
Community Differentiation and Ecological Influencing Factors along Environmental Gradients: Evidence from 1200 km Belt Transect across Inner Mongolia Grassland, China

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
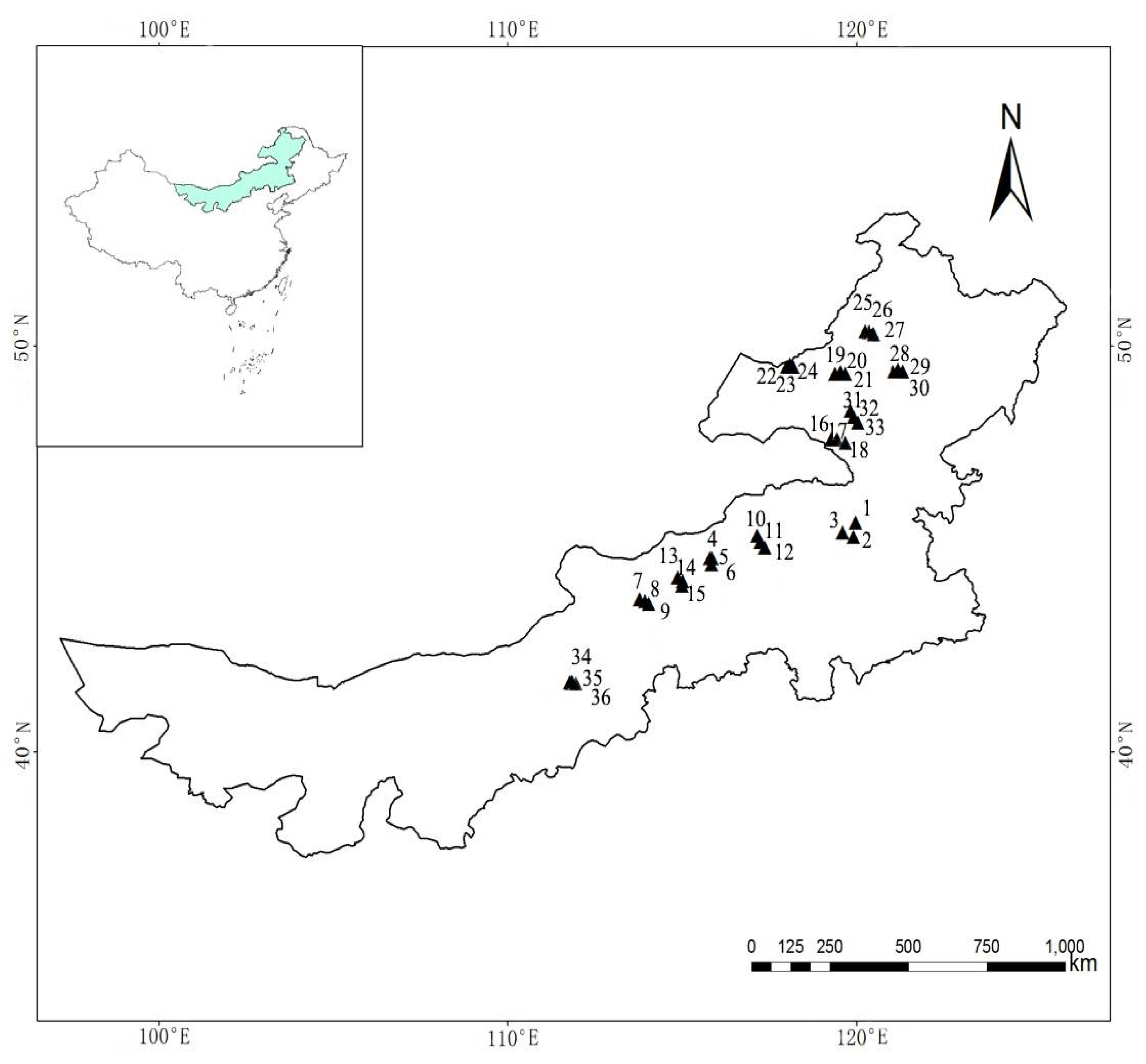
2.1. Study Area
2.2. Sampling Design
2.3. Measurement and Acquisition of Ecological Factors
2.4. Data Processing and Analysis
3. Results
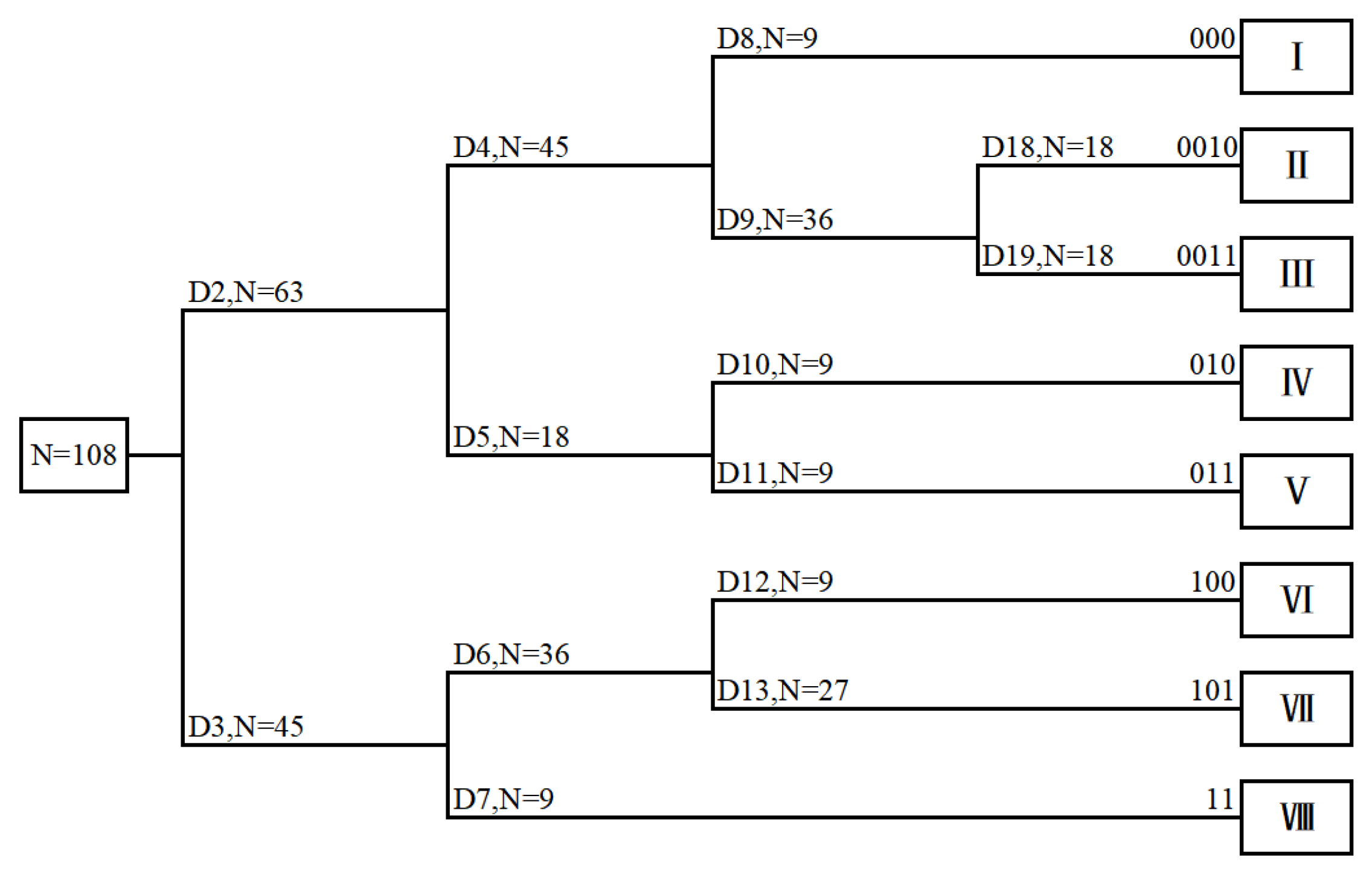
3.1. TWINSPAN Classification of Grassland Communities
- I.
- Stipa sareptana var. krylovii + Dysphania aristata, included three sites (S34, S35, S36), with low species richness, and a total of 16 plants. S. sareptana var. krylovii was the dominant species, D. aristata and S. bungeana were the suboptimal species. The total vegetation cover was 70−76%, the diversity and aboveground biomass were at the medium levels, while the evenness was relatively high.
- II.
- Stipa grandis + Leymus chinensis, included three sites (S10, S11, S12), with medium species richness, and a total of 19 plants. S. grandis was the dominant species, L. chinensis was the suboptimal species. The total vegetation cover was 78%−82%, the species diversity, and evenness were both high, while aboveground biomass was at a medium level.
- III.
- Stipa sareptana var. krylovii + Leymus chinensis, included nine sites (S4, S5, S6, S7, S8, S9, S13, S14, S15), with a total of 33 plants. S. sareptana var. krylovii was the dominant species, L. chinensis, C. squarrosa, and Allium ramosum were suboptimal species. The total vegetation cover was 52−76%, and the species diversity, evenness, and aboveground biomass were relatively low.
- IV.
- Stipa grandis+ Cleistogenes squarrosa, included three sites (S22, S23, S24), with a total of 16 plants. S. baicalensis was the dominant species, C. squarrosa was the suboptimal species. The total vegetation cover was 88%−92%. The species richness, species diversity, and evenness were all at the medium levels, while aboveground biomass was high.
- V.
- Stipa grandis + Carex duriuscula, included three sites (S19, S20, S21), with low species richness, and a total of 15 plants. S. grandis and C. duriuscula were both dominant species. The total vegetation cover was 90−95%. The species diversity, evenness, and aboveground biomass were all low.
- VI.
