Sustainable Intensification? Increased Production Diminishes Omega-3 Content of Sheep Milk

 , ,
, ,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Systems
2.2. Experimental Design and Sampling Procedure
2.3. Measurements and Analytical Methods
2.4. Statistical Analyses
3. Results
3.1. Environmental Conditions
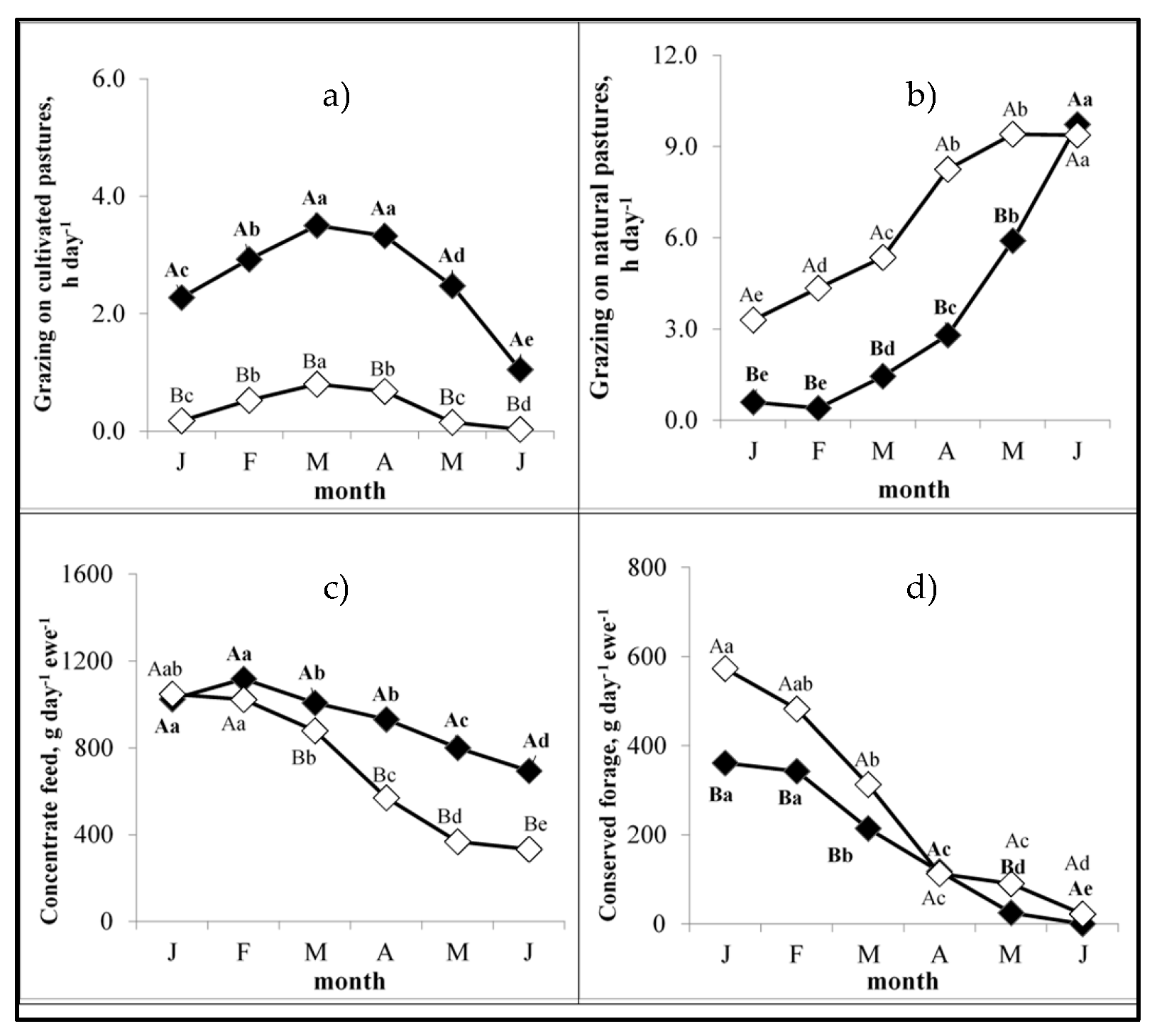
3.2. Feeding Regimes
3.3. Milk Yield and Basic Composition
3.4. Fatty Acid Profile
3.4.1. Major Saturated and Monounsaturated Fatty Acids.
3.4.2. Major Polyunsaturated Fatty Acids
3.4.3. Fatty Acid Groups and the Omega-6/Omega-3 Ratio
3.4.4. Odd-Chain Fatty Acids
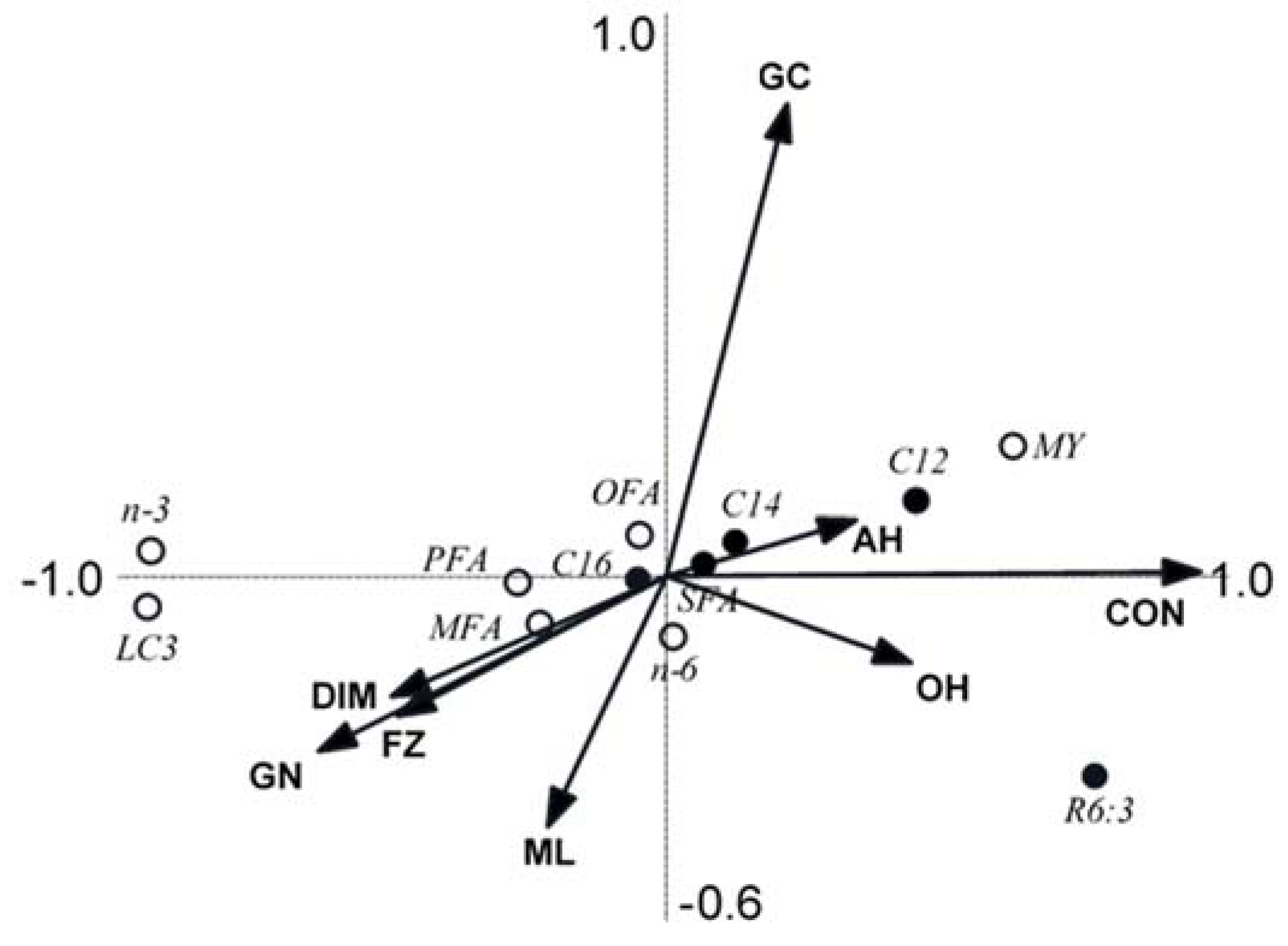
3.5. Redundancy Analyses
4. Discussion
4.1. Differences in Milk Yield and Composition
4.2. Possible Explanations
4.3. Potential Consequences
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- FAO. Proceedings of the Expert Meeting on How to Feed the World 2050; Food and Agriculture Organisation of the United Nations: Rome, Italy, 2009; ISBN 978-92-5-106363-7. [Google Scholar]
- Schader, C.; Baumgart, L.; Landert, J.; Muller, A.; Ssebunya, B.; Blockeel, J.; Weisshaidinger, R.; Petrasek, R.; Mészáros, D.; Padel, S.; et al. Using the Sustainability Monitoring and Assessment Routine (SMART) for the systematic analysis of trade-offs and synergies between sustainability dimensions and themes at farm level. Sustainability 2016, 8, 274. [Google Scholar] [CrossRef]
- Ripoll-Bosch, R.; Díez-Unquera, B.; Ruiz, R.; Villalba, D.; Molina, E.; Joy, M.; Olaizola, A.; Bernués, A. An integrated sustainability assessment of mediterranean sheep farms with different degrees of intensification. Agric. Syst. 2012, 105, 46–56. [Google Scholar] [CrossRef]
- FAO. Sustainable Diets and Biodiversity; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2012; ISBN 978-92-5-107288-2. [Google Scholar]
- Średnicka-Tober, D.; Barański, M.; Seal, C.; Sanderson, R.; Benbrook, C.; Steinshamn, H.; Gromadzka-Ostrowska, J.; Rembiałkowska, E.; Skwarło-Sońta, K.; Eyre, M.; et al. Composition differences between organic and conventional meat: A systematic literature review and meta-analysis. Br. J. Nutr. 2016, 115, 994–1011. [Google Scholar] [CrossRef] [PubMed]
- Barański, M.; Średnicka-Tober, D.; Volakakis, N.; Seal, C.; Sanderson, R.; Stewart, G.B.; Benbrook, C.; Biavati, B.; Markellou, E.; Giotis, C.; et al. Higher antioxidant and lower cadmium concentrations and lower incidence of pesticide residues in organically grown crops: A systematic literature review and meta-analyses. Br. J. Nutr. 2014, 112, 794–811. [Google Scholar] [CrossRef] [PubMed]
