Response of Bacterial Community Structure to Different Biochar Addition Dosages in Karst Yellow Soil Planted with Ryegrass and Daylily

Abstract
1. Introduction
2. Materials and Methods
2.1. Soil and Pretreatment
2.2. Experimental Design
2.3. Soil Chemical Properties
2.4. Soil DNA Extraction and PCR (Polymerase Chain Reaction) Amplification
2.5. Sequence Processing and Bioinformatics Analysis
2.6. Statistical Analysis
3. Results
3.1. Soil Chemical Properties
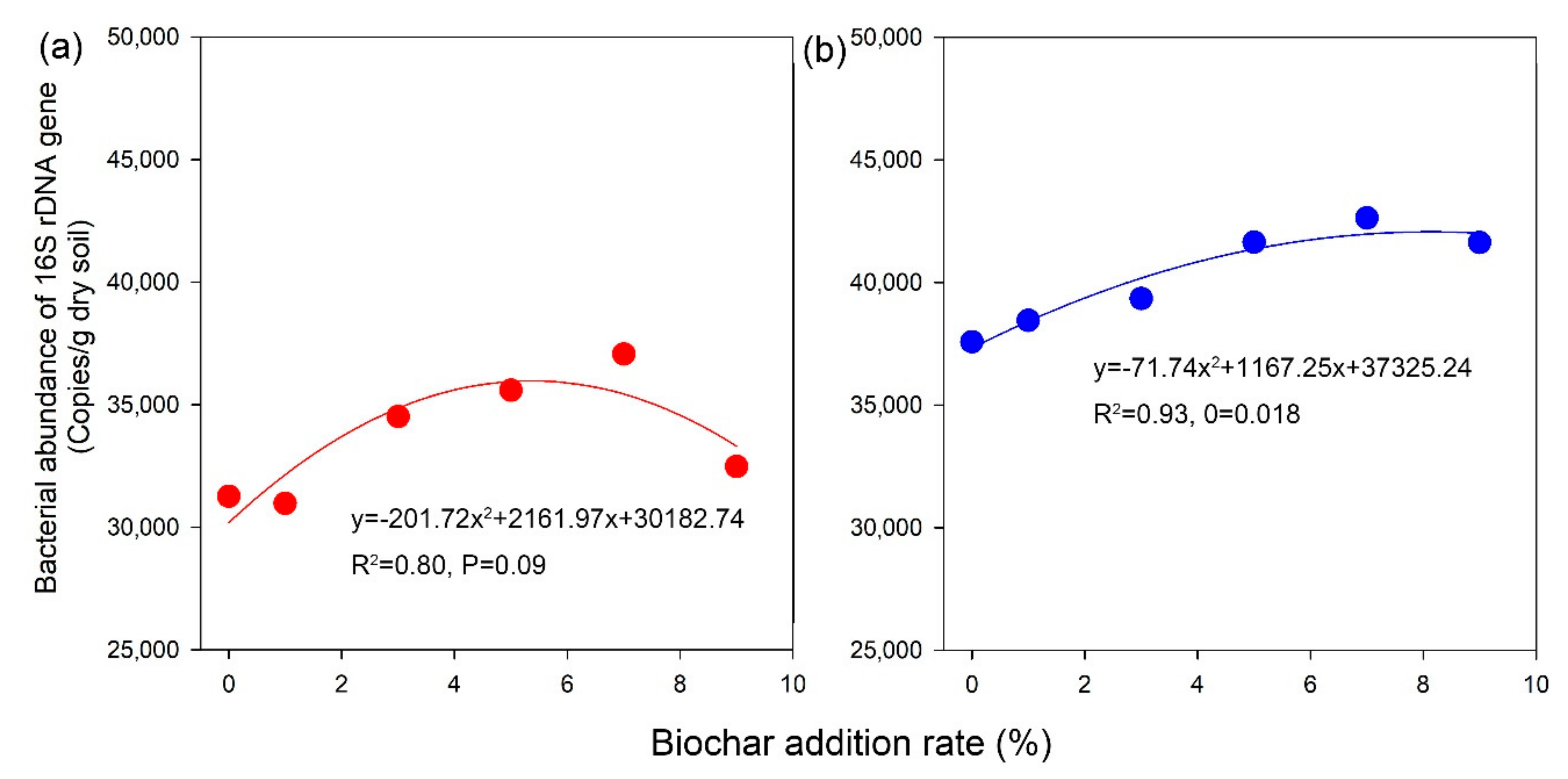
3.2. Soil Bacterial Abundance
3.3. Soil Bacterial Community Diversity
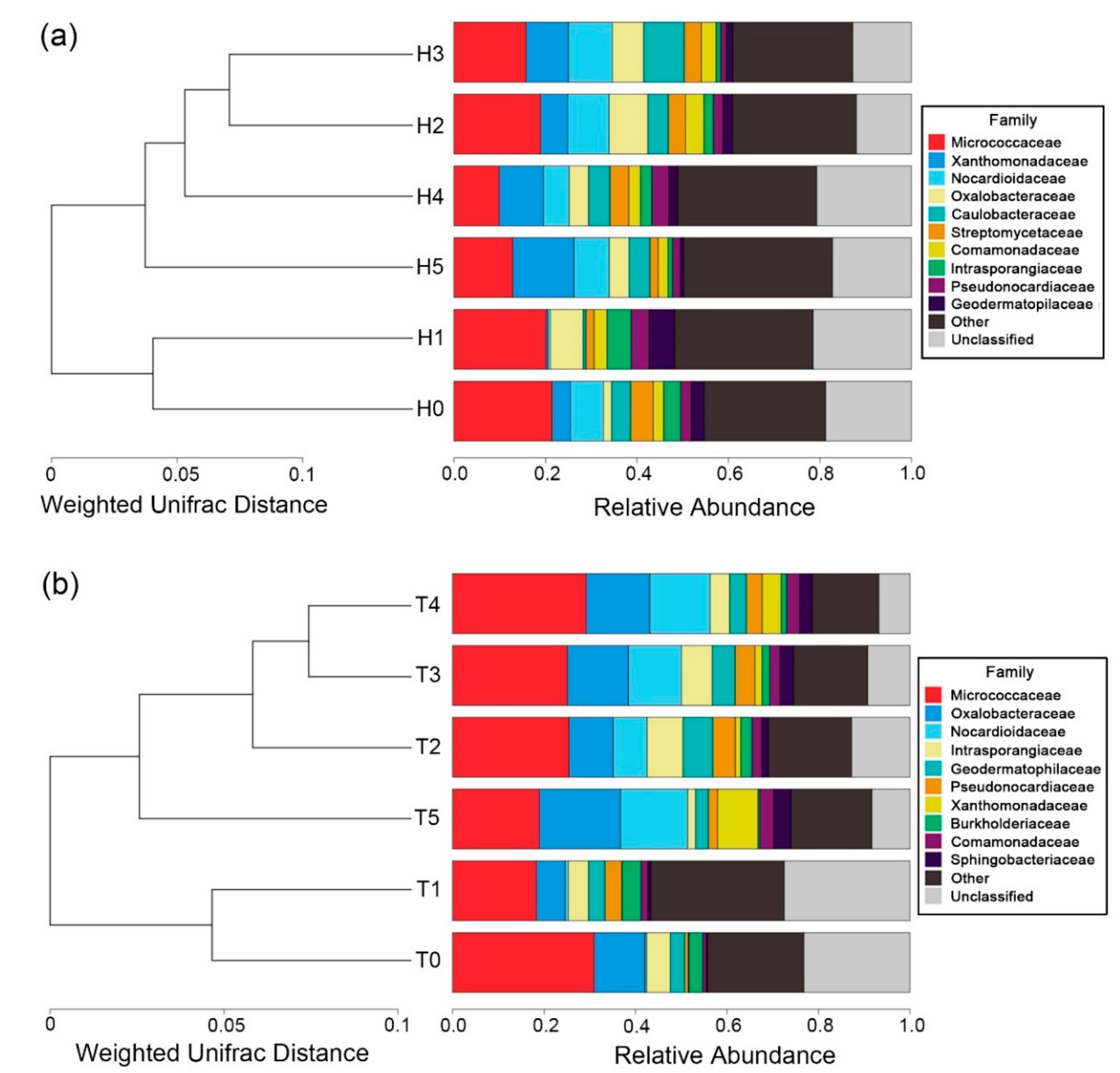
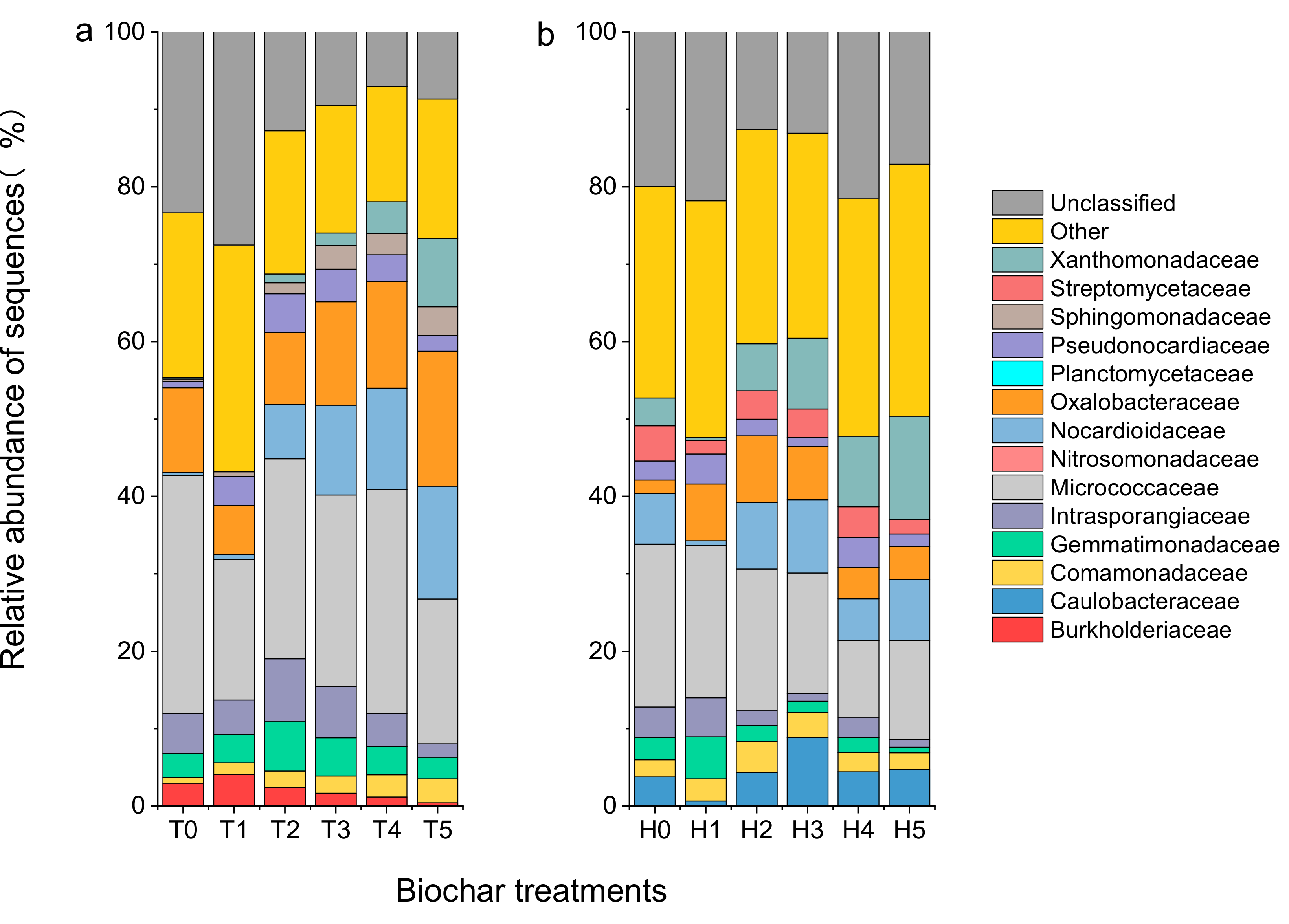
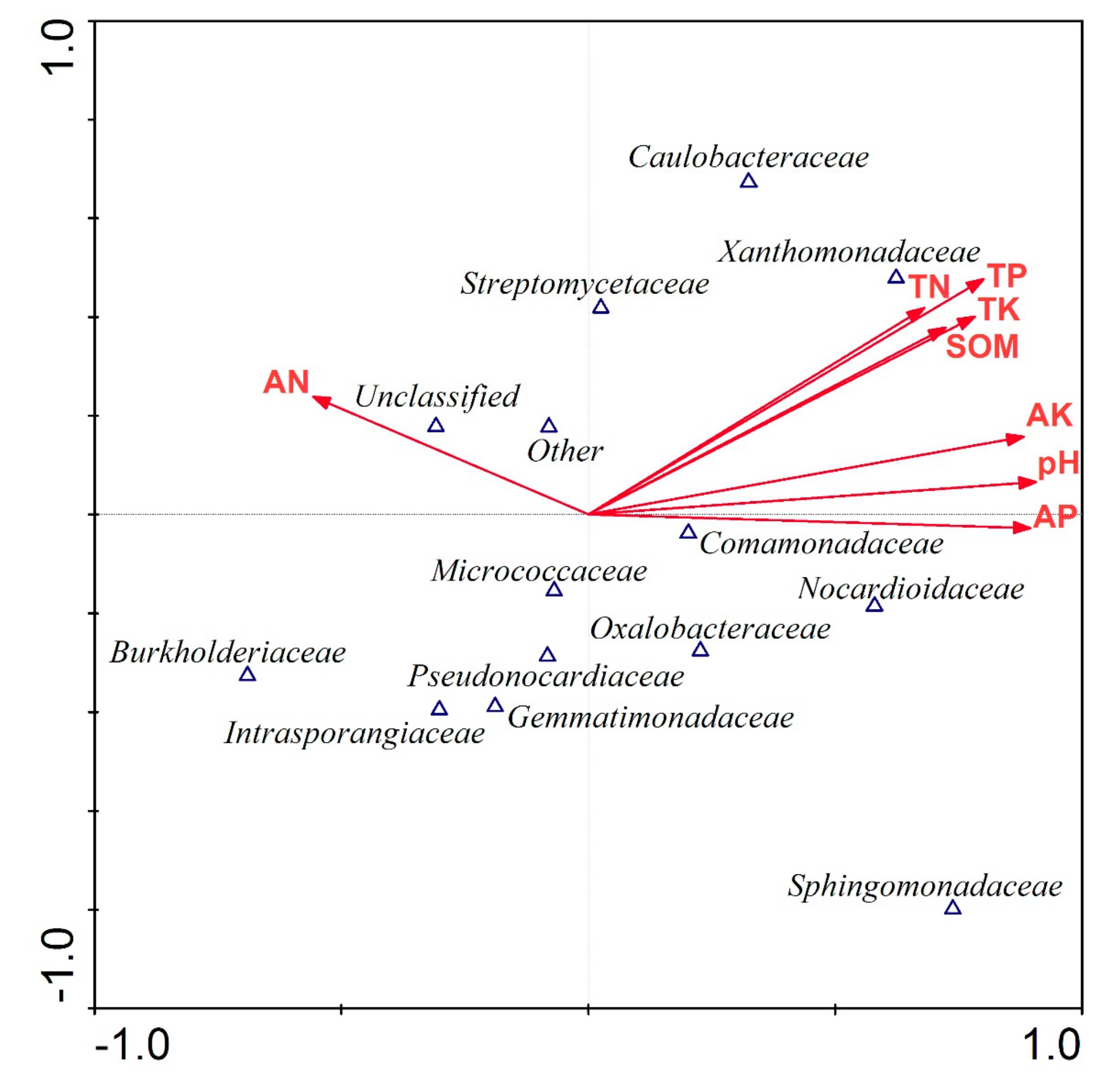
3.4. Soil Bacterial Community Structure
4. Discussion
4.1. Effect of Biochar Addition on Soil Chemical Properties
4.2. Effect of Biochar Addition on Soil Bacterial Abundance
4.3. Effect of Biochar Addition on Soil Bacterial Community Structure
4.4. Effect of Biochar Addition on Plants and Bacterial Community Composition
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- O’Laughlin, J.; Mcelligott, K. Biochar for environmental management: Science and technology; Lehman, J., Joseph, S., Eds.; Earthscan: London, UK, 2009; pp. 535–536. [Google Scholar]
- International Biochar Initiative. Standardized product definition and product testing guidelines for biochar that is used in soil; International Biochar Initiative: Canandaigua, NY, US, 2012. [Google Scholar]
- Jeffery, S.; Verheijen, F.G.A.; Velde, M.V.D.; Bastos, A.C.A. Quantitative review of the effects of biochar application to soils on crop productivity using meta-analysis. Agric. Ecosyst. Environ. 2011, 144, 175–187. [Google Scholar] [CrossRef]
- Lehmann, J.; Rillig, M.C.; Thies, J.; Masiello, C.A.; Hockaday, W.C.; Crowley, D. Biochar effects on soil biota-a review. Soil Biol. Biochem. 2011, 43, 1812–1836. [Google Scholar] [CrossRef]
- Park, J.H.; Ok, Y.S.; Kim, S.H.; Kang, S.W.; Cho, J.S.; Heo, J.S.; Delaune, R.D.; Seo, D.C. Characteristics of biochars derived from fruit tree pruning wastes and their effects on lead adsorption. J. Korean Soc. Appl. Biol. Chem. 2015, 58, 751–760. [Google Scholar] [CrossRef]
- Rajapaksha, A.; Ahmad, M.; Vithanage, M.; Kim, K.R.; Chang, J.; Lee, S.; Ok, Y.S. The role of biochar, natural iron oxides, and nanomaterials as soil amendments for immobilizing metals in shooting range soil. Environ. Geochem. Health 2015, 37, 931–942. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, J. Bio-energy in the black. Front Ecol. Environ. 2007, 5, 381–387. [Google Scholar] [CrossRef]
- Shrestha, G.; Traina, S.J.; Swanston, C.W. Black carbon’s properties and role in the environment: A comprehensive review. Sustainability 2010, 2, 294–320. [Google Scholar] [CrossRef]
