Clean and Green Urban Water Bodies Benefit Nocturnal Flying Insects and Their Predators, Insectivorous Bats


 ,
,
Abstract
1. Introduction
2. Methods
2.1. Study Area and Selection of Study Sites
2.2. Data Collection
2.3. Landscape- and Water Body Scale Variables
2.4. Data analyses and Model Selection
3. Results
3.1. Landscape Factors Driving Nocturnal Flying Insects in the Urban Environment
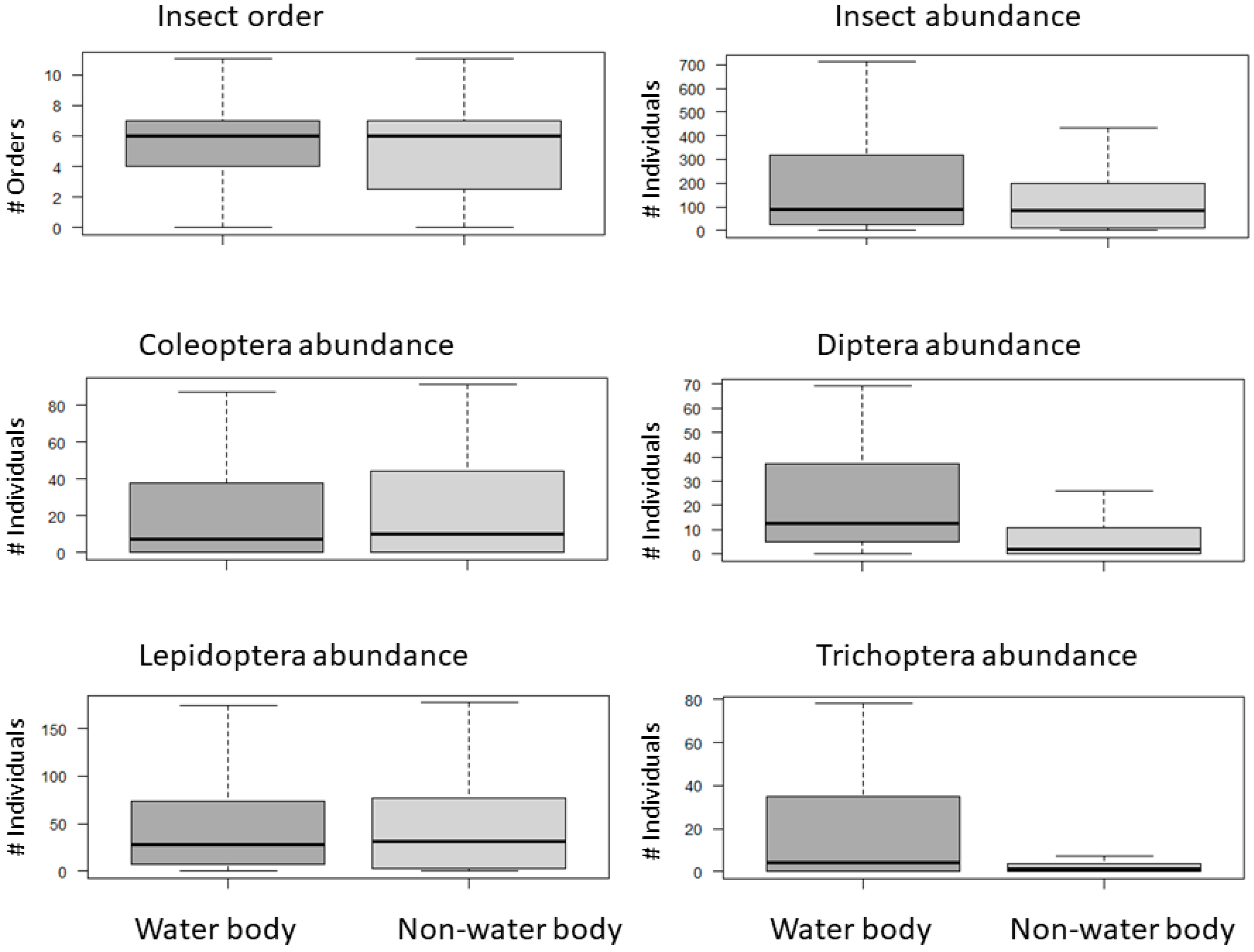
3.2. Water Body Characteristics Driving Nocturnal Flying Insects
3.3. Relationship between Insect Order Richness and Insectivorous Bat Species Richness and Activity
4. Discussion
4.1. Landscape Factors Driving Nocturnal Flying Insects in the Urban Environment
4.2. Water Body Characteristics Driving Nocturnal Flying Insects
4.3. Relationship among Insects, Landscape Variables and Insectivorous Bats
4.4. Implications for Insectivorous Bats
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Batzer, D.P.; Wissinger, S.A. Ecology of insect communities in nontidal wetlands. Annu. Rev. Entomol. 1996, 41, 75–100. [Google Scholar] [CrossRef]
- Colless, D.H.; McAlpine, D.K. Diptera. In The Insects of Australia: A Textbook for Students and Research Workers, 2nd ed.; Naumann, I.D., Came, P.B., Lawrence, J.F., Nielsen, E.S., Spradbery, J.P., Taylor, R.W., Whitten, M.J., Littlejohn, M.J., Eds.; Melbourne University Press: Melbourne, Australia; Cornell University Press: Ithaca, NY, USA, 1991; Volume 1, pp. 717–786. [Google Scholar]
- Neboiss, A. Trichoptera. In The Insects of Australia: A Textbook for Students and Research Workers, 2nd ed.; Naumann, I.D., Came, P.B., Lawrence, J.F., Nielsen, E.S., Spradbery, J.P., Taylor, R.W., Whitten, M.J., Littlejohn, M.J., Eds.; Melbourne University Press: Melbourne, Australia; Cornell University Press: Ithaca, NY, USA, 1991; Volume 1, pp. 787–816. [Google Scholar]
- Warren, P.H. Spatial and temporal variation in the structure of a freshwater food web. Oikos 1989, 55, 299–311. [Google Scholar] [CrossRef]
- Racey, P.A.; Swift, S.M. Feeding ecology of Pipistrellus pipistrellus (Chiroptera: Vespertilionidae) during pregnancy and lactation. I. Foraging behaviour. J. Anim. Ecol. 1985, 45, 205–215. [Google Scholar] [CrossRef]
- Wickramasinghe, L.P.; Harris, S.; Jones, G.; Jennings, N.V. Abundance and species richness of nocturnal insects on organic and conventional farms: Effects of agricultural intensification on bat foraging. Conserv. Biol. 2004, 18, 1283–1292. [Google Scholar] [CrossRef]
- Vindigni, M.A.; Morris, A.D.; Miller, D.A.; Kalcounis-Rueppell, M.C. Use of modified water sources by bats in a managed pine landscape. Forest Ecol. Manag. 2009, 258, 2056–2061. [Google Scholar] [CrossRef]
- Stahlschmidt, P.; Pätzold, A.; Ressl, L.; Schulz, R.; Brühl, C.A. Constructed wetlands support bats in agricultural landscapes. Basic Appl. Ecol. 2012, 13, 196–203. [Google Scholar] [CrossRef]
