Defining Dry Rivers as the Most Extreme Type of Non-Perennial Fluvial Ecosystems


 and
and
Abstract
1. Introduction
2. Contextualization of Dry Rivers
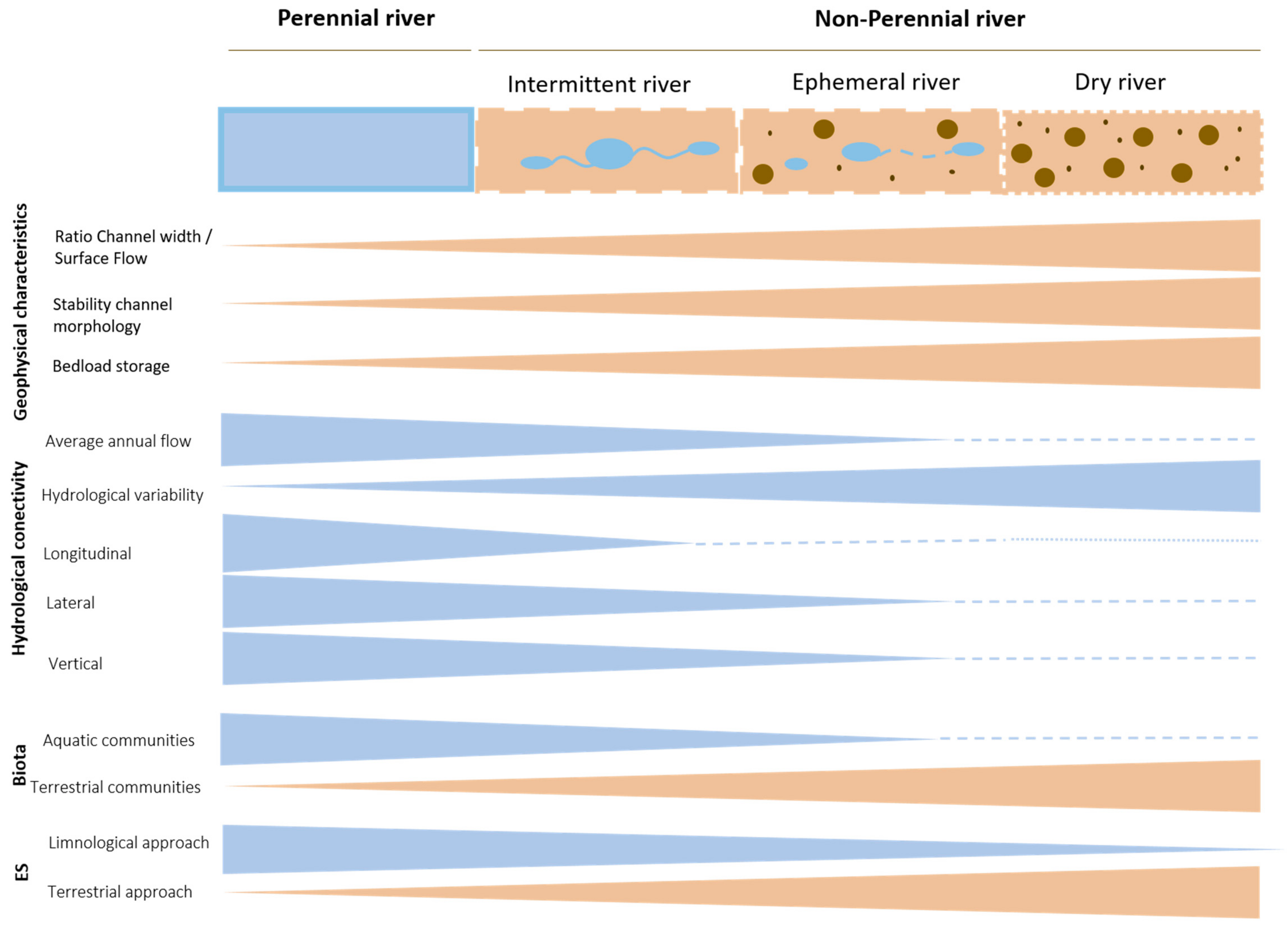
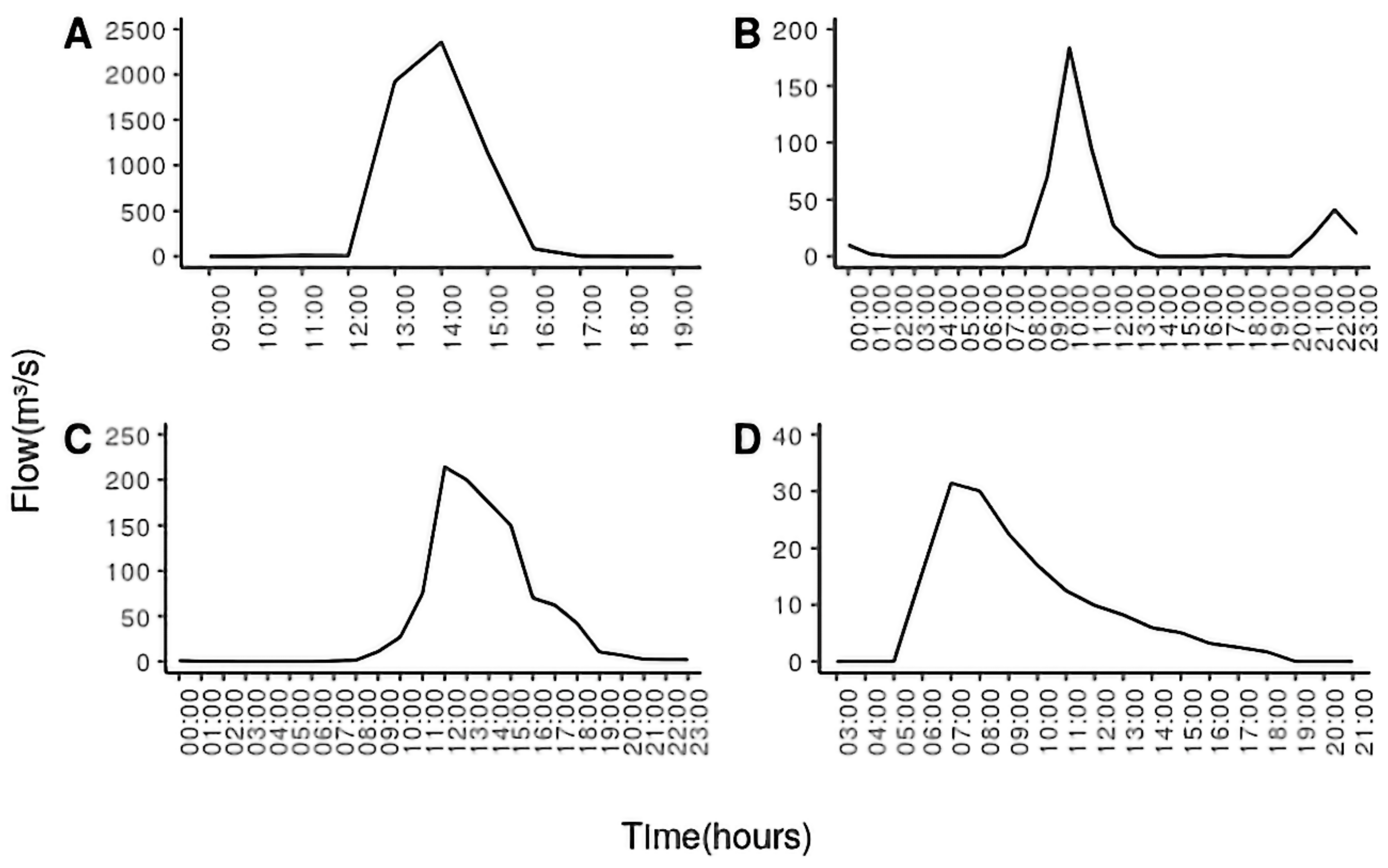
3. Distribution, Geophysical and Hydrological Characteristics of Dry Rivers
4. Biota of Dry Rivers
5. Biogeochemical Processing in Dry Rivers
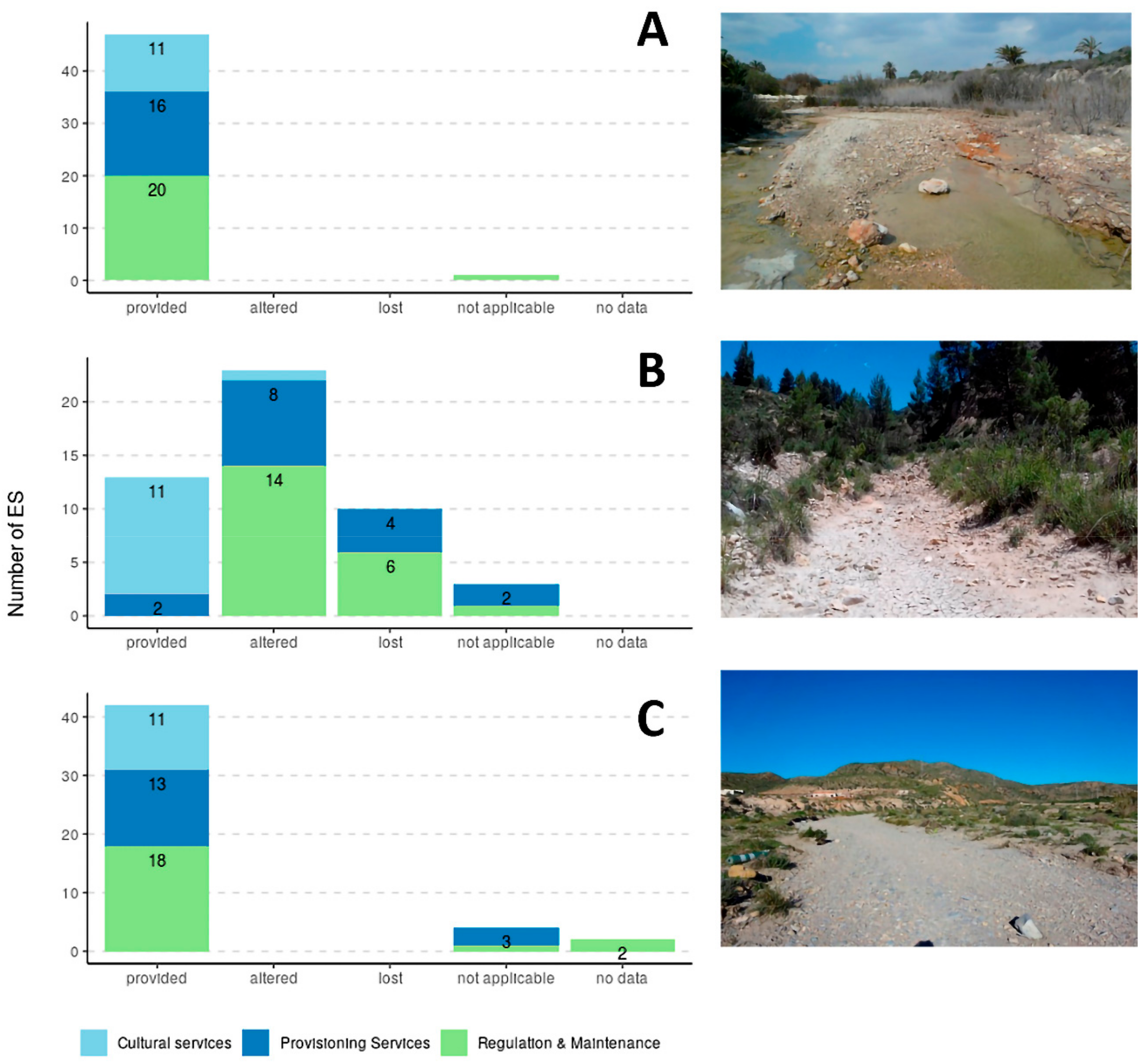
6. Ecosystem Services of Dry Rivers
7. Anthropogenic Threats to Dry Rivers
8. Management Challenges
9. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Leopold, L.B.; Miller, J.P. Ephemeral Streams—Hydraulic Factors and Their Relation to the Drainage Net; United States Geological Survey Professional Paper 282; U.S Government Printing Office: Washington, DC, USA, 1956. [Google Scholar]
- Rodier, J.A. Aspects of arid zones hydrology. In Facets of Hydrology; Rodda, J.C., Ed.; John Wiley & Sons: Chichester, UK, 1985; Volume II, pp. 205–247. [Google Scholar]
- Boulton, A.J.; Suter, P. Ecology of temporary streams—An Australian perspective. In Limnology in Australia; De Deckker, P., Williams, W.D., Eds.; CSIRO 61: Melbourne, Australia, 1986; pp. 313–327. [Google Scholar] [CrossRef]
- Graf, W.J. Fluvial Processes in Dryland Rivers; Springer: Berlin/Heidelberg, Germany; Verlag: Zurich, Switzerland, 1988. [Google Scholar]
- Grimm, N.B.; Valett, H.M.; Stanley, E.H.; Fisher, S.G. Contribution of the hyporheic zone to stability of an arid-land stream. Verh. Intern. Ver. Limnol. 1991, 24, 1595–1599. [Google Scholar] [CrossRef]
- Stanley, E.H.; Buschman, D.L.; Boulton, A.J.; Grimm, N.B.; Fisher, S.G. Invertebrate resistance and resilience to intermittency in a desert stream. Am. Midl. Nat. 1994, 131, 288–300. [Google Scholar] [CrossRef]
- Thornes, J.B. Channel processes, evolution and history. In Geomorphology of Desert Environments; Abrahams, A.D., Parsons, A.J., Eds.; Chapman & Hall: London, UK, 1994; pp. 288–317. [Google Scholar]
- Jacobson, P.J.; Jacobson, K.M.; Seely, M.K. Ephemeral Rivers and Their Catchments: Sustaining People and Development in Western Namibia; Desert Research Foundation of Namibia: Windhoek, Namibia, 1995. [Google Scholar]
- Jacobson, P.J.; Jacobson, K.M.; Angermeier, P.L.; Cherry, D.S. Transport, retention and ecological significance of woody debris within a large ephemeral river. J. N Am. Benthol. Soc. 1999, 18, 429–444. [Google Scholar] [CrossRef]