- Major, J.; Rondon, M.; Molina, D.; Riha, S.J.; Lehmann, J. Maize yield and nutrition during 4 years after biochar application to a Colombian savanna oxisol. Plant Soil 2010, 333, 117–128. [Google Scholar] [CrossRef]
- Nguyen, T.T.N.; Wallace, H.M.; Xu, C.Y.; Zwieten, L.; Weng, Z.H.; Xu, Z.; Che, R.; Tahmasbian, I.; Hu, H.W.; Bai, S.H. The effects of short term, long term and reapplication of biochar on soil bacteria. Sci. Total Environ. 2018, 636, 142–151. [Google Scholar] [CrossRef]
- Beluri, K.; Laxma, R.P.V.; Bojeong, K.; Soo, L.S.; Kumar, P.S.; Ki-Hyun, K. Benefits and limitations of biochar amendment in agricultural soils: A review. J. Environ. Manag. 2018, 227, 146–154. [Google Scholar]
- Devare, M.; Londono, R.L.M.; Thies, J.E. Neither transgenic Bt maize (MON863) nor tefluthrin insecticide adversely affect soil microbial activity or biomass: A 3-year field analysis. Soil Biol. Biochem. 2007, 39, 2038–2047. [Google Scholar] [CrossRef]
- Kolton, M.; Harel, Y.M.; Pasternak, Z.; Graber, E.R.; Elad, Y.; Cytryn, E. Impact of biochar application to soil on the root-associated bacterial community structure of fully developed greenhouse pepper plants. Appl. Environ. Microbiol. 2011, 77, 4924–4930. [Google Scholar] [CrossRef] [PubMed]
- Joseph, S.; Xu, C.; Wallace, H.; Farrar, M.; Nguyen, T.N.; Bai, S.; Solaiman, Z. Biochar Production from Agricultural and Forestry Wastes and Microbial Interactions. In Current Developments in Biotechnology and Bioengineering: Solid Waste Management; Wong, J.W.C., Tyagi, R.D., Pandey, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 443–473. [Google Scholar]
- Spokas, K.A.; Novak, J.M.; Stewart, C.E.; Cantrell, K.B.; Uchimiya, M.; DuSaire, M.G.; Ro, K.S. Qualitative analysis of volatile organic compounds on biochar. Chemosphere 2011, 85, 869–882. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Chen, B.; Zhu, L.; Xing, B. Effects and mechanisms of biochar-microbe interactions in soil improvement and pollution remediation: A review. Environ. Pollut. 2017, 227, 98–115. [Google Scholar] [CrossRef]
- Wang, X.B.; Song, D.L.; Liang, G.Q.; Zhang, Q.; Ai, C.; Zhou, W. Maize biochar addition rate influences soil enzyme activity and microbial community composition in a fluvoaquic soil. Appl. Soil Ecol. 2015, 96, 265–272. [Google Scholar] [CrossRef]
- Graber, E.R.; Harel, Y.M.; Kolton, M.; Cytryn, E.; Silber, A.; David, D.R.; Tsechansky, L.; Borenshtein, M.; Elad, Y. Biochar impact on development and productivity of pepper and tomato grown in fertigated soilless media. Plant Soil 2010, 337, 481–496. [Google Scholar] [CrossRef]
- Quilliam, R.S.; Glanville, H.C.; Wade, S.C.; Jones, D.L. Life in the ‘charosphere’ - does biochar in agricultural soil provide a significant habitat for microorganisms? Soil Biol. Biochem. 2013, 65, 287–293. [Google Scholar] [CrossRef]