- Piano, E.; Souffreau, C.; Merckx, T.; Baardsen, L.; Backeljau, T.; Bonte, D.; Brans, K.I.; Cours, M.; Dahirel, M.; Debortoli, N.; et al. Urbanization drives cross-taxon declines in abundance and diversity at multiple spatial scales. Glob. Chang. Biol. 2019. [Google Scholar] [CrossRef]
- Göbel, P.; Dierkes, C.; Coldewey, W.G. Storm water runoff concentration matrix for urban areas. J. Contam. Hydrol. 2007, 91, 26–42. [Google Scholar] [CrossRef]
- Pettigrove, V.; Hoffmann, A. Impact of urbanisation on heavy metal contamination in urban stream sediments: Influence of catchment geology. Australas. J. Ecotoxicol. 2003, 9, 119–128. [Google Scholar]
- Ehrenfeld, J.G. Exotic invasive species in urban wetlands: Environmental correlates and implications for wetland management. J. App. Ecol. 2008, 45, 1160–1169. [Google Scholar] [CrossRef]
- Owen, C. Hydrology and history: Land use changes and ecological responses in an urban wetland. Wetl. Ecol. Manag. 1998, 6, 209–219. [Google Scholar] [CrossRef]
- Lundquist, M.J.; Zhu, W. Aquatic insect functional diversity and nutrient content in urban streams in a medium-sized city. Ecosphere 2018, 9, e02284. [Google Scholar] [CrossRef]
- Pettigrove, V.; Hoffmann, A. A field-based microcosm method to assess the effects of polluted urban stream sediments on aquatic macroinvertebrates. Environ. Toxicol. Chem. 2005, 24, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Walsh, C.J.; Sharpe, A.K.; Breen, P.F.; Sonneman, J.A. Effects of urbanization on streams of the Melbourne region, Victoria, Australia. I. Benthic macroinvertebrate communities. Freshw. Biol. 2001, 46, 535–551. [Google Scholar] [CrossRef]
- Naidoo, S.; Vosloo, D.; Schoeman, M.C. Foraging at wastewater treatment works increases the potential for metal accumulation in an urban adapter, the banana bat (Neoromicia nana). Afr. Zool. 2013, 48, 39–55. [Google Scholar] [CrossRef]
- Jung, K.; Threlfall, C.G. Urbanisation and its effects on bats—A global meta-analysis. In Bats in the Anthropocene: Conservation of Bats in a Changing World; Voigt, C.C., Kingston, T., Eds.; Springer: Cham, Switzerland, 2016; pp. 13–33. [Google Scholar]
- Russo, D.; Ancillotto, L. Sensitivity of bats to urbanization: A review. Mamm. Biol. 2015, 80, 205–212. [Google Scholar] [CrossRef]
- Jong, J.; Ahlén, I. Factors affecting the distribution pattern of bats in Uppland, central Sweden. Ecography 1991, 14, 92–96. [Google Scholar] [CrossRef]
- Jones, G.; Jacobs, D.S.; Kunz, T.H.; Willig, M.R.; Racey, P.A. Carpe noctem: The importance of bats as bioindicators. Endanger. Species Res. 2009, 8, 93–115. [Google Scholar] [CrossRef]
- Russo, D.; Jones, G. Bats as bioindicators: An introduction. Mamm. Biol. Z. Säugetierkd. 2015, 80, 157–158. [Google Scholar] [CrossRef]
- Hill, M.J.; Biggs, J.; Thornhill, I.; Briers, R.A.; Gledhill, D.G.; White, J.C.; Wood, P.J.; Hassall, C. Urban ponds as an aquatic biodiversity resource in modified landscapes. Glob. Chang. Biol. 2017, 23, 986–999. [Google Scholar] [CrossRef]