- Giller, P.S. Floods and droughts: The effects of variations in water flow on streams and rivers. In Disturbance and Recovery of Ecological Systems: Proceedings of a Seminar Held on 14–15 February 1995; Myers, A.A., Giller, P.S., Eds.; Royal Irish Academy: Dublin, Ireland, 1996; pp. 1–19. [Google Scholar]
- Uys, M.C.; O’Keeffe, J.H. Simple words and fuzzy zones: Early directions for temporary river research in South Africa. Environ. Manag 1997, 21, 517–531. [Google Scholar] [CrossRef]
- Reid, I.; Frostick, L.E. Channel form flows and sediments in deserts. In Arid Zone Geomorphology: Process, Form and Change in Drylands, 2nd ed.; John Wiley & Sons: Chichester, UK, 1997; pp. 205–219. [Google Scholar]
- Costigan, K.H.; Jaeger, K.L.; Goss, C.W.; Fritz, K.M.; Goebel, P.C. Understanding control of flow permanence in intermittent rivers to aid ecological research: Integrating meteorology, geology and land cover. Ecohydrology 2016, 9, 1141–1153. [Google Scholar] [CrossRef]
- Datry, T.; Bonada, N.; Boulton, A. Intermittent rivers and ephemeral streams. Ecology and Management; Academic Press: London, UK, 2017. [Google Scholar]
- Raymond, P.A.; Hartmann, J.; Lauerwald, R.; Sobek, S.; McDonald, C.; Hoover, M.; Butman, D.; Striegl, R.; Mayorga, E.; Humborg, C.; et al. Global carbon dioxide emissions from inland waters. Nature 2013, 503, 355–359. [Google Scholar] [CrossRef]
- Larned, S.T.; Datry, T.; Arscott, D.B.; Tockner, K. Emerging concepts in temporary-river ecology. Freshw. Biol. 2010, 55, 717–738. [Google Scholar] [CrossRef]
- Steward, A.L.; von Schiller, D.; Tockner, K.; Marshall, J.C.; Bunn, S.E. When the river runs dry: Human and ecological values of dry riverbeds. Front. Ecol Environ. 2012, 10, 202–209. [Google Scholar] [CrossRef]
- Acuña, V.; Datry, T.; Marshall, J.; Barceló, D.; Dahm, C.N.; Ginebreda, A.; McGregor, G.; Sabater, S.; Tockner, K.; Palmer, M.A. Why should we care about temporary waterways? Science 2014, 343, 1080–1081. [Google Scholar] [CrossRef]
- Vidal-Abarca, M.R. Los ríos de las cuencas áridas y semiáridas: Una perspectiva comparativa y de síntesis. Sci. Gerund. 1990, 16, 219–228. [Google Scholar]
- Dent, C.L.; Grimm, N.B. Spatial heterogeneity of stream water nutrient concentrations over successional time. Ecology 1999, 80, 2283–2298. [Google Scholar] [CrossRef]
- Lake, P.S. Ecological effects of perturbation by drought in flowing waters. Freshw. Biol. 2003, 48, 1161–1172. [Google Scholar] [CrossRef]
- Vidal-Abarca, M.R.; Gómez, R.; Suárez, M.L. Los ríos de las regiones semiáridas. Ecosistemas 2004, 13. Available online: https://www.redalyc.org/articulo.oa?id=54013113 (accessed on 27 March 2020).
