CityScapeLab Berlin: A Research Platform for Untangling Urbanization Effects on Biodiversity

 ,
,
Abstract
1. Introduction
- exploring the effects of urbanization and rapid transitions in urban land-use patterns on biodiversity and ecosystem functioning at different spatial and temporal scales;
- developing and testing theories on the intersection between urbanization and biodiversity;
- supporting policies on the integration of biodiversity and the associated ecosystem services into urban landscapes, as a contribution to sustainable, livable, and resilient cities.
2. Fundamental Approach of the CityScapeLab Berlin
2.1. Model Ecosystem
- ■
- ■
- Urban grassland patches are usually subject to a varying intensity of anthropogenic recreational activities associated with trampling, soil disturbance, or nutrient influxes—or they exist without such interferences, e.g., in conservation areas;
- As a low productivity ecosystem, urban dry grassland is particularly sensitive to nutrient influxes from the urban matrix or from local human activities [63];
- Urban grassland is often invaded by alien plant species [57,67] as is the urban dry grassland in Berlin, with an average proportion of 25% of alien species [68]. Grassland, as a shared habitat of many native and alien species, allows analyses of multiple biotic interactions (e.g., native‒alien, plant‒animal, or belowground‒aboveground interactions);
- Urban grassland has a limited structural heterogeneity due to the prevalence of grasses and herbs and a similar management intensity on low productivity sites—dry grassland in Berlin, for example, is usually mown up to two times a year, or less;
- Urban grassland often spans a large gradient of ecological novelty, from near-natural sites to designed greenspaces to novel ecosystems on vacant land [66], encompassing sites with natural and anthropogenic soils and including patches with different land-use legacies and current or historical habitat connectivity [61].
2.2. Bridging Spatial Scales
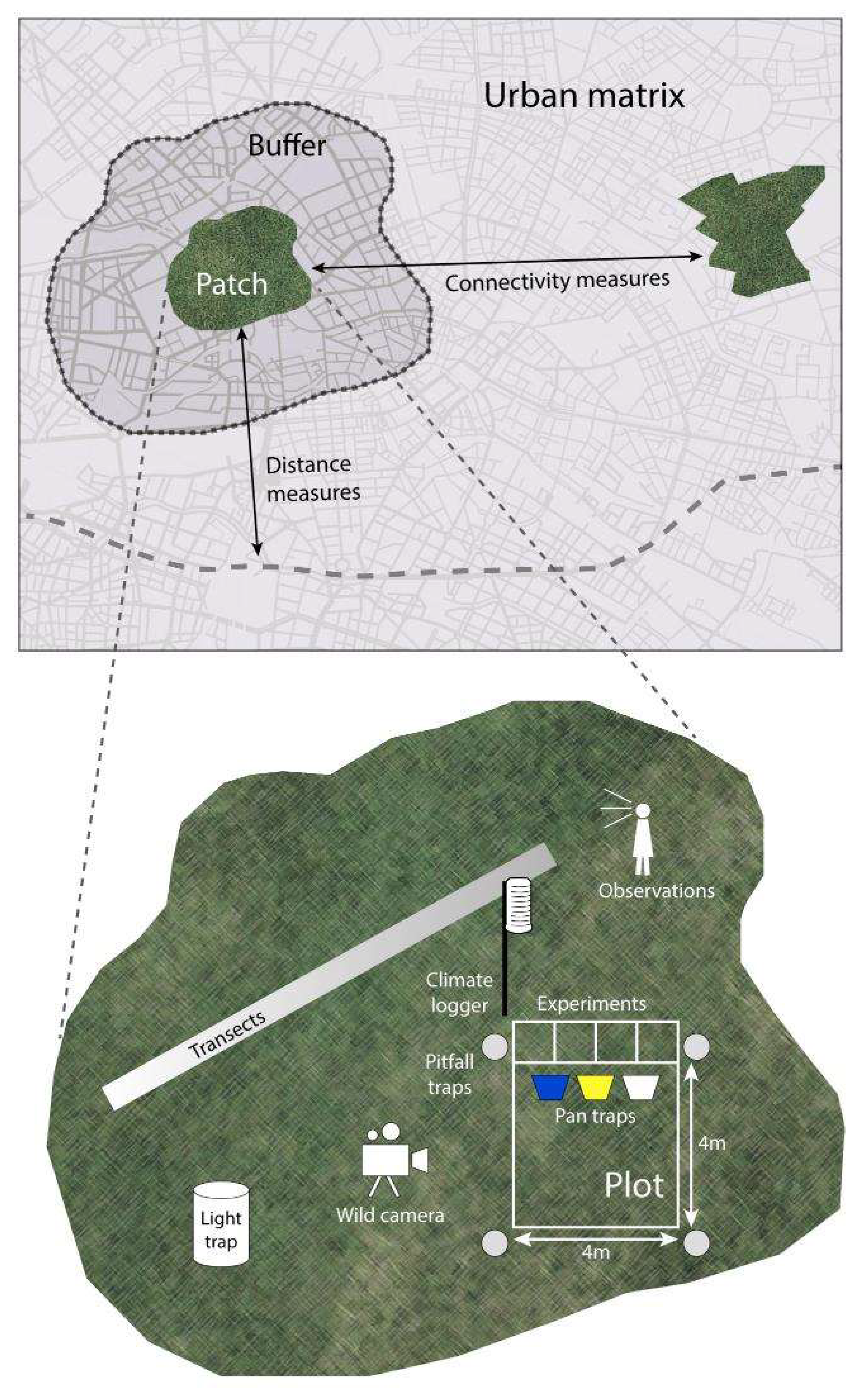
- The basic survey units of the CityScapeLab are patches of the selected model ecosystem, i.e., grassland patches. These share a rather homogeneous vegetation structure but usually differ in size, adjacency to other, and connectivity to similar ecosystems. A minimum patch size of 100 m² still ensures that biodiversity measurements of taxa of different spatial range can be linked to each other and the grassland patch. For example, measures of bat activity from an automated bat recorder usually cover a radius between 12 and 75 m but can still be reasonably related to light traps of nocturnal insects of the same patch. Moreover, this patch-centered approach allows for analyses of biotope transitions at the edge to surrounding habitats and different land uses;
- Each urban grassland patch encompasses one randomly located plot with a standardized size (4 × 4 m) for sampling environmental variables and some taxa at the plot scale (e.g., plants and arthropods). Other taxa, including grasshoppers, can be sampled along transects at the patch level, spatially linked with the plots or by nearby exposed camera traps;
- Next to the plot and within the patch, an area is foreseen where destructive investigations (e.g., biomass measurements e.g., [63]) or experiments can be performed (e.g., plant-pollinator interactions [75]). While the plots remain undisturbed, results from the adjacent experimental area can be related to the environmental variables measured at the plot or patch level;
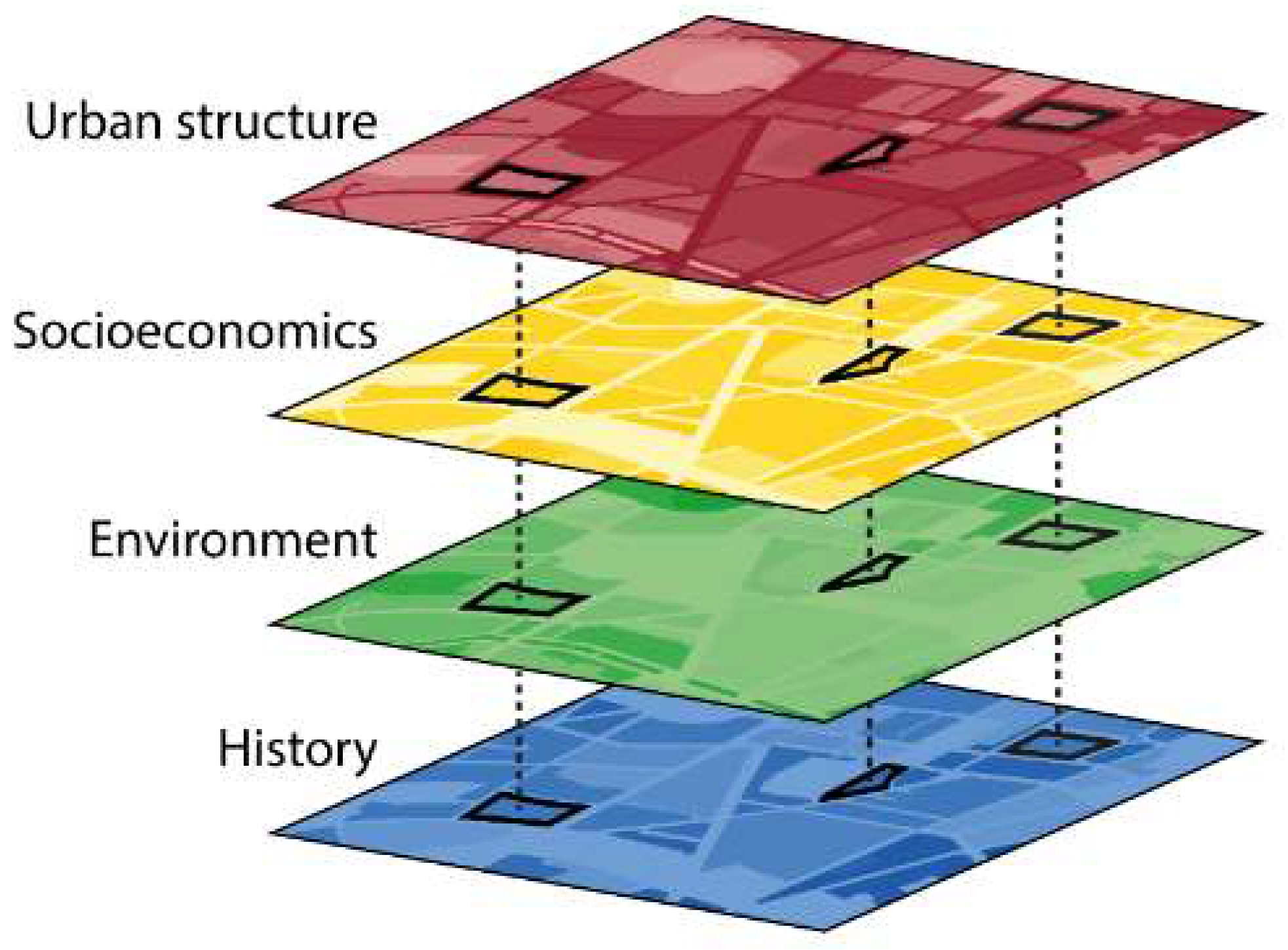
- A range of environmental and socioeconomic data is available for spatial buffers in the surroundings of the patches thus allowing us to elucidate relationships between the urban matrix and the biodiversity patterns at the plot or patch scale;
- To ensure analyses of functional spatial relations between the plot or patch scale, the city area, and the surrounding countryside, the research platform not only covers the entire metropolitan region of Berlin but also includes typical rural landscapes within a buffer of approximately ten kilometers beyond the administrative borders of Berlin (Figure 3). Results from the biodiversity analyses, thus, can be synthesized, or differentiated, on a regional scale. Citywide spatial data sets on all significant environmental and urbanization effects enable the linking of the local and supra-local level to regional drivers of biodiversity;