- Thornhill, I.; Batty, L.; Death, R.G.; Friberg, N.R.; Ledger, M.E. Local and landscape scale determinants of macroinvertebrate assemblages and their conservation value in ponds across an urban land-use gradient. Biodivers. Conserv. 2017, 26, 1065–1086. [Google Scholar] [CrossRef] [PubMed]
- Mata, L.; Threlfall, C.G.; Williams, N.S.; Hahs, A.K.; Malipatil, M.; Stork, N.E.; Livesley, S.J. Conserving herbivorous and predatory insects in urban green spaces. Sci. Rep. 2017, 7, 40970. [Google Scholar] [CrossRef] [PubMed]
- Threlfall, C.G.; Mata, L.; Mackie, J.; Hahs, A.K.; Stork, N.E.; Williams, N.S.G.; Livesley, S.J. Increasing biodiversity in urban green spaces through simple vegetation interventions. J. Appl. Ecol. 2017, 54, 1874–1883. [Google Scholar] [CrossRef]
- Knop, E.; Zoller, L.; Ryser, R.; Gerpe, C.; Hörler, M.; Fontaine, C. Artificial light at night as a new threat to pollination. Nature 2017, 548, 206. [Google Scholar] [CrossRef]
- Owens, A.C.; Lewis, S.M. The impact of artificial light at night on nocturnal insects: A review and synthesis. Ecol. Evol. 2018, 8, 11337–11358. [Google Scholar] [CrossRef]
- Avila-Flores, R.; Fenton, M.B. Use of spatial features by foraging insectivorous bats in a large urban landscape. J. Mammal. 2005, 86, 1193–1204. [Google Scholar] [CrossRef]
- Ober, H.K.; Hayes, J.P. Influence of forest riparian vegetation on abundance and biomass of nocturnal flying insects. Forest Ecol. Manag. 2008, 256, 1124–1132. [Google Scholar] [CrossRef]
- Carew, M.E.; Pettigrove, V.; Cox, R.L.; Hoffmann, A.A. The response of Chironomidae to sediment pollution and other environmental characteristics in urban wetlands. Freshw. Biol. 2007, 52, 2444–2462. [Google Scholar] [CrossRef]
- Ruiz-García, A.; Márquez-Rodríguez, J.; Ferreras-Romero, M. Implications of anthropogenic disturbance factors on the Trichoptera assemblage in a Mediterranean fluvial system: Are Trichoptera useful for identifying land-use alterations? Ecol. Indic. 2012, 14, 114–123. [Google Scholar] [CrossRef]
- Straka, T.M.; Lentini, P.E.; Lumsden, L.F.; Wintle, B.A.; van der Ree, R. Urban bat communities are affected by wetland size, quality, and pollution levels. Ecol. Evol. 2016, 6, 4761–4774. [Google Scholar] [CrossRef]
- Oertli, B.; Parris, K.M. Review: Toward management of urban ponds for freshwater biodiversity. Ecosphere 2019, 10, e02810. [Google Scholar] [CrossRef]
- Yen, A.L. Melbourne’s terrestrial invertebrate biodiversity: Losses, gains and the new perspective. Vic. Nat. 2011, 128, 201–208. [Google Scholar]
- Van der Ree, R.; McCarthy, M.A. Inferring persistence of indigenous mammals in response to urbanisation. Anim. Conserv. 2005, 8, 309–319. [Google Scholar]
- Australian Bureau of Statistics (ABS). Regional Population Growth, Australia 2018–2019. Available online: https://www.abs.gov.au/AUSSTATS/abs@.nsf/mf/3218.0 (accessed on 30 October 2019).