- Bonada, N.; Rieradevall, M.; Prat, N. Macroinvertebrate community structure and biological traits related to flow permanence in a Mediterranean river network. Hydrobiologia 2007, 589, 91–106. [Google Scholar] [CrossRef]
- Sánchez-Montoya, M.M.; Puntí, T.; Suárez, M.L.; Vidal-Abarca, M.R.; Rieradevall, M.; Poquet, J.M.; Zamora, C.; Robles, S.; Álvarez, M.; Alba, J.; et al. Concordance between ecotypes and macroinvertebrate assemblages in Mediterranean streams. Freshw. Biol. 2007, 52, 2240–2255. [Google Scholar] [CrossRef]
- Leigh, C.; Datry, T. Drying as a primary hydrological determinant of biodiversity in river systems: A broad-scale analisis. Ecography 2016, 39, 001–013. [Google Scholar] [CrossRef]
- Soria, M.; Leigh, C.; Datry, T.; Bini, L.M.; Bonada, N. Biodiversity in perennial and intermittent rivers: A meta-analysis. Oikos 2017, 126, 1078–1089. [Google Scholar] [CrossRef]
- Bunn, S.E.; Balcombe, S.R.; Davies, P.M.; Fellows, C.S.; Mc Kenzie-Smith, F.J. Aquatic productivity and food webs of desert river ecosystems. In Ecology of Desert Rivers; Kingsford, R.T., Ed.; Cambridge 490; Cambridge University Press: Cambridge, UK, 2006; pp. 76–99. [Google Scholar]
- Bernal, S.; von Schiller, D.; Sabater, F.; Martí, E. Hydrological extremes modulate nutrient dynamics in mediterranean climate streams across different spatial scales. Hydrobiologia 2013, 719, 21–42. [Google Scholar] [CrossRef]
- Arce, M.I.; Mendoza-Lera, C.; Almagro, M.; Catalan, N.; Romani, A.M.; Marti, E.; Gomez, R.; Bernal, S.; Foulquier, A.; Mutz, M.; et al. A conceptual framework for understanding the biogeochemistry of dry riverbeds through the lens of soil science. Earth-Sci. Rev. 2019, 188, 441–453. [Google Scholar] [CrossRef]
- Von Schiller, D.; Datry, T.; Corti, R.; Foulquier, A.; Tockner, K.; Marcé, R.; García-Baquero, G.; Odriozola, I.; Obrador, B.; Elosegi, A.; et al. Sediment respiration pulses in intermittent rivers and ephemeral streams. Glob. Biogeochem. Cycles 2019, 33, 1251–1263. [Google Scholar] [CrossRef]
- Marcé, R.; Obrador, B.; Gómez-Gener, L.; Catalán, N.; Koschorreck, M.; Arce, M.I.; Singere, G.; von Schiller, D. Emissions from dry inland waters are a blind spot in the global carbon cycle. Earth-Sci. Rev. 2019, 188, 240–248. [Google Scholar] [CrossRef]
- Datry, T.; Larned, S.T.; Tockner, K. Intermittent rivers: A challenge for freshwater ecology. Bioscience 2014, 64, 229–235. [Google Scholar] [CrossRef]
- Sánchez-Montoya, M.M.; von Schiller, D.; Barbera, G.G.; Diaz, A.M.; Arce, M.I.; del Campo, R.; Tockner, K. Understanding the effects of predictability, duration, and spatial pattern of drying on benthic invertebrate assemblages in two contrasting intermittent streams. PLoS ONE 2018, 13, e0193933. [Google Scholar] [CrossRef] [PubMed]
- Gordon, N.D.; McMahon, T.A.; Finlayson, B.L.; Gippel, C.J.; Nathan, R.J. Stream Hydrology: An Introduction for Ecologists, 2nd ed.; Wiley: Chichester, UK, 2004. [Google Scholar]
- Stanley, E.H.; Fisher, S.G.; Grimm, N.B. Ecosystem expansion and contraction in streams. Bioscience 1997, 47, 427–435. [Google Scholar] [CrossRef]
- Datry, T. Benthic and hyporheic invertebrate assemblages along a flow intermittence gradient: Effects of duration of dry events. Freshw. Biol. 2012, 57, 563–574. [Google Scholar] [CrossRef]
- Gómez, R.; Arce, M.I.; Sánchez, J.J.; Sánchez-Montoya, M.M. The effects of drying on sediment nitrogen content in a Mediterranean intermittent stream: A microcosms study. Hydrobiologia 2012, 679, 43–59. [Google Scholar] [CrossRef]
- Bogan, M.T.; Boersma, K.S.; Lytle, D.A. Resistance and resilience of invertebrate communities to seasonal and supraseasonal drought in arid-land headwater streams. Freshw. Biol. 2015, 60, 2547–2558. [Google Scholar] [CrossRef]
- Arce, M.I.; von Schiller, D.; Bengtsson, M.M.; Hinze, C.; Jung, H.; Eloy Alves, R.J.; Urich, T.; Singer, G. Drying and rainfall shape the structure and functioning of nitrifying microbial communities in riverbed sediments. Front. Microbiol. 2018, 9, 2794. [Google Scholar] [CrossRef]
- Colls, M.; Timoner, X.; Font, C.; Sabater, S.; Acuña, V. Effects of duration, frequency, and severity of the non-flow period on stream biofilm metabolism. Ecosystems 2019, 22, 1393–1405. [Google Scholar] [CrossRef]
- Mabbutt, J.A. Desert Landforms; MIT Press: Cambridge, UK, 1977. [Google Scholar]
- Le Houérou, H.N. North Africa. In Aridland Ecosystems: Structure, Functioning and Management; Goodalld, W., Perryr, A., Eds.; Cambridge University Press: Cambridge, UK, 1979. [Google Scholar]
- Hedman, E.R.; Osterkamp, W.R. Streamflow Characteristics Related to Channel Geometry of Streams in Western United States; U.S. Geological Survey Water-Supply Paper; USGPO: Washington, DC, USA, 1982; Volume 2193, pp. 1–17. [Google Scholar]
- Williams, D.D. The Ecology of Temporary Waters; Croom Helm: New South Wales, Australia, 1987. [Google Scholar]
- Boulton, A.J.; Lake, P.S. Australian temporary streams—Some ecological characteristics. Verh Intern. Ver. Theor. Angew. Limnol. 1988, 23, 1380–1383. [Google Scholar] [CrossRef]
- Matthews, W.J. North American prairie streams as systems for ecological study. J. N. Am. Benthol. Soc. 1988, 7, 387–409. [Google Scholar] [CrossRef]
- Comin, F.A.; Williams, W.D. Parched continents: Our common future? In Limnology Now: A Paradigm of Planetary Problems; Margalef, R., Ed.; Elsevier: Amsterdam, The Netherlands, 1994; pp. 473–527. [Google Scholar]
- Bull, L.J.; Kirkby, M.J. Dryland river characteristics and concept. In Dryland Rivers: Hydrology and Geomorphology of the Semi-Arid Channels; Bull, L.J., Kirkby, M.J., Eds.; John Wiley & Sons: Chichester, UK, 2002; pp. 3–15. [Google Scholar]
- Gómez, R.; Hurtado, I.; Suárez, M.L.; Vidal-Abarca, M.R. Ramblas in south-east Spain: Threatened and valuable ecosystems. Aquat. Conserv. 2005, 15, 387–402. [Google Scholar] [CrossRef]
- Fritz, K.M.; Johnson, B.R.; Walters, D.M. Field Operations Manual for Assessing the Hydrologic Permanence and Ecological Condition of Headwater Streams; EPA/600/R-06/126; U.S. Environmental Protection Agency, Office of Research and Development: Washington, DC, USA, 2006. [Google Scholar] [CrossRef]
- Levick, L.R.; Goodrich, D.C.; Hernandez, M.; Fonseca, J.; Semmens, D.J.; Stromberg, J.C.; Leidy, R.; Scianni, M.; Guertin, D.P.; Tluczek, M.; et al. The Ecological and Hydrological Significance of Ephemeral and Intermittent Streams in the Arid and Semi-Arid American Southwest; US Environmental Protection Agency, Office of Research and Development: Washington, DC, USA, 2008. [Google Scholar]
- Brooks, R.T. Potential impacts of global climate change on the hydrology and ecology of ephemeral freshwater systems of the forests of the northeastern United States. Clim. Chang. 2009, 95, 469–483. [Google Scholar] [CrossRef]
- Dodds, W.K.; Whiles, M.T. Freshwater Ecology. Concepts and Environmental Applications of Limnology, 2nd ed.; Academic Press: : Cambridge, MA, USA; Elsevier Inc.: Amsterdam, The Netherlands, 2010. [Google Scholar]
- Lake, P.S. Drought and aquatic ecosystems. Effects and Responses; Wiley-Blackwell: West Sussex, UK, 2011. [Google Scholar]
