas sp. and Bacillus sp. | Soil salinity | Pot | Plant growth promotion | India | [47] | |
| Rice (Oryza sativa) | Bacillus and Pseudomonas spp. | Saline soil | Field | Phytostimulation | India | [48] |
| Bacillus amyloliquefaciensRWL-1 | 120 | Pot | Increase in essential amino acids | Korea | [49] | |
| Pseudomonas aeruginosa [PRR1 and PHL3] and Lysinibacillus sp. [BPC2] | 100 mM | Pot | Germination percentage and root length | India | [50] | |
| Curtobacterium albidumSRV4 | 100–300 | Pot | Improve photosynthetsis, osmolytes, and antioxidative enzymes | India | [51] | |
| Enterobactersp. P23 | 0 to 200 | Invitro | Promote rice-seedling growth | India | [52] | |
| Enterobactersp. PR14 | 150–900 | Invitro | Growth promotion | India | [53] | |
| Maize (Zea mays) | Pseudomonas | 150 | Pot | Improve POD activity, proline, and soil moisture | Pakistan | [54] |
| Enterobactercloacae [KP226569] | 100–200 | In vitro | Increased growth parameters | India | [55] | |
| Azotobacternigricans [KP966496] | 100–200 | In vitro | Enlargement, growth parameters, and germination | India | [56] | |
| Bacillus subtilis | 0–200 | Plastic pots | Increased the relative water content in leaves | Brazil | [57] | |
| Rhizobium tropici and Azospirillum brasilense | 170 | Leonard jars | Antioxidant enzymes | Brazil | [58] | |
| Azospirillum lipoferum and Azotobacter chroococcum | 100 | Pot | Significantly enhanced growth parameters and pigments | S.Arabia | [59] | |
| Millets | E. cloacae [KP226575] | 100–200 | Greenhouse | Increase in seed germination andenhanced root and shoot elongation | India | [60] |
| Sorghum (Sorghum bicolor) | Azotobacter salinestris NBRC 102,611 | 50 | Pot | Improved plant growth, carbohydrate, proline, and macroelements | Egypt | [61] |
| Mung bean (Vigna radiata) | Enterobacter cloacae strain KBPD | 80 | Pot | Increase growth parameters, fresh and dry weights | India | [62] |
| Tomato (Solanumly copersicum) | Azotobacter chroococcum | 100 | Jar | Employed as bioeffectors | China | [63] |
| Azotobacter chroococcum 76A | 100 | Pot | Promoted plant growth | Italy | [64] | |
| Pseudomonas spp. OFT2 and OFT5 | 75 | pot | Promoted shoot uptake of other macronutrients and micronutrients | Japan | [65] | |
| Pea (Pisum sativum) | Bacillus subtilis RhStr_71, Bacillus safensis RhStr_223, and Bacillus cereus RhStr_JH5 | 10 | Pot | Enhanced the antioxidant enzymes | India | [66] |
| Enterobacter ludwigii Acinetobacter bereziniae, and Alcaligenes faecalis | 150 | Field trials | Seedling enlargement and yield | India | [67] | |
| Chickpea (Cicerarietinum) | Bacillus subtilis [BERA 71] | 200 | Plastic pots | Increased the synthesis of photosynthetic pigments and plant biomass | Saudi Arabia | [68] |
| Rhizobium sp. | 150 | Pot | Influenced photosynthesis and improved yield attributes | India | [69] | |
| Capsicum (Capsicum annuum L.) | Bacillus WU-9, WU-5, and WU-13 | 60 | Pot | Growth parameters and freshand dry weights | china | [70] |
| Red Pepper (Capsicum annuum) | Pseudomonas frederiksbergensis OB139 and vancouverensisOB155 | 150 | Plastic pots | Increased content of photosynthetic pigments | Republic of Korea | [71] |