- Barański, M.; Rempelos, L.; Iversen, P.O.; Leifert, C. Effects of organic food consumption on human health; the jury is still out! Food Nutr. Res. 2017, 61, 1287333. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, B.; Du, Y.; Snetselaar, L.G.; Sun, Q.; Hu, F.B.; Bao, W. Inverse association between organic food purchase and diabetes mellitus in US adults. Nutrients 2018, 10, 1877. [Google Scholar] [CrossRef]
- Baudry, J.; Lelong, H.; Adriouch, S.; Julia, C.; Allès, B.; Hercberg, S.; Touvier, M.; Lairon, D.; Galan, P.; Kesse-Guyot, E. Association between organic food consumption and metabolic syndrome: Cross-sectional results from the NutriNet-Santé study. Eur. J. Nutr. 2018, 57, 2477–2488. [Google Scholar] [CrossRef]
- Sanchez-Villegas, A.; Martinez, J.A.; Prattala, R.; Toledo, E.; Roos, G.; Martinez-Gonzalez, M.A.; Group, F. A systematic review of socioeconomic differences in food habits in Europe: Consumption of cheese and milk. Eur. J. Clin. Nutition 2003, 57, 917–929. [Google Scholar] [CrossRef]
- Schaefer, E.J. Lipoproteins, nutrition, and heart disease. Am. J. Clin. Nutition 2002, 75, 191–212. [Google Scholar] [CrossRef]
- American Heart Association Nutrition Committee; Lichtenstein, A.H.; Appel, L.J.; Brands, M.; Carnethon, M.; Daniels, S.; Franch, H.A.; Franklin, B.; Kris-Etherton, P.; Harris, W.S.; et al. Diet and lifestyle recommendations revision 2006: A scientific statement from the American Heart Association Nutrition Committee. Circulation 2006, 114, 82–96. [Google Scholar]
- Elwood, P.C.; Givens, D.I.; Beswick, A.D.; Fehily, A.M.; Pickering, J.E.; Gallacher, J. The survival advantage of milk and dairy consumption: An overview of evidence from cohort studies of vascular diseases, diabetes and cancer. J. Am. Coll. Nutr. 2008, 27, 723S–734S. [Google Scholar] [CrossRef] [PubMed]
- Lordan, R.; Zabetakis, I. Invited review: The anti-inflammatory properties of dairy lipids. J. Dairy Sci. 2016, 100, 4197–4212. [Google Scholar] [CrossRef] [PubMed]
- Zervas, G.; Tsiplakou, E. The effect of feeding systems on the characteristics of products from small ruminants. Small Rumin. Res. 2011, 101, 140–149. [Google Scholar] [CrossRef]
- Markiewicz-Kęszycka, M.; Czyżak-Runowska, G.; Lipińska, P.J.W. Fatty Acid Profile of Milk-A Review. Bull. Vet. Inst. Pulawy 2013, 57, 135–139. [Google Scholar] [CrossRef]
- Scharf, R.J.; Demmer, R.T.; DeBoer, M.D. Longitudinal evaluation of milk type consumed and weight status in preschoolers. Arch. Dis. Child. 2013, 98, 335–340. [Google Scholar] [CrossRef]
