
Root Characteristics and Water Erosion-Reducing Ability of Alpine Silver Grass and Yushan Cane for Alpine Grassland Soil Conservation

Abstract
:1. Introduction
2. Materials and Methods
2.1. Site and Vegetation
2.2. Plant Raising
2.3. Growth Characteristics
2.4. Uprooting Test
2.5. Root Tensile Test
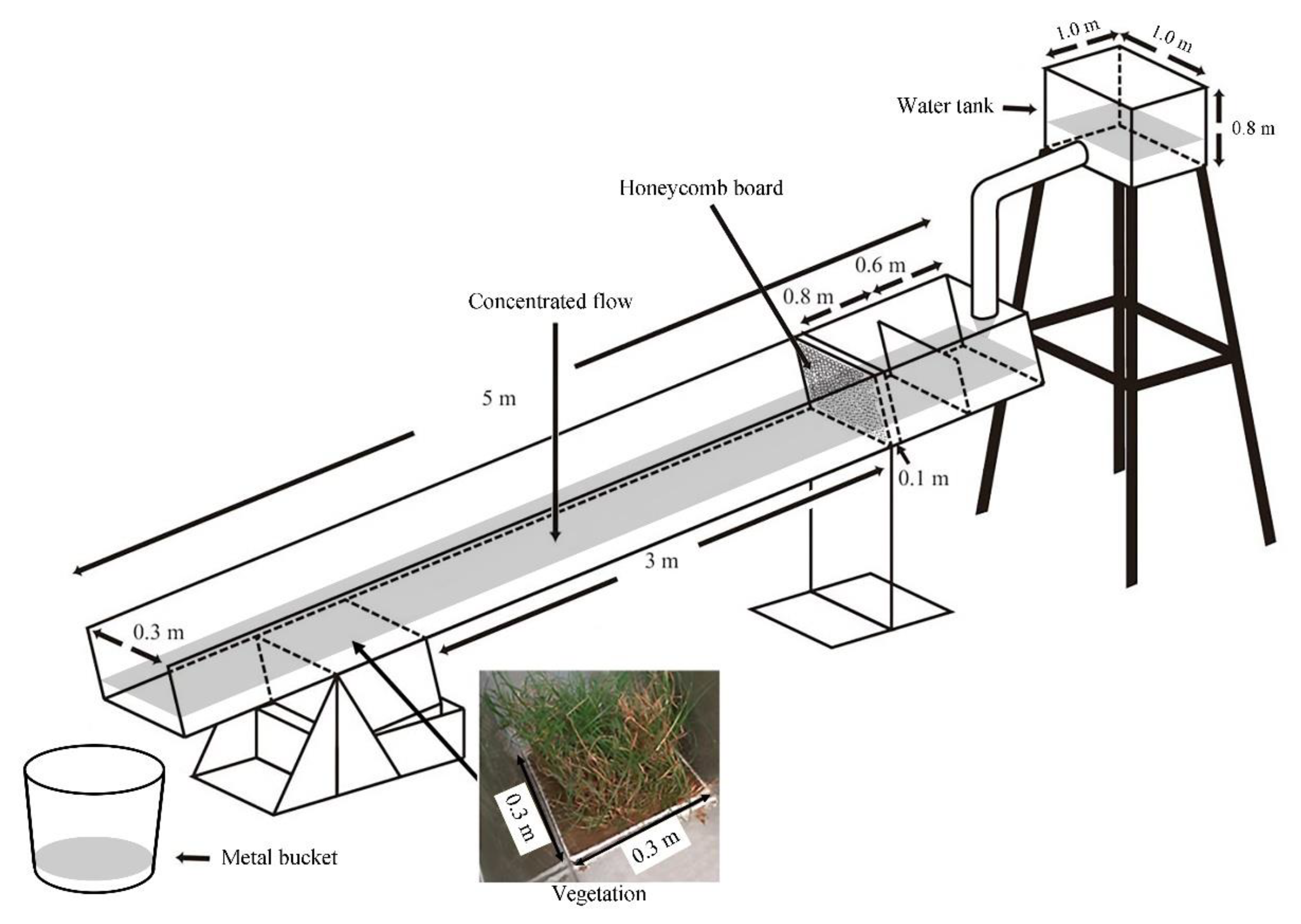
2.6. Hydraulic Flume Experiment
2.7. Statistical Analysis
3. Results
3.1. Root System Architecture
3.2. Growth Characteristics
3.3. Root Anchorage Ability
3.4. Root Tensile Strength
3.5. Water Erosion-Reducing Ability
4. Discussion
4.1. Root System Architecture
4.2. Growth Characteristics
4.3. Root Anchorage Ability
4.4. Root Tensile Strength
4.5. Water Erosion-Reducing Ability
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Juárez-Orozco, S.M.; Siebe, C.; Fernández, D.F. Causes and effects of forest fires in tropical rainforests: A bibliometric approach. Trop. Conserv. Sci. 2017, 10, 1–14. [Google Scholar] [CrossRef]
- Jhu, J.J.; Li, F.Q. Forest degradation/decline: Research and practice. Chin. J. Appl. Ecol. 2007, 18, 1601–1609. [Google Scholar]
- Ruokolainen, L.; Salo, K. The effect of fire intensity on vegetation succession on a sub-xeric heath during ten years after wildfire. Ann. Bot. Fennici 2009, 46, 30–42. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Jiang, J.; Li, G.X.; Ma, X.Y. Characteristics of pulsed runoff-erosion events under typical rainstorms in a small watershed on the Loess Plateau of China. Sci. Rep. 2018, 8, 3672. [Google Scholar] [CrossRef]
- Abbate, A.; Longoni, L.; Ivanov, V.I.; Papini, M. Wildfire impacts on slope stability triggering in mountain areas. Geoscience 2019, 9, 417. [Google Scholar] [CrossRef] [Green Version]
- Girard, F.; Payette, S.; Delwaide, A. Patterns of early postfire succession of alpine, subalpine and lichen-woodland vegetation: 21 years of monitoring from permanent plots. Forests 2017, 8, 346. [Google Scholar] [CrossRef] [Green Version]
- Ivanova, G.A.; Ivanova, V.A.; Kovaleva, N.M.; Conard, S.G.; Zhila, S.V.; Tauasov, P.A. Succession of vegetation after a high-intensity fire in a pine forest with lichens. Contemp. Probl. Ecol. 2017, 10, 52–61. [Google Scholar] [CrossRef]
- Chou, C.H.; Lee, Y.F. Allelopathic dominance of Miscanthus transmorrisonensis in an alpine grassland community in Taiwan. J. Chem. Ecol. 1991, 17, 2267–2281. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.A.; Chen, W.C.; Tseng, Y.H.; Liao, M.C.; Wang, W.; Tseng, H.Y. Vegetation recovery after fire of subalpine grassland on the east peak of Mt. Shei. Q. J. For. Res. 2015, 37, 5–18. Available online: https://www.researchgate.net/publication/321010797_Vegetation_Recovery_after_Fire_of_Subalpine_Grassland_on_the_East_Peak_of_Mt_Shei (accessed on 1 June 2021).