- Melbourne Water. Constructed Wetlands Guidelines; Melbourne Water: Melbourne, Australia, 2010. [Google Scholar]
- Kuechly, H.U.; Kyba, C.C.; Ruhtz, T.; Lindemann, C.; Wolter, C.; Fischer, J.; Hölker, F. Aerial survey and spatial analysis of sources of light pollution in Berlin, Germany. Remote Sens. Environ. 2012, 126, 39–50. [Google Scholar] [CrossRef]
- Leopold, L.B. Hydrology for Urban Land Planning—A Guidebook on the Hydrologic Effects of Urban Land Use; US Geological Survey: Washington, DC, USA, 1968; Volume 554. [Google Scholar]
- McGeachie, W.J. The effects of moonlight illuminance, temperature and wind speed on light-trap catches of moths. Bull. Entomol. Res. 1989, 79, 185–192. [Google Scholar] [CrossRef]
- Zahn, A.; Maier, S. Hunting activity of bats at streams and ponds. Z. Saugetierkd. 1997, 62, 1–11. [Google Scholar]
- Lumsden, L.F.; Bennett, A.F. Scattered trees in rural landscapes: Foraging habitat for insectivorous bats in south-eastern Australia. Biol. Conserv. 2005, 122, 205–222. [Google Scholar] [CrossRef]
- Scanlon, A.; Petit, S. Biomass and biodiversity of nocturnal aerial insects in an Adelaide City park and implications for bats (Microchiroptera). Urban Ecosyst. 2008, 11, 91–106. [Google Scholar] [CrossRef]
- O’Donnell, C.F.; Christie, J.E.; Simpson, W. Habitat use and nocturnal activity of lesser short-tailed bats (Mystacina tuberculata) in comparison with long-tailed bats (Chalinolobus tuberculatus) in temperate rainforest. N. Z. J. Zool. 2006, 33, 113–124. [Google Scholar] [CrossRef][Green Version]
- Naumann, I.D.; Came, P.B.; Lawrence, J.F.; Nielsen, E.S.; Spradbery, J.P.; Taylor, R.W.; Whitten, M.J.; Littlejohn, M.J. The Insects of Australia: A Textbook for Students and Research Workers, 2nd ed.; Melbourne University Press: Melbourne, Australia; Cornell University Press: Ithaca, NY, USA, 1991; Volume 1. [Google Scholar]
- Kunz, T.H. Methods for assessing insect availability for insectivorous bats. In Ecological and Behavioural Methods for the Study of Bats; Kunz, T.H., Ed.; Smithsonian Institute Press: Washington, DC, USA, 1988; pp. 191–201. [Google Scholar]
- Threlfall, C.G.; Law, B.; Banks, P.B. Influence of landscape structure and human modifications on insect biomass and bat foraging activity in an urban landscape. PLoS ONE 2012, 7, e38800. [Google Scholar] [CrossRef]
- Adams, M.D.; Law, B.S.; Gibson, M.S. Reliable automation of bat call identification for eastern New South Wales, Australia, using classification trees and AnaScheme software. Acta Chiropterol. 2010, 12, 231–245. [Google Scholar] [CrossRef]
- Yela, J.L.; Holyoak, M. Effects of moonlight and meteorological factors on light and bait trap catches of noctuid moths (Lepidoptera: Noctuidae). Environ. Entomol. 1997, 26, 1283–1290. [Google Scholar] [CrossRef]
- Board, J.E.; Maka, V.; Price, R.; Knight, D.; Baur, M.E. Development of vegetation indices for identifying insect infestations in soybean. Agron. J. 2007, 99, 650–656. [Google Scholar] [CrossRef]
- Stokeld, D.; Hamer, A.J.; van der Ree, R.; Pettigrove, V.; Gillespie, G. Factors influencing occurrence of a freshwater turtle in an urban landscape: A resilient species? Wildl. Res. 2014, 41, 163–171. [Google Scholar] [CrossRef]
- Australian and New Zealand Environment and Conservation Council/Agriculture and resource Management Council of Australia and New Zealand (ANZECC/ARMCANZ). Australian and New Zealand Guidelines for Fresh and Marine Water Quality—The Guidelines; Australia Water Association: Artarmon, NSW, Australia, 2000; Volume 1.