- Robinson, J.G.; Ijioma, N.; Harris, W. Omega-3 fatty acids and cognitive function in women. Women’s Health 2010, 6, 119–134. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Banu, J.; Rahman, M.; Causey, J.; Fernandes, G. Biological effects of conjugated linoleic acids in health and disease. J. Nutr. Biochem. 2006, 17, 789–810. [Google Scholar] [CrossRef]
- Jenkins, B.; West, J.A.; Koulman, A. A review of odd-chain fatty acid metabolism and the role of pentadecanoic acid (C15:0) and heptadecanoic acid (C17:0) in health and disease. Molecules 2015, 20, 2425–2444. [Google Scholar] [CrossRef]
- Fievez, V.; Colman, E.; Castro-Montoya, J.M.; Stefanov, I.; Vlaeminck, B. Milk odd- and branched-chain fatty acids as biomarkers of rumen function—An update. Anim. Feed Sci. Technol. 2012, 172, 51–65. [Google Scholar] [CrossRef]
- Dewhurst, R.J.; Shingfield, K.J.; Lee, M.R.F.; Scollan, N.D. Increasing the concentrations of beneficial polyunsaturated fatty acids in milk produced by dairy cows in high-forage systems. Anim. Feed Sci. Technol. 2006, 131, 168–206. [Google Scholar] [CrossRef]
- Butler, G.; Stergiadis, S.; Seal, C.; Eyre, M.; Leifert, C. Fat composition of organic and conventional retail milk in northeast England. J. Dairy Sci. 2011, 94, 24–36. [Google Scholar] [CrossRef] [PubMed]
- Stergiadis, S.; Leifert, C.; Seal, C.J.; Eyre, M.D.; Nielsen, J.H.; Larsen, M.K.; Slots, T.; Steinshamn, H.; Butler, G. Effect of feeding intensity and milking system on nutritionally relevant milk components in dairy farming systems in the north east of England. J. Agric. Food Chem. 2012, 60, 7270–7281. [Google Scholar] [CrossRef] [PubMed]
- Sanz Sampelayo, M.R.; Chilliard, Y.; Schmidely, P.; Boza, J. Influence of type of diet on the fat constituents of goat and sheep milk. Small Rumin. Res. 2007, 68, 42–63. [Google Scholar] [CrossRef]
- Biondi, L.; Valvo, M.A.; Di Gloria, M.; Tenghi, E.S.; Galofaro, V.; Priolo, A. Changes in ewe milk fatty acids following turning out to pasture. Small Rumin. Res. 2008, 75, 17–23. [Google Scholar] [CrossRef]
- Martini, M.; Liponi, G.B.; Salari, F. Effect of forage:concentrate ratio on the quality of ewe’s milk, especially on milk fat globules characteristics and fatty acids composition. J. Dairy Res. 2010, 77, 239–244. [Google Scholar] [CrossRef]
- Tsiplakou, E.; Kominakis, A.; Zervas, G. The interaction between breed and diet on CLA and fatty acids content of milk fat of four sheep breeds kept indoors or at grass. Small Rumin. Res. 2008, 74, 179–187. [Google Scholar] [CrossRef]