- Yu, J.; Deem, L.M.; Crow, S.E.; Deenik, J.L.; Penton, C.R. Biochar application influences microbial assemblage complexity and composition due to soil and bioenergy crop type interactions. Soil Biol. Biochem. 2018, 117, 97–107. [Google Scholar] [CrossRef]
- Chen, J.H.; Liu, X.Y.; Zheng, J.W.; Zhang, B.; Lu, H.F.; Chi, Z.Z.; Pan, G.X.; Li, L.Q.; Zheng, J.F.; Zhang, X.H.; et al. Biochar soil amendment increased bacterial but decreased fungal gene abundance with shifts in community structure in a slightly acid rice paddy from Southwest China. Appl. Soil. Ecol. 2013, 71, 33–44. [Google Scholar] [CrossRef]
- Gul, S.; Whalen, J.K.; Thomas, B.W.; Sachdeva, V.; Deng, H. Physico-chemical properties and microbial responses in biochar-amended soils: Mechanisms and future directions. Agric. Ecosyst. Environ. 2015, 206, 46–59. [Google Scholar] [CrossRef]
- Lu, H.F.; Lashari, M.S.; Liu, X.Y.; Ji, H.S.; Li, L.Q.; Zheng, J.F.; Kibue, G.W.; Joseph, S.; Pan, G.X. Changes in soil microbial community structure and enzyme activity with amendment of biochar-manure compost and pyroligneous solution in a saline soil from central China. Eur. J. Soil Biol. 2015, 70, 67–76. [Google Scholar] [CrossRef]
- Hu, L.; Cao, L.X.; Zhang, R.D. Bacterial and fungal taxon changes in soil microbial community composition induced by short-term biochar amendment in red oxidized loam soil. World J. Microb. Biot. 2014, 30, 1085–1092. [Google Scholar] [CrossRef] [PubMed]
- Hale, L.; Luth, M.; Crowley, D. Biochar characteristics relate to its utility as an alternative soil inoculum carrier to peat and vermiculite. Soil Biol. Biochem. 2015, 81, 228–235. [Google Scholar] [CrossRef]
- Rutigliano, F.A.; Romano, M.; Marzaioli, R.; Baglivo, I.; Baronti, S.; Miglietta, F.; Castaldi, S. Effect of biochar addition on soil microbial community in a wheat crop. Eur. J. Soil Biol. 2014, 60, 9–15. [Google Scholar] [CrossRef]
- Manolikaki, I.; Diamadopoulos, E. Ryegrass yield and nutrient status after biochar application in two Mediterranean soils. Arch. Agro. Soil Sci. 2016, 63, 1093–1107. [Google Scholar] [CrossRef]
- Liu, C.C.; Liu, Y.G.; Guo, K.; Wang, S.J.; Liu, H.M.; Zhao, H.W.; Qiao, X.G.; Hou, D.J.; Li, S.B. Aboveground carbon stock, allocation and sequestration potential during vegetation recovery in the karst region of southwestern China: A case study at a watershed scale. Agric. Ecosyst. Environ. 2016, 235, 91–100. [Google Scholar] [CrossRef]
- Bai, X.Y.; Wang, S.J.; Xiong, K.N. Assessing spatial-temporal evolution processes of karst rocky desertification land: Indications for restoration strategies. Land Degrad. Dev. 2013, 24, 47–56. [Google Scholar] [CrossRef]
- Yan, Y.J.; Dai, Q.H.; Yuan, Y.F.; Peng, X.D.; Zhao, L.S.; Yang, J. Effects of rainfall intensity on runoff and sediment yields on bare slopes in a karst area, SW China. Geoderma 2018, 330, 30–40. [Google Scholar] [CrossRef]