- Stipa baicalensis + Leymus chinensis, included three sites (S1, S2, S3), with a total of 31 plants. The dominant species was S. baicalensis, L. chinensis was the suboptimal species. The total vegetation cover was 90−95%. The species diversity and evenness were in the medium levels, while the mean aboveground biomass was prominent.
- VII.
- Carex pediformis + Stipa baicalensis, included nine sites (S16, S17, S18, S25, S26, S27, S31, S32, S33), with very high species richness, and a total of 69 plants. C. pediformis and S. baicalensis were both dominant species, the Agropyron cristatum was the suboptimal species. The total vegetation cover was 89−99%. The evenness was low, while the aboveground biomass was the highest (230.67 g·m−2).
- VIII.
- Leymus chinensis + Elymus dahuricus, included three sites (S28, S29, S30), with a total of 30 plants. L. chinensis and E. dahuricus were both the dominant species. The total vegetation cover was 97−98%. The species diversity, evenness, and aboveground biomass were relatively high.
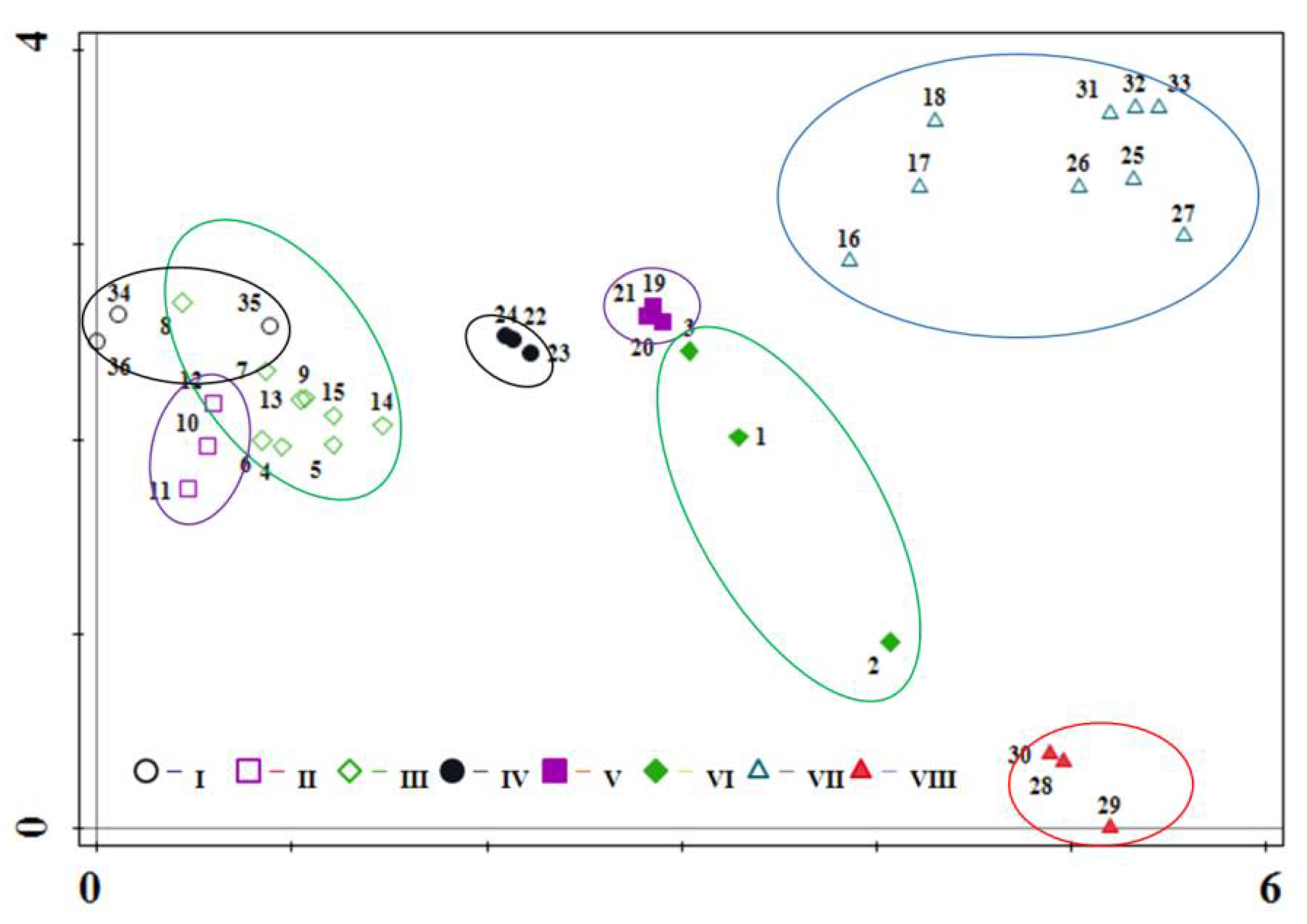
3.2. DCA Ordination
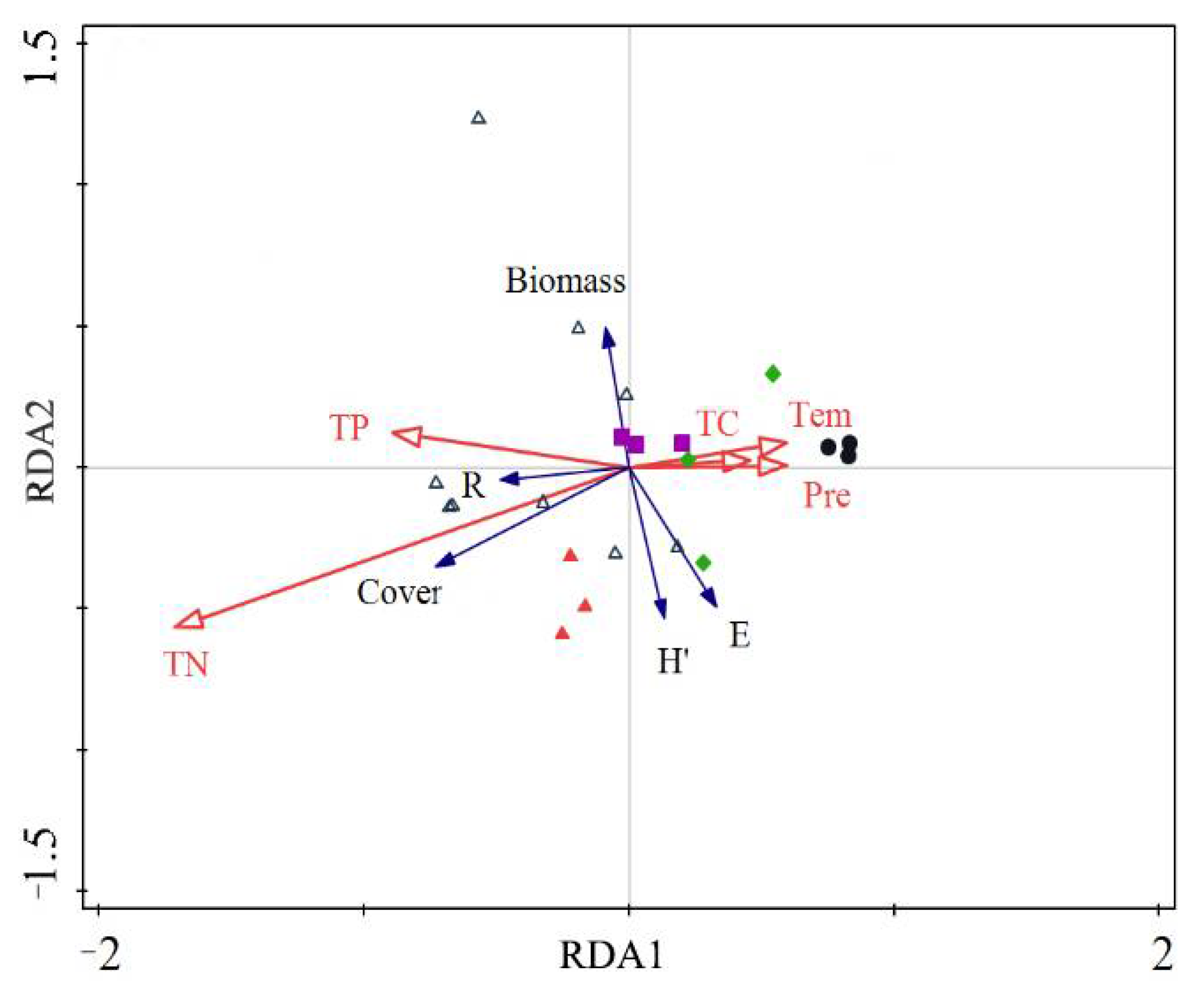
3.3. RDA Analysis in the Meadow Steppe
3.4. RDA Analysis in the Typical Steppe
4. Discussion
4.1. Spatial Differentiation Characteristics of Plant Community in the Inner Mongolia Grassland
4.2. Influencing Factors of Plant Community Characteristics in the Inner Mongolia Grassland
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hooper, D.U.; Chapin, F.S.; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S.; et al. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Jochum, M.; Fischer, M.; Isbell, F.; Roscher, C.; Plas, F.V.D.; Boch, S.; Boenisch, G.; Buchmann, N.; Catford, J.A.; Jeannine Cavender-Bares, J.; et al. The results of biodiversity−ecosystem functioning experiments are realistic. Nat. Ecol. Evol. 2020, 4, 1485–1494. [Google Scholar] [CrossRef] [PubMed]
- Li, W.H.; Hooper, D.U.; Wu, L.J.; Bakker, J.; Gianuca, A.T.; Wu, X.B.; Taube, F.; Wang, C.N.; Bai, Y.F. Grazing regime alters plant community structure via patch-scale diversity in semiarid grasslands. Ecosphere 2021, 12, e03547. [Google Scholar] [CrossRef]
- Parton, W.J.; Scurlock, J.M.O.; Ojima, D.S.; Gilmanov, T.G.; Scholes, R.J.; Schimel, D.S.; Kirchner, T.; Menaut, J.C.; Seastedt, T.; Moya, E.G.; et al. Observation and modeling of biomass and soil organic matter dynamics for the grassland biome worldwide. Glob. Biogeochem. Cycles 1993, 7, 785–809. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.F.; Lesi, M.C.; Zhang, M.M. Ecosystem pattern change and its influencing factors of “two barriers and three belts”. Chin. J. Ecol. 2019, 38, 2138–2148. [Google Scholar]
- Kong, Y.Y.; Luo, Q.Y.; Chen, Z. Study on the characteristic of vegetation composition and community succession of alpine meadow under three management modes. J. Yunnan Agric. Univ. Nat. Sci. 2020, 35, 1046–1053. [Google Scholar]