- Mele, M.; Contarini, G.; Cercaci, L.; Serra, A.; Buccioni, A.; Povolo, M.; Conte, G.; Funaro, A.; Banni, S.; Lercker, G.; et al. Enrichment of Pecorino cheese with conjugated linoleic acid by feeding dairy ewes with extruded linseed: Effect on fatty acid and triglycerides composition and on oxidative stability. Int. Dairy J. 2011, 21, 365–372. [Google Scholar] [CrossRef]
- Gómez-Cortés, P.; Toral, P.G.; Frutos, P.; Juárez, M.; de la Fuente, M.A.; Hervás, G. Effect of the supplementation of dairy sheep diet with incremental amounts of sunflower oil on animal performance and milk fatty acid profile. Food Chem. 2011, 125, 644–651. [Google Scholar] [CrossRef]
- Volanis, M.; Kominakis, A.; Rogdakis, E. Genetic analysis of udder score and milk traits in test day records of Sfakia dairy ewes. Archiv. fur Tierzucht Dummerstorf 2002, 45, 69–77. [Google Scholar] [CrossRef]
- Volanis, M.; Stefanakis, A.; Hadjigeorgiou, I.; Zoiopoulos, P. Supporting the extensive dairy sheep smallholders of the semi-arid region of Crete through technical intervention. Trop. Anim. Health Prod. 2007, 39, 325–334. [Google Scholar] [CrossRef]
- Stefanakis, A.; Volanis, M.; Zoiopoulos, P.; Hadjigeorgiou, I. Assessing the potential benefits of technical intervention in evolving the semi-intensive dairy-sheep farms in Crete. Small Rumin. Res. 2007, 72, 66–72. [Google Scholar] [CrossRef]
- Pinheiro, J.C.; Bates, D.M. Mixed-Effects Models in S and S-PLUS; Springer: New York, NY, USA, 2000. [Google Scholar]
- Crawley, M.J. The R Book; Wiley: Chichester, UK; Hoboken, NJ, USA, 2007. [Google Scholar]
- ter Braak, C.J.F.; Smilauer, P. CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (version 4.5); Biometris: Wageningen, The Netherlands, 2002. [Google Scholar]
- Hibbeln, J.R.; Nieminen, L.R.; Blasbalg, T.L.; Riggs, J.A.; Lands, W.E. Healthy intakes of n-3 and n-6 fatty acids: Estimations considering worldwide diversity. Am. J. Clincal Nutr. 2006, 83, 1483s–1493s. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (EFSA). Panel on Dietetic Products, Nutrition, and Allergies, Scientific Opinion on Dietary Reference Values for fats, including saturated fatty acids, polyunsaturated fatty acids, monounsaturated fatty acids, trans fatty acids, and cholesterol. Eur. Food Saf. Auth. J. 2010, 8, 107. [Google Scholar]
- Parente, G. Grazing systems and biodiversity in Mediterranean areas: Spain, Italy and Greece. Grass Forage Sci. 2011, 66, 606. [Google Scholar] [CrossRef]