- In parallel, biodiversity analyses from the study sites can be related to data at higher spatial scales. For few taxa, grid-based data exist in Berlin (e.g., for vascular plants [76]), while total species inventories are available for a broad range of plant and animal groups on the city scale. As these are often coupled with Red Lists of species endangered or extinct in Berlin (Table 1), biodiversity responses to urbanization can be specified for species of particular conservation concern.
2.3. Bridging Temporal Scales
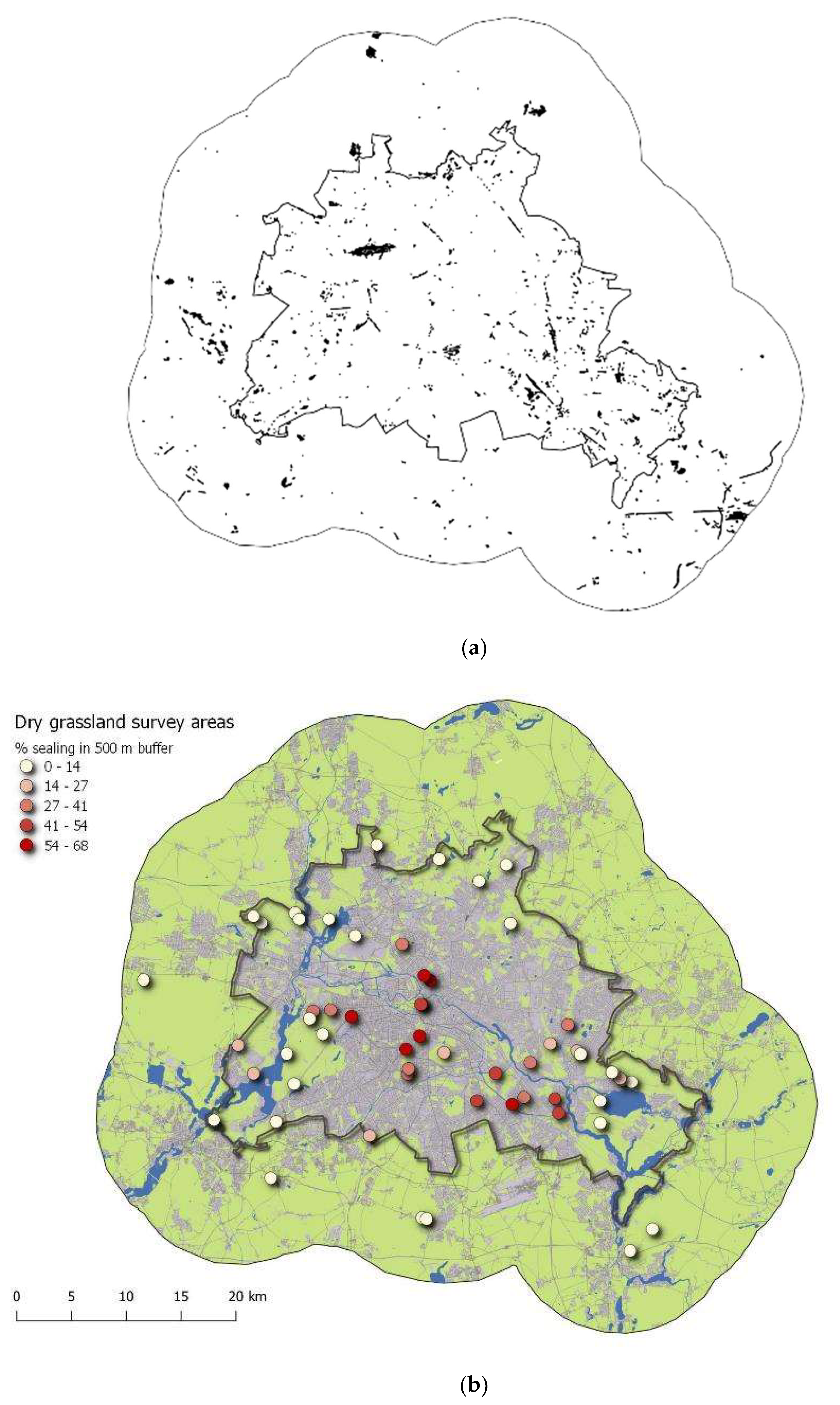
- For the total area of Berlin, the historical land use for the settlement area, woodland, and grassland has been traced back from analyses of georeferenced historical maps from three periods of time since the beginning of the 19th century. This allowed for the incorporation of both “old” grassland patches (established more than 100 years ago) and “new” grassland patches (established since 1945) into the set of survey areas. This enables testing for the relevance of land-use continuity and for habitat connectivity at different periods of time for current biodiversity patterns;
- All study areas were established as permanent monitoring sites, with a plot precisely mapped at centimeter accuracy, allowing for the temporal dynamics in biodiversity patterns and processes to be captured in the future. Some of the established monitoring plots overlap with previous vegetation surveys, dating back between 10 and 30 years. These data well support historical comparisons because they were georeferenced at sub-meter accuracy;
- Ultimately, the survey patches also stretch along a gradient of ecological novelty, both in terms of biotic novelty and abiotic novelty. As illustrated by Figure 4, the patches span from sites with near-natural soils (e.g., grassland in a forest or agricultural context), to areas with anthropogenic soils (e.g., in historical parks), to novel sites with artificial soils, such as in transportation corridors (e.g., motorway embankments), or vacant land (e.g., abandoned rail yards). This allows for the unraveling of the relationships between different levels of ecological novelty and ecological and evolutionary patterns in urban environments—with significant implications for conservation strategies in the face of global change [80,81].
2.4. Bridging Taxonomical and Functional Groups
- The focus on a standardized ecosystem type allows for the identification of the relative importance of urbanization drivers versus local environmental conditions in shaping community assembly and modulating biotic interactions;
- The multi-scale approach (see Section 2.2) provides a flexible research infrastructure to incorporate taxa with starkly varying activity radii into the analyses. Thus, different taxa representing different trophic levels can be analyzed with regard to urbanization responses in terms of biodiversity measures and biotic interactions;
- Dry grassland as a species-rich model ecosystem is ideal for investigating a large variety of interactions as most trophic levels are represented by different taxonomic groups with a potentially large number of native and alien species.
2.5. Integrating the Human Dimension
- The study areas are intentionally not fenced in to allow analyses of a wide range of everyday interactions of city dwellers with the grassland;
- On a neighborhood level, relationships between biodiversity patterns and human influences can be elucidated by incorporating socioeconomic biodiversity predictors. High-resolution spatial data on socioeconomic features are available from the Berlin Social Atlas on the basis of neighborhoods that cover on average 7500 inhabitants. Further spatial data on environmental justice make it possible to address citizens not only as drivers of ecological change but also as those affected by environmental pressures in the city;
- Free access to the research sites also allows the application of methods from the social sciences, for example, observations of the behavior of humans staying on or passing through the study plots [102].