| Okra (Abelmoschuse sculentus) | Enterobacter sp. UPMR18 | 75 | Plastic pots | Increased antioxidant enzyme activities | Malaysia | [72] |
| Shalgam (Brassica rapa) | Pseudomonas stutzeri ISE12 | 200 | Pot | Antioxidant defense system | Poland | [73] |
| Soybean (Glycine max) | Bacillus subtilis and P.pseudoalcaligenes | 100 | Hydroponics | Enhancement of growth parameters | India | [74] |
| Soybean | ALT29 and ALT43 | 80–2400 | Pot | Improvement in agronomic parameters, fresh and chlorophyll content | Korea | [75] |
| Canola (Brassica napus) | Enterobacter cloacae [HSNJ4] | 50–100 | Pot | Enhancement of IAA and ethylene content | China | [76] |
| Canola (Brassica napus) | Rhizobium sp. | 90 | GH | The increased dry weight of the root | Iran | [77] |
| Faba bean (Viciafaba) | Rhizobium leguminosarum | 0–120 | Marrakech-Haouz region | Increased plant biomass, nodule number, and nitrogen content | Morocco | [78] |
| Snap Bean (Phaseolus vulgaris) | Rhizobium–arbuscularmycorrhizal fungi [AMF] | 6–200 | Organic garden-shade house | Increased yield | Florida | [79] |
| Pegion Pea (Cajanus cajan) | Funneliformis mosseae and Rhizophagus irregularis | 0–100 | Pot | Symbiosis | India | [80] |
| Hopbush Shrub (Dodonae aviscosa L.) | Azospirillumlipoferum and Azotobacterchroococcum | 150–200 | Pot | Improved seedling growth parameters | Iran | [81] |
| Artemisia annuaL. | Piriformospora indica [Pi] and Azotobacter chroococcum [Az] | 200 | Pot | Enhanced activities of antioxidant enzymes | India | [82] |
| Sunflower (Helianthus) | Pseudomonas citronellolis SLP6 | 80 | pot | Increase in antioxidant enzymes and chlorophyll content | India | [83] |
| Arabidopsis thaliana | Pseudomonas putida | 200 | Pot | Auxin Activity RAR shows tryptophan trust for growth | India | [84] |
| Alfalfa (Medicago sativa L.) | S. meliloti ARh29, K. cowanii A37, Klebsiella sp. A36, and rhizobial strain | 1200 | Greenhouse and invitro | Plant growth | Iran | [85] |
| Alfalfa (Medicago sativa L.) | B. subtilis NRCB002, NRCB002, NRCB003 [NRCB003] | 130 | Greenhouse | Significantly increased dry weight | China | [86] |
| Medicago sativa | Klebsiella sp., Alcaligenes sp., Pseudomonas cedrina, and Pseudomonas putida | 20–60 | Pot | Plant development, chlorophyll content, and root AM colonization | Morocco | [87] |
| Pseudomonas aeruginosa | 170 | Field experiments | Significant increase in shoot length | Pakistan | [88] | |
| Talh Tree Acacia gerrardiiBenth. | AMF and B. subtilis | 250 | Pot | Increase in phenols, fiber content, and total lipids | Riyadh, KSA | [89] |
| Crop | Halotolerant PGPB | Tolerance Mechanism | Reference |
|---|---|---|---|
| Cereals | |||
| Rice | Thalassobacillus denorans and Oceanobacillus kapialis | Increase in germination % | [148] |
| strains S6 and S7 | Yield increased | [149] | |
| Enterobacterudwigii [AF134] and P. Putida [AF137] | Increased root and shoot biomass | [150] | |
| Enterobacter cloacae [KP226569] | Increase in germination percentage, root, and shoot | [151] | |
| Azotobacter sp. [KH-2] | Enlargement of plant height, yield, and biomass | [152] | |
| Alcaligenes sp. [AF7] | Increasingly support the vegetative growth parameters of crop | [153] | |