- Molle, G.; Decandia, M.; Ligios, S.; Fois, N. Grazing management and stocking rate with particular reference to Mediterranean environments. In Dairy Sheep Nutrition; Pulina, G., Bencini, R., Eds.; CABI Pub: Oxfordshire, UK, 2004; pp. 191–211. [Google Scholar]
- Nudda, A.; Battacone, G.; Boaventura Neto, O.; Cannas, A.; Francesconi, A.H.D.; Atzori, A.S.; Pulina, G. Feeding strategies to design the fatty acid profile of sheep milk and cheese. Rev. Bras. de Zootec. 2014, 43, 445–456. [Google Scholar] [CrossRef]
- Cabiddu, A.; Decandia, M.; Addis, M.; Piredda, G.; Pirisi, A.; Molle, G. Managing Mediterranean pastures in order to enhance the level of beneficial fatty acids in sheep milk. Small Rumin. Res. 2005, 59, 169–180. [Google Scholar] [CrossRef]
- Dervishi, E.; Joy, M.; Sanz, A.; Alvarez-Rodriguez, J.; Molino, F.; Calvo, J.H. Forage preservation (grazing vs. hay) fed to ewes affects the fatty acid profile of milk and CPT1B gene expression in the sheep mammary gland. BMC Vet. Res. 2012, 8, 106. [Google Scholar] [CrossRef]
- Carloni, M.; Fedeli, D.; Roscioni, T.; Gabbianelli, R.; Falcioni, G. Seasonal variation of fat composition in sheep’s milk from areas of central Italy. Mediterr. J. Nutr. Metab. 2009, 3, 55–60. [Google Scholar] [CrossRef]
- Chilliard, Y.; Ferlay, A.; Rouel, J.; Lamberet, G. A review of nutritional and physiological factors affecting goat milk lipid synthesis and lipolysis. J. Dariy Sci. 2003, 86, 1751–1770. [Google Scholar] [CrossRef]
- Virto, M.; Bustamante, M.; Ruiz de Gordoa, J.C.; Amores, G.; Fernandez-Caballero, P.N.; Mandaluniz, N.; Arranz, J.; Najera, A.I.; Albisu, M.; Perez-Elortondo, F.J.; et al. Interannual and geographical reproducibility of the nutritional quality of milk fat from commercial grazing flocks. J. Dairy Res. 2012, 79, 485–494. [Google Scholar] [CrossRef]
- Martin, B.; Buchin, S.; Hurtaud, C. Conditions in milk production and the sensory qualitites of cheese. Prod. Anim. 2003, 16, 283–288. [Google Scholar]



{kind=link}
{kind=link}
| Milk Yield | Fat | Protein | Lactose | Somatic Cell Count | Colony-Forming Units | |
|---|---|---|---|---|---|---|
| Factors | l/ewe/day | g/100 mL | g/100 mL | g/100 mL | 1000 cells/mL | 1000 units/mL |
| Management | ||||||
| Semi-Intensive | 1.00 ± 0.03 | 5.43 ± 0.11 | 5.17 ± 0.03 | 4.78 ± 0.02 | 98 ± 6 | 477 ± 54 |
| Extensive | 0.63 ± 0.02 | 6.05 ± 0.11 | 5.24 ± 0.03 | 4.73 ± 0.03 | 108 ± 8 | 537 ± 60 |
| Sampling Months | ||||||
| January | 1.02 ± 0.06 a | 6.22 ± 0.17 a | 5.17 ± 0.06 | 4.78 ± 0.03 ab | 107 ± 10 abc | 331 ± 74 b |
| February | 0.92 ± 0.05 ab | 6.53 ± 0.16 a | 5.28 ± 0.06 | 4.77 ± 0.04 ab | 126 ± 15 a | 435 ± 93 b |