- Zhou, B.Z.; Fu, M.Y.; Xie, J.Z.; Yang, X.S.; Li, Z.C. Ecological functions of bamboo forest: Research and Application. J. For. Res. 2005, 16, 143–147. [Google Scholar]
- Reubens, B.; Poesen, J.; Danjon, F.; Geudens, G.; Muys, B. The role of fine and coarse roots in shallow slope stability and soil erosion control with a focus on root system architecture: A review. Trees 2007, 21, 385–402. [Google Scholar] [CrossRef]
- Yen, C.P. The major patterns of root growth of the useful plants for soil conservation in Taiwan. J. Chin. Soil Water Conserv. 1973, 4, 65–85. [Google Scholar]
- Pohl, M.; Stroude, R.; Buttler, A.; Rixen, C. Functional traits and root morphology of alpine plants. Ann. Bot. 2011, 108, 537–545. [Google Scholar] [CrossRef] [Green Version]
- Hudek, C.; Sturrock, C.J.; Atkinson, B.S.; Stanchi, S.; Freppaz, M. Root morphology and biomechanical characteristics of high altitude alpine plant species and their potential application in soil stabilization. Ecol. Eng. 2017, 109, 228–239. [Google Scholar] [CrossRef]
- Mickovski, S.B.; Van Beek, L.P.H.; Salin, F. Uprooting of vetiver uprooting resistance of vetiver grass (Vetiveria zizanioides). Plant Soil 2005, 278, 33–41. [Google Scholar] [CrossRef]
- Burylo, M.; Rey, F.; Roumet, C.; Buisson, E.; Dutoit, T. Linking plant morphological traits to uprooting resistance in eroded marly lands (Southern Alps, France). Plant Soil 2009, 324, 31–42. [Google Scholar] [CrossRef]
- Lee, J.T.; Chu, M.Y.; Lin, Y.S.; Kung, K.N.; Lin, W.C.; Lee, M.J. Root traits and biomechanical properties of three tropical pioneer tree species for forest restoration in landslide areas. Forests 2020, 11, 179. [Google Scholar] [CrossRef] [Green Version]
- Pal, A.K.; Ahmed, A.; Panday, V.K. Soil binding capability of different forage grasses in terms of reinforcement ability toward soil slope stabilization. Ind. J. Hill Farming 2019, 32, 137–143. [Google Scholar]
- Comino, E.; Druetta, A. The effect of Poaceae roots on the shear strength of soils in the Italian alpine environment. Soil Tillage Res. 2010, 106, 194–201. [Google Scholar] [CrossRef]
- De Baets, S.; Poesen, J.; Reubens, B.; Wemans, K.; De Baerdemaeker, J.; Muys, B. Root tensile strength and root distribution of typical Mediterranean plant species and their contribution to soil shear strength. Plant Soil 2008, 305, 207–226. [Google Scholar] [CrossRef]
- De Baets, S.; Poesen, J.; Gyssels, G.; Knapen, A. Effects of grass roots on the erodibility of topsoils during concentrated flow. Geomorphology 2006, 76, 54–67. [Google Scholar] [CrossRef]
- De Baets, S.; Poesen, J.; Knapen, A.; Galindo, P. Impact of root architecture on the erosion-reducing potential of roots during concentrated flow. Earth Surf. Process. Landf. 2007, 32, 1323–1345. [Google Scholar] [CrossRef]
