Bioclimatology, Structure, and Conservation Perspectives of Quercus pyrenaica, Acer opalus subsp. Granatensis, and Corylus avellana Deciduous Forests on Mediterranean Bioclimate in the South-Central Part of the Iberian Peninsula

 ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Subjects
2.2. Origin and Structure of the Vegetation Data
2.3. Climatic, Geographic, and Topographic Data
- T: Mean annual temperature in degrees centigrade
- M: Mean temperature of the maximums
- m: Mean temperature of the minimums
- Tmax: Mean temperature of the warmest month of the year
- Tmin: Mean temperature of the coldest month of the year
- Tcmax: Mean maximum temperature of the month with the greatest temperature difference in the year
- Tcmin: Mean minimum temperature of the month with the greatest temperature difference in the year
- Tn: Negative annual temperature (sum of the months with a mean temperature below 0 °C in tenths of degrees centigrade)
- Tp: Positive annual temperature (sum of the months with a mean temperature above 0 °C in tenths of degrees centigrade)
- Tpd: Positive temperature of the driest quarter of the year, in tenths of degrees centigrade
- Tpd1: Positive temperature of the driest month of the year, in tenths of degrees centigrade
- Tpd2: Positive temperature of the driest two months of the year, in tenths of degrees centigrade
- Pp: Positive annual precipitation (of the months in Ti above 0 °C)
- PE: Thornthwaite’s potential annual evapotranspiration index
- PEs: Potential evapotranspiration index of the summer quarter
- PE1−12: Potential monthly evapotranspiration index 1 = January, 2 = February, … 12 = December
- Annual aridity index (Iar);
- Diurnality index (Id): the difference between the mean maximum temperature (Tc max) and the mean minimum temperature (Tc min) of the months with the greatest temperature differences in the year; that is, the month with the greatest daily swing or interval between maximum and minimum temperatures.
- Humidity index: the percentage of precipitation above or below the annual precipitation (P) compared to the annual evapotranspiration (PE)
- Mediterraneity index (Im): the coefficient between the value for Thornthwaite’s mean summer evapotranspiration (PEs), and precipitation in mm in the same period (Ps).
- Mediterraneity index for the month of July (Im1) and for July and August (Im2);
- Annual evapotranspiration index (Ioe): the coefficient between mean precipitation (P) and potential annual evapotranspiration (PE, Thornthwaite).
- Ombrothermic index of the warmest month of the summer quarter (Ios1): ombrothermic index of the warmest two months of the summer quarter (Ios2), ombrothermic index of the summer quarter (Ios3), and ombrothermic index of the summer quarter and the previous month (Ios4).
- Monthly ombrothermic indices (Iom1–12), with i = 1 = January 2 = February, …, 12 = December.
- Annual ombrothermic index (Io): the coefiicient between the total mean precipitation in mm in the months with a mean temperature above zero degrees centigrade (Pp) and the sum of the mean monthly temperatures above zero degrees in tenths of a degree (Tp).
- Simple continentality index (Ic): the continentality index expresses in degrees centigrade the difference or oscillation between the mean temperature of the warmest month (Tmax) and the mean temperature of the coldest month of the year (Tmin).
- Thermicity index (It): this index describes the intensity of cold, which is a limiting factor for many plants and plant communities.