- Zhang, X.Q.; Hu, M.C.; Guo, X.Y.; Yang, H.; Zhang, Z.K.; Zhang, K.L. Effects of topographic factors on runoff and soil loss in Southwest China. Catena 2018, 160, 394–402. [Google Scholar] [CrossRef]
- Carlson, J.; Saxena, J.; Basta, N.; Hundal, L.; Busalacchi, D.; Dick, R.P. Application of organic amendments to restore degraded soil: Effects on soil microbial properties. Environ. Monit. Assess. 2015, 187, 1–15. [Google Scholar] [CrossRef]
- Dai, Q.; Peng, X.; Yang, Z.; Zhao, L. Runoff and erosion processes on bare slopes in the karst rocky desertification area. Catena 2017, 152, 218–226. [Google Scholar] [CrossRef]
- El-Naggar, A.; Lee, S.S.; Rinklebe, J.; Farooq, M.; Song, H.; Sarmah, A.K.; Zimmerman, A.R.; Ahmad, M.; Shaheen, S.M.; Ok, Y.S. Biochar application to low fertility soils: A review of current status, and future prospects. Geoderma 2019, 337, 536–554. [Google Scholar] [CrossRef]
- Prendergast-Miller, M.T.; Duvall, M.; Sohi, S.P. Biochar-root interactions are mediated by biochar nutrient content and impacts on soil nutrient availability. Eur. J. Soil Sci. 2014, 65, 173–185. [Google Scholar] [CrossRef]
- Yu, H.W.; Zou, W.X.; Chen, J.J.; Chen, H.; Yu, Z.B.; Huang, J.; Tang, H.R.; Wei, X.Y.; Gao, B. Biochar amendment improves crop production in problem soils: A review. J. Environ. Manag. 2019, 232, 8–21. [Google Scholar] [CrossRef] [PubMed]
- Verheijen, F.G.A.; Jeffery, S.; Bastos, A.C.; Velde, M.V.D.; Diafas, I. Biochar application to soils: A critical scientific review of effects on soil properties, processes and functions. In EUR 24099 EN; Office for the Official Publications of the European Communities: Luxembourg, 2009. [Google Scholar]
- Sun, C.X.; Chen, X.; Cao, M.; Li, Q.; Zhang, Y.L. Growth and metabolic responses of maize roots to straw biochar application at different rates. Plant Soil 2017, 416, 487–502. [Google Scholar] [CrossRef]
- Gale, N.V.; Thomas, S.C. Dose-dependence of growth and ecophysiological responses of plants to biochar. Sci. Total Environ. 2019, 658, 1344–1354. [Google Scholar] [CrossRef]
- Lu, R.K. Analytical Methods of Soil Agrochemistry; China Agricultural Science and Technology Press: Beijing, China, 1999. (In Chinese) [Google Scholar]
- Institute of Soil Sciences, Chinese Academy of Sciences (ISSCAS). Physical and Chemical Analysis Methods of Soils; Shanghai Science Technology Press: Shanghai, China, 1978. (In Chinese) [Google Scholar]
- Miranda, K.M.; Espey, M.G.; Wink, D.A. A rapid, simple spectrophotometric method for simultaneous detection of nitrate and nitrite. Nitric. Oxide 2001, 5, 62–71. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996. [Google Scholar] [CrossRef]