- McDonough, O.T.; Hosen, J.D.; Palmer, M.A. Temporary streams: The Hydrology, Geography, and Ecology of non-perennially flowing waters. In Rivers Ecosystems: Dynamics, Management and Conservation; Elliot, H.S., Martin, L.E., Eds.; Nova Science Publishers, Inc.: New York, NY, USA, 2011; pp. 259–289. [Google Scholar]
- Armstrong, A.; Stedman, R.C.; Bishop, J.A.; Sullivan, P.J. What’s a Stream without Water? Disproportionality in Headwater Regions Impacting Water Quality. Environ. Manag. 2012, 50, 849–860. [Google Scholar] [CrossRef] [PubMed]
- Gallart, F.; Prat, N.; García-Roger, E.M.; Latron, J.; Rieradevall, M.; Llorens, P.; Barbera, G.G.; Brito, D.; De Girolamo, A.M.; Lo Porto, A.; et al. A novel approach to analysing the regimes of temporary streams in relation to their controls on the composition and structure of aquatic biota. Hydrol. Earth Syst. Sci. 2012, 16, 3165–3182. [Google Scholar] [CrossRef]
- Martin, J.; Kurc, S.A.; Zaimes, G.; Crimmins, M.; Hutmacher, A.; Green, D. Elevated air temperatures in riparian ecosystems along ephemeral streams: The role of housing density. J. Arid Environ. 2012, 84, 9–18. [Google Scholar] [CrossRef]
- Arthington, A.H.; Bernardo, J.M.; Ilhéu, M. Temporary rivers: Linking Ecohydrology, Ecological Quality and Reconciliation Ecology. River Res. Applic. 2014, 30, 1209–1215. [Google Scholar] [CrossRef]
- Boulton, A.J. Conservation of ephemeral streams and their ecosystem services: What are we missing? Aquat. Conserv. 2014, 24, 733–738. [Google Scholar] [CrossRef]
- De Girolamo, A.M.; Gallart, F.; Pappagallo, G.; Santese, G.; Lo Porto, A. An eco-hydrological assessment method for temporary rivers. The Celone and Salsola rivers case study (SE, Italy). Ann. Limnol. Int. J. Lim. 2015, 51, 1–10. [Google Scholar] [CrossRef]
- Oueslati, O.; De Girolamo, A.M.; Abouabdillah, A.; Kjeldsen, T.R.; Lo Porto, A. Classifying the flow regimes of Mediterranean streams using multivariate analysis. Hydrol. Process. 2015, 29, 4666–4682. [Google Scholar] [CrossRef]
- Moosavi, S. Where the Rivers Run Dry: An Inquiry into the Design of Wadis in the Arabian Peninsula. Ph.D. Thesis, University of Melbourne, Melbourne, Australia, 2016. [Google Scholar]
- Datry, T.; Boulton, A.J.; Bonada, N.; Fritz, K.; Leigh, C.; Sauquet, E.; Tockner, K.; Hugueny, B.; Dahm, C.N. Flow intermittence and ecosystem services in rivers of the Anthropocene. J. Appl. Ecol. 2017, 1–12. [Google Scholar] [CrossRef] [PubMed]
- DeLucio, J.V.; Múgica, M. Landscape preferences and behaviour of visitors to Spanish national parks. Landsc. Urban. Plan. 1994, 29, 145–160. [Google Scholar] [CrossRef]
- García-Llorente, M.; Martín-López, B.; Iniesta-Arandia, I.; López-Santiago, C.A.; Aguilera, P.A.; Montes, C. The role of multi-functionality in social preferences toward semi-arid rural landscapes: An ecosystem service approach. Environ. Sci. Policy 2012, 19–20, 136–146. [Google Scholar] [CrossRef]
- Schneider, A.; Jost, A.; Coulon, C.; Silvestre, M.; Thery, S.; Ducharne, A.C.G. Global-scale river network extraction based on high-resolution topography and constrained by lithology, climate, slope, and observed drainage density. Geophys Res. Lett. 2017, 44, 2773–2781. [Google Scholar] [CrossRef]
- Gautier, E.F. Sahara: The Great Desert; Octagon Books: New York, NY, USA, 1970. [Google Scholar]
- Davies, B.R.; O’Keeffe, J.H.; Snaddon, C.D. A Synthesis of the Ecological Functioning, Conservation and Management of South African River Ecosystems; Water Research Commission Report No. TT62/93; Water Research Commission: Pretoria, South Africa, 1993. [Google Scholar]
- Sheldon, F.; Bunn, S.E.; Hughes, J.M.; Arthington, A.H.; Balcombe, S.R.; Fellows, C.S. Ecological roles and threats to aquatic refugia in arid landscapes: Dryland river waterholes. Mar. Freshw. Res. 2010, 61, 885–895. [Google Scholar] [CrossRef]
- Snelder, T.H.; Datry, T.; Lamouroux, N.; Larned, S.T.; Sauquet, E.; Pella, H.; Catalogne, C. Regionalization of patterns of flow intermittence from gauging station records. Hydrol. Earth Syst. Sci. 2013, 17, 2685–2699. [Google Scholar] [CrossRef]
- Nanson, G.C.; Tooth, S. Arid-zones rivers as indicators of climate change. In Paleoenvironmental Reconstruction in Arid Lands; Singhvi, A.K., Derbyshire, E., Eds.; A.A. Balkema: Rotterdam, The Netherlands, 1999; pp. 175–216. [Google Scholar]
- Mateu, J.F. Ríos y ramblas mediterráneos. In Avenidas Fluviales e Inundaciones en la Cuenca del Mediterráneo; Gil Olcina, A., Morales Gil, A., Eds.; Instituto Universitario de Geografía, Universidad de Alicante, Caja de Ahorros del Mediterráneo: Murcia, Spain, 1989; pp. 113–150. [Google Scholar]
- Bremner, J.M.; Rogers, J.; Willis, J.P. Sedimentological aspects of the 1988 Orange River floods. Trans. R. Soc. S. Afr. 1990, 47, 247–294. [Google Scholar] [CrossRef]
- Reid, I. Sediment dynamics of ephemeral channels. In Dryland Rivers. Hydrology and Geomorphology of Semi-Arid Channels; Bull, L.J., Kirkby, M.J., Eds.; John Wiley & Sons: Chischester, UK, 2002; pp. 107–128. [Google Scholar]
- García, C.; Laronne, J.B.; Sala, M. Variable source areas of bedload flux in a gravel-bed stream. J. Sediment. Res. 1999, 69, 27–31. [Google Scholar] [CrossRef]
- Griffin, G.F.; Smith, D.M.S.; Morton, S.R.; Allan, G.E.; Masters, K.A. Status and implications of the invasion of Tamarisk (Tamarix aphylla) on the Finke River, Northern Territory, Australia. J. Environ. Manag. 1989, 29, 297–315. [Google Scholar]
- Tooth, S.; Nanson, G.C. The role of vegetation in the formation of anabranching channels in an ephemeral river, Northern Plains, arid central Australia. Hydrol Proc. 2000, 14, 3099–3117. [Google Scholar] [CrossRef]
- Jaeger, K.L.; Sutfin, N.A.; Tooth, S.; Michaelides, K.; Singer, M. Geomorphology and sediment regimes of intermittent rivers and ephemeral streams. In Intermittent Rivers and Ephemeral Streams, Ecology and Management; Datry, T., Bonada, N., Boulton, A., Eds.; Academic Press: London, UK, 2017; pp. 21–49. [Google Scholar]
- Wolman, M.G.; Gerson, R. Relative scale of time and effectiveness of climate in watershed geomorphology. Earth Surf. Proc. Land 1978, 3, 189–208. [Google Scholar] [CrossRef]
- Schumm, S.A. The Fluvial System; John Wiley & Sons: New York, NY, USA, 1977. [Google Scholar]
- Gordon, N.D.; McMahon, T.A.; Finlayson, B.L. Stream Hydrology: An Introduction for Ecologists; John Wiley & Sons: Chichester, UK, 1993. [Google Scholar]
- Boulton, A.J.; Rolls, R.J.; Jaeger, K.L.; Datry, T. Hydrological connectivity in intermittent rivers and ephemeral streams. In Intermittent Rivers and Ephemeral Streams. Ecology and Management; Datry, T., Bonada, N., Boulton, A., Eds.; Academic Press: London, UK, 2017; pp. 79–108. [Google Scholar]
- Nanson, G.C.; Tooth, S.; Knighton, A.D. A global perspective on dryland rivers: Perceptions, misconceptions and distinctions. In Dryland Rivers. Hydrology and Geomorphology of Semi-Arid Channels; John Wiley & Sons: Chischester, UK, 2002; pp. 17–54. [Google Scholar]
- Shannon, J.; Richardson, R.; Thornes, J. Modelling event-based fluxes in ephemeral streams. In Dryland Rivers. Hydrology and Geomorphology of Semi-Arid Channels; Bull, L.J., Kirkby, M.J., Eds.; John Wiley & Sons: Chischester, UK, 2002; pp. 129–172. [Google Scholar]
- Knighton, A.D.; Nanson, G.C. Distinctiveness, diversity and uniqueness in arid zone river systems. In Arid Zone Geomorphology: Process Form and Change in Drylands; Thomas, D.S.G., Ed.; John Wiley & Sons: Chichester, UK, 1997; pp. 185–203. [Google Scholar]