- Roscher, C.; Schumacher, J.; Gubsch, M.; Lipowsky, A.; Weigelt, A.; Buchmann, N.; Schmid, B.; Schulze, E.D. Using plant functional traits to explain diversity-productivity relationships. PLoS ONE 2012, 7, e36760. [Google Scholar] [CrossRef] [PubMed]
- Fraser, L.H.; Pither, J.; Jentsch, A.; Sternberg, M.; Zobel, M.; Askarizadeh, D.; Bartha, S.; Beierkuhnlein, C.; Bennett, J.A.; Bittel, A.; et al. Worldwide evidence of a unimodal relationship between productivity and plant species richness. Science 2015, 349, 302–305. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.T.; Jiang, L.; Zhang, Y. Relationships between functional diversity and aboveground biomass production in the Northern Tibetan alpine grasslands. Sci. Rep. 2016, 6, 34105. [Google Scholar] [CrossRef] [Green Version]
- Dong, S.K.; Tang, L.; Zhang, X.F.; Liu, S.L.; Liu, Q.R.; Su, X.K.; Zhang, Y.; Wu, X.Y.; Zhao, Z.Z.; Li, Y.; et al. Relationship between plant species diversity and functional diversity in alpine grasslands. Acta Ecol. Sin. 2017, 37, 1472–1483. [Google Scholar]
- Zhang, Q.; Buyantuev, A.; Li, F.Y.H.; Jiang, L.; Niu, J.M.; Ding, Y.; Kang, S.; Ma, W.J. Functional dominance rather than taxonomic diversity and functional diversity mainly affects community aboveground biomass in the Inner Mongolia grassland. Ecol. Evol. 2017, 7, 1605–1615. [Google Scholar] [CrossRef] [PubMed]
- Dai, C.; Kang, M.Y.; Ji, W.Y.; Jiang, Y. Responses of belowground biomass and biomass allocation to environmental factors in central grassland of Inner Mongolia. Acta Agrestia Sin. 2012, 20, 268–274. [Google Scholar]
- Dorji, T.; Moe, S.R.; Klein, J.A.; Totland, Ø. Plant species richness, evenness, and composition along environmental gradients in an alpine meadow grazing ecosystem in central Tibet, China. Arct. Antarct. Alp. Res. 2014, 46, 308–326. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Hou, X.Y.; Li, F.Y.H.; Niu, J.M.; Zhou, Y.L.; Ding, Y.; Zhao, L.Q.; Li, X.; Ma, W.J.; Kang, S. Alpha, beta and gamma diversity differ in response to precipitation in the Inner Mongolia grassland. PLoS ONE 2014, 9, e935183. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.J.; Su, P.X.; Zhou, Z.J.; Shi, R. Belowground bud bank distribution and aboveground community characteristics along different moisture gradients of alpine meadow in the Zoige plateau, China. Sustainability 2019, 11, 2602. [Google Scholar] [CrossRef] [Green Version]
- Adomou, A.C.; Akoègninou, A.; Sinsin, B.; Foucault, B.D.; Maesen, L.J.G.V.D. Notulae Florae Beninensis, 13-Biogeographical analysis of the vegetation in Benin. Acta Bot. Gall. 2007, 154, 221–233. [Google Scholar] [CrossRef]
- Wang, Y.H.; Zhao, M.F.; Kang, M.Y. Distribution pattern of main steppe communities in Inner Mongolia region and environmental interpretation. J. Beijing Norm. Univ. Nat. Sci. 2016, 52, 83–90. [Google Scholar]