- Windsor, F.M.; Pereira, M.G.; Tyler, C.R.; Ormerod, S.J. Persistent contaminants as potential constraints on the recovery of urban river food webs from gross pollution. Water Res. 2019, 163, 114858. [Google Scholar] [CrossRef]
- Sharley, D.J.; Hoffmann, A.A.; Pettigrove, V. Effects of sediment quality on macroinvertebrates in the Sunraysia region of the Murray—Darling rivers, Australia. Environ. Pollut. 2008, 156, 689–698. [Google Scholar] [CrossRef]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–51. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach; Springer Science & Business Media: New York, NY, USA, 2002. [Google Scholar]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Zuur, A.; Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R.; Springer: New York, NY, USA, 2009. [Google Scholar]
- Rhodes, J.R.; McAlpine, C.A.; Zuur, A.; Smith, G.; Ieno, E. GLMM applied on the spatial distribution of koalas in a fragmented landscape. In Mixed Effects Models and Extensions in Ecology with R.; Zuur, A., Ieno, E.N., Walker, N., Saveliev, A.A., Smith, G.M., Eds.; Springer: New York, NY, USA, 2009; pp. 469–492. [Google Scholar]
- Gresens, S.E.; Belt, K.T.; Tang, J.A.; Gwinn, D.C.; Banks, P.A. Temporal and spatial responses of Chironomidae (Diptera) and other benthic invertebrates to urban stormwater runoff. Hydrobiologia 2007, 575, 173–190. [Google Scholar] [CrossRef]
- Gołdyn, R.; Szpakowska, B.; Świerk, D.; Domek, P.; Buxakowski, J.; Dondajewska, R.; Barałkiewicz, D.; Sajnóg, A. Influence of stormwater runoff on macroinvertebrates in a small urban river and a reservoir. Sci. Total Environ. 2018, 625, 743–751. [Google Scholar] [CrossRef]
- Adler, P.H.; Courtney, G.W. Ecological and societal services of aquatic diptera. Insects 2019, 10, 70. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Rich, P.M.; Price, K.P.; Kettle, W.D. Relations between NDVI and tree productivity in the central Great Plains. Int. J. Remote Sens. 2004, 25, 3127–3138. [Google Scholar] [CrossRef]
- Strong, D.R.; Lawton, J.H.; Southwood, R.S. Insects on Plants: Community Patterns and Mechanisms; Blackwell Scientific: Oxford, UK, 1984. [Google Scholar]
- McGeoch, M.A.; Chown, S.L. Impact of urbanization on a gall-inhabiting Lepidoptera assemblage: The importance of reserves in urban areas. Biodivers. Conserv. 1997, 6, 979–993. [Google Scholar] [CrossRef]
- Beninde, J.; Veith, M.; Hochkirch, A. Biodiversity in cities needs space: A meta-analysis of factors determining intra-urban biodiversity variation. Ecol. Lett. 2015, 18, 581–592. [Google Scholar] [CrossRef]
- Plummer, K.E.; Hale, J.D.; O’Callaghan, M.J.; Sadler, J.P.; Siriwardena, G.M. Investigating the impact of street lighting changes on garden moth communities. J. Urban Ecol. 2016, 2, 1–10. [Google Scholar] [CrossRef]
- Briers, R.A.; Gee, J.H.R. Riparian forestry management and adult stream insects. Hydrol. Earth Syst. Sci. 2004, 3, 545–549. [Google Scholar] [CrossRef]
- Wakefield, A.; Broyles, M.; Stone, E.L.; Harris, S.; Jones, G. Quantifying the attractiveness of broad-spectrum street lights to aerial nocturnal insects. J. Appl. Ecol. 2018, 55, 714–722. [Google Scholar] [CrossRef]