- Larkin, Z.T.; Ralph, T.J.; Tooth, S.; Fryirs, K.A.; Carthey, A.J.R. Identifying threshold responses of Australian dryland rivers to future hydroclimatic change. Sci. Rep. 2020, 10, 6653. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, K.M.; Boulton, A.J. Detecting impacts and setting restoration targets in arid-zone rivers: Aquatic micro-invertebrate responses to reduced floodplain inundation. J. Appl. Ecol. 2007, 44, 833–842. [Google Scholar] [CrossRef]
- Robson, B.J.; Chester, E.T.; Austin, C.M. Why life history information matters: Drought refuges and macroinvertebrate persistence in non-perennial streams subject to a drier climate. Mar. Freshw. Res. 2011, 62, 801–810. [Google Scholar] [CrossRef]
- Stubbington, R.; Datry, T. The macroinvertebrate seedbank promotes community persistence in temporary rivers across climate zones. Freshw. Biol. 2013, 58, 1202–1220. [Google Scholar] [CrossRef]
- Pennak, R.W. Freshwater Invertebrates of the United States, 3rd ed.; John Wiley and Sons: New York, NY, USA, 1989. [Google Scholar]
- Carvalho, G.H.; Wolf, H.G. Resting eggs of lake Daphnia I. Distribution, abundance and hatching of eggs collected from various depths in lake sediments. Freshw. Biol. 1989, 22, 459–470. [Google Scholar] [CrossRef]
- Moritz, C. A note on the hatching and viability of Ceriodaphnia ephippia collected from lake sediment. Hydrobiologia 1987, 145, 309–314. [Google Scholar] [CrossRef]
- De Stasio, B.T.J. The seed bank of a freshwater crustacean: Copepodology for the plant ecologist. Ecology 1989, 70, 1377–1389. [Google Scholar] [CrossRef]
- Sánchez-Montoya, M.M.; Tockner, K.; von Schiller, D.; Miñano, J.; Catarineu, C.; Lencian, J.L.; Barbera, G.G.; Ruhí, A. Dynamics of ground-dwelling arthropod metacommunities in intermittent streams: The key role of dry riverbeds. Biol. Conserv. 2019. [Google Scholar] [CrossRef]
- Wishart, M.J. The terrestrial invertebrate fauna of a temporary stream in southern Africa. Afr. Zool. 2000, 35, 193–200. [Google Scholar] [CrossRef]
- Steward, A.L.; Marshall, J.C.; Sheldon, F.; Harch, B.; Choy, S.; Bunn, S.E.; Tockner, K. Terrestrial invertebrates of dry river beds are not simply subsets of riparian assemblages. Aquat. Sci. 2011, 73, 551–566. [Google Scholar] [CrossRef]
- Steward, A.L.; Negus, P.; Marshall, J.C.; Clifford, S.E.; Dent, C. Assessing the ecological health of rivers when they are dry. Ecol. Indic. 2018, 85, 537–547. [Google Scholar] [CrossRef]
- Corti, R.; Datry, T. Drying of a temperate, intermittent river has little effect on adjacent riparian arthropod communities. Freshw. Biol. 2014, 59, 666–678. [Google Scholar] [CrossRef]
- Sánchez-Montoya, M.M.; von Schiller, D.; Ruhí, S.P.G.; Proia, L.; Miñano, J.; Vidal-Abarca, M.R.; Suárez, M.L.; Tockner, K. Responses of ground-dwelling arthropods to surface flow drying in channels and adjacent habitats along Mediterranean streams. Ecohydrology 2016, 9, 1376–1387. [Google Scholar] [CrossRef]
- Sánchez-Montoya, M.M.; Guerrero-Brotons, M.; Miñano, J.; Gómez, R. Effects of debris piles and pools along dry riverbeds on nutrients, microbial activity, and ground-dwelling arthropods: A Namibian ephemeral river case. J. Arid Environ. 2019, 175. [Google Scholar] [CrossRef]
- Steward, A.L.; Langhans, S.D.; Corti, R.; Datry, T. The biota of intermittent rivers and ephemeral streams: Terrestrial and semiaquatic invertebrates. In Intermittent Rivers and Ephemeral Streams, Ecology and Management; Datry, T., Bonada, N., Boulton, A., Eds.; Academic Press: London, UK, 2017; pp. 245–271. [Google Scholar]
- Segev, O.; Ziv, M.; Lubin, Y. The male mating system in a desert widow spider. J. Arachnol. 2003, 31, 379–393. [Google Scholar] [CrossRef]
- Crist, T.O.; Guertin, D.S.; Wiens, J.A.; Milne, B.T. Animal movement in heterogeneous landscapes: An experiment with Eleodes beetles in short grass prairie. Funct. Ecol. 1992, 6, 536–544. [Google Scholar] [CrossRef]
- Jopp, F.; Reuter, H. Dispersal of carabid beetles: Emergence of distribution patterns. Ecol. Model. 2005, 186, 389–405. [Google Scholar] [CrossRef]
- Sánchez-Montoya, M.M.; Moleón, M.; Sánchez-Zapata, J.A.; Escoriza, D. The biota of intermittent and ephemeral rivers: Amphibians, reptiles, birds and mammals. In Intermittent Rivers and Ephemeral Streams, Ecology and Management; Datry, T., Bonada, N., Boulton, A., Eds.; Academic Press: London, UK, 2017; pp. 299–322. [Google Scholar]
- Leggett, K.E.A.; Fennessy, J.; Schneider, S. Seasonal distributions and social dynamics of elephants in the Hoanib River catchment, north-western Namibia. Afr. Zool. 2003, 38, 305–316. [Google Scholar]
- Leggett, K.E.A.; Fennessy, J.; Schneider, S. A study of animal movement in the Hoanib River catchment, northwestern Namibia. Afr. Zool. 2004, 39, 1–11. [Google Scholar]
- Krueper, D.J. Conservation Priorities in Naturally Fragmented and Human-Altered Riparian Habitats of the Arid West; General Technical Report RM-43; U.S. Department of Agriculture, Cornell Laboratory of Ornithology, Cornell University: Ithaca, NY, USA, 1993. [Google Scholar]
- Melton, D.A. The biology of aardvark (Tubulidentata-Orycteropodidae). Mammal Rev. 1976, 6, 75–88. [Google Scholar] [CrossRef]
- Haynes, G. Elephants (and extinct relatives) as earth-movers and ecosystem engineers. Geomorphology 2012, 157–158, 99–107. [Google Scholar] [CrossRef]
- Ramey, E.M.; Ramey, R.R.; Brown, L.M.; Scott, T.K. Desert elephants (Loxodonta africana) in Namibia dig wells to purify drinking water. Pachyderm 2013, 53, 66–72. [Google Scholar]
- Sabater, S.; Timoner, X.; Bornette, G.; de Wilde, M.; Stromberg, J.C.; Stella, J.C. The biota of intermittent rivers and ephemeral streams: Algae and vascular plants. In Intermittent Rivers and Ephemeral Streams, Ecology and Management; Datry, T., Bonada, N., Boulton, A., Eds.; Academic Press: London, UK, 2017; pp. 189–216. [Google Scholar]
- Sponseller, R.; Fisher, S. Drainage size, stream intermittency, and ecosystem function in a Sonoran Desert landscape. Ecosystems 2006, 9, 344–356. [Google Scholar] [CrossRef]
- Fossati, J.; Pautou, G.; Peltier, J.P. Water as resource and disturbance for wadi vegetation in a hyperarid area (Wadi Sannur, Eastern Desert, Egypt). J. Arid Environ. 1999, 43, 63–77. [Google Scholar] [CrossRef]
- Tooth, S.; Nanson, G.C. Anabranching rivers on the Northern Plains of arid central Australia. Geomorphology 1999, 29, 211–233. [Google Scholar] [CrossRef]
- Jacobson, P.J.; Jacobson, K.M.; Angermeier, P.L.; Cherry, D.S. Hydrologic influences on soil properties along ephemeral rivers in the Namib Desert. J. Arid Environ. 2000, 45, 21–34. [Google Scholar] [CrossRef]
- Jacobson, P.J.; Jacobson, K.M.; Angermeier, P.L.; Cherry, D.S. Variation in material transport and water chemistry along a large ephemeral river in the Namib Desert. Freshw. Biol. 2000, 44, 481–491. [Google Scholar] [CrossRef]
- Schaffers, A.P.; Raemaker, I.P.; Sýkora, K.V.; Ter Braak, C.J. Arthropod assemblages are best predicted by plant species composition. Ecology 2008, 89, 782–794. [Google Scholar] [CrossRef] [PubMed]
- Dudley, J.P. Seed dispersal of Acacia erioloba by African bush elephants in Hwange National Park, Zimbabwe. Afr. J. Ecol. 1999, 37, 375–385. [Google Scholar] [CrossRef]