3. Study Region and Linkages to Urban Biodiversity Research
3.1. Berlin as a Model of a Metropolitan Region
3.2. Links to Research Traditions in Berlin
4. Methodological Approaches
4.1. Dry Grassland Survey Areas
4.2. Plot design for Multi-Taxon Analyses
4.3. Analyses of Local Environmental Conditions
4.3.1. Microclimate
4.3.2. Sky View Factor
4.3.3. Chemical Soil Properties
4.3.4. Vegetation
4.3.5. Human Interferences
4.3.6. Light Pollution
4.4. Establishment of a Sensor Network
4.5. Analyses of the Urban Matrix
4.5.1. Urban Structure
4.5.2. Environmental Dimension
4.5.3. Socioeconomic Dimension
4.5.4. Historical Dimension
5. Outlook
- Within the already established model ecosystem (i.e., grassland), the covered urbanization gradient can be extended toward more extreme settings in terms of aridity or spatial isolation (e.g., dry grassland on roof tops, road verges, and block courtyards);
- The suite of model ecosystems can be expanded successively, e.g., with urban ponds to support analyses on the role of urbanization for aquatic biodiversity patterns. A further extension might cover emerging urban woodlands as a model system that allows insights into community assembly and ecosystem functioning in “wild” ecosystems that largely develop without direct human interferences in urban regions [148];
- The approach could also be extended toward the other end of the urbanization gradient by including very remote patches of the same model ecosystem(s) outside the Berlin metropolitan area as control sites without minor urban impact;
- Results generated from the CityScapeLab Berlin could be a reference for testing theories or generalisations through comparisons with other cities at European or global scales.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ellis, E.C. Ecology in an anthropogenic biosphere. Ecol. Monogr. 2015, 85, 287–331. [Google Scholar] [CrossRef]
- Haase, D.; Larondelle, N.; Andersson, E.; Artmann, M.; Borgström, S.; Breuste, J.; Gomez-Baggethun, E.; Gren, A.; Hamstead, Z.; Hansen, R.; et al. A Quantitative Review of Urban Ecosystem Service Assessments: Concepts, Models, and Implementation. Ambio 2014, 43, 413–433. [Google Scholar] [CrossRef]
- Hartig, T.; Mitchell, R.; de Vries, S.; Frumkin, H. Nature and Health. Annu. Rev. Publ. Health 2014, 35, 207–228. [Google Scholar] [CrossRef]
- Schebella, M.F.; Weber, D.; Schultz, L.; Weinstein, P. The Wellbeing Benefits Associated with Perceived and Measured Biodiversity in Australian Urban Green Spaces. Sustainability 2019, 11, 802. [Google Scholar] [CrossRef]
- Güneralp, B.; Seto, K.C. Futures of global urban expansion: Uncertainties and implications for biodiversity conservation. Environ. Res. Lett. 2013, 8, 014025. [Google Scholar] [CrossRef]
- McKinney, M.L. Urbanization, biodiversity, and conservation. Bioscience 2002, 52, 883–890. [Google Scholar] [CrossRef]
- Aronson, M.F.J.; La Sorte, F.A.; Nilon, C.H.; Katti, M.; Goddard, M.A.; Lepczyk, C.A.; Warren, P.S.; Williams, N.S.G.; Cilliers, S.; Clarkson, B.; et al. A global analysis of the impacts of urbanization on bird and plant diversity reveals key anthropogenic drivers. Proc. R. Soc. B Biol. Sci. 2014, 281, 2013330. [Google Scholar] [CrossRef]
- Hall, D.M.; Camilo, G.R.; Tonietto, R.K.; Ollerton, J.; Ahrne, K.; Arduser, M.; Ascher, J.S.; Baldock, K.C.R.; Fowler, R.; Frankie, G.; et al. The city as a refuge for insect pollinators. Conserv. Biol. 2017, 31, 24–29. [Google Scholar] [CrossRef]
- Ives, C.D.; Lentini, P.E.; Threlfall, C.G.; Ikin, K.; Shanahan, D.F.; Garrard, G.E.; Bekessy, S.A.; Fuller, R.A.; Mumaw, L.; Rayner, L.; et al. Cities are hotspots for threatened species. Glob. Ecol. Biogeogr. 2016, 25, 117–126. [Google Scholar] [CrossRef]
- Kowarik, I.; von der Lippe, M. Plant population success across urban ecosystems: A framework to inform biodiversity conservation in cities. J. Appl. Ecol. 2018, 55, 2354–2361. [Google Scholar] [CrossRef]
- Soanes, K.; Lentini, P.E. When cities are the last chance for saving species. Front. Ecol. Environ. 2019, 17, 225–231. [Google Scholar] [CrossRef]
- Aronson, M.F.J.; Lepczyk, C.A.; Evans, K.L.; Goddard, M.A.; Lerman, S.B.; MacIvor, J.S.; Nilon, C.H.; Vargo, T. Biodiversity in the city: Key challenges for urban green space management. Front. Ecol. Environ. 2017, 15, 189–196. [Google Scholar] [CrossRef]
- Nilon, C.H.; Aronson, M.F.J.; Cilliers, S.S.; Dobbs, C.; Frazee, L.J.; Goddard, M.A.; O’Neill, K.M.; Roberts, D.; Stander, E.K.; Werner, P.; et al. Planning for the Future of Urban Biodiversity: A Global Review of City-Scale Initiatives. Bioscience 2017, 67, 331–341. [Google Scholar] [CrossRef]
- Parris, K.M.; Amati, M.; Bekessy, S.A.; Dagenais, D.; Fryd, O.; Hahs, A.K.; Hes, D.; Imberger, S.J.; Livesley, S.J.; Marshall, A.J.; et al. The seven lamps of planning for biodiversity in the city. Cities 2018, 83, 44–53. [Google Scholar] [CrossRef]
- Fuller, R.A.; Irvine, K.N.; Devine-Wright, P.; Warren, P.H.; Gaston, K.J. Psychological benefits of greenspace increase with biodiversity. Biol. Lett. 2007, 3, 390–394. [Google Scholar] [CrossRef]
- Schwarz, N.; Moretti, M.; Bugalho, M.N.; Davies, Z.G.; Haase, D.; Hack, J.; Hof, A.; Melero, Y.; Pett, T.J.; Knapp, S. Understanding biodiversity-ecosystem service relationships in urban areas: A comprehensive literature review. Ecosyst. Serv. 2017, 27, 161–171. [Google Scholar] [CrossRef]
- Fischer, L.K.; Honold, J.; Cvejic, R.; Delshammar, T.; Hilbert, S.; Lafortezza, R.; Nastran, M.; Nielsen, A.B.; Pintar, M.; van der Jagt, A.P.N.; et al. Beyond green: Broad support for biodiversity in multicultural European cities. Glob. Environ. Chang. 2018, 49, 35–45. [Google Scholar] [CrossRef]
- Tzoulas, K.; Korpela, K.; Venn, S.; Yli-Pelkonen, V.; Kaźmierczak, A.; Niemela, J.; James, P. Promoting ecosystem and human health in urban areas using Green Infrastructure: A literature review. Landsc. Urban Plan. 2007, 81, 167–178. [Google Scholar] [CrossRef]
- Ramalho, C.E.; Hobbs, R.J. Time for a change: Dynamic urban ecology. Trends Ecol. Evol. 2012, 27, 179–188. [Google Scholar] [CrossRef]
- Pickett, S.T.A.; Cadenasso, M.L.; Grove, J.M.; Nilon, C.H.; Pouyat, R.V.; Zipperer, W.C.; Costanza, R. Urban ecological systems: Linking terrestrial ecological, physical, and socioeconomic components of metropolitan areas. Annu. Rev. Ecol. Syst. 2001, 32, 127–157. [Google Scholar] [CrossRef]
- Liu, J.G.; Dietz, T.; Carpenter, S.R.; Alberti, M.; Folke, C.; Moran, E.; Pell, A.N.; Deadman, P.; Kratz, T.; Lubchenco, J.; et al. Complexity of coupled human and natural systems. Science 2007, 317, 1513–1516. [Google Scholar] [CrossRef] [PubMed]
- Alberti, M.; Marzluff, J.M.; Shulenberger, E.; Bradley, G.; Ryan, C.; Zumbrunnen, C. Integrating humans into ecology: Opportunities and challenges for studying urban ecosystems. Bioscience 2003, 53, 1169–1179. [Google Scholar] [CrossRef]
- Grimm, N.B.; Faeth, S.H.; Golubiewski, N.E.; Redman, C.L.; Wu, J.G.; Bai, X.M.; Briggs, J.M. Global change and the ecology of cities. Science 2008, 319, 756–760. [Google Scholar] [CrossRef] [PubMed]
- Kühn, I.; Brandl, R.; Klotz, S. The flora of German cities is naturally species rich. Evol. Ecol. Res. 2004, 6, 749–764. [Google Scholar]