- De Baets, S.; Poesen, J.; Meersman, J.; Serlet, L. Cover crops and their erosion-reducing effects during concentrated flow erosion. Catena 2011, 85, 237–244. [Google Scholar] [CrossRef]
- Vannoppen, W.; De Baets, S.; Keeble, J.; Dong, Y.; Poesen, J. How do root and soil characteristics affect the erosion-reducing potential of plant species? Ecol. Eng. 2017, 109, 186–195. [Google Scholar] [CrossRef] [Green Version]
- Vannoppen, W.; Vannoppen, W.; De Baets, S.; Poesen, J. A review of the mechanical effects of plant roots on concentrated flow erosion rates. Earth Sci. Rev. 2015, 150, 666–678. [Google Scholar] [CrossRef] [Green Version]
- Gyssels, G.; Poesen, J. The importance of plant root characteristics in controlling concentrated flow erosion rates. Earth Surf. Process. Landf. 2003, 28, 371–384. [Google Scholar] [CrossRef]
- Burylo, M.; Rey, F.; Mathys, N.; Dutoit, T. Plant root traits affecting the resistance of soils to concentrated flow erosion. Earth Surf. Process. Landf. 2012, 37, 1463–1470. [Google Scholar] [CrossRef]
- King, H.B. Genesis and classification of soils developed under Yushan cane (Yushania niitakayamensis) grassland in the Hohuan Mountain area. Bull. Taiwan For. Res. Inst. New Ser. 1993, 8, 21–38. [Google Scholar]
- Böhm, W. Excavation methods. In Methods of Studying Root Systems; Billings, W.D., Colley, F., Lange, O.L., Olson, J.S., Eds.; Springer: Berlin, Germany, 1979; pp. 5–18. [Google Scholar]
- Bouma, T.J.; Nielsen, K.L.; Koutstaal, K. Sample preparation and scanning protocol for computerized analysis of root length and diameter. Plant Soil 2000, 218, 185–196. [Google Scholar] [CrossRef]
- Pang, W.; Crow, W.T.; Luc, J.E.; McSorley, R.; Giblin-Davis, R.M.; Kruse, J.K. Comparison of water displacement and WinRHIZO software for plant root parameter assessment. Plant Dis. 2011, 95, 1308–1310. [Google Scholar] [CrossRef] [Green Version]
- Gould, I.J.; Quinton, J.N.; Weigelt, A.; De Deyn, G.B.; Bardgett, R.D. Plant diversity and root traits benefit physical properties key to soil function in grasslands. Ecol. Lett. 2016, 19, 1140–1149. [Google Scholar] [CrossRef]
- Loades, K.W.; Bengough, A.G.; Bransby, M.F.; Hallett, P.D. Biomechanics of nodal, seminal and lateral roots of barley: Effects of diameter, waterlogging and mechanical impedance. Plant Soil 2013, 370, 407–418. [Google Scholar] [CrossRef]
- Lee, J.T.; Yen, L.Z.; Chu, M.Y.; Lin, Y.S.; Chang, C.C.; Lin, R.S.; Chao, K.H.; Lee, M.J. Growth characteristics and anti-wind erosion ability of three tropical fordune pioneer species for sand dune stabilization. Sustainability 2020, 12, 3353. [Google Scholar] [CrossRef] [Green Version]