- Chen, B.R.; Zhu, Y.X.; Zhang, H.B.; Zhou, L.; Xin, X.P. Quantitative classification and ordination of meadow grassland vegetations in Hulunber. J. Wuhan Bot. Res. 2008, 26, 476–481. [Google Scholar]
- Chen, B.R.; Li, H.S.; Zhu, Y.X.; Li, G.; Xin, X.P.; Zhang, H.B.; Zhou, L. The spatial pattern and environmental interpretation of the plant community of Hulunber grassland. Acta Ecol. Sin. 2010, 30, 1265–1271. [Google Scholar]
- Sha, W.; Dong, S.K.; Liu, S.L.; Liu, Q.R.; Shi, J.B.; Li, X.W.; Su, X.K.; Wu, Y. Spatial patterns of plant community biomass and species diversity in Aerjin Mountain Nature Reserve and their influencing factors. Chin. J. Ecol. 2016, 35, 330–337. [Google Scholar]
- Fu, Z.Y.; Jiang, H.; Wang, G.Q.; Xue, B.L.; Wang, H.Q. Effects of soil properties on plant community structure in a semi−arid grassland. Chin. J. Ecol. 2018, 37, 823–830. [Google Scholar]
- Wang, Y.T.; Bi, Y.T.; Wang, Q.; Hao, H.C.; Xu, M.H. Spatial differentiation of plant species diversity in subalpine meadows in Shanxi Province. Chin. Agric. Sci. Bull. 2018, 34, 77–83. [Google Scholar]
- Shan, D.; Zhu, Y.J.; Liu, Y.S.; Shi, Z.J.; Yang, X.H. Differentiation of vegetation types and biodiversity in the southern edge of Hulunbuir grassland. Chin. J. Ecol. 2019, 38, 619–626. [Google Scholar]
- Klaus, V.H.; Whittingham, M.J.; Báldi, A.; Eggers, S.; Francksen, R.M.; Hiron, M.; Lellei-Kovacs, E.; Rhymer, C.M.; Buchmann, N. Do biodiversity−ecosystem functioning experiments inform stakeholders how to simultaneously conserve biodiversity and increase ecosystem service provisioning in grasslands? Biol. Conserv. 2020, 245, 108552. [Google Scholar] [CrossRef]
- Chinese Academy of Sciences. Vegetation Atlas of China; Science Press: Beijing, China, 2001. [Google Scholar]
- Bai, Y.F.; Han, X.G.; Wu, J.G.; Chen, Z.Z.; Li, L.H. Ecosystem stability and compensatory effects in the Inner Mongolia grassland. Nature 2004, 431, 181–184. [Google Scholar] [CrossRef]
- Ma, K.P.; Huang, J.H.; Yu, S.L.; Chen, L.Z. Plant community diversity in Dongling Mountain, Beijing, China: II Species richness, evenness and species diversity. Acta Ecol. Sin. 1995, 15, 268–277. [Google Scholar]
- Hájek, M.; Hájková, P.; Roleček, J. A novel dataset of permanent plots in extremely species-rich temperate grasslands. Folia Geobot. 2020, 55, 257–268. [Google Scholar] [CrossRef]
- Yang, Y.H.; Fang, J.Y.; Ma, W.H.; Guo, D.L.; Mohammat, A. Large-scale pattern of biomass partitioning across China’s grasslands. Glob. Ecol. Biogeogr. 2010, 19, 268–277. [Google Scholar] [CrossRef]
- Bao, S.D. Agricultural and Chemistry Analysis of Soil; Chinese Agriculture Press: Beijing, China, 2000. [Google Scholar]