| March | 0.83 ± 0.04 bc | 5.41 ± 0.16 b | 5.24 ± 0.06 | 4.88 ± 0.03 a | 97 ± 12 abc | 423 ± 90 b |
| April | 0.79 ± 0.04 cd | 5.33 ± 0.21 b | 5.24 ± 0.06 | 4.85 ± 0.03 ab | 91 ± 13 bc | 387 ± 88 b |
| May | 0.71 ± 0.05 de | 5.21 ± 0.20 b | 5.23 ± 0.05 | 4.76 ± 0.05 b | 76 ± 12 c | 786 ± 114 a |
| June | 0.60 ± 0.05 e | 5.68 ± 0.17 b | 5.06 ± 0.05 | 4.47 ± 0.08 c | 121 ± 14 ab | 714 ± 117 a |
| Milking Year | ||||||
| First | 0.90 ± 0.03 | 5.41 ± 0.11 | 5.16 ± 0.03 | 4.85 ± 0.03 | 83 ± 6 | 389 ± 50 |
| Second | 0.74 ± 0.03 | 6.06 ± 0.11 | 5.24 ± 0.03 | 4.66 ± 0.03 | 123 ± 9 | 623 ± 61 |
| ANOVA (p-values) 1 | ||||||
| Main effects | ||||||
| Management (MS) | *** | *** | ns | ns | ns | ns |
| Sample Month (SM) | *** | *** | t | *** | * | *** |
| Year (Y) | *** | *** | t | *** | *** | ** |
| Interactions | ||||||
| MS × SM 2 | ns | ns | ns | *** | ns | ns |
| MS × Y | ns | t | ns | ns | ns | ns |
| SM × Y | ns | ns | ns | ns | ns | ns |
| MS × SM × Y | ns | ns | ns | ns | ns | ns |
| Lauric Acid C12:0 | Myristic Acid C14:0 | Palmitic Acid C16:0 | Stearic Acid C18:0 | Oleic Acid C18:1 Cis9 | Vaccenic Acid C18:1 trans11 | |
|---|---|---|---|---|---|---|
| Factors | g/100 g total FAs | |||||
| Management | ||||||
| Semi-Intensive | 4.92 ± 0.13 | 12.4 ± 0.15 | 27.2 ± 0.20 | 8.72 ± 0.19 | 19.3 ± 0.33 | 1.15 ± 0.05 |
| Extensive | 4.06 ± 0.12 | 11.3 ± 0.15 | 27.6 ± 0.24 | 9.60 ± 0.18 | 21.3 ± 0.35 | 1.01 ± 0.05 |
| Sampling Months | ||||||
| January | 4.93 ± 0.18 ab | 11.6 ± 0.21 ab | 26.9 ± 0.27 b | 9.13 ± 0.30 bc | 19.7 ± 0.51 bc | 1.18 ± 0.08 ab |
| February | 5.14 ± 0.19 a | 12.0 ± 0.24 a | 26.6 ± 0.36 b | 8.83 ± 0.32 bc | 19.5 ± 0.48 bc | 1.19 ± 0.09 a |
| March | 5.06 ± 0.21 ab | 12.3 ± 0.26 a | 27.0 ± 0.30 b | 8.34 ± 0.33 c | 19.1 ± 0.60 c | 1.14 ± 0.07 ab |
| April | 4.56 ± 0.22 b | 12.1 ± 0.30 a | 26.8 ± 0.36 b | 9.19 ± 0.35 bc | 19.0 ± 0.49 c | 1.18 ± 0.09 ab |
| May | 3.97 ± 0.22 c | 12.0 ± 0.31 a | 28.5 ± 0.46 a | 9.36 ± 0.34 ab | 20.6 ± 0.61 a | 1.00 ± 0.07 b |
| June | 3.19 ± 0.17 d | 11.2 ± 0.30 b | 28.7 ± 0.41 a | 10.15 ± 0.29 a | 24.0 ± 0.60 a | 0.76 ± 0.07 c |
| Milking Year | ||||||
| First | 5.18 ± 0.13 | 13.0 ± 0.12 | 28.7 ± 0.20 | 8.38 ± 0.18 | 19.0 ± 0.33 | 0.78 ± 0.04 |
| Second | 3.85 ± 0.11 | 10.8 ± 0.13 | 26.1 ± 0.16 | 9.89 ± 0.18 | 21.5 ± 0.34 | 1.36 ± 0.04 |
| ANOVA (p-values) 1 | ||||||
| Main Effects | ||||||
| Management (MS) | *** | *** | ** | *** | *** | *** |
| Sample Month (SM) | *** | *** | *** | *** | *** | *** |
| Year (Y) | *** | *** | *** | *** | *** | *** |
| Interactions | ||||||
| MS × SM 2 | *** | *** | * | * | ** | ** |