- Bischetti, G.B.; Chiaradia, E.A.; Simonato, T.; Speziali, B.; Vitali, B.; Vullo, P.; Zocco, A. Root strength and root area ratio of forest species in Lombardy (Northern Italy). Plant Soil 2005, 278, 11–22. [Google Scholar] [CrossRef]
- Boldrin, D.; Leung, A.K.; Bengough, A.G. Effects of root dehydration on biomechanical properties of woody roots of Ulex europaeus. Plant Soil 2018, 431, 347–369. [Google Scholar] [CrossRef] [Green Version]
- Sidorchuk, A.; Sidorchuk, A. Model for estimating gully morphology. In Modelling Soil Erosion, Sediment Transport and Closely Related Hydrological Processes, Proceedings of the International Symposium on Modelling Soil Erosion, Sediment Transport and Closely Related Hydrological Processes, Vienna, Austria, 13–17 July 1998; IAHS Publications: Wallingford, UK, 1998. [Google Scholar]
- Gyssels, G.; Poesen, J.; Bochet, E.; Li, Y. Impact of plant roots on the resistance of soils to erosion by water: A review. Prog. Phys. Geogr. 2005, 29, 189–217. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.C. Miscanthus transmorrisonensis (Hayata). In Flora of Taiwan, 1st ed.; Editorial Committee of Flora of Taiwan: Taipei, Taiwan, 1978; Volume 5, p. 681. [Google Scholar]
- Lin, W.C. Yushania niitakayamensis (Hayata) Keng f. In Flora of Taiwan, 2nd ed.; Editorial Committee of Flora of Taiwan: Taipei, Taiwan, 2000; Volume 5, pp. 651–654. [Google Scholar]
- Truong, P.; Loch, R. Vetiver system for erosion and sediment control. In Proceedings of the 13th International Soil Conservation Organization Conference, Brisbane, QLD, Australia, 1–4 July 2004; pp. 1–6. [Google Scholar]
- Katuwal, S.; Vermang, J.; Cornelis, W.M.; Gabriels, D.; Moldrup, P.; De Jonge, L.W. Effect of root density on erosion and erodibility of a loamy soil under simulated rain. Soil Sci. 2013, 178, 29–36. [Google Scholar] [CrossRef]
- Guo, M.M.; Wang, W.L.; Kang, H.L.; Yang, B. Changes in soil properties and erodibility of gully heads induced by vegetation restoration on the Loess Plateau, China. J. Arid Land 2018, 10, 712–725. [Google Scholar] [CrossRef] [Green Version]
- Nwoke, H.U.; Dike, B.U.; Nwite, S.A.; Nwakwasi, N.L. Relating uprooting resistance to stem basal diameters of plants for erosion mitigation. Int. J. Civ. Eng. Technol. 2016, 7, 129–135. [Google Scholar]
- Roering, J.J.; Schmidt, K.M.; Stock, J.D.; William, E.D.; David, R.M. Shallow landsliding, root reinforcement, and the spatial distribution of trees in the Oregon Coast Range. Can. Geotech. J. 2003, 40, 237–253. [Google Scholar] [CrossRef] [Green Version]
- Tsige, D.; Senadheera, S.; Talema, A. Stability analysis of plant-root-reinforced shallow slopes along mountainous road corridors based on numerical modeling. Geosciences 2020, 10, 19. [Google Scholar] [CrossRef] [Green Version]