- Urban, M.C.; Skelly, D.K.; Burchsted, D.; Price, W.; Lowry, S. Stream communities across a rural–urban landscape gradient. Divers. Distrib. 2006, 12, 337–350. [Google Scholar] [CrossRef]
- Piccolo, J.J.; Wipfli, M.S. Does red alder (Alnus rubra) in upland riparian forests elevate macroinvertebrate and detritus export from headwater streams to downstream habitats in southeastern Alaska? Can. J. Fish. Aquat. Sci. 2002, 59, 503–513. [Google Scholar] [CrossRef]
- Harrison, S.S.; Harris, I.T. The effects of bankside management on chalk stream invertebrate communities. Freshw. Biol. 2002, 47, 2233–2245. [Google Scholar] [CrossRef]
- Allan, J.D.; Wipfli, M.S.; Caouette, J.P.; Prussian, A.; Rodgers, J. Influence of streamside vegetation on inputs of terrestrial invertebrates to salmonid food webs. Can. J. Fish. Aquat. Sci. 2003, 60, 309–320. [Google Scholar] [CrossRef]
- French, B.W.; Elliott, N.C. Spatial and temporal distribution of ground beetle (Coleoptera: Carabidae) assemblages in riparian strips and adjacent wheat fields. Environ. Entomol. 1999, 28, 597–607. [Google Scholar] [CrossRef]
- Tszydel, M.; Markowski, M.; Majecki, J.; Błońska, D.; Zieliński, M. Assessment of water quality in urban streams based on larvae of Hydropsyche angustipennis (Insecta, Trichoptera). Environ. Sci. Pollut. Res. 2015, 22, 14687–14701. [Google Scholar] [CrossRef] [PubMed]
- Molineri, C.; Tejerina, E.G.; Torrejón, S.E.; Pero, E.J.; Hankel, G.E. Indicative value of different taxonomic levels of Chironomidae for assessing the water quality. Ecol. Indic. 2020, 108, 105703. [Google Scholar] [CrossRef]
- Chantaramongkol, P. Light-trapped caddisflies (trichoptera) as water quality indicators in large rivers: Results from the danube at veröce, hungary. Aquat. Insects 1983, 5, 33–37. [Google Scholar] [CrossRef]
- Blicharska, M.; Andersson, J.; Bergsten, J.; Bjelke, U.; Hilding-Rydevik, T.; Johansson, F. Effects of management intensity, function and vegetation on the biodiversity in urban ponds. Urban For. Urban Green. 2016, 20, 103–112. [Google Scholar] [CrossRef]
- Nag, A.; Nath, P. Effect of moon light and lunar periodicity on the light trap catches of cutworm Agrotis ipsilon (Hufn.) moths. J. Appl. Entomol. 1991, 111, 358–360. [Google Scholar] [CrossRef]
- De Oliveira, L.Q.; Marciente, R.; Magnusson, W.E.; Bobrowiec, P.E.D. Activity of the insectivorous bat Pteronotus parnellii relative to insect resources and vegetation structure. J. Mammal. 2015, 96, 1036–1044. [Google Scholar] [CrossRef]
- Gonsalves, L.; Bicknell, B.; Law, B.; Webb, C.; Monamy, V. Mosquito consumption by insectivorous bats: Does size matter? PLoS ONE 2013, 8, e77183. [Google Scholar] [CrossRef]
- Burgar, J.M.; Murray, D.C.; Craig, M.D.; Haile, J.; Houston, J.; Stokes, V.; Bunce, M. Who’s for dinner? High-throughput sequencing reveals bat dietary differentiation in a biodiversity hotspot where prey taxonomy is largely undescribed. Mol. Ecol. 2014, 23, 3605–3617. [Google Scholar] [CrossRef]
- Whitaker, J.O., Jr.; Neefus, C.; Kunz, T.H. Dietary variation in the Mexican free-tailed bat (Tadarida brasiliensis mexicana). J. Mammal. 1996, 77, 716–724. [Google Scholar] [CrossRef]
- Ancillotto, L.; Bosso, L.; Salinas-Ramos, V.B.; Russo, D. The importance of ponds for the conservation of bats in urban landscapes. Landsc. Urban Plann. 2019, 190, 103607. [Google Scholar] [CrossRef]