- Datry, T.; Foulquier, A.; Corti, R.; von Schiller, D.; Tockner, K.; Mendoza-Lera, C.; Clement, J.C.; Gessner, M.O.; Moleón, M.; Stubbington, R.; et al. A global analysis of terrestrial plant litter dynamics in non-perennial waterways. Nat. Geosci. 2018, 11, 497–503. [Google Scholar] [CrossRef]
- Shumilova, O.; Zak, D.; Datry, T.; von Schiller, D.; Corti, R.; Foulquier, A.; Obrador, B.; Tockner, K.; Allan, D.C.; Altermatt, F.; et al. Simulating rewetting events in intermittent rivers and ephemeral streams: A global analysis of leached nutrients and organic matter. Glob. Chang. Biol. 2019, 1–21. [Google Scholar] [CrossRef]
- Keller, P.S.; Catalán, N.; von Schiller, D.; Grossart, H.P.; Koschorreck, M.; Obrador, B.; Frassl, M.A.; Karakaya, N.; Barros, N.; Howitt, J.A.; et al. Global CO2 emissions from dry inland waters share common drivers across ecosystems. Nat. Commun. 2020, 11, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Romaní, A.M.; Fischer, H.; Mille-Lindblom, C.; Tranvik, L.J. Interactions of bacteria and fungi on decomposing litter: Differential extracellular enzyme activities. Ecology 2006, 87, 2559–2569. [Google Scholar] [CrossRef]
- Ylla, I.; Sanpera-Calbet, I.; Vazquez, E.; Romani, A.M.; Munoz, I.; Butturini, A.; Sabater, S. Organic matter availability during pre- and post-drought periods in a Mediterranean stream. Hydrobiologia 2010, 657, 217–232. [Google Scholar] [CrossRef]
- Gallo, M.E.; Porras-Alfaro, A.; Odenbach, K.J.; Sinsabaugh, R.L. Photoacceleration of plant litter decomposition in an arid environment. Soil Biol. Biochem. 2009, 41, 1433–1441. [Google Scholar] [CrossRef]
- Del Campo, R.; Gómez, R. Exposure of wood in floodplains affects its chemical quality and its subsequent breakdown in streams. Sci. Total Environ. 2016, 543, 652–661. [Google Scholar] [CrossRef] [PubMed]
- Ortega, M.; Vidal-Abarca, M.R.; Suárez, M.L.; González-Beseran, J.L.; Ramírez-Díaz, L. Caraterísticas físico-químicas de las aguas superficiales de la Rambla del Moro después de una riada (Cuenca del Río Segura, SE de España). Limnética 1988, 4, 19–26. [Google Scholar]
- Skoulikidis, N.T.; Amaxidis, Y. Origin and dynamics of dissolved and particulate nutrients in a minimally disturbed Mediterranean river with intermittent flow. J. Hydrol. 2009, 373, 218–229. [Google Scholar] [CrossRef]
- Al-Qudah, O.M.; Walton, J.C.; Woocay, A. Chemistry and evolution of desert ephemeral stream runoff. J. Arid Environ. 2015, 122, 169–179. [Google Scholar] [CrossRef]
- Von Schiller, D.; Marcé, R.; Obrador, B.; Gómez, L.; Casas, J.P.; Acuña, V.; Koschorreck, M. Carbon dioxide emissions from dry watercourses. Inland Waters 2014, 4, 377–382. [Google Scholar] [CrossRef]
- Gómez-Gener, L.; Obrador, B.; Marcé, R.; Acuña, V.; Catalán, N.; Casas-Ruiz, J.P.; Sabater, S.; Muñoz, I.; von Schiller, D. When water vanishes: Magnitude and regulation of carbon dioxide emissions from dry temporary streams. Ecosystems 2016, 19, 710. [Google Scholar] [CrossRef]
- Austin, B.J.; Strauss, E.A. Nitrification and denitrification response to varying periods of desiccation and inundation in a western Kansas stream. Hydrobiologia 2011, 658, 183–195. [Google Scholar] [CrossRef]
- Arce, M.I.; Sánchez-Montoya, M.M.; Vidal-Abarca, M.R.; Suárez, M.L.; Gómez, R. Implications of flow intermittency on sediment nitrogen availability and processing rates in a Mediterranean head water stream. Aquat. Sci. 2014, 76, 173–186. [Google Scholar] [CrossRef]
- Arce, M.I.; Sánchez-Montoya, M.M.; Gómez, R. Nitrogen processing following experimental sediment rewetting in isolated pools in an agricultural stream of a semiarid region. Ecol. Eng. 2015, 77, 233–241. [Google Scholar] [CrossRef]
- Merbt, S.N.; Proia, L.; Prosser, J.I.; Marti, E.; Casamayor, E.O.; von Schiller, D. Stream drying drives microbial ammonia oxidation and first-flush nitrate export. Ecology 2016, 97, 2192–2198. [Google Scholar] [CrossRef]
- Schimel, J.P. Life in dry soils: Effects of drought on soil microbial communities and processes. Annu. Rev. Ecol. Evol. Syst. 2018, 49, 409–432. [Google Scholar] [CrossRef]
- Gionchetta, G.; Oliva, F.; Menéndez, M.; Lopez Laseras, P.; Romaní, A.M. Key role of streambed moisture and flash storms for microbial resistance and resilience to long-term drought. Freshw. Biol. 2019, 64, 306–322. [Google Scholar] [CrossRef]
- Costanza, R.; d’Arge, R.; de Groot, R.; Farber, S.; Grasso, M.; Hannon, B.; Limburg, K.; Naeem, S.; Oneill, R.V.; Paruelo, J.; et al. The value of the world’s ecosystem services and natural capital. Nature 1997, 387, 253–260. [Google Scholar] [CrossRef]
- Millennium Ecosystem Assessment (MA). Ecosystems and Human Well-Being: Biodiversity Synthesis; World Resources Institute: Washington, DC, USA, 2005. [Google Scholar]
- Spanish National Ecosystem Assessment. Ecosystems and Biodiversity for Human Wellbeing. Synthesis of the Key Findings; Biodiversity Foundation of the Spanish Ministry of Agriculture, Food and Environment: Madrid, Spain, 2013. Available online: http://www.ecomilenio.es/ecosystems-and-biodiversity-for-human-wellbeing-snea-synthesis-of-key-findings-download/3661 (accessed on 1 September 2020).
- Díaz, S.; Demissew, S.; Carabias, J.; Joly, C.; Lonsdale, M.; Ash, N.; Larigauderie, A.; Adhikari, J.R.; Arico, S.; Baldi, A.; et al. The IPBES conceptual framework—Connecting nature and people. Curr. Opin. Environ. Sustain. 2015, 14, 1–16. [Google Scholar] [CrossRef]
- De Groot, R.S.; Wilson, M.A.; Boumans, R.M.J. A typology for the classification, description and valuation of ecosystem functions, goods and services. Ecol. Econ. 2002, 41, 393–408. [Google Scholar] [CrossRef]
- Powell, D.M.; Brazie, R.; Parsons, A.; Wainwright, J.; Nichols, M. Sediment transfer and storage in dryland headwater streams. Geomorphology 2007, 88, 152–166. [Google Scholar] [CrossRef]
- Sánchez, M.A.; Sánchez-Zapata, J.A.; Díez de Revenga, E. El conejo (Oryctolagus cuniculus) en la Región de Murcia. In Actas del II Congreso de la Naturaleza de la Región de Murcia; ANSE, Ed.; ANSE: Murcia, Spain, 2004; pp. 169–179. Available online: https://www.researchgate.net/profile/Jose_Sanchez-Zapata/publication/267832938_El_conejo_Oryctolagus_cuniculus_en_la_Region_de_Murcia/links/54dcfae80cf25b09b912e3c0/El-conejo-Oryctolagus-cuniculus-en-la-Region-de-Murcia.pdf (accessed on 1 September 2020).
- Salinas, M.J.; Guirado, J. Riparian Plant Restoration in Summer-Dry Riverbeds of Southeastern Spain. Restor. Ecol. 2002, 10, 695–702. [Google Scholar] [CrossRef]
- Burns, R.G.; DeForest, J.L.; Marxsen, J.; Sinsabaugh, R.L.; Stromberger, M.E.; Wallenstein, M.D.; Weintraub, M.N.; Zoppini, A. Soil enzymes in a changing environment: Current knowledge and future directions. Soil Biol. Biochem. 2013, 58, 216–234. [Google Scholar] [CrossRef]
- Montes, C.; García, M.R. La Evaluación de los Ecosistemas del Milenio en Andalucía Haciendo Visibles los vínculos entre la Naturaleza y el Bienestar Humano. Junta de Andalucía. 2012. Available online: http://www.juntadeandalucia.es/medioambiente/site/portalweb/menuitem.7e1cf46ddf59bb227a9ebe205510e1ca/?vgnextoid=de5b4a6acd72f210VgnVCM1000001325e50aRCRD&vgnextchannel=a349e156217d4310VgnVCM2000000624e50aRCRD (accessed on 27 March 2020).