| Exiguobacterium sp., Stenotrophomonas sp, Enterobacter sp., Microbacterium sp., and Achromobacter sp. | Significantly improve proline, total chlorophyll, and total phenol | [154] | |
| Glutamicibacter sp. [YD01] | Enhanced stress-responsive gene expression and photosynthetic | [155] | |
| Bacillus tequilensis [UPMRB9], Providencia stuartii [UPMRG1], and B. aryabhattai [UPMRE6] | Improved chlorophyll contentand reduced electrolyte leakage | [156] | |
| Wheat | P. fluorescens, B. pumilus, Exiguobacterium aurantiacum | Enhancement of agronomic traits | [157] |
| Klebsiella sp. | Increase in total protein content and proline | [158] | |
| Azospirillum brasilense 65B [Abr65B] | Enhanced enlargement and nutrient access | [159] | |
| Erwinia sp. | Enhanced growth and yield parameters | [160] | |
| Bacillus megaterium, P fluorescens and B. subtilis | Increased grain yield | [161] | |
| Azospirillum sp., Azoarcus sp., and Azorhizobium sp. | Increase in biomass and the activity of enzymes | [162] | |
| Azotobacter sp. [Az1-Az6] | Improved agronomic characteristics | [163] | |
| Maize | Staphylococcus sciuri | Significantly increased growth parameters and biochemical characteristics | [164] |
| Pseudomonas sp. P8, Peribacillus sp. P10, Streptomyces sp. X52 | promoted the growth | [165] | |
| Bacillus aquimaris DY-3 | Increased chlorophyll content antioxidant enzymes and osmotic regulation | [166] | |
| Bacillus sp. SR-2-1/1 | increased contents of chlorophyll, total phenolics and proline | [167] | |
| Serratialiquefaciens [KM4] | Antioxidant enzymes, decrease in ABA biosynthesis, and nutrient uptake | [168] | |
| Bacillus sp. [HL3RS14] | Increased dry weights of roots and shoots | [169] | |
| B.safensis [NBRI 12 M] | Increased amounts of chlorophyll, proline, and soluble sugar | [170] | |
| Bacillus sp. [NBRI YN4.4] | Improved biochemical traits and soil enzymes | [171] | |
| Oat | Klebsiella sp. [IG 3] | Enhanced plant growth | [172] |
| Oil-yielding grains | |||
| Soybean | Bradyrhizobium sp. | Increased grain yield | [173] |
| P. fluorescens [LBUM677] | Improved plant biomass, lipid composition, and oil content | [174] | |
| Bradyrhizobium sp and L. adecarcoxylata | Enhanced plant growth and productivity | [175] | |
| B. velezensis [S141], and B.diazoefficiens [USDA110] | Improved nodulation and N2-fixing efficiency | [176,177] | |
| Sunflower | Bradyrhizobium sp. | nodY/K and nifH gene expression for salinity tolerance | [178] |
| Groundnut | Klebsiella, Pseudomonas, Agrobacterium, and Ochrobactrum | Enhanced PGP traits | [179] |
| Cotton seed | B. amyloliquefaciens, Curtobacterium oceanosedimentum, and Pseudomonas oryzihabitans | Seed germination | [180] |
| Legumes | |||
| Chickpea | Bradyrhizobium and Actinomadura, Paenibacillusgraminis | Induced increased antioxidant enzymes, and | [181] |
| Peanut (Arachish ypogaea L.) | Serratia sp. S119; Acinetobacter sp. L176; | Increased growth parameters and P content | [182] |
| Planomicrobium sp. MSSA-10 | Increased growth | [183] | |
| Black gram | P. fluorescens | Improved photosynthetic content | [184] |
| Faba bean | Pseudomonas anguilliseptica [SAW 24] | EPS production and biofilm production | [185] |
| Vegetables | |||