- Liu, C.R.; Ma, K.P.; Yu, S.L.; Wang, W. Plant community diversity in Dongling Mountain, Beijing, China: IV. Effects of sample size on diversity measures. Acta Ecol. Sin. 1997, 17, 584–592. [Google Scholar]
- Nie, Y.Y.; Chen, J.Q.; Xin, X.P.; Xu, L.J.; Yang, G.X.; Wang, X. Responses of niche characteristics and species diversity of main plant populations to duration of enclosure in the Hulun Buir meadow steppe. Acta Prataculturae Sin. 2021, 30, 15–25. [Google Scholar]
- Grandin, U. PC-ORD version 5: A user-friendly toolbox for ecologists. J. Veg. Sci. 2005, 17, 843–844. [Google Scholar] [CrossRef]
- Lai, J.S. Canoco 5: A new version of an ecological multivariate data ordination program. Biodiversity. Sci. 2013, 21, 765–768. [Google Scholar]
- Bai, Y.F.; Li, L.H.; Wang, Q.B.; Zhang, L.X.; Zhang, Y.; Chen, Z.Z. Changes in plant species diversity and productivity along gradients of precipitation and elevation in the Xinlin River Basin. Acta Phytoecol. Sin. 2000, 24, 667–673. [Google Scholar]
- Chytrý, M.; Otýpková, Z. Plot sizes used for phytosociological sampling of European vegetation. J. Veg. Sci. 2003, 14, 563–570. [Google Scholar] [CrossRef]
- Shen, X.J.; Jiang, M.; Lu, X.G.; Liu, X.T.; Liu, B.; Zhang, J.Q.; Wang, X.W.; Tong, S.Z.; Lei, G.C.; Wang, S.Z.; et al. Aboveground biomass and its spatial distribution pattern of herbaceous marsh vegetation in China. Sci. China Earth Sci. 2021, 64, 1115–1125. [Google Scholar] [CrossRef]
- Xu, Y.J.; Zhang, Y.; Li, W.; Liu, W.Y.; Gu, X.; Guan, Z.H.; Yang, J.; Lu, Z.Y. Effects of tree functional diversity and environmental gradients on belowground biomass in a natural old-growth forest ecosystem. Can. J. For. Res. 2019, 49, 1623–1632. [Google Scholar] [CrossRef]
- Zuo, W.Q.; Wang, Y.H.; Wang, F.Y.; Shi, G.X. Effects of enclosure on the community characteristics of Leymus chinensis in degenerated steppe. Acta Prataculturae Sin. 2009, 18, 12–19. [Google Scholar]
- Lu, S.Z.; Qiao, X.G.; Zhao, L.Q.; Wang, Z.; Gao, C.G.; Wang, J.; Guo, K. Basic characteristics of Stipa sareptana var. krylovii communities in China. Chin. J. Plant Ecol. 2020, 44, 1087–1094. [Google Scholar] [CrossRef]
- Mokany, K.; Raison, R.J.; Prokushkin, A.S. Critical analysis of root: Shoot ratios in terrestrial biomes. Global Change Biol. 2006, 12, 84–96. [Google Scholar] [CrossRef]
- Manning, P.; Vries, F.T.D.; Tallowin, J.R.B.; Smith, R.; Mortimer, S.R.; Pilgrim, E.S.; Harrison, K.A.; Wright, D.G.; Quirk, H.; Benson, J.; et al. Simple measures of climate, soil properties and plant traits predict national-scale grassland soil carbon stocks. J. Appl. Ecol. 2015, 52, 1188–1196. [Google Scholar] [CrossRef] [Green Version]
- Siefert, A.; Ravenscroft, C.; Althoff, D.; Alvarez−Yépiz, J.C.; Carter, B.E.; Glennon, K.L.; Heberling, J.M.; Jo, I.S.; Pontes, A.; Sauer, A.; et al. Scale dependence of vegetation−environment relationships: A meta-analysis of multivariate data. J. Veg. Sci. 2012, 23, 942–951. [Google Scholar] [CrossRef]