- Godefroid, S.; Koedam, N. Distribution pattern of the flora in a peri-urban forest: An effect of the city-forest ecotone. Landsc. Urban Plan. 2003, 65, 169–185. [Google Scholar] [CrossRef]
- Lundholm, J.T.; Richardson, P.J. Habitat analogues for reconciliation ecology in urban and industrial environments. J. Appl. Ecol. 2010, 47, 966–975. [Google Scholar] [CrossRef]
- Kowarik, I. Novel urban ecosystems, biodiversity, and conservation. Environ. Pollut. 2011, 159, 1974–1983. [Google Scholar] [CrossRef]
- Linkola, K. Studien über den Einfluss der Kultur auf die Flora in den Gegenden nördlich vom Ladogasee. Acta Soc. Fauna Flora Fenn. 1916, 45, 1–429. [Google Scholar]
- Pyšek, P. Alien and native species in Central European urban floras: A quantitative comparison. J. Biogeogr. 1998, 25, 155–163. [Google Scholar] [CrossRef]
- Čeplová, N.; Kalusová, V.; Lososová, Z. Effects of settlement size, urban heat island and habitat type on urban plant biodiversity. Landsc. Urban Plan. 2017, 159, 15–22. [Google Scholar] [CrossRef]
- Kunick, W. Zonierung des Stadtgebietes von Berlin West. Ergebnisse Floristischer Untersuchungen; Landschaftsentwicklung und Umweltforschung: Berlin, Germany, 1982; Volume 14, p. 304. [Google Scholar]
- Sukopp, H.; Weiler, S. Biotope mapping and nature monservation strategies in urban areas of the Federal Republic of Germany. Landsc. Urban Plan. 1988, 15, 39–58. [Google Scholar] [CrossRef]
- Löfvenhaft, K.; Björn, C.; Ihse, M. Biotope patterns in urban areas: A conceptual model integrating biodiversity issues in spatial planning. Landsc. Urban Plan. 2002, 58, 223–240. [Google Scholar] [CrossRef]
- Lososová, Z.; Chytrý, M.; Tichý, L.; Danihelka, J.; Fajmon, K.; Hájek, O.; Kintrová, K.; Kühn, I.; Laníková, D.; Otýpková, Z.; et al. Native and alien floras in urban habitats: A comparison across 32 cities of central Europe. Glob. Ecol. Biogeogr. 2012, 21, 545–555. [Google Scholar] [CrossRef]
- McDonnell, M.J.; Pickett, S.T.A. Ecosystem Structure and Function Along Urban Rural Gradients—An Unexploited Opportunity for Ecology. Ecology 1990, 71, 1232–1237. [Google Scholar] [CrossRef]
- McDonnell, M.J.; Hahs, A.K. The use of gradient analysis studies in advancing our understanding of the ecology of urbanizing landscapes: Current status and future directions. Landsc. Ecol. 2008, 23, 1143–1155. [Google Scholar] [CrossRef]
- Niemela, J.; Kotze, D.J. Carabid beetle assemblages along urban to rural gradients: A review. Landsc. Urban Plan. 2009, 92, 65–71. [Google Scholar] [CrossRef]
- von der Lippe, M.; Kowarik, I. Do cities export biodiversity? Traffic as dispersal vector across urban-rural gradients. Divers. Distrib. 2008, 14, 18–25. [Google Scholar] [CrossRef]
- Niggemann, M.; Jetzkowitz, J.; Brunzel, S.; Wichmann, M.C.; Bialozyt, R. Distribution patterns of plants explained by human movement behaviour. Ecol. Model. 2009, 220, 1339–1346. [Google Scholar] [CrossRef]
- Hahs, A.K.; McDonnell, M.J.; McCarthy, M.A.; Vesk, P.A.; Corlett, R.T.; Norton, B.A.; Clemants, S.E.; Duncan, R.P.; Thompson, K.; Schwartz, M.W.; et al. A global synthesis of plant extinction rates in urban areas. Ecol. Lett. 2009, 12, 1165–1173. [Google Scholar] [CrossRef]
- Knapp, S.; Kühn, I.; Stolle, J.; Klotz, S. Changes in the functional composition of a Central European urban flora over three centuries. Perspect. Plant Ecol. 2010, 12, 235–244. [Google Scholar] [CrossRef]
- Chocholoušková, Z.; Pyšek, P. Changes in composition and structure of urban flora over 120 years: A case study of the city of Plzeň. Flora 2003, 198, 366–376. [Google Scholar] [CrossRef]
- Williams, N.S.G.; Hahs, A.K.; Vesk, P.A. Urbanisation, plant traits and the composition of urban floras. Perspect. Plant Ecol. 2015, 17, 78–86. [Google Scholar] [CrossRef]
- Buchholz, S.; Gathof, A.K.; Grossmann, A.J.; Kowarik, I.; Fischer, L.K. Wild bees in urban grasslands: Urbanisation, functional diversity and species traits. Landsc. Urban Plan. 2020, 196, 103731. [Google Scholar] [CrossRef]
- Shwartz, A.; Turbe, A.; Julliard, R.; Simon, L.; Prevot, A.C. Outstanding challenges for urban conservation research and action. Glob. Environ. Chang. 2014, 28, 39–49. [Google Scholar] [CrossRef]
- Beninde, J.; Veith, M.; Hochkirch, A. Biodiversity in cities needs space: A meta-analysis of factors determining intra-urban biodiversity variation. Ecol. Lett. 2015, 18, 581–592. [Google Scholar] [CrossRef]
- Pickett, S.T.A.; Cadenasso, M.L.; Grove, J.M.; Irwin, E.G.; Rosi, E.J. (Eds.) Science for the Sustainable City: Empirical Insights from the Baltimore School of Urban Ecology; Yale University Press: New Haven, CT, USA, 2019. [Google Scholar]
- Cadenasso, M.L.; Pickett, S.T.A.; Schwarz, K. Spatial heterogeneity in urban ecosystems: Reconceptualizing land cover and a framework for classification. Front. Ecol. Environ. 2007, 5, 80–88. [Google Scholar] [CrossRef]
- Malkinson, D.; Kopel, D.; Wittenberg, L. From rural-urban gradients to patch—Matrix frameworks: Plant diversity patterns in urban landscapes. Landsc. Urban Plan. 2018, 169, 260–268. [Google Scholar] [CrossRef]
- Godefroid, S.; Monbaliu, D.; Koedam, N. The role of soil and microclimatic variables in the distribution patterns of urban wasteland flora in Brussels, Belgium. Landsc. Urban Plan. 2007, 80, 45–55. [Google Scholar] [CrossRef]
- Westermann, J.R.; von der Lippe, M.; Kowarik, I. Seed traits, landscape and environmental parameters as predictors of species occurrence in fragmented urban railway habitats. Basic Appl. Ecol. 2011, 12, 29–37. [Google Scholar] [CrossRef]
- Anderson, E.C.; Minor, E.S. Assessing social and biophysical drivers of spontaneous plant diversity and structure in urban vacant lots. Sci. Total Environ. 2019, 653, 1272–1281. [Google Scholar] [CrossRef]
- Klaus, V.H. Urban Grassland Restoration: A Neglected Opportunity for Biodiversity Conservation. Restor. Ecol. 2013, 21, 665–669. [Google Scholar] [CrossRef]
- Fischer, L.K.; von der Lippe, M.; Kowarik, I. Urban land use types contribute to grassland conservation: The example of Berlin. Urban Urban Gree 2013, 12, 263–272. [Google Scholar] [CrossRef]
- van der Walt, L.; Cilliers, S.S.; Kellner, K.; du Toit, M.J.; Tongway, D. To what extent does urbanisation affect fragmented grassland functioning? J. Environ. Manag. 2015, 151, 517–530. [Google Scholar] [CrossRef] [PubMed]
- Filibeck, G.; Petrella, P.; Cornelini, P. All ecosystems look messy, but some more so than others: A case-study on the management and acceptance of Mediterranean urban grasslands. Urban Urban Gree 2016, 15, 32–39. [Google Scholar] [CrossRef]
- Zeeman, B.J.; Minden, V.; Morgan, J.W. Non-native plant cover and functional trait composition of urban temperate grasslands in relation to local- and landscape-scale road density. Biol. Invasions 2018, 20, 3025–3036. [Google Scholar] [CrossRef]
- Norton, B.A.; Bending, G.D.; Clark, R.; Corstanje, R.; Dunnett, N.; Evans, K.L.; Grafius, D.R.; Gravestock, E.; Grice, S.M.; Harris, J.A.; et al. Urban meadows as an alternative to short mown grassland: Effects of composition and height on biodiversity. Ecol. Appl. 2019, 29, e01946. [Google Scholar] [CrossRef] [PubMed]
- Maurer, U.; Peschel, T.; Schmitz, S. The flora of selected urban land-use types in Berlin and Potsdam with regard to nature conservation in cities. Landsc. Urban Plan. 2000, 46, 209–215. [Google Scholar] [CrossRef]