| Tomato | Bacillus pumilus | Significantly improved the fresh shoot weight and dry weight | [186] |
| B. thuringiensis | Seed germination and shoot elongation | [187] | |
| Pseudomonas jessenii and Pseudomonas synxantha | Significantly enhanced plant growth | [188] | |
| Pseudomonas sp. KP966497 [PR21] | Seed germination and shoot elongation | [189] | |
| Pseudomonas sp. UW4 | Enhanced plant growth | [190] | |
| Chinese cabbage, Lettuce, Radish | Lactobacillus sp. and P. putida, A. chroococcum | Increased the plumule and radicle lengths of germinated seeds | [191] |
| Capsicum | B. fortis strain [SSB21] | Reduced phytohormones | [184] |
| Potato (SolanumtuberosumL.) | Bacillus strains | Enhancement of plant growth | [192] |
| Halotolerant PGPB | Crops | Gene | Gene Product/Protein/ Enzyme | Significance | Reference |
|---|---|---|---|---|---|
| Bacillus megaterium | Tomato | Expression of MT2 and GR1 | Metallothionein Glutathione reductase enzyme synthesis | Antioxidant enzyme production, tolerance to salinity | [203] |
| Bacillus megaterium strain A12 [BMA12] | Tomato | PsbA and PBGD | Increased biosynthesis of chlorophyll | [203] | |
| Pseudomonas putida PS01 | A. thaliana | Upregulation of LOX2 | Jasmonic acid synthesis pathway, SOS pathway, abscisic acid [ABA] production | ROS activity, abscisic acid | [139] |
| Arthrobacter nitroguajacolicus | Wheat | Upregulation of AA0618700, AA0359620, APX, and GPX Expression ofAA0410390, AA1982260, AA0412840, andAA1872340 | SOS pathway and antioxidant enzyme | Plant-cell-wall biosynthesis, ROS activityosmotic balance | [117] |
| ALT29 and ALT43 | Soybean | [GmFLD19 and GmNARK] | Antioxidants [GSH, LPO, TPP, PPO, and POD], ion uptake [ Na and K] | Significantly increased growth parameters and biochemical traits | [75] |
| Bacillus amyloliquefaciens | Pepper | Upregulation of XTH [Xyloglucan Endotrans glucosylase/Hydrolase] | High biochemical traitsand antioxidant activities | Enhanced plant growth and tolerance for salinity and drought conditions | [201] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sagar, A.; Rai, S.; Ilyas, N.; Sayyed, R.Z.; Al-Turki, A.I.; El Enshasy, H.A.; Simarmata, T. Halotolerant Rhizobacteria for Salinity-Stress Mitigation: Diversity, Mechanisms and Molecular Approaches. Sustainability 2022, 14, 490. https://doi.org/10.3390/su14010490
Sagar A, Rai S, Ilyas N, Sayyed RZ, Al-Turki AI, El Enshasy HA, Simarmata T. Halotolerant Rhizobacteria for Salinity-Stress Mitigation: Diversity, Mechanisms and Molecular Approaches. Sustainability. 2022; 14(1):490. https://doi.org/10.3390/su14010490
Chicago/Turabian StyleSagar, Alka, Shalini Rai, Noshin Ilyas, R. Z. Sayyed, Ahmad I. Al-Turki, Hesham Ali El Enshasy, and Tualar Simarmata. 2022. "Halotolerant Rhizobacteria for Salinity-Stress Mitigation: Diversity, Mechanisms and Molecular Approaches" Sustainability 14, no. 1: 490. https://doi.org/10.3390/su14010490
APA StyleSagar, A., Rai, S., Ilyas, N., Sayyed, R. Z., Al-Turki, A. I., El Enshasy, H. A., & Simarmata, T. (2022). Halotolerant Rhizobacteria for Salinity-Stress Mitigation: Diversity, Mechanisms and Molecular Approaches. Sustainability, 14(1), 490. https://doi.org/10.3390/su14010490