2.4. Analysis of the Subjects (Relevés from Deciduous Forests)
3. Results
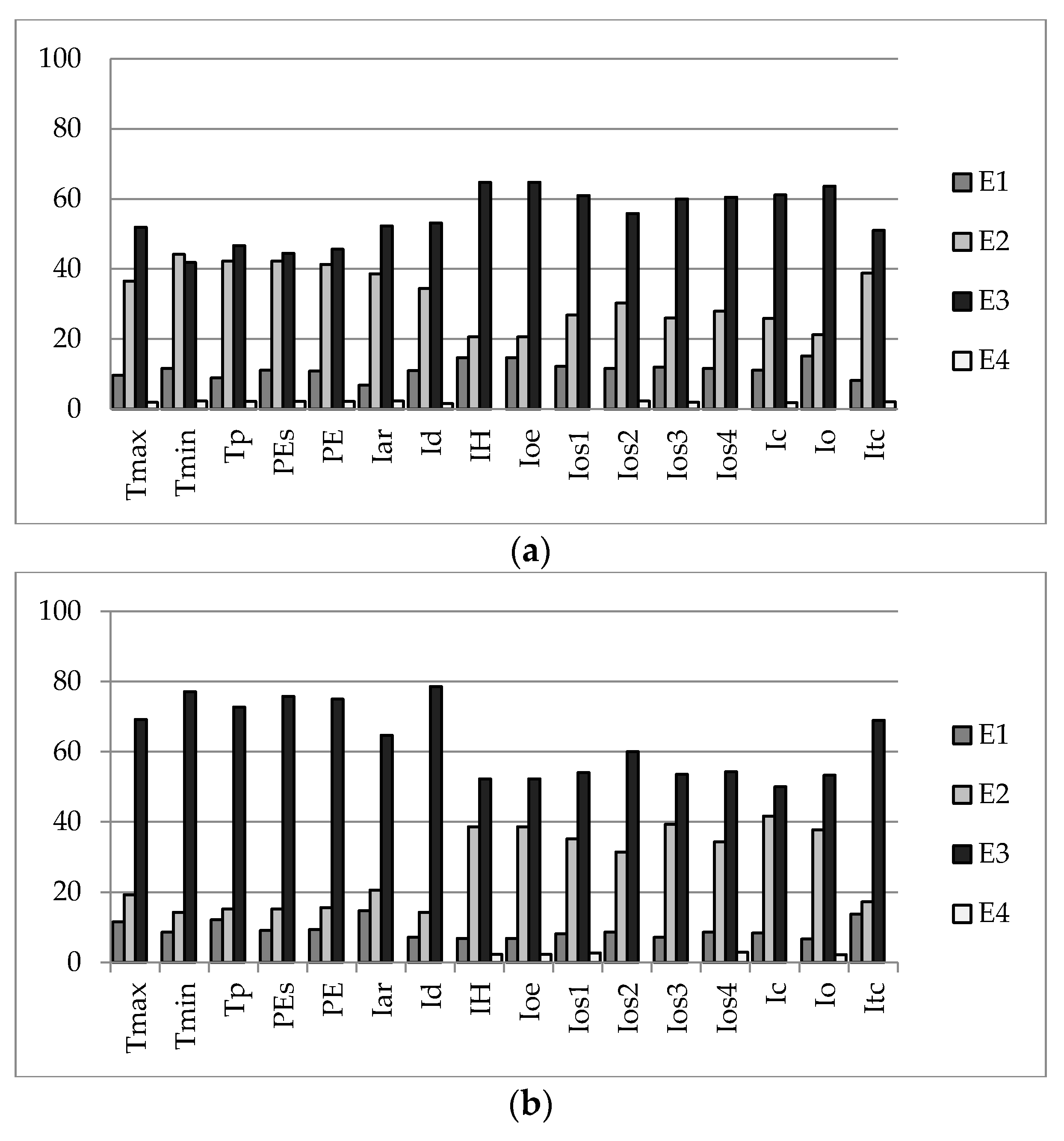
3.1. Correlation between Characteristic Taxa, Stratum, and Bioclimatic Variables
3.1.1. Spearman’s Correlation Analysis
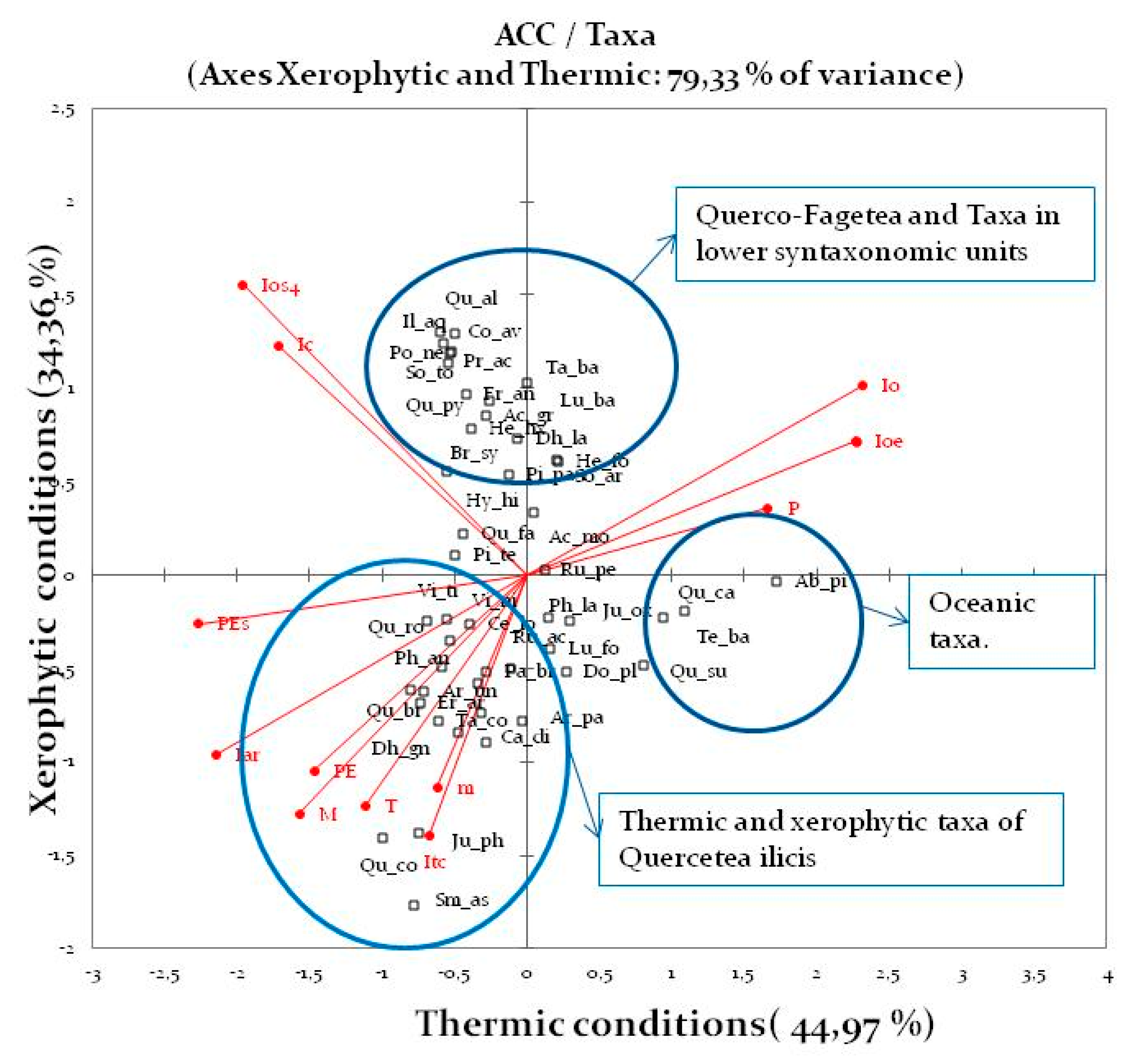
3.1.2. Canonical Correlation Analysis (CCA)
3.2. ANOVA Analysis of Introgression of Quercetea ilicis Species in Deciduous Forests
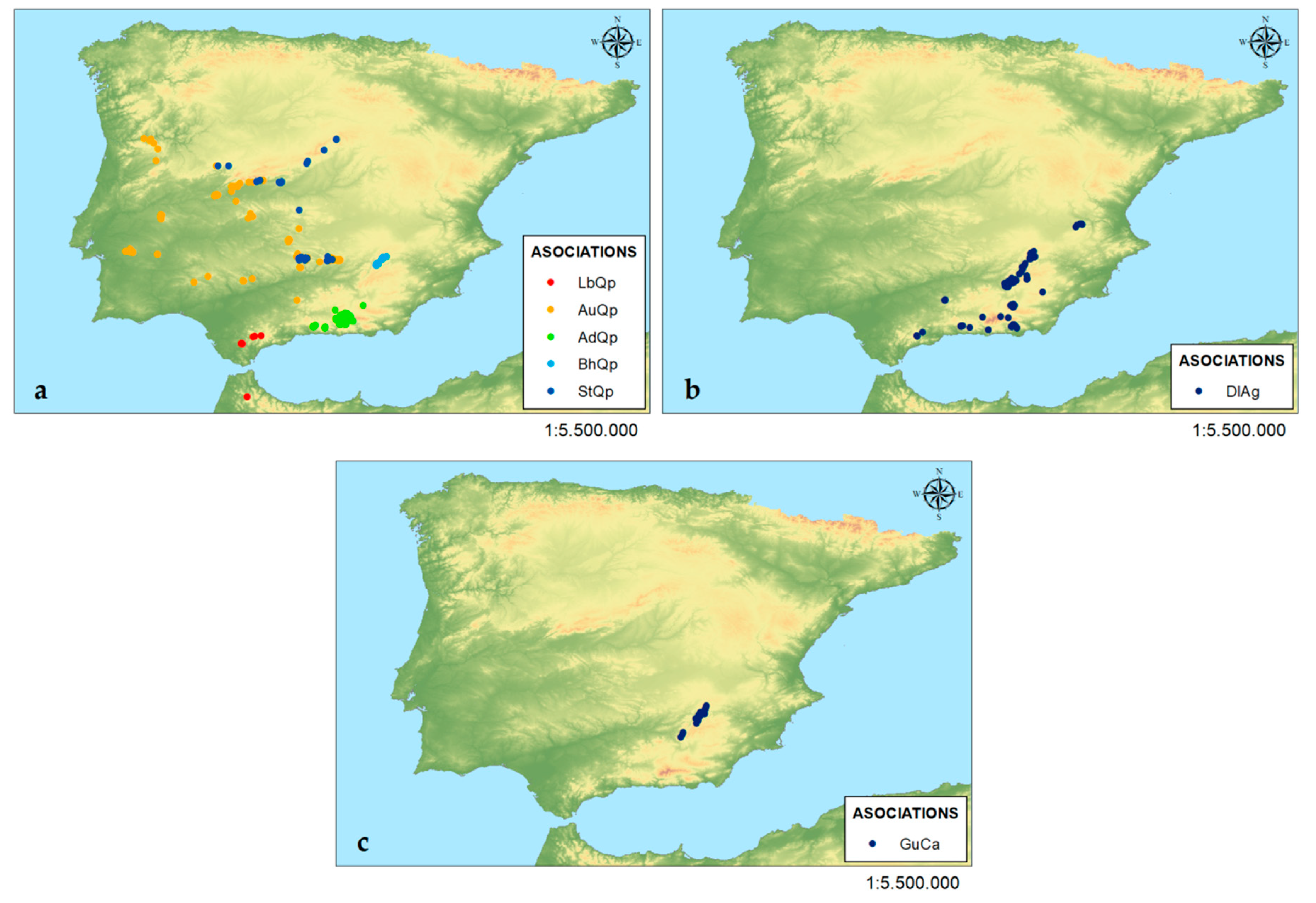