- Williams, N.S.G.; McDonnell, M.J.; Seager, E.J. Factors influencing the loss of an endangered ecosystem in an urbanising landscape: A case study of native grasslands from Melbourne, Australia. Landsc. Urban Plan. 2005, 71, 35–49. [Google Scholar] [CrossRef]
- du Toit, M.J.; Kotze, D.J.; Cilliers, S.S. Landscape history, time lags and drivers of change: Urban natural grassland remnants in Potchefstroom, South Africa. Landsc. Ecol. 2016, 31, 2133–2150. [Google Scholar] [CrossRef]
- Zeeman, B.J.; McDonnell, M.J.; Kendal, D.; Morgan, J.W. Biotic homogenization in an increasingly urbanized temperate grassland ecosystem. J. Veg. Sci. 2017, 28, 550–561. [Google Scholar] [CrossRef]
- Onandia, G.; Schittko, C.; Ryo, M.; Bernard-Verdier, M.; Heger, T.; Joshi, J.; Kowarik, I.; Gessler, A. Ecosystem functioning in urban grasslands: The role of biodiversity, plant invasions and urbanization. PLoS ONE 2019, 14, e0225438. [Google Scholar] [CrossRef] [PubMed]
- Cilliers, S.S.; Müller, N.; Drewes, E. Overview on urban nature conservation: Situation in the western-grassland biome of South Africa. Urban Urban Gree 2004, 3, 49–62. [Google Scholar] [CrossRef]
- Threlfall, C.G.; Walker, K.; Williams, N.S.G.; Hahs, A.K.; Mata, L.; Stork, N.; Livesley, S.J. The conservation value of urban green space habitats for Australian native bee communities. Biol. Conserv. 2015, 187, 240–248. [Google Scholar] [CrossRef]
- Planchuelo, G.; von der Lippe, M.; Kowarik, I. Untangling the role of urban ecosystems as habitats for endangered plant species. Landsc. Urban Plan. 2019, 189, 320–334. [Google Scholar] [CrossRef]
- Cilliers, S.S.; Williams, N.S.G.; Barnard, F.J. Patterns of exotic plant invasions in fragmented urban and rural grasslands across continents. Landsc. Ecol. 2008, 23, 1243–1256. [Google Scholar] [CrossRef]
- Kowarik, I. On the Role of Alien Species in Urban Flora and Vegetation. In Urban Ecology: An International Perspective on the Interaction Between Humans and Nature; Marzluff, J.M., Shulenberger, E., Endlicher, W., Alberti, M., Bradley, G., Ryan, C., Simon, U., ZumBrunnen, C., Eds.; Springer: Boston, MA, USA, 2008; pp. 321–338. [Google Scholar]
- Alberti, M. The effects of urban patterns on ecosystem function. Int. Reg. Sci. Rev. 2005, 28, 168–192. [Google Scholar] [CrossRef]
- Hahs, A.K.; McDonnell, M.J. Selecting independent measures to quantify Melbourne’s urban-rural gradient. Landsc. Urban Plan. 2006, 78, 435–448. [Google Scholar] [CrossRef]
- Seress, G.; Lipovits, A.; Bokony, V.; Czuni, L. Quantifying the urban gradient: A practical method for broad measurements. Landsc. Urban Plan. 2014, 131, 42–50. [Google Scholar] [CrossRef]
- Johnson, A.L.; Tauzer, E.C.; Swan, C.M. Human legacies differentially organize functional and phylogenetic diversity of urban herbaceous plant communities at multiple spatial scales. Appl. Veg. Sci. 2015, 18, 513–527. [Google Scholar] [CrossRef]
- Stillfried, M.; Fickel, J.; Börner, K.; Wittstatt, U.; Heddergott, M.; Ortmann, S.; Kramer-Schadt, S.; Frantz, A.C. Do cities represent sources, sinks or isolated islands for urban wild boar population structure? J. Appl. Ecol. 2017, 54, 272–281. [Google Scholar] [CrossRef]
- Piano, E.; Souffreau, C.; Merckx, T.; Baardsen, L.F.; Backeljau, T.; Bonte, D.; Brans, K.I.; Cours, M.; Dahirel, M.; Debortoli, N.; et al. Urbanization drives cross-taxon declines in abundance and diversity at multiple spatial scales. Glob. Chang. Biol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Buchholz, S.; Kowarik, I. Urbanisation modulates plant-pollinator interactions in invasive vs. native plant species. Sci. Rep. 2019, 9, 6375. [Google Scholar] [CrossRef] [PubMed]
- Seitz, B.; Ristow, M.; Prasse, R.; Machatzi, B.; Klemm, G.; Böcker, R.; Sukopp, H. Der Berliner Florenatlas; Botanischer Verein von Berlin und Brandenburg: Rangsdorf, Germany, 2012; Beiheft 7; p. 533. [Google Scholar]
- Aey, W. Historical approaches to urban ecology. In Urban Ecology: Plants and Plant Communities in Urban Environments; Sukopp, H., Hejný, S., Kowarik, I., Eds.; SPB Academic Publishing: The Hague, the Netherlands, 1990; pp. 113–129. [Google Scholar]
- Lepczyk, C.A.; Aronson, M.F.J.; Evans, K.L.; Goddard, M.A.; Lerman, S.B.; Macivor, J.S. Biodiversity in the City: Fundamental Questions for Understanding the Ecology of Urban Green Spaces for Biodiversity Conservation. Bioscience 2017, 67, 799–807. [Google Scholar] [CrossRef]
- Lindborg, R.; Eriksson, O. Historical landscape connectivity affects present plant species diversity. Ecology 2004, 85, 1840–1845. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Higgs, E.S.; Hall, C. Novel Ecosystems: Intervening in the New Ecological World Order; Wiley-Blackwell: Hoboken, NJ, USA, 2013; p. 380. [Google Scholar]
- Heger, T.; Bernard-Verdier, M.; Gessler, A.; Greenwood, A.D.; Grossart, H.P.; Hilker, M.; Keinath, S.; Kowarik, I.; Kueffer, C.; Marquard, E.; et al. Towards an Integrative, Eco-Evolutionary Understanding of Ecological Novelty: Studying and Communicating Interlinked Effects of Global Change. Bioscience 2019, 69, 888–899. [Google Scholar] [CrossRef]
- Johnson, A.L.; Borowy, D.; Swan, C.M. Land use history and seed dispersal drive divergent plant community assembly patterns in urban vacant lots. J. Appl. Ecol. 2018, 55, 451–460. [Google Scholar] [CrossRef]
- Roman, L.A.; Pearsall, H.; Eisenman, T.S.; Conway, T.M.; Fahey, R.T.; Landry, S.; Vogt, J.; van Doorn, N.S.; Grove, J.M.; Locke, D.H.; et al. Human and biophysical legacies shape contemporary urban forests: A literature synthesis. Urban Urban Gree 2018, 31, 157–168. [Google Scholar] [CrossRef]
- Bornkamm, R. Spontaneous development of urban woody vegetation on differing soils. Flora 2007, 202, 695–704. [Google Scholar] [CrossRef]
- Rebele, F.; Lehmann, C. Twenty years of woodland establishment through natural succession on a sandy landfill site in Berlin, Germany. Urban Urban Gree 2016, 18, 182–189. [Google Scholar] [CrossRef]
- Brazel, A.; Selover, N.; Vose, R.; Heisler, G. The tale of two climates—Baltimore and Phoenix urban LTER sites. Clim. Res. 2000, 15, 123–135. [Google Scholar] [CrossRef]
- Ripplinger, J.; Franklin, J.; Collins, S.L. When the economic engine stalls—A multi-scale comparison of vegetation dynamics in pre- and post-recession Phoenix, Arizona, USA. Landsc. Urban Plan. 2016, 153, 140–148. [Google Scholar] [CrossRef]
- McKinney, M. Effects of urbanization on species richness: A review of plants and animals. Urban Ecosyst. 2008, 11, 161–176. [Google Scholar] [CrossRef]
- Seibold, S.; Cadotte, M.W.; MacIvor, J.S.; Thorn, S.; Müller, J. The Necessity of Multitrophic Approaches in Community Ecology. Trends Ecol. Evol. 2018, 33, 754–764. [Google Scholar] [CrossRef] [PubMed]
- Jaganmohan, M.; Knapp, S.; Buchmann, C.M.; Schwarz, N. The Bigger, the Better? The Influence of Urban Green Space Design on Cooling Effects for Residential Areas. J. Environ. Qual. 2016, 45, 134–145. [Google Scholar] [CrossRef] [PubMed]
- McPhearson, T.; Pickett, S.T.A.; Grimm, N.B.; Niemela, J.; Alberti, M.; Elmqvist, T.; Weber, C.; Haase, D.; Breuste, J.; Qureshi, S. Advancing Urban Ecology toward a Science of Cities. Bioscience 2016, 66, 198–212. [Google Scholar] [CrossRef]