- Stokes, A.; Atger, C.; Bengough, A.G.; Fourcaud, T.; Sidle, R.C. Desirable plant root traits for protecting natural and engineered slopes against landslides. Plant Soil 2009, 324, 1–30. [Google Scholar] [CrossRef]
- Genet, M.; Stokes, A.; Salin, F.; Mickovski, S.B. The influence of cellulose content on tensile strength in tree roots. Plant Soil 2005, 278, 1–9. [Google Scholar] [CrossRef]
- Zhang, C.B.; Chen, L.H.; Jiang, J. Why fine tree roots are stronger than thicker roots: The role of cellulose and lignin in relation to slope stability. Geomorphology 2014, 206, 196–202. [Google Scholar] [CrossRef]
- Zhao, X.; Zhao, L.; Li, Q.; Chen, H.; Zhou, H.; Xu, S.; Dong, Q.; Wu, G.; He, Y. Using balance of seasonal herbage supply and demand to inform sustainable grassland management on the Qinghai–Tibetan Plateau. Front. Agric. Sci. Eng. 2018, 5, 1–8. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Properties | Soil |
|---|---|
| pH (water) | 6.21 |
| Conductivity (dS m−1) | 0.11 |
| Organic carbon (g kg−1) | 1.95 |
| Total nitrogen (%) | 0.27 |
| Phosphorus (mg kg−1) | 20 |
| Potassium (mg kg−1) | 78 |
| Calcium (mg kg−1) | 1359 |
| Magnesium (mg kg−1) | 162 |
| Zn (ppm) | 6.87 |
| Mn (ppm) | 143 |
| Fe (ppm) | 350 |
| Cu (ppm) | 1.56 |
| Cd (ppm) | 0.00 |
| Cr (ppm) | 1.00 |
| Ni (ppm) | 1.4 |
| Pb (ppm) | 6.4 |
| Properties | Soil |
|---|---|
| Skeletal fraction (%) | 68 |
| Sand (%) | 17 |
| Silt (%) | 11 |
| Clay (%) | 4 |
| Bulk density (g cm−3) | 1.1 |
| Porosity (%) | 46.2 |
| Particle density (g cm−3) | 2.3 |
| Species | Root Area Ratio (%) | ||||
|---|---|---|---|---|---|
| 0–5 cm | 5–10 cm | 10–15 cm | 15–20 cm | 20–25 cm | |
| Miscanthus transmorrisonensis | 0.17 ± 0.04 a | 0.08 ± 0.03 b | 0.02 ± 0.01 b | 0.01 ± 0.01 a | 0.00 ± 0.00 a |
| Yushania niitakayamensis | 0.26 ± 0.02 a | 1.13 ± 0.20 a | 0.17 ± 0.04 a | 0.04 ± 0.02 a | 0.00 ± 0.00 a |
| Growth Characteristics | M. transmorrisonensis | Y. niitakayamensis | t Value |
|---|---|---|---|
| RCD (mm) | 4.07 ± 0.21 b | 9.17 ± 0.58 a | 8.228 *** |
| RT | 1371.65 ± 130.28 a | 1125.78 ± 175.2 a | −1.118 |
| TRL (cm) | 332.08 ± 73.36 b | 1369.6 ± 121.14 a | −7.326 *** |
| RB (g) | 5.84 ± 1.12 b | 13.33 ± 2.65 a | 3.066 ** |
| SB (g) | 3.73 ± 0.39 b | 38.56 ± 3.59 a | 9.653 *** |
| RD (kg m−3) | 2.33 ± 0.45 b | 5.64 ± 1.39 a | 2.263 * |
| RLD (km m−3) | 1.2 ± 0.27 b | 5.48 ± 0.48 a | −7.705 *** |
| RSA (cm2) | 333.68 ± 64.12 b | 1271.83 ± 161.03 a | −5.412 *** |
| RTD (g cm−3) | 0.24 ± 0.02 a | 0.35 ± 0.06 a | 1.773 |
| RV (cm3) | 26.47 ± 5.03 b | 45.23 ± 3.15 a | 1.855 * |
| SRL (m g−1) | 0.33 ± 0.1 b | 3.48 ± 0.51 a | −6.052 *** |
| Root Anchorage Ability | M. transmorrisonensis | Y. niitakayamensis | t-Value |