- R Development Core Team. R: A language and environment for statistical computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glöckner, F.O. SILVA: A comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Knight, R. Qiime allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Brewer, C.E.; Brown, R.C. Biochar. In Comprehensive Renewable Energy; Sayigh, A., Ed.; Elsevier: Oxford, UK, 2012; pp. 357–384. [Google Scholar]
- Lal, R. Soils and food sufficiency: A review. Agron. Sustain. Dev. 2009, 29, 113–133. [Google Scholar] [CrossRef]
- Zheng, H.; Wang, Z.Y.; Deng, X.; Herbert, S.; Xing, B.S. Impacts of adding biochar on nitrogen retention and bioavailability in agricultural soil. Geoderma 2013, 206, 32–39. [Google Scholar] [CrossRef]
- Liu, L.; Wang, Y.; Yan, X.; Li, J.; Jiao, N.; Hu, S. Biochar amendments increase the yield advantage of legume-based intercropping systems over monoculture. Agric. Ecosyst. Environ. 2017, 237, 16–23. [Google Scholar] [CrossRef]
- Arif, M.; Ilyas, M.; Riaz, M.; Ali, K.; Shah, K.; Ul Haq, I.; Fahad, S. Biochar improves phosphorus use efficiency of organic-inorganic fertilizers, maize-wheat productivity and soil quality in a low fertility alkaline soil. Field Crop. Res. 2017, 214, 25–37. [Google Scholar] [CrossRef]
- Zhou, Z.D.; Gao, T.; Zhu, Q.; Yan, T.T.; Li, D.C.; Xue, J.H.; Wu, Y.B. Increases in bacterial community network complexity induced by biochar-based fertilizer amendments to karst calcareous soil. Geoderma 2019, 337, 691–700. [Google Scholar] [CrossRef]
- Pietikäinen, J.; Kiikkilä, O.; Fritze, H. Charcoal as a habitat for microbes and its effects on the microbial community of the underlying humus. Oikos 2000, 89, 231–242. [Google Scholar] [CrossRef]
- Smith, J.L.; Collins, H.P.; Bailey, V.L. The effect of young biochar on soil respiration. Soil Biol. Biochem. 2010, 42, 2345–2347. [Google Scholar] [CrossRef]
- Pietri, J.C.A.; Brookes, P.C. Relationships between soil pH and microbial properties in a UK arable soil. Soil Biol. Biochem. 2008, 40, 1856–1861. [Google Scholar] [CrossRef]
- Ameloot, N.; Graber, E.R.; Verheijen, F.G.A.; DeNeve, S. Interactions between biochar stability and soil organisms: Review and research needs. Eur. J. Soil. Sci. 2013, 64, 379–390. [Google Scholar] [CrossRef]
- Li, C.J.; Xiong, Y.W.; Qu, Z.Y.; Xu, X.; Huang, Q.Z.; Huang, G.H. Impact of biochar addition on soil properties and water-fertilizer productivity of tomato in semi-arid region of Inner Mongolia, China. Geoderma 2018, 331, 100–108. [Google Scholar] [CrossRef]
- Gomez, J.D.; Denef, K.; Stewart, C.E.; Zheng, J.; Cotrufo, M.F. Biochar addition rate influences soil microbial abundance and activity in temperate soils. Eur. J. Soil Sci. 2014, 65, 28–39. [Google Scholar] [CrossRef]
- Mitchell, P.J.; Simpson, A.J.; Soong, R.; Simpson, M.J. Shifts in microbial community and water-extractable organic matter composition with biochar amendment in a temperate forest soil. Soil Biol. Biochem. 2015, 81, 244–254. [Google Scholar] [CrossRef]