- Turrini, T.; Sanders, D.; Knop, E. Effects of urbanization on direct and indirect interactions in a tri-trophic system. Ecol. Appl. 2016, 26, 664–675. [Google Scholar] [CrossRef]
- Luder, K.; Knop, E.; Menz, M.H.M. Contrasting responses in community structure and phenology of migratory and non-migratory pollinators to urbanization. Divers. Distrib. 2018, 24, 919–927. [Google Scholar] [CrossRef]
- Kowarik, I. Some responses of flora and vegetation to urbanization in central Europe. In Urban ecology: Plants and Plant Communities in Urban Environments; Sukopp, H., Hejný, S., Kowarik, I., Eds.; SPB Academic Publishing: The Hague, The Netherlands, 1990; pp. 45–74. [Google Scholar]
- Hill, M.O.; Roy, D.B.; Thompson, K. Hemeroby, urbanity and ruderality: Bioindicators of disturbance and human impact. J. Appl. Ecol. 2002, 39, 708–720. [Google Scholar] [CrossRef]
- Redman, C.L.; Grove, J.M.; Kuby, L.H. Integrating social science into the long-term ecological research (LTER) network: Social dimensions of ecological change and ecological dimensions of social change. Ecosystems 2004, 7, 161–171. [Google Scholar] [CrossRef]
- Hope, D.; Gries, C.; Zhu, W.X.; Fagan, W.F.; Redman, C.L.; Grimm, N.B.; Nelson, A.L.; Martin, C.; Kinzig, A. Socioeconomics drive urban plant diversity. Proc. Natl. Acad. Sci. USA 2003, 100, 8788–8792. [Google Scholar] [CrossRef]
- Petřík, P.; Sádlo, J.; Hejda, M.; Štajerová, K.; Pyšek, P.; Pergl, J. Composition patterns of ornamental flora in the Czech Republic. Neobiota 2019, 52, 87–109. [Google Scholar] [CrossRef]
- Čeplová, N.; Lososová, Z.; Kalusová, V. Urban ornamental trees: A source of current invaders; a case study from a European City. Urban Ecosyst. 2017, 20, 1135–1140. [Google Scholar] [CrossRef]
- Buchholz, S.; Hannig, K.; Moller, M.; Schirmel, J. Reducing management intensity and isolation as promising tools to enhance ground-dwelling arthropod diversity in urban grasslands. Urban Ecosyst. 2018, 21, 1139–1149. [Google Scholar] [CrossRef]
- Thompson, G.L.; Kao-Kniffin, J. Urban Grassland Management Implications for Soil C and N Dynamics: A Microbial Perspective. Front. Ecol. Evol. 2019, 7, 315. [Google Scholar] [CrossRef]
- Palliwoda, J.; Kowarik, I.; von der Lippe, M. Human-biodiversity interactions in urban parks: The species level matters. Landsc. Urban Plan. 2017, 157, 394–406. [Google Scholar] [CrossRef]
- Kowarik, I. Urban wilderness: Supply, demand, and access. Urban Urban Gree 2018, 29, 336–347. [Google Scholar] [CrossRef]
- Senate Department for Urban Development and Housing. Berlin Environmental Atlas. 06.02.1 Actual Use and Vegetation Cover. 2016. Available online: https://www.stadtentwicklung.berlin.de/umwelt/umweltatlas/eke601.htm (accessed on 2 March 2020).
- Sukopp, H. (Ed.) Stadtökologie: Das Beispiel Berlin; D. Reimer: Berlin, Germany, 1990; p. 455. [Google Scholar]
- Krausch, H.D. Die Sandtrockenrasen (Sedo-Scleranthetea) in Brandenburg. Mitt. Flor.-Soz. Arbeitsgem. 1968, NF-13, 71–100. [Google Scholar]
- Kowarik, I. The “Green Belt Berlin”: Establishing a greenway where the Berlin Wall once stood by integrating ecological, social and cultural approaches. Landsc. Urban Plan. 2019, 184, 12–22. [Google Scholar] [CrossRef]
- Cubasch, U.; Kadow, C. Global Climate Change and Aspects of Regional Climate Change in the Berlin-Brandenburg Region. Erde 2011, 142, 3–20. [Google Scholar]
- Quanz, J.A.; Ulrich, S.; Fenner, D.; Holtmann, A.; Eimermacher, J. Micro-Scale Variability of Air Temperature within a Local Climate Zone in Berlin, Germany, during Summer. Climate 2018, 6, 5. [Google Scholar] [CrossRef]
- Sukopp, H. On the history of plant geography and plant ecology in Berlin. Englera 1987, 7, 85–103. [Google Scholar]
- Krausch, H.; Sukopp, H. Geschichte der Erforschung von Flora und Vegetation in Berlin und Brandenburg; Beiheft 6; Botanischer Verein von Berlin und Brandenburg: Berlin, Germany, 2009; pp. 5–155. [Google Scholar]
- Seitz, B.; Ristow, M.; Meißner, J.; Machatzki, B.; Sukopp, H. Rote Liste und Gesamtartenliste der etablierten Farn- und Blütenpflanzen von Berlin. In Rote Listen der Gefährdeten Pflanzen, Pilze und Tiere von Berlin; Der Landesbeauftrage für Naturschutz und Landschaftspflege & Senatsverwaltung für Umwelt, Verkehr und Klimaschutz: Berlin, Germany, 2018. [Google Scholar]
- Lachmund, J. Greening Berlin: The Co-Production of Science, Politics, and Urban Nature; The MIT Press: Cambridge, MA, USA; London, UK, 2013; p. 320. [Google Scholar]
- Kowarik, I. Herbert Sukopp—An inspiring pioneer in the field of urban ecology. Urban Ecosyst. 2020. [Google Scholar]
- Arbeitsgemeinschaft Biotopkartierung. Beschreibung der Biotoptypen; Senatsverwaltung für Stadtentwicklung und Umwelt: Berlin, Germany, 2005; Available online: https://www.berlin.de/senuvk/natur_gruen/naturschutz/biotopschutz/download/biotopkartierung/biotoptypen.pdf (accessed on 2 March 2020).
- van der Maarel, E.; Franklin, J. (Eds.) Vegetation Ecology; John Wiley & Sons: Chichester, UK, 2012; p. 572. [Google Scholar]
- Schubert, R.; Hilbig, W.; Klotz, S. Bestimmungsbuch der Pflanzengesellschaften Deutschlands; Spektrum Akademischer Verlag: Heidelberg, Germany, 2009; p. 488. [Google Scholar]
- QGIS Development Team. QGIS Geographic Information System; Open Source Geospatial Foundation Project: 2016. Available online: http://qgis.osgeo.org (accessed on 2 March 2020).
- Senate Department for Urban Development and Housing. Berlin Environmental Atlas. 05.08 Biotope Types. 2014. Available online: https://www.stadtentwicklung.berlin.de/umwelt/umweltatlas/ek508.htm (accessed on 2 March 2020).
- Landesamt für Umwelt Brandenburg. Flächendeckende Biotop- und Landnutzungskartierung (BTLN) im Land Brandenburg—CIR-Biotoptypen 2009. 2009. Available online: https://lfu.brandenburg.de/cms/detail.php/bb1.c.359429.de (accessed on 2 March 2020).
- Senate Department for Urban Development and Housing. Berlin Environmental Atlas. 06.01 Actual Use of Built-up Areas/06.02 Inventory of Green and Open Spaces 2010. 2011. Available online: https://www.stadtentwicklung.berlin.de/umwelt/umweltatlas/ekd601.htm (accessed on 2 March 2020).
- Hill, D.; Fasham, M.; Tucker, G.; Shewry, M.; Shaw, P. (Eds.) Handbook of Biodiversity Methods: Survey, Evaluation and Monitoring; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Der Landesbeauftrage für Naturschutz und Landschaftspflege; Senatsverwaltung für Umwelt, Verkehr und Klimaschutz (Eds.) Rote Listen der gefährdeten Pflanzen, Pilze und Tiere von Berlin; Universitätsverlag TU Berlin: Berlin, Germany, 2018. [Google Scholar]
- Holmer, B.; Postgard, U.; Eriksson, M. Sky view factors in forest canopies calculated with IDRISI. Theor. Appl. Climatol. 2001, 68, 33–40. [Google Scholar] [CrossRef]
- Hanski, I. A Practical Model of Metapopulation Dynamics. J. Anim. Ecol. 1994, 63, 151–162. [Google Scholar] [CrossRef]
- Senate Department for Urban Development and Housing. Monitoring Soziale Stadtentwicklung. 2017. Available online: https://www.stadtentwicklung.berlin.de/planen/basisdaten_stadtentwicklung/monitoring/index.shtml (accessed on 2 March 2020).
- Staatsbibliothek zu Berlin, Preußischer Kulturbesitz. Preußische Uraufnahme (1831-71); Landesamt für Vermessung und Geobasisinformationen: Berlin, Germany; pp. 1831–1871.