|---|---|---|---|
| Ultimate uprooting force (kN) | 0.11 ± 0.02 b | 0.8 ± 0.09 a | 7.607 *** |
| Morphological Traits | Species | Regression Equation | R2 | p |
|---|---|---|---|---|
| RCD (mm) | M. transmorrisonensis Y. niitakayamensis | Ur = 0.05RCD − 0.093 Ur = 0.141RCD − 0.509 | 0.762 ∗∗ 0.8218 ∗∗ | 0.01 0.01 |
| RSA (cm2) | M. transmorrisonensis Y. niitakayamensis | Ur = −0.0004RSA + 0.05 Ur = −0.001RSA + 0.407 | 0.738 ∗ 0.803 ∗ | 0.023 0.034 |
| Species | Regression Equation | R2 | p | VIF |
|---|---|---|---|---|
| M. transmorrisonensis | Ur = 0.042RCD + 0.00028RSA − 0.097 | 0.715 ∗∗ | 0.002 | 1.139 |
| Y.niitakayamensis | Ur = 0.122RCD + 0.00RSA − 0.581 | 0.85 ∗∗∗ | 0.0004 | 1.078 |
| Parameters | M. transmorrisonensis | Y. niitakayamensis | t Value |
|---|---|---|---|
| Root diameters (mm) | 0.71 ± 0.06 b | 1.73 ± 0.17 a | −5.608 *** |
| Tensile resistance force (N) | 15.30 ± 1.94 b | 61.25 ± 8.96 a | −5.012 *** |
| Tensile strength (MPa) | 408.55 ± 22.71 a | 24.96 ± 1.13 b | 16.871 *** |
| Slope (°) | Soil Loss Amount (g min−1) | ANOVA (p) | ||
|---|---|---|---|---|
| Miscanthus transmorrisonensis | Yushania niitakayamensis | Bare Soil | ||
| 2.86 | 49 ± 10.02 b | 7.73 ± 1.15 c | 224.5 ± 41.5 a | 0.000 *** |
| 16.7 | 267.67 ± 35.45 b | 82.25 ± 12.61 c | 666.1 ± 66.3 a | 0.000 *** |
| Slope (°) | Relative Soil Detachment Rate (%) | p | |
|---|---|---|---|
| Miscanthus transmorrisonensis | Yushania niitakayamensis | ||
| 2.86 | 21.83 ± 4.46 a | 3.44 ± 0.51 b | 0.005 ** |
| 16.7 | 40.19 ± 5.32 a | 12.35 ± 1.89 b | 0.003 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.-T.; Tsai, S.-M.; Wu, Y.-J.; Lin, Y.-S.; Chu, M.-Y.; Lee, M.-J. Root Characteristics and Water Erosion-Reducing Ability of Alpine Silver Grass and Yushan Cane for Alpine Grassland Soil Conservation. Sustainability 2021, 13, 7633. https://doi.org/10.3390/su13147633
Lee J-T, Tsai S-M, Wu Y-J, Lin Y-S, Chu M-Y, Lee M-J. Root Characteristics and Water Erosion-Reducing Ability of Alpine Silver Grass and Yushan Cane for Alpine Grassland Soil Conservation. Sustainability. 2021; 13(14):7633. https://doi.org/10.3390/su13147633
Chicago/Turabian StyleLee, Jung-Tai, Shun-Ming Tsai, Yu-Jie Wu, Yu-Syuan Lin, Ming-Yang Chu, and Ming-Jen Lee. 2021. "Root Characteristics and Water Erosion-Reducing Ability of Alpine Silver Grass and Yushan Cane for Alpine Grassland Soil Conservation" Sustainability 13, no. 14: 7633. https://doi.org/10.3390/su13147633
APA StyleLee, J.-T., Tsai, S.-M., Wu, Y.-J., Lin, Y.-S., Chu, M.-Y., & Lee, M.-J. (2021). Root Characteristics and Water Erosion-Reducing Ability of Alpine Silver Grass and Yushan Cane for Alpine Grassland Soil Conservation. Sustainability, 13(14), 7633. https://doi.org/10.3390/su13147633