- Chase, J.M. Drought mediates the importance of stochastic community assembly. Proc. Natl. Acad. Sci. USA 2007, 104, 17430–17434. [Google Scholar] [CrossRef] [Green Version]
- Qiao, X.G. Community Ecology of Stipa Steppes in China; Institute of Botany, Chinese Academy of Sciences: Beijing, China, 2019. [Google Scholar]
- Fang, J.Y.; Song, Y.C.; Liu, H.Y.; Piao, S.L. Vegetation climate relationship and its application in the division of vegetation zone in China. Acta Bot. Sin. 2002, 44, 1105–1122. [Google Scholar]
- Bai, Y.F.; Wu, J.G.; Pan, Q.M.; Huang, J.H.; Wang, Q.B.; Li, F.S.; Buyantuyev, A.; Han, X.G. Positive linear relationship between productivity and diversity: Evidence from the Eurasian Steppe. J. Appl. Ecol. 2007, 44, 1023–1034. [Google Scholar] [CrossRef]
- Bai, Y.F.; Wu, J.G.; Xing, Q.; Pan, Q.M.; Huang, J.H.; Yang, D.L.; Han, X.G. Primary production and rain use efficiency across a precipitation gradient on the Mongolia Plateau. Ecology 2008, 89, 2140–2153. [Google Scholar] [CrossRef]
- Divíšek, J.; Hájek, M.; Jamrichová, E.; Petr, L.; Večeřa, M.; Tichý, L.; Willner, W.; Horsák, M. Holocene matters: Landscape history accounts for current species richness of vascular plants in forests and grasslands of eastern Central Europe. J. Biogeogr. 2020, 47, 721–735. [Google Scholar] [CrossRef]
- Ma, J.J.; Yao, H.; Feng, Z.Y.; Zhang, S.L. Changes in plant functional groups and species diversity under three grassland using modes in typical grassland area of Inner Mongolia, China. Chin. J. Plant Ecol. 2012, 36, 20200675. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Li, S.Y. Relationship between soil phosphorus and plant. World For. Res. 2013, 26, 19–24. [Google Scholar]
- Xian, Y.N.; Chen, C.; Fang, X.; Wang, L.F.; Chen, J.L.; Gu, X.; Zhang, S.J. Variation characteristics of soil total phosphorus and effective phosphorus concentrations in four vegetation type soils in the central hilly area of Hunan Province, China. J. Cent. South Univ. For. Technol. 2019, 39, 79–88. [Google Scholar]
- Long, M.; Wu, H.H.; Smith, M.D.; Pierre, K.J.L.; Lü, X.T.; Zhang, H.Y.; Han, X.G.; Yu, Q. Nitrogen deposition promotes phosphorus uptake of plants in a semi−arid temperate grassland. Plant Soil. 2016, 408, 475–484. [Google Scholar] [CrossRef]
- Dobben, H.F.V.; Wamelink, G.W.W.; Slim, P.A.; Kamiński, J.; Piórkowski, H. Species−rich grassland can persist under nitrogen−rich but phosphorus-limited conditions. Plant Soil 2017, 411, 451–466. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetation Types | TVC (%) | R | H′ | E | AB (g·m−2) |
|---|---|---|---|---|---|
| Stipa sareptana var. krylovii + Dysphania aristata | 73.7 ± 3.2 | 9.7 ± 1.2 | 1.58 ± 0.21 | 0.70 ± 0.06 | 111.25 ± 17.8 |
| Stipa sareptana var. krylovii + Leymus chinensis | 80.3 ± 2.1 | 11.3 ± 3.1 | 1.78 ± 0.17 | 0.75 ± 0.09 | 101.68 ± 7.72 |