- Staatsbibliothek zu Berlin, Preußischer Kulturbesitz. Preußische Neuaufnahme (1927-40); Landesamt für Vermessung und Geobasisinformationen: Berlin, Germany, 1927–1940. [Google Scholar]
- Senate Department for Urban Development and Housing. Berlin Environmental Atlas. 04.02 Long-term Mean Air Temperatures 1961–1990. 2001. Available online: https://www.stadtentwicklung.berlin.de/umwelt/umweltatlas/eka402.htm (accessed on 2 March 2020).
- Senate Department for Urban Development and Housing. Berlin Environmental Atlas. 04.05 Urban Climate Zones. 2001. Available online: https://www.stadtentwicklung.berlin.de/umwelt/umweltatlas/eka405.htm (accessed on 2 March 2020).
- Göteborg Urban Climate Group. SOLWEIG1D Version 2015a; University of Gothenburg: Gothenburg, Sweden, 2015; Available online: https://gvc.gu.se/english/research/climate/urban-climate/software/solweig (accessed on 2 March 2020).
- Falchi, F.; Cinzano, P.; Duriscoe, D.; Kyba, C.C.M.; Elvidge, C.D.; Baugh, K.; Portnov, B.A.; Rybnikova, N.A.; Furgoni, R. The new world atlas of artificial night sky brightness. Sci. Adv. 2016, 2, e1600377. [Google Scholar] [CrossRef]
- Kuechly, H.U.; Kyba, C.C.M.; Ruhtz, T.; Lindemann, C.; Wolter, C.; Fischer, J.; Holker, F. Aerial survey and spatial analysis of sources of light pollution in Berlin, Germany. Remote Sens. Environ. 2012, 126, 39–50. [Google Scholar] [CrossRef]
- Blume, H.P.; Karl, S.; Peter, L. Bodenkundliches Praktikum—Eine Einführung in pedologisches Arbeiten für Ökologen, Land- und Forstwirte, Geo- und Umweltwissenschaftler; Spektrum Akademischer Verlag: Heidelberg, Germany, 2011. [Google Scholar]
- ESRI. ArcGIS 10.3.1 for Desktop; Environmental Systems Research Institute: Redlands, CA, USA, 2015; Available online: https://www.esri.com/en-us/arcgis/products/arcgis-desktop/overview (accessed on 2 March 2020).
- Senate Department for Urban Development and Housing. Berlin Environmental Atlas. 06.06 Population Density 2015. 2016. Available online: http://www.stadtentwicklung.berlin.de/umwelt/umweltatlas/ekl606.htm (accessed on 2 March 2020).
- Senate Department for Urban Development and Housing. Berlin Environmental Atlas. 06.09 Urban Structural Density. 2012. Available online: https://www.stadtentwicklung.berlin.de/umwelt/umweltatlas/ek609.htm (accessed on 2 March 2020).
- GmbH, G. OpenStreetMap Shapefile “osm_roads_line_2015_25833”. 2015. Available online: https://www.geofabrik.de/de/data/shapefiles.html (accessed on 2 March 2020).
- Senate Department for Urban Development and Housing. Berlin Environmental Atlas. Protected Areas by Nature Conservation Legislation (incl. Natura 2000). 2016. Available online: https://www.berlin.de/senuvk/natur_gruen/naturschutz/schutzgebiete/ (accessed on 2 March 2020).
- Senate Department for Urban Development and Housing. Geoportal Berlin. Arbeitslosenanteil 2016. 2016. Available online: https://www.stadtentwicklung.berlin.de/planen/basisdaten_stadtentwicklung/monitoring/de/2017/index.shtml (accessed on 24 March 2020).
- Senate Department for Urban Development and Housing. Geoportal Berlin. Einwohnerinnen und Einwohner mit Migrationshintergrund 2016. 2016. Available online: https://www.stadtentwicklung.berlin.de/planen/basisdaten_stadtentwicklung/monitoring/de/2017/index.shtml (accessed on 24 March 2020).
- Senate Department for Urban Development and Housing. Geoportal Berlin. Wohnfläche 2016. 2016. Available online: https://www.stadtentwicklung.berlin.de/planen/basisdaten_stadtentwicklung/monitoring/de/2017/index.shtml (accessed on 24 March 2020).
- Senate Department for Urban Development and Housing. Geoportal Berlin. Wohndauer über 5 Jahre 2016. 2016. Available online: https://www.stadtentwicklung.berlin.de/planen/basisdaten_stadtentwicklung/monitoring/de/2017/index.shtml (accessed on 24 March 2020).
- Straka, T.M.; Wolf, M.; Gras, P.; Buchholz, S.; Voigt, C.C. Tree Cover Mediates the Effect of Artificial Light on Urban Bats. Front. Ecol. Evol. 2019, 7, 91. [Google Scholar] [CrossRef]
- Schittko, C.; Bernard-Verdier, M.; Heger, T.; Buchholz, S.; Kowarik, I.; von der Lippe, M.; Seitz, B.; Joshi, J.; Jeschke, J. A multidimensional framework for measuring biotic novelty: How novel is a community? bioRxiv 2019. [Google Scholar] [CrossRef]
- Möller, M.; Blick, T.; Buchholz, S. Spiders of dry grasslands in and around Berlin—Diversity, distribution and endangerment. Arachnol. Lett. 2019, 58, 52–61. [Google Scholar] [CrossRef]
- Gathof, A.K.; Grossmann, A.J.; Buchholz, S. Wildbienen (Hymenoptera: Apidae) auf Berliner Trockenrasen. Märkische Entomol. Nachr. 2019, 21, 140–162. [Google Scholar]
- Kowarik, I.; Hiller, A.; Planchuelo, G.; Seitz, B.; von der Lippe, M.; Buchholz, S. Emerging Urban Forests: Opportunities for Promoting the Wild Side of the Urban Green Infrastructure. Sustainability 2019, 11, 6318. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Spatial Scale in Survey Areas | Red Lists/Checklists at City Scale | Citywide Distribution Data | |
|---|---|---|---|---|
 | Plants (Plantae) |  | ✔ | ✔ |
 | Wild bees (Anthophila) | | ✔ | |
 | Hoverflies (Syrphidae) | | ✔ | |
 | Butterflies (Lepidoptera) |  | ✔ | |
 | Moths (Lepidoptera) | | ||
 | Grasshoppers (Orthoptera) | | ✔ | ✔ |
 | Ground beetles (Carabidae) | | ✔ | |
 | Rove beetles (Staphylinidae) | | ✔ | |
 | Woodlice (Isopoda) | | ||
 | Myriapods (Myriapoda) | | ||
 | Spiders (Araneae) | | ✔ | |
 | Harvestmen (Opiliones) | | ✔ | |
 | Sand lizard (Lacerta agilis) | | ✔ | |
 | Bats (Chiroptera) |  | ✔ | |
| Variable | Unit/Range/Categories | Spatial Scale | Year | Method/Equipment/Software | Data Source/Reference |
|---|---|---|---|---|---|
| 1. Abiotic/Physical | |||||
| a. Microclimate | |||||
| Air temperature zone longtime medium 1961-1990 | °C | patch | 2016 | Software: QGIS Version 2.18.0 [118] | Berlin Environmental Atlas/Long-term Mean Air Temperatures 1961–1990 [129] |
| Urban climatic zone changes in temperature, air humidity, and wind regime compared to open land conditions | 0 = no; 1 = very low; 2 = low; 3 = medium; 4 = high changes | patch | 2016 | Software: QGIS Version 2.18.0 [118] | Berlin Environmental Atlas/Urban Climate Zones [130] |
| Air temperature values measured at 2 m height | ° C | plot | 2017 | Data logger: EasyLog EL-USB-2+, Lascar Electronics | |
| Relative air humidity measured at 2 m height | % | plot | 2017 | ||
| Dew point measured at 2 m height | ° C | plot | 2017 | ||
| Sky view factor share of open sky | 0–1 | plot | 2017 | Analysis of fisheye photos; Camera: Canon EOS 700D; Fisheye lens: SIGMA 4.5mm F2.8 EX DC HSM Circular Fisheye; Software: SOLWEIG1D Version 2015a [131] | Holmer et al. 2001 [124] |
| b. Light | |||||
| Skyglow value at a maximum cloudiness of 12.5 % | µcd/m² | plot | 2017 | Sky quality meter: Sky Quality Meter—L, Unihedron, Ontario, Canada | |
| Maximum illuminance at a maximum cloudiness of 12.5 % | lux | plot | 2017 | Lux meter: LX-1108, VOLTCRAFT, Hirschau, Germany | |
| Mean skyglow based on satellite data | µcd/m² | buffer | 2017 | Software: QGIS Version 2.14 [118] | Falchi et al. 2016 [132] |
| Mean light pollution based on aerial high resolution (1 m²) mosaic image | 0–… | buffer | 2017 | Software: QGIS Version 2.14 [118] | Kuechly et al. 2012 [133] |
| c. Soil chemistry | |||||
| pH value | 0–14 | plot | 2017 | Analysis of soil samples; Inductively coupled plasma optical emission spectrometry (ICP-OES): iCAP 6000 ICP Spectrometer, Thermo Fisher Scientific, Dreieich, Germany | Blume et al. 2011 [134] |
| Organic carbon (C) content | g/kg | plot | 2017 | ||