- Zhang, C.S.; Lin, Y.; Tian, X.Y.; Xu, Q.; Chen, Z.H.; Lin, W. Tobacco bacterial wilt suppression with biochar soil addition associates to improved soil physiochemical properties and increased rhizosphere bacteria abundance. Appl. Soil Ecol. 2017, 112, 90–96. [Google Scholar] [CrossRef]
- O’Neil, B.; Grossman, J.; Tsai, M.T.; Gomes, J.E.; Lehmann, J.; Peterson, J.; Neves, E.; Thies, J.E. Bacterial community composition in Brazilian Anthrosols and adjacent soils characterized using culturing and molecular identification. Microb. Ecol. 2009, 58, 23–35. [Google Scholar] [CrossRef]
- Prayogo, C.; Jones, J.E.; Baeyens, J.; Bending, G.D. Impact of biochar on mineralisation of C and N from soil and willow litter and its relationship with microbial community biomass and structure. Biol. Fertil. Soils 2014, 50, 695–702. [Google Scholar] [CrossRef]
- Xu, N.; Tan, G.C.; Wang, H.Y.; Gai, X.P. Effect of biochar additions to soil on nitrogen leaching, microbial biomass and bacterial community structure. Eur. J. Soil Biol. 2016, 74, 1–8. [Google Scholar] [CrossRef]
- Ren, C.J.; Wang, T.; Xu, Y.D.; Deng, J.; Zhao, F.Z.; Yang, G.H.; Han, X.H.; Feng, Y.Z.; Ren, G.X. Differential soil microbial community responses to the linkage of soil organic carbon fractions with respiration across land-use changes. For. Ecol. Manag. 2018, 409, 170–178. [Google Scholar] [CrossRef]
- Jones, D.L.; Murphy, D.V.; Khalid, M.; Ahmad, W.; Edwards-Jones, G.; DeLuca, T.H. Short-term biochar-induced increase in soil CO2 release is both biotically and abiotically mediated. Soil Biol. Biochem. 2011, 43, 1723–1731. [Google Scholar] [CrossRef]
- Jones, D.L.; Edwards-Jones, G.; Murphy, D.V. Biochar mediated alterations in herbicide breakdown and leaching in soil. Soil Biol. Biochem. 2011, 43, 804–813. [Google Scholar] [CrossRef]
- McCormack, S.A.; Ostle, N.; Bardgett, R.D.; Hopkins, D.W.; Vanbergen, A.J. Biochar in bioenergy cropping systems:impacts on soil faunal communities and linked ecosystem processes. Gcb. Bioenergy 2013, 5, 81–95. [Google Scholar] [CrossRef]
- Paz-Ferreiro, J.; Fu, S.L. Biological indices for soil quality evaluation: Perspectives and limitations. Land Degrad. Dev. 2016, 27, 14–25. [Google Scholar] [CrossRef]
- Sheng, Y.Q.; Zhu, L.Z. Biochar alters microbial community and carbon sequestration potential across different soil pH. Sci. Total Environ. 2018, 622, 1391–1399. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Y.; Ye, Y.X.; Liu, Y.M.; Zhang, A.F.; Zhang, X.H.; Li, L.Q.; Pan, G.X.; Kibue, G.W.; Zheng, J.F.; Zheng, J.W. Sustainable biochar effects for low carbon crop production: A 5-crop season field experiment on a low fertility soil from Central China. Agric. Syst. 2014, 129, 22–29. [Google Scholar] [CrossRef]