| Stipa sareptana var. krylovii + Leymus chinensis | 65.3 ± 8.6 | 11.8 ± 3.0 | 1.36 ± 0.34 | 0.56 ± 0.12 | 63.28 ± 24.55 |
| Stipa grandis + Cleistogenes squarrosa | 90.0 ± 2.0 | 12.0 ± 1.0 | 1.56 ± 0.52 | 0.62 ± 0.19 | 168.26 ± 53.84 |
| Stipa grandis + Carex duriuscula | 92.3 ± 2.5 | 9.7 ± 2.5 | 1.05 ± 0.10 | 0.47 ± 0.10 | 91.95 ± 29.57 |
| Stipa baicalensis + Leymus chinensis | 92.3 ± 2.5 | 13.7 ± 0.6 | 1.44 ± 0.14 | 0.55 ± 0.05 | 190.15 ± 31.52 |
| Carex pediformis + Stipa baicalensis | 95.4 ± 0.3 | 18.7 ± 3.2 | 1.20 ± 0.33 | 0.41 ± 0.10 | 230.67 ± 107.57 |
| Leymus chinensis + Elymus dahuricus | 97.7 ± 0.6 | 20.0 ± 1.0 | 2.11 ± 0.02 | 0.71 ± 0.01 | 189.49 ± 36.53 |
| Ecological Factors | Simple Effects (%) | Conditional Effects (%) | Contribution (%) | Pseudo−F | p |
|---|---|---|---|---|---|
| TN | 18.8 | 18.8 | 48 | 4.4 | 0.006 ** |
| Tem | 2.2 | 8.4 | 21.5 | 0.4 | 0.12 |
| Pre | 2.1 | 8 | 20.4 | 2 | 0.126 |
| TC | 1.3 | 2.2 | 5.7 | 2 | 0.676 |
| TP | 4.9 | 1.7 | 4.4 | 0.6 | 0.784 |
| Ecological Factors | Simple Effects (%) | Conditional Effects (%) | Contribution (%) | Pseudo–F | p |
|---|---|---|---|---|---|
| Pre | 42.5 | 42.5 | 63.6 | 9.6 | 0.002 ** |
| TP | 39.6 | 15.3 | 22.9 | 4.4 | 0.006 ** |
| Tem | 19.7 | 7.5 | 11.2 | 2.4 | 0.068 |
| TN | 39.4 | 1.3 | 2 | 0.4 | 0.788 |
| TC | 32.2 | 0.2 | 0.3 | <0.1 | 0.992 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, Z.; Wang, F.; Lu, Z.; Zhang, M.; Zhang, L.; Hao, W.; Zhao, L.; Jiang, Y.; Gao, B.; Chen, R.; et al. Community Differentiation and Ecological Influencing Factors along Environmental Gradients: Evidence from 1200 km Belt Transect across Inner Mongolia Grassland, China. Sustainability 2022, 14, 361. https://doi.org/10.3390/su14010361
Fu Z, Wang F, Lu Z, Zhang M, Zhang L, Hao W, Zhao L, Jiang Y, Gao B, Chen R, et al. Community Differentiation and Ecological Influencing Factors along Environmental Gradients: Evidence from 1200 km Belt Transect across Inner Mongolia Grassland, China. Sustainability. 2022; 14(1):361. https://doi.org/10.3390/su14010361
Chicago/Turabian StyleFu, Zhanyong, Fei Wang, Zhaohua Lu, Meng Zhang, Lin Zhang, Wenyue Hao, Ling Zhao, Yang Jiang, Bing Gao, Rui Chen, and et al. 2022. "Community Differentiation and Ecological Influencing Factors along Environmental Gradients: Evidence from 1200 km Belt Transect across Inner Mongolia Grassland, China" Sustainability 14, no. 1: 361. https://doi.org/10.3390/su14010361
APA StyleFu, Z., Wang, F., Lu, Z., Zhang, M., Zhang, L., Hao, W., Zhao, L., Jiang, Y., Gao, B., Chen, R., & Wang, B. (2022). Community Differentiation and Ecological Influencing Factors along Environmental Gradients: Evidence from 1200 km Belt Transect across Inner Mongolia Grassland, China. Sustainability, 14(1), 361. https://doi.org/10.3390/su14010361