- Campbell, S. So long as it’s near water: Variable roosting behaviour of the large-footed myotis (Myotis macropus). Aust. J. Zool. 2009, 57, 89–98. [Google Scholar] [CrossRef]
- Verboom, B.; Spoelstra, K. Effects of food abundance and wind on the use of tree lines by an insectivorous bat, Pipistrellus pipistrellus. Can. J. Zool. 1999, 77, 1393–1401. [Google Scholar] [CrossRef]
- Townsend, K.R.; Pettigrove, V.J.; Carew, M.E.; Hoffmann, A.A. The effects of sediment quality on benthic macroinvertebrates in the River Murray, Australia. Mar. Freshw. Res. 2009, 60, 70–82. [Google Scholar] [CrossRef]
- Lindqvist, L. Accumulation of cadmium, copper, and zinc in five species of phytophagous insects. Environ. Entomol. 1992, 21, 160–163. [Google Scholar] [CrossRef]
- Shwartz, A.; Turbé, A.; Simon, L.; Julliard, R. Enhancing urban biodiversity and its influence on city-dwellers: An experiment. Biol. Conserv. 2014, 171, 82–90. [Google Scholar] [CrossRef]
- Hall, D.M.; Camilo, G.R.; Tonietto, R.K.; Ollerton, J.; Ahrné, K.; Arduser, M.; Ascher, J.S.; Baldock, K.C.R.; Fowler, R.; Frankie, G.; et al. The city as a refuge for insect pollinators. Conserv. Biol. 2017, 31, 24–29. [Google Scholar] [CrossRef]
- Sanford, M.P.; Manley, P.N.; Murphy, D.D. Effects of urban development on ant communities: Implications for ecosystem services and management. Conserv. Biol. 2009, 23(1), 131–141. [Google Scholar]
- Parsons, S.E.; Kerner, L.M.; Frank, S.D. Effects of native and exotic congeners on diversity of invertebrate natural enemies, available spider biomass, and pest control services in residential landscapes. Biol. Conserv. 2020. [Google Scholar] [CrossRef]
- Potts, S.G.; Imperatriz-Fonseca, V.L.; Ngo, H.T.; Aizen, M.A.; Biesmeijer, J.C.; Breeze, T.D.; Dicks, L.V.; Garibaldi, L.A.; Hill, R.; Settele, J.; et al. Safeguarding pollinators and their values to human well-being. Nature 2016, 540, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Hochkirch, A. The insect crisis we can’t ignore. Nature 2016, 539, 141. [Google Scholar] [CrossRef] [PubMed]
- Harvey, J.A.; Heinen, R.; Armbrecht, I.; Basset, Y.; Baxter-Gilbert, J.; Bezemer, T.M.; Böhm, M.; Bommarco, R.; Borges, P.A.V.; Cardoso, P.; et al. International scientists formulate a roadmap for insect conservation and recovery. Nat. Ecol. Evol. 2020. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
 Habitat (Effect Shown for Non-Water Body Sites) (Effect Size ± SE) |  NDVI 500 m (Effect Size ± SE) |  VNIR 500 m (Effect Size ± SE) |  Temperature (Effect Size ± SE) |  Moon(Effect Size ± SE) | Deviance Reduction (%) | |
|---|---|---|---|---|---|---|
| Coleoptera abundance | −0.25 ± 0.40 | 0.57 ** ± 0.21 | 0.20 ± 0.21 | 0.90 ± 0.02 | 0.47 ± 0.06 | 52.7 |
| Diptera abundance | −1.69 *** ± 0.31 | 0.33 * ± 0.16 | 0.20 ± 0.15 | 0.42 *** ± 0.02 | 0.80 *** ± 0.06 | 39.7 |
| Lepidoptera abundance | −0.10 ± 0.33 | 0.31 ± 0.17 | 0.08 ± 0.17 | 0.32 *** ± 0.02 | 0.38 *** ± 0.04 | 39.0 |
| Trichoptera abundance | −2.60 *** ± 0.50 | NA | 0.14 ± 0.24 | 0.94 *** ± 0.03 | −0.27 *** ± 0.07 | 22.9 |
| Total insect abundance | −0.82 * ± 0.36 | 0.50 ** ± 0.18 | 0.23 ± 0.18 | 0.62 *** ± 0.01 | 0.13 *** ± 0.02 | 15.3 |