- Nielsen, S.; Minchin, T.; Kimber, S.; Van Zwieten, L.; Gilbert, J.; Munroe, P.; Joseph, S.; Thomas, T. Comparative analysis of the microbial communities in agricultural soil amended with enhanced biochars or traditional fertilisers. Agric. Ecosyst. Environ. 2014, 191, 73–82. [Google Scholar] [CrossRef]
- Shaaban, M.; Zwieten, L.V.; Bashir, S.; Younas, A.; Núñez-Delgado, A.; Chhajro, M.A.; Kubar, K.A.; Ali, U.; Rana, M.S.; Mehmood, M.A.; et al. A concise review of biochar application to agricultural soils to improve soil conditions and fight pollution. J. Environ. Manag. 2018, 228, 429–440. [Google Scholar] [CrossRef]
- Akhter, A.; Hage-Ahmed, K.; Soja, G.; Steinkellner, S. Compost and biochar alter mycorrhization, tomato root exudation, and development of Fusarium oxysporum f. sp. lycopersici. Front. Plant Sci. 2015, 6, 529. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | TP g kg−1 | AP g kg−1 | TK g kg−1 | AK g kg−1 | CEC cmol kg−1 | TCa g kg−1 | TMg g kg−1 | Surface Area m2 g−1 |
|---|---|---|---|---|---|---|---|---|
| 10.09 ± 0.05 | 16.17 ± 0.19 | 0.20 ± 0.01 | 60.11 ± 0.31 | 16.55 ± 0.07 | 10.26 ± 0.39 | 37.88 ± 1.16 | 11.43 ± 0.16 | 17.20 ± 1.31 |
| Treatments | pH | SOM g kg−1 | TN g kg−1 | AN g kg−1 | TP g kg−1 | AP cmol kg−1 | TK g kg−1 | AK g kg−1 |
|---|---|---|---|---|---|---|---|---|
| T0 (CK) | 5.42 ± 0.02e | 5.88 ± 0.75d | 0.52 ± 0.01c | 38.25 ± 2.79a | 0.19 ± 0.01f | 0.34 ± 0.05f | 20.4 ± 0.15c | 0.05 ± 0.01f |
| T1 | 5.49 ± 0.06e | 7.71 ± 0.23d | 0.44 ± 0.02d | 26.02 ± 2.31b | 0.21 ± 0.01e | 0.76 ± 0.09e | 20.2 ± 0.1cd | 0.42 ± 0.01e |
| T2 | 6.00 ± 0.02d | 7.98 ± 0.61d | 0.55 ± 0.01c | 22.40 ± 2.41bc | 0.21 ± 0.01d | 1.07 ± 0.05d | 19.9 ± 0.22d | 1.06 ± 0.02d |
| T3 | 7.31 ± 0.02c | 22.39 ± 1.14c | 0.70 ± 0.01b | 21.05 ± 0.55bc | 0.25 ± 0.01c | 1.83 ± 0.05c | 22.2 ± 0.31b | 2.06 ± 0.05c |
| T4 | 7.57 ± 0.04b | 29.99 ± 0.68a | 1.03 ± 0.01a | 20.59 ± 3.24c | 0.38 ± 0.02a | 2.13 ± 0.09b | 22.4 ± 0.14b | 2.40 ± 0.31b |
| T5 | 7.90 ± 0.04a | 25.13 ± 2.18b | 0.69 ± 0.01b | 21.72 ± 2.54bc | 0.34 ± 0.01b | 2.32 ± 0.11a | 23.8 ± 0.27a | 3.57 ± 0.08a |
| H0 (CK) | 5.45 ± 0.02e | 3.25 ± 0.32f | 0.50 ± 0.01f | 52.27 ± 0.56a | 0.23 ± 0.06c | 0.34 ± 0.05d | 19.4 ± 0.06c | 0.06 ± 0.01f |
| H1 | 5.48 ± 0.06e | 8.47 ± 0.95e | 0.60 ± 0.03e | 46.16 ± 0.01a | 0.23 ± 0.01c | 0.87 ± 0.09c | 20.0 ± 0.10c | 0.51 ± 0.07e |
| H2 | 6.24 ± 0.02d | 16.37 ± 0.90d | 0.71 ± 0.01d | 24.43 ± 1.11b | 0.29 ± 0.02bc | 1.63 ± 0.14b | 20.7 ± 0.43c | 1.28 ± 0.03d |
| H3 | 7.17 ± 0.02c | 21.46 ± 2.87c | 0.81 ± 0.02c | 23.53 ± 6.23b | 0.33 ± 0.03b | 1.56 ± 0.09b | 23.4 ± 0.55b | 2.12 ± 0.03c |
| H4 | 7.44 ± 0.03b | 31.96 ± 0.74b | 0.88 ± 0.06b | 21.04 ± 2.77b | 0.32 ± 0.01b | 1.79 ± 0.09b | 23.3 ± 0.25b | 2.53 ± 0.07b |
| H5 | 7.76 ± 0.01a | 46.71 ± 2.17a | 1.22 ± 0.01a | 26.02 ± 2.09b | 0.46 ± 0.03a | 2.09 ± 0.19a | 25.2 ± 1.65a | 3.58 ± 0.05a |
| Sample ID | Bacterial Sequences | Effective Bacterial Sequences | Effective (%) | Number of Species | ACEc | Chao 1 Richness | Shannon’s Diversity | Simpson’s Diversity | Coverage (%) |