| MS × Y 3 | * | ns | * | ns | * | ns |
| SM × Y 4 | ns | ns | ** | ns | ns | ns |
| MS × SM × Y | * | ns | ns | ns | ** | ns |
| α-Linolenic Acid C18:3 Cis9.Cis12.Cis15 | Conjugated Linoleic Acid C18:2 Cis9.Trans11 | Eicosapentaenoic Acid C20:5 N-3 | Docosapentaenoic Acid C22:5 N-3 | Docosahexaenoic Acid C22:6 N-3 | |
|---|---|---|---|---|---|
| Factors | g/100 g total FA | ||||
| Management | |||||
| Semi-Intensive | 0.55 ± 0.02 | 1.04 ± 0.02 | 0.046 ± 0.003 | 0.075 ± 0.004 | 0.019 ± 0.002 |
| Extensive | 0.75 ± 0.05 | 0.99 ± 0.03 | 0.061 ± 0.003 | 0.092 ± 0.005 | 0.020 ± 0.002 |
| Sampling Months | |||||
| January | 0.47 ± 0.03 d | 0.87 ± 0.04 c | 0.041 ± 0.003 c | 0.076 ± 0.007 bc | 0.019 ± 0.003 |
| February | 0.48 ± 0.05 d | 0.98 ± 0.05 bc | 0.037 ± 0.003 c | 0.074 ± 0.007 c | 0.019 ± 0.003 |
| March | 0.54 ± 0.03 cd | 1.07 ± 0.04 ab | 0.039 ± 0.004 c | 0.068 ± 0.005 c | 0.017 ± 0.003 |
| April | 0.88 ± 0.09 a | 1.13 ± 0.05 a | 0.056 ± 0.005 b | 0.099 ± 0.008 a | 0.020 ± 0.003 |
| May | 0.84 ± 0.08 ab | 1.08 ± 0.05 ab | 0.066 ± 0.006 b | 0.095 ± 0.009 ab | 0.023 ± 0.003 |
| June | 0.68 ± 0.06 bc | 0.96 ± 0.04 bc | 0.084 ± 0.005 a | 0.088 ± 0.008 abc | 0.020 ± 0.003 |
| Milking Year | |||||
| First | 0.53 ± 0.03 | 0.90 ± 0.03 | 0.044 ± 0.003 | 0.049 ± 0.002 | 0.003 ± 0.000 |
| Second | 0.75 ± 0.04 | 1.12 ± 0.03 | 0.062 ± 0.003 | 0.116 ± 0.004 | 0.036 ± 0.001 |
| ANOVA (p-values) 1 | |||||
| Main effects | |||||
| Management (MS) | *** | ns | *** | *** | ns |
| Sample Month (SM) | *** | *** | *** | *** | ns |
| Year (Y) | *** | *** | *** | *** | *** |
| Interactions | |||||
| MS × SM 2 | *** | * | *** | *** | * |
| MS × Y 3 | * | ns | * | ns | ns |
| SM × Y 4 | ns | * | ns | ns | ns |
| MS × SM × Y | ns | ns | ns | ns | ns |
| SFAs | MUFAs | PUFAs | Omega-3 PUFAs | Omega-6 PUFAs | Omega-6/Omega-3 Ratio | |
|---|---|---|---|---|---|---|
| Factors | g/100 g Total FA | |||||
| Management | ||||||
| Semi-Intensive | 69.8 ± 0.40 | 24.5 ± 0.36 | 5.73 ± 0.09 | 0.97 ± 0.04 | 3.06 ± 0.07 | 3.9 ± 0.20 |
| Extensive | 67.6 ± 0.49 | 26.3 ± 0.36 | 5.88 ± 0.14 | 1.18 ± 0.07 | 3.03 ± 0.05 | 3.7 ± 0.25 |
| Sampling Months | ||||||
| January | 69.8 ± 0.61 a | 24.8 ± 0.54 a | 5.40 ± 0.15 c | 0.85 ± 0.06 d | 3.06 ± 0.10 | 4.6 ± 0.39 ab |
| February | 69.4 ± 0.63 a | 24.9 ± 0.53 a | 5.72 ± 0.19 bc | 0.86 ± 0.08 cd | 3.21 ± 0.12 | 4.9 ± 0.56 a |
| March | 70.1 ± 0.75 a | 24.2 ± 0.65 a | 5.64 ± 0.17 bc | 0.94 ± 0.06 cd | 2.95 ± 0.09 | 3.7 ± 0.28 bc |
| April | 68.8 ± 0.96 a | 24.2 ± 0.52 a | 6.28 ± 0.26 a | 1.40 ± 0.12 a | 2.94 ± 0.11 | 2.7 ± 0.25 c |