| Insect order richness | −0.08 ± 0.08 | 0.15 *** ± 0.04 | 0.04 ± 0.04 | 0.12 ± 0.04 | 0.16 ± 0.08 | 28.2 |
 Tree cover (Effect size ± SE) |  Understorey vegetation (Effect size ± SE) |  Size (Effect size ± SE) |  SQQ (Effect size ± SE) |  Temperature (Effect size ± SE) |  Moon (Effect size ± SE) | AICc | Deviance reduction (%) | |
|---|---|---|---|---|---|---|---|---|
| Coleoptera abundance | 0.48 * ± 0.22 | 0.41 ± 0.21 | NA | −0.28 ± 0.21 | 0.97 *** ± 0.03 | 0.70 *** ± 0.06 | 2964.6 | 61.4 |
| Diptera abundance | NA | NA | NA | 0.05 ± 0.16 | 0.47 *** ± 0.02 | 0.97 *** ± 0.05 | 3861.8 | 38.6 |
| Lepidoptera abundance | 0.30 ± 0.15 | NA | NA | −0.12 ± 0.14 | 0.38 *** ± 0.02 | 1.01 *** ± 0.05 | 4201.3 | 55.7 |
| Trichoptera abundance | NA | NA | 0.66 * ± 0.27 | −0.16 ± 0.27 | 0.91 ± 0.03 | −0.20 ± 0.07 | 3201.3 | 44.6 |
| Total insect abundance | NA | NA | NA | −0.04 ± 0.15 | 0.69 *** ± 0.01 | 0.40 *** ± 0.02 | 13687.7 | 52.9 |
| Insect order richness | 0.07 ± 0.05 | NA | NA | −0.12 * ± 0.05 | 0.17 *** ± 0.04 | 0.12 ± 0.09 | 552.8 | 45.4 |
| Diptera (Effect Size ± SE) | Coleoptera (Effect Size ± SE) | Lepidoptera (Effect Size ± SE) | Trichoptera (Effect Size ± SE) | Insect Order Richness (Effect Size ± SE) | Habitat (Effect Size ± SE) | NDVI (500 m) (Effect Size ± SE) | VNIR (500 m) (Effect Size ± SE) | Deviance Reduction (%) | |
|---|---|---|---|---|---|---|---|---|---|
| Austronomus australis | + *** | - *** | + *** | + *** | - *** | + | + *** | - | 44.0 |
| Chalinolobus gouldii | + *** | - *** | + *** | - *** | + *** | - | + | + | 29.4 |
| Chalinolobus morio | - * | + *** | + * | - *** | - *** | - | + *** | - | 36.0 |
| Vespadelus darlingtoni | - *** | NA | - *** | + *** | + *** | - | + ** | - | 40.6 |
| Vespadelus regulus | NA | - * | NA | + * | + *** | - | + ** | - | 33.3 |
| Vespadelus vulturnus | - *** | + *** | + *** | - *** | - * | - | + *** | - | 32.6 |
| Total bat activity | + *** | - * | + *** | - *** | + *** | - * | + ** | - | 26.1 |
| Bat species richness | + | NA | + | - | + | - | + *** | + | 25.1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Straka, T.M.; Lentini, P.E.; Lumsden, L.F.; Buchholz, S.; Wintle, B.A.; van der Ree, R. Clean and Green Urban Water Bodies Benefit Nocturnal Flying Insects and Their Predators, Insectivorous Bats. Sustainability 2020, 12, 2634. https://doi.org/10.3390/su12072634
Straka TM, Lentini PE, Lumsden LF, Buchholz S, Wintle BA, van der Ree R. Clean and Green Urban Water Bodies Benefit Nocturnal Flying Insects and Their Predators, Insectivorous Bats. Sustainability. 2020; 12(7):2634. https://doi.org/10.3390/su12072634
Chicago/Turabian StyleStraka, Tanja M., Pia E. Lentini, Linda F. Lumsden, Sascha Buchholz, Brendan A. Wintle, and Rodney van der Ree. 2020. "Clean and Green Urban Water Bodies Benefit Nocturnal Flying Insects and Their Predators, Insectivorous Bats" Sustainability 12, no. 7: 2634. https://doi.org/10.3390/su12072634
APA StyleStraka, T. M., Lentini, P. E., Lumsden, L. F., Buchholz, S., Wintle, B. A., & van der Ree, R. (2020). Clean and Green Urban Water Bodies Benefit Nocturnal Flying Insects and Their Predators, Insectivorous Bats. Sustainability, 12(7), 2634. https://doi.org/10.3390/su12072634