- Montes, C.; Santos-Martin, F.; Benayas, J. Ecosistemas y Biodiversidad para el Bienestar Humano. Evaluación de los Ecosistemas del Milenio de España. Fundación Biodiversidad; Ministerio de Medio Ambiente y Medio Rural y Marino: Madrid, Spain, 2011. [CrossRef]
- UK NEA. The UK National Ecosystem Assessment: Synthesis of the Key Findings; UNEP-WCMC: Cambridge, UK, 2011; Available online: https://www.researchgate.net/publication/317236524_UK_National_Ecosystem_Assessment_understanding_nature’s_value_to_society_Synthesis_of_key_findings (accessed on 1 September 2020).
- Landers, D.H.; Nahlik, A.M. Final Ecosystem Goods and Services Classification System (FEGS-CS)(EPA/600/R-13/ORD-004914); Office of Research and Development, U.S. Environmental Protection Agency: Washington, DC, USA, 2013. Available online: http://refhub.elsevier.com/B978-0-12-803835-2.00018-8/rf0255 (accessed on 27 March 2020).
- Vidal-Abarca, M.R.; Suárez, M.L. Which are, what is their status and what can we expect from ecosystem services provided by Spanish rivers and riparian areas? Biodivers. Conserv. 2013, 22, 2469–2503. [Google Scholar] [CrossRef]
- Harrison, P.A.; Vandewalle, M.; Sykes, M.T.; Berry, P.M.; Bugter, R.; de Bello, F.; Feld, C.K.; Grandin, U.; Harrington, R.; Haslett, J.R.; et al. Identifying and prioritising services in European terrestrial and freshwater ecosystems. Biodivers. Conserv. 2010, 19, 2791–2821. [Google Scholar] [CrossRef]
- IPBES. Summary for Policymakers of the Regional Assessment Report on Biodiversity and Ecosystem Services for Africa of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services; Archer, E., Dziba, L.E., Mulongoy, K.J., Maoela, M.A., Walters, M., Eds.; IPBES: Bonn, Germany; IPBES Secretariat: Bonn, Germany, 2018. [Google Scholar]
- IPBES. Summary for Policymakers of the Regional Assessment Report on Biodiversity and Ecosystem Services for Asia and the Pacific of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services; Karki, M., Senaratna Sellamuttu, S., Okayasu, S., Suzuki., W., Eds.; IPBES: Bonn, Germany; IPBES Secretariat: Bonn, Germany, 2018. [Google Scholar]
- IPBES. Summary for Policymakers of the Regional Assessment Report on Biodiversity and Ecosystem Services for the Americas of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services; Rice, J., Seixas, S., Zaccagnini, M.E., Bedoya-Gaitán, M., Valderrama, N., Eds.; IPBES Secretariat: Bonn, Germany, 2018. [Google Scholar]
- IPBES. Summary for Policymakers of the Regional Assessment Report on Biodiversity and Ecosystem Services for Europe and Central Asia of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services; Fischer, M., Rounsevell, M., Torre-Marin, A., Mader, A., Eds.; IPBES Secretariat: Bonn, Germany, 2018. [Google Scholar]
- Koundouri, P.; Boulton, A.J.; Datry, T.; Souliotis, I. Ecosystem services, values, and societal perceptions of intermittent rivers and ephemeral streams. In Intermittent Rivers and Ephemeral Streams, Ecology and Management; Datry, T., Bonada, N., Boulton, A., Eds.; Academic Press: London, UK, 2017; pp. 455–476. [Google Scholar]
- Shackleton, C.; Shackleton, S.; Gambiza, J.; Nel, E.; Rowntree, K.; Urquhart, P. Links between Ecosystem Services and Poverty Alleviation: Situation Analysis for Arid and Semi-Arid Lands in Southern Africa; Consortium on Ecosystems and Poverty in Sub-Saharan (CEPSA). 2008. Available online: uct.ac.za/sites/default/files/image_tool/images/192/Shackleton%20et%20al%20%202008%20Southern%20African%20Ecosystem%20Services%20and%20Poverty%20report_CEPSA.pdf (accessed on 1 September 2020).
- Iniesta-Arandia, I.; García del Amo, D.; García-Nieto, A.P.; Piñeiro, C.; Montes, C.; Martín-López, B. Factors influencing local ecological knowledge maintenance in Mediterranean watersheds: Insights for environmental policies. Ambio 2014, 44, 285–296. [Google Scholar] [CrossRef]
- Alkhaddar, R.M. Water harvesting in Jordan using earth ponds. Waterlines 2003, 22, 19–21. [Google Scholar] [CrossRef]
- Nasiri, F.; Mafakheri, M.S. Qanat water supply systems: A revisit of sustainability perspectives. Environ. Syst. Res. 2015, 4, 13. [Google Scholar] [CrossRef]
- Seely, M.; Henderson, J.; Heyns, P.; Jacobson, P.; Nakale, T.; Nantanga, K.; Schachtschneider, K. Ephemeral and endoreic river systems: Relevance and management challenges. In Transboundary Rivers, Sovereignty and Development: Hydropolitical Drivers in the Okavango River Basin; Turton, A., Ashton, P., Cloete, E., Eds.; African Water Issues Research Unit: Pretoria, South Africa, 2003; pp. 187–212. [Google Scholar]
- Ito, A. Distribution of the Nara plant that affect the life of the Topnaar people in the lower Kuiseb River, Namib Desert. Afr. Study Monogr. Suppl. 2005, 30, 65–75. [Google Scholar]
- Ffolliott, P.F.; DeBano, L.F. Riparian areas of the southwestern United States: Hydrology, Ecology, and Management; CRC Press: Boca Raton, FL, USA, 2003. [Google Scholar]
- Mizuno, K.; Yamagata, K. Vegetation succession and plant use in relation to environmental changes along the Kuiseb River in the Namib Desert. Afr. Study Monogr. 2005, 30, 3–14. [Google Scholar] [CrossRef]
- Yamagata, K. Recent grain-size coarsening of floodplain deposits and forest decline along the Kuiseb River, Namib Desert, Namibia. Afr. Study Monogr. 2010, 40, 19–30. [Google Scholar]
- Jacobson, P.J.; Jacobson, K.M. Hydrologic controls of physical and ecological processes in Namib Desert ephemeral rivers: Implications for conservation and management. J. Arid Environ. 2013, 93, 80–93. [Google Scholar] [CrossRef]
- Bond, N.R.; Cottingham, P. Ecology and Hydrology of Temporary Streams: Implications for Sustainable Water Management; eWater Technical Report; eWater Cooperative Research Centre: Canberra, Australia, 2008; Available online: https://www.researchgate.net/profile/Nick_Bond2/publication/242107251_Ecology_and_Hydrology_of_Temporary_Streams_Implications_for_Sustainable_Water_Management/links/548b83780cf214269f1dd657/Ecology-and-Hydrology-of-Temporary-Streams-Implications-for-Sustainable-Water-Management.pdf (accessed on 1 September 2020).