| May | 68.2 ± 0.79 a | 25.7 ± 0.64 a | 6.08 ± 0.22 ab | 1.31 ± 0.11 ab | 3.00 ± 0.10 | 3.0 ± 0.29 c |
| June | 65.6 ± 0.81 b | 28.7 ± 0.70 b | 5.74 ± 0.18 abc | 1.08 ± 0.09 bc | 3.10 ± 0.09 | 3.8 ± 0.40 bc |
| Milking Year | ||||||
| First | 71.6 ± 0.31 | 23.3 ± 0.28 | 5.07 ± 0.09 | 0.78 ± 0.04 | 2.93 ± 0.07 | 4.9 ± 0.28 |
| Second | 65.9 ± 0.42 | 27.4 ± 0.35 | 6.51 ± 0.10 | 1.35 ± 0.05 | 3.15 ± 0.05 | 2.7 ± 0.10 |
| ANOVA (p-values) 1 | ||||||
| Main effects | ||||||
| Management(MS) | *** | *** | ns | *** | ns | ns |
| Sample Month (SM) | *** | *** | *** | *** | ns | *** |
| Year (Y) | *** | *** | *** | *** | ** | *** |
| Interactions | ||||||
| MS × SM 2 | *** | *** | *** | *** | ns | *** |
| MS × Y 3 | ns | ns | * | * | ns | ns |
| SM × Y 4 | ns | ns | ns | ns | ns | ns |
| MS × SM × Y | ns | ns | ns | ns | ns | ns |
| Factor | Years | Stage of Lactation | Management System |
|---|---|---|---|
| Liquid yield | + | +++ | ++ |
| Solids yield | 0 | +++ | ++ |
| SFAs | 0 | 0 | 0 |
| MUFAs | + | + | 0 |
| PUFAs | + | + | 0 |
| Omega-3 | +++ | +++ | + |
| LC-omega-3 | +++ | ++ | + |
| Omega-6 | 0 | 0 | 0 |
| Ratio of n-6/n-3 | +++ | +++ | 0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Voutzourakis, N.; Tzanidakis, N.; Stergiadis, S.; Rempelos, L.; Eyre, M.; Atsali, I.; Franceschin, E.; Leifert, C.; Stefanakis, A.; Sotiraki, S.; et al. Sustainable Intensification? Increased Production Diminishes Omega-3 Content of Sheep Milk. Sustainability 2020, 12, 1228. https://doi.org/10.3390/su12031228
Voutzourakis N, Tzanidakis N, Stergiadis S, Rempelos L, Eyre M, Atsali I, Franceschin E, Leifert C, Stefanakis A, Sotiraki S, et al. Sustainable Intensification? Increased Production Diminishes Omega-3 Content of Sheep Milk. Sustainability. 2020; 12(3):1228. https://doi.org/10.3390/su12031228
Chicago/Turabian StyleVoutzourakis, Nikolaos, Nikolaos Tzanidakis, Sokratis Stergiadis, Leonidas Rempelos, Mick Eyre, Ioanna Atsali, Enrica Franceschin, Carlo Leifert, Alexandros Stefanakis, Smaragda Sotiraki, and et al. 2020. "Sustainable Intensification? Increased Production Diminishes Omega-3 Content of Sheep Milk" Sustainability 12, no. 3: 1228. https://doi.org/10.3390/su12031228
APA StyleVoutzourakis, N., Tzanidakis, N., Stergiadis, S., Rempelos, L., Eyre, M., Atsali, I., Franceschin, E., Leifert, C., Stefanakis, A., Sotiraki, S., & Butler, G. (2020). Sustainable Intensification? Increased Production Diminishes Omega-3 Content of Sheep Milk. Sustainability, 12(3), 1228. https://doi.org/10.3390/su12031228