| Nitrogen (N) content | g/kg | plot | 2017 | ||
| Sulfur (S) content | g/kg | plot | 2017 | ||
| Phosphorus (P) content | mg/kg | plot | 2017 | ||
| Potassium (K) content | mg/kg | plot | 2017 | ||
| Copper (Cu) content | mg/kg | plot | 2017 | ||
| Zinc (Zn) content | mg/kg | plot | 2017 | ||
| Cadmium (Cd) content | mg/kg | plot | 2017 | ||
| Lead (Pb) content | mg/kg | plot | 2017 | ||
| Nickel (Ni) content | mg/kg | plot | 2017 | ||
| Gravimetric water content | % | plot | 2017 | ||
| Electrical conductivity in suspension | mS/m | plot | 2017 | ||
| Cation exchange capacity | µmolc/g | plot | 2017 | ||
| Carbon-to-nitrogen (C/N) ratio | % | plot | 2017 | ||
| Carbon-to-sulfur (C/S) ratio | % | plot | 2017 | ||
| 2. Spatial | |||||
| a. Continuity/Connectivity | |||||
| Historical continuity as grassland biotope | O = old (established before 1940); N = new (established after 1940) | patch | 2016 | Intersection of current dry grassland biotopes with historical grassland biotopes (from digitized and georeferenced historical land-use maps); Software: QGIS Version 2.18.0 [118] | Historical land-use maps: Preußische Uraufnahme (1831–71) [127], Preußische Neuaufnahme (1927–40) [128]; Berlin Environmental Atlas/Biotope Types [119], Biotope mapping Brandenburg [120] |
| Patch size of dry grassland | m² | patch | 2017 | Software: QGIS Version 2.18.0 [118] | Berlin Environmental Atlas/Biotope Types [119], Biotope mapping Brandenburg [120] |
| Share of dry grassland in the surrounding | 0–1 | buffer | 2016 | Software: QGIS Version 2.18.0 [118] | Berlin Environmental Atlas/Biotope Types [119], Biotope mapping Brandenburg [120] |
| Hanski’s connectivity index (HCI) based on pairwise distances (d) between plot (i) and dry grassland patches (j) and area (A) of the patches | 0–… | distance | 2020 | HCI = ∑j≠i exp(−α dij) Aj; Software: ArcGIS 10.3.1 for Desktop [135]; Tool: Generate Near Table (Analysis) | Berlin Environmental Atlas/Biotope Types [119], Biotope mapping Brandenburg [120]; Hanski 1994 [125] |
| b. Urbanity | |||||
| Sealed surface | % | buffer | 2017 | Software: QGIS Version 2.18.0 [118]; Tool: Zonal statistics | Berlin Environmental Atlas/Actual Use of Built-up Areas, Inventory of Green and Open Spaces 2010 [121] |
| Population size | number of inhabitants | buffer | 2017 | Software: QGIS Version 2.18.0 [118] | Berlin Environmental Atlas/Population Density 2015 [136] |
| Population density | inhabitants/ ha | buffer | 2017 | Software: QGIS Version 2.18.0 [118]; Tool: Zonal statistics | Berlin Environmental Atlas/Population Density 2015 [136] |
| Floor space index (FSI) | m² floor area/ m² lot area | buffer | 2017 | Software: QGIS Version 2.18.0 [118]; Tool: Zonal statistics | Berlin Environmental Atlas/Urban Structural Density [137] |
| Urban land cover (ULC) Built-up areas and traffic areas (biotope type 12 according to the Berlin classification of biotope types) | % | buffer | 2018 | Software: QGIS Version 2.18.0 [118] | Berlin Environmental Atlas/Biotope Types [119], Biotope mapping Brandenburg [120]; Hahs & McDonnell 2006 [70] |
| People per unit Urban Land Cover (PULC) Ratio of people divided by the proportion of urban land cover | inhabitants/ ULC | buffer | 2018 | PULC = Population/(ULC + 0.5); Software: QGIS Version 2.18.0 [118] | |
| Road density Total length of roads | km | buffer | 2017 | Software: QGIS Version 2.18.0 [118]; Tool: Sum line lengths | OpenStreetMap shapefile “osm_roads_line_2015_25833” [138] |
| Road distance Shortest distance from plot midpoint to nearest road | m | distance | 2017 | Software: QGIS Version 2.18.0 [118]; Tool: v.distance | |
| Railway density Total length of railways | km | buffer | 2017 | Software: QGIS Version 2.18.0 [118]; Tool: Sum line lengths | |
| Railway distance Shortest distance from plot midpoint to nearest railway | m | distance | 2017 | Software: QGIS Version 2.18.0 [118]; Tool: v.distance | |
| Distance to central business district Shortest distance from plot midpoint to Friedrichstraße station | km | distance | 2018 | Software: QGIS Version 2.18.0 [118]; Tool: v.distance | Hahs & McDonnell 2006 [70] |
| c. Landscape variables/Environment | |||||
| Number of biotope patches | number | buffer | 2018 | Software: QGIS Version 2.18.0 [118] | Berlin Environmental Atlas/Biotope Types [119], Biotope mapping Brandenburg [120]; Hahs & McDonnell 2006 [70] |
| Size of largest biotope patch | ha | buffer | 2018 | Software: QGIS Version 2.18.0 [118] | |
| Land cover richness Number of different biotope types (based on the highest level of the Berlin classification of biotope types) | 1–12 | buffer | 2018 | Software: QGIS Version 2.18.0 [118] | |
| Landscape shape index (LSI) Ratio of sum of edge length to total area for a landscape measured against a circle standard | 1–… | buffer | 2018 | LSI = Edge length/(2 * √(Pi * Area)); Software: QGIS Version 2.18.0 [118] | |
| Type of protected area | LCA = Landscape Conservation Area; NP = Nature Park; FFH = Flora Fauna Habitat Area; SPA = Special Protected Area according to Bird Conservation Directive; None | patch | 2016 | Software: QGIS Version 2.18.0 [118] | Berlin Environmental Atlas/Protected Areas by Nature Conservation Legislation (incl. Natura 2000) [139] |
| 3. Habitat structure | |||||
| Total vegetation cover | % | plot | 2017 | Estimation | |
| Cover of herbal layer | % | plot | 2017 | Estimation | |
| Cover of moss layer | % | plot | 2017 | Estimation | |
| Cover of litter layer | % | plot | 2017 | Estimation | |
| Height of herbal layer | cm | plot | 2017 | Estimation | |
| Degree of slope | ° | plot | 2017 | Estimation | |
| Aspect of sloped plots | 90° = East; 180° = South; 270° = West; 360° = North | plot | 2017 | Estimation | |
| Wild boars Digging traces | 0 = no; 1 = yes | plot | 2017 | Estimation | |
| Regular mowing at least once a year | 0 = no; 1 = yes | plot | 2017 | Estimation | |
| 4. Socioeconomic | |||||
| Unemployed people | % | buffer | 2018 | Software: QGIS Version 2.18.0 [118] | Geoportal Berlin/unemployment rate 2016 [140] |
| Inhabitants with migration background | % | buffer | 2018 | Software: QGIS Version 2.18.0 [118] | Geoportal Berlin/residents with migration background 2016 [141] |
| Mean living space per inhabitant | m² | buffer | 2018 | Software: QGIS Version 2.18.0 [118] | Geoportal Berlin/living space 2016 [142] |
| Occupancy of inhabitants People who have lived at the same address for at least five years | % | buffer | 2018 | Software: QGIS Version 2.18.0 [118] | Geoportal Berlin/residence over 5 years 2016 [143] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
von der Lippe, M.; Buchholz, S.; Hiller, A.; Seitz, B.; Kowarik, I. CityScapeLab Berlin: A Research Platform for Untangling Urbanization Effects on Biodiversity. Sustainability 2020, 12, 2565. https://doi.org/10.3390/su12062565
von der Lippe M, Buchholz S, Hiller A, Seitz B, Kowarik I. CityScapeLab Berlin: A Research Platform for Untangling Urbanization Effects on Biodiversity. Sustainability. 2020; 12(6):2565. https://doi.org/10.3390/su12062565
Chicago/Turabian Stylevon der Lippe, Moritz, Sascha Buchholz, Anne Hiller, Birgit Seitz, and Ingo Kowarik. 2020. "CityScapeLab Berlin: A Research Platform for Untangling Urbanization Effects on Biodiversity" Sustainability 12, no. 6: 2565. https://doi.org/10.3390/su12062565
APA Stylevon der Lippe, M., Buchholz, S., Hiller, A., Seitz, B., & Kowarik, I. (2020). CityScapeLab Berlin: A Research Platform for Untangling Urbanization Effects on Biodiversity. Sustainability, 12(6), 2565. https://doi.org/10.3390/su12062565