- Hassan, M.A.; Egozi, R. Impact of wastewater discharge on the channel morphology of ephemeral streams. Earth Surf. Proc. Land. 2001, 26, 1285–1302. [Google Scholar] [CrossRef]
- Skoulikidis, N.T.; Sabater, S.; Datry, T.; Morais, M.M.; Buffagni, A.; Dörflinger, G.; Zogaris, S.; Sánchez-Montoya, M.M.; Bonada, N.; Kalogianni, E.; et al. Non-perennial Mediterranean rivers in Europe: Status, pressures, and challenges for research and management. Sci. Total Environ. 2017, 577, 1–18. [Google Scholar] [CrossRef]
- National Research Council. Riparian Areas: Functions and Strategies for Management; The National Academies Press: Washington, DC, USA, 2002. [Google Scholar] [CrossRef]
- Krueper, D.J. Effects of livestock management on Southwestern riparian ecosystems. In Shaw DW, Finch DM (Tech. Coords) Desired Future Conditions for Southwestern Riparian Ecosystems: Bringing Interests and Concerns Together. Gen. Tech. Rep. RM-GTR-272; US Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: Fort Collins, CO, USA, 1996; pp. 281–301. [Google Scholar]
- Maldonado, M.B.; Aranibar, J.N.; Serrano, A.M.; Chacoff, N.P.; Vázquez, D.P. Dung beetles and nutrient cycling in a dryland environment. Catena 2019, 179, 66–73. [Google Scholar] [CrossRef]
- Zaragoza, E.B. Anthropic occupation of ravines and gullies in the Bajo Segura county (Alicante): Analysis and diagnosis of the situation in some of its towns. Investig. Geogr. 1997, 17, 149–162. [Google Scholar] [CrossRef]
- Sánchez-Montoya, M.M.; Moleón, M.; Sánchez-Zapata, J.A.; Tockner, K. Dry riverbeds: Corridors for terrestrial vertebrates. Ecosphere 2016, 7, e01508. [Google Scholar] [CrossRef]
- Chiu, M.C.; Leigh, C.; Mazor, R.; Cid, N.; Resh, V. Anthropogenic threats to intermittent rivers and ephemeral streams. In Intermittent Rivers and Ephemeral Streams, Ecology and Management; Datry, T., Bonada, N., Boulton, A., Eds.; Academic Press: London, UK, 2017; pp. 433–454. [Google Scholar]
- Milton, S.J.; Dean, W.R.J. Plant invasions in arid areas: Special problems and solutions: A South African perspective. Biol. Invasions 2010, 12, 3935–3948. [Google Scholar] [CrossRef]
- Coffman, G.C. Factors Influencing Invasion of Giant Reed (Arundo donax) in Riparian Ecosystems of Mediterranean-Type Climate Regions. Ph.D. Thesis, University of California, Los Angeles, CA, USA, 2007. [Google Scholar] [CrossRef]
- Norström, A.V.; Cvitanovic, C.; Löf, M.F.; West, S.; Wyborn, C.; Balvanera, P.; Bednarek, A.T.; Bennett, E.M.; Biggs, R.; de Bremond, A.; et al. Principles for knowledge co-production in sustainability research. IJEWE 2020, 3. [Google Scholar] [CrossRef]
- Hall, C.M.; Boyd, S.W. Nature-Based Tourism in Peripheral Areas: Development or Disaster? Channel View Publications: Bristol, UK, 2005; Volume 21. [Google Scholar]
- Stubbington, R.; Chadd, R.; Cid, N.; Csabai, Z.; Miliša, M.; Morais, M.; Munné, A.; Pařil, P.; Pešić, V.; Tziortzis, I.; et al. Biomonitoring of intermittent rivers and ephemeral streams in Europe: Current practice and priorities to enhance ecological status assessments. Sci. Total Environ. 2018, 618, 1096–1113. [Google Scholar] [CrossRef] [PubMed]
- Ollero, A.; Ballarín, D.; Díaz Bea, E.; Mora, D.; Sánchez Fabre, M.; Acín, V.; Echeverría, M.T.; Granado, D.; González de Matauco, A.I.; Sánchez, L.; et al. IHG: Un índice para la valoración hidrogeomorfológica de sistemas fluviales. Limnetica 2008, 27, 171–188. [Google Scholar] [CrossRef]
- Suarez, M.L.; Vidal-Abarca, M.R. Índice para valorar el estado de conservación de las ramblas mediterráneas (Índice de alteración de ramblas, IAR). Tecnol. Agua 2008, 293, 67–78. [Google Scholar]
- Suárez, M.L.; Vidal-Abarca, M.R.; Navarro, I.; Gómez, R.; López-Bermúdez, F. Los arroyos de Baja California Sur (México): Caracterización ambiental y aplicación de un índice de Estado de Alteración (IAR). C y G 2010, 24, 63–77. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Definition | References |
|---|---|
| Ephemeral washes are those watercourses that flow briefly in direct response to precipitation and are distinguished from intermittent streams because they are always above the phreatic level. | [1] |
| The ephemeral river, entirely dependent upon desert storms for surface flow. | [41] |
| Wadi: Course that only transports water a few hours after rain. | [42] |
| An ephemeral stream was one flowing only in direct response to precipitation; with measurable discharge occurring less than 10% of the time. | [43] |
| Ephemeral streams are apparent only after periods of high storm intensity or snow melts; they seldom contain any animal or plant life. | [44] |
| They distinguished ‘temporary’ or ‘intermittent’ rivers, with more-or-less regular, seasonally intermittent discharge, from ‘ephemeral’ or ‘episodic’ rivers, which flow only after unpredictable rainfall. | [45] |
| The author provided a definition based upon annual flow duration, distinguishing ephemeral systems, flowing < 20% of the year, from intermittent systems flowing between 20–80% annually. | [46] |
| The episodic systems, restricted to arid and hyper-arid regions, contain water on an unpredictable basis. The authors discouraged the use of the term ephemeral as a synonym for episodic and the use of the term temporary as a synonym for intermittent. | [47] |
| Ephemeral: Rivers that flow for less time than they are dry. Flow or flood for short periods of most years in a five-year period, in response to unpredictable high rainfall events. Support a series of pools in parts of the channel. Episodic: Highly flashy systems that flow or flood only in response to extreme rainfall events, usually high in their catchments. May not flow in a five-year period or may flow only once in several years. | [11] |
| Term ‘ephemeral’ used by hydrogeologists to encompass running waters that are permanently disconnected from the GWT (groundwater table), for example, dryland rivers that flow only for a short period during and after rainstorms. | [48] |
| Ramblas can be dry for many years and only transport water for a few days as a result of heavy rainfall. | [49] |
| Ephemeral (or episodic) streams are usually dry except for several days immediately following precipitation. | [50] |
| Ephemeral (dry washes): A stream or portion of a stream which flows briefly in direct response to precipitation in the immediate vicinity, and whose channel is, at all times, above the groundwater reservoir. Ephemeral streams are unique in that they lack permanent flow except in response to rainfall events. | [51] |
| Ephemeral freshwater systems (EFS). In the highest reaches of these segments, stream flow is generally less than permanent. | [52] |
| Streams can be classified as perennial, flowing all the time or, at least at all times, except extreme droughts; intermittent, flowing some of the time and receiving water from groundwater; or ephemeral, flowing rarely and not receiving input from groundwater. Ephemeral, not holding surface water most of the year. | [53] |
| Temporary systems cover those that are: ephemeral—ones which receive water for a short period very occasionally and highly unpredictably; episodic—those that fill occasionally, and which may last months or even years. | [54] |
| Ephemeral streams are those that only flow during and in immediate response to precipitation events. The groundwater table is situated below the streambed so that the channel never receives groundwater discharge. Ephemeral streams are always losing streams. | [55] |
| Intermittent (periodically dry, fed from seasonal flow) and ephemeral (periodically dry, event-based flow) streams are predominantly located in the uppermost reaches of a watershed. | [56] |
| Episodic-Ephemeral: Water flow and pools are short-lived and occasional. Most of the organisms found are opportunistic, adapted to a quick development of their biological cycle. Biological quality assessment needs other methods beyond the customary study of aquatic fauna (e.g., desiccation-resistant stages of aquatic fauna or terrestrial fauna). | [57] |
| The flow in ephemeral streams is episodic, with limited and stochastic water supply. | [58] |
| Episodic or ephemeral—those that flow and maintain water on a largely unpredictable basis, depending on precipitation events, and with hydrological continuity for only a short period of time (usually days to weeks). | [59] |
| The author use ‘temporary’ as a collective adjective to refer to all waterways that cease surface flow at some points in time and space along their course, and ‘ephemeral’ for the subset of temporary streams that flow only briefly (days to weeks), usually after rainfall and runoff. | [60] |
| The authors use the term intermittent rivers to refer to all temporary, ephemeral, seasonal, and episodic streams and rivers in defined channels. | [32] |
| Ephemeral stream: Flow duration < 2 months; Pool duration: variable; dry period: >10 months/year. | [61] |
| Perennial and ephemeral streams constitute the limits of a continuum composed of a multitude of intermediate flow regimes, which can be described in a different way. | [62] |
| Wadis are dynamic systems with hydrological regimes shifting between long periods of drought and sudden flash flooding. | [63] |
| The term ephemeral implies a shorter flow duration and lower predictability than intermittent but there are not fixed boundaries. | [64] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vidal-Abarca, M.R.; Gómez, R.; Sánchez-Montoya, M.M.; Arce, M.I.; Nicolás, N.; Suárez, M.L. Defining Dry Rivers as the Most Extreme Type of Non-Perennial Fluvial Ecosystems. Sustainability 2020, 12, 7202. https://doi.org/10.3390/su12177202
Vidal-Abarca MR, Gómez R, Sánchez-Montoya MM, Arce MI, Nicolás N, Suárez ML. Defining Dry Rivers as the Most Extreme Type of Non-Perennial Fluvial Ecosystems. Sustainability. 2020; 12(17):7202. https://doi.org/10.3390/su12177202
Chicago/Turabian StyleVidal-Abarca, María Rosario, Rosa Gómez, María Mar Sánchez-Montoya, María Isabel Arce, Néstor Nicolás, and María Luisa Suárez. 2020. "Defining Dry Rivers as the Most Extreme Type of Non-Perennial Fluvial Ecosystems" Sustainability 12, no. 17: 7202. https://doi.org/10.3390/su12177202
APA StyleVidal-Abarca, M. R., Gómez, R., Sánchez-Montoya, M. M., Arce, M. I., Nicolás, N., & Suárez, M. L. (2020). Defining Dry Rivers as the Most Extreme Type of Non-Perennial Fluvial Ecosystems. Sustainability, 12(17), 7202. https://doi.org/10.3390/su12177202