|---|---|---|---|---|---|---|---|---|---|
| T0 | 46,154 ± 3199 | 43,822 ± 2470 | 95.03 | 703 ± 102 | 886.29 ± 82.11nsb | 878 ± 91b | 5.72 ± 0.27b | 0.9385 ± 0.0106b | 99.57 |
| T1 | 44,976 ± 2410 | 43,761 ± 2512 | 97.28 | 657 ± 50 | 799.87 ± 79.75 | 788 ± 68aa | 6.46 ± 0.11a | 0.9727 ± 0.0033a | 99.65 |
| T2 | 43,317 ± 7064 | 40,795 ± 6620 | 94.25 | 639 ± 183 | 793.40 ± 209.10 | 797 ± 205ab | 6.00 ± 0.44ab | 0.9515 ± 0.0117ab | 99.60 |
| T3 | 44,017 ± 3616 | 40,557 ± 3517 | 92.11 | 662 ± 28 | 807.73 ± 30.99 | 830 ± 51ab | 6.00 ± 0.13ab | 0.9513 ± 0.0017ab | 99.59 |
| T4 | 45,186 ± 1675 | 41,522 ± 942 | 91.95 | 770 ± 80 | 946.15 ± 99.97 | 974 ± 91a | 5.93 ± 0.22ab | 0.9387 ± 0.0169ab | 99.51 |
| T5 | 39,326 ± 1526 | 36,783 ± 1427 | 93.54 | 687 ± 65 | 819.76 ± 79.48 | 823 ± 72ab | 6.24 ± 0.16ab | 0.9560 ± 0.0049ab | 99.57 |
| H0 | 53,187 ± 8790 | 48,863 ± 5699 | 92.63 | 1033 ± 175 | 1260.92 ± 220.03b | 1282 ± 193b | 6.64 ± 0.12ns | 0.9578 ± 0.0138ns | 99.45 |
| H1 | 60,186 ± 5879 | 56,018 ± 5218 | 93.11 | 1129 ± 87 | 1340.28 ± 96.76b | 1341 ± 84b | 6.65 ± 0.15 | 0.9594 ± 0.0073 | 99.52 |
| H2 | 58,624 ± 5592 | 52,736 ± 5727 | 89.85 | 1441 ± 49 | 1690.14 ± 68.64a | 1693 ± 67a | 7.09 ± 0.31 | 0.9616 ± 0.0112 | 99.34 |
| H3 | 53,098 ± 1299 | 49,754 ± 1606 | 93.69 | 1152 ± 49 | 1388.98 ± 67.24ab | 1408 ± 85ab | 7.01 ± 0.22 | 0.9664 ± 0.0061 | 99.44 |
| H4 | 53,083 ± 5527 | 50,094 ± 3767 | 94.68 | 1056 ± 181 | 1286.53 ± 238.19b | 1311 ± 233b | 7.08 ± 0.27 | 0.9773 ± 0.0057 | 99.48 |
| H5 | 58,344 ± 7146 | 56,106 ± 7622 | 96.02 | 918 ± 65 | 1128.02 ± 66.25b | 1135 ± 74b | 7.03 ± 0.11 | 0.9747 ± 0.0023 | 99.60 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, S.; He, B.; Song, D.; Li, T.; Wu, Y.; Yang, L. Response of Bacterial Community Structure to Different Biochar Addition Dosages in Karst Yellow Soil Planted with Ryegrass and Daylily. Sustainability 2020, 12, 2124. https://doi.org/10.3390/su12052124
Luo S, He B, Song D, Li T, Wu Y, Yang L. Response of Bacterial Community Structure to Different Biochar Addition Dosages in Karst Yellow Soil Planted with Ryegrass and Daylily. Sustainability. 2020; 12(5):2124. https://doi.org/10.3390/su12052124
Chicago/Turabian StyleLuo, Songping, Binghui He, Dandan Song, Tianyang Li, Yaopeng Wu, and Lei Yang. 2020. "Response of Bacterial Community Structure to Different Biochar Addition Dosages in Karst Yellow Soil Planted with Ryegrass and Daylily" Sustainability 12, no. 5: 2124. https://doi.org/10.3390/su12052124
APA StyleLuo, S., He, B., Song, D., Li, T., Wu, Y., & Yang, L. (2020). Response of Bacterial Community Structure to Different Biochar Addition Dosages in Karst Yellow Soil Planted with Ryegrass and Daylily. Sustainability, 12(5), 2124. https://doi.org/10.3390/su12052124