3.3. Frequency of Occurrence and Average Cover in the Floristic Composition of Each Plant Community
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | IVI | Phytosociological Class | Stratum | Relative Frequency (%) | Average Coverage (%) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AdQp | AuQp | BhQp | LbQp | StQp | DlAg | GuCa | AdQp | AuQp | BhQp | LbQp | StQp | DlAg | GuCa | ||||
| Quercus pyrenaica Willd. | 12.59 | Q-F | E1 | 100.00 | 97.20 | 100.00 | 100.00 | 100.00 | 1.79 | - | 53.38 | 53.20 | 45.83 | 65.00 | 72.91 | 18.50 | - |
| Acer opalus subsp. granatense (Boiss.) Font Quer and Rothm. | 6.29 | Q-F | E2 | 26.23 | 0.70 | 38.89 | - | - | 83.04 | 29.41 | 22.38 | 45.00 | 10.29 | - | - | 35.27 | 14.10 |
| Corylus avellana L. | 5.00 | Q-F | E2 | - | - | - | - | 4.65 | 5.36 | 100.00 | - | - | - | - | 6.00 | 19.67 | 46.65 |
| Quercus faginea subsp. faginea Lam. | 4.85 | Q-F | E1 | 3.28 | 13.29 | 5.56 | - | 4.65 | 48.21 | 47.06 | 12.00 | 21.00 | 25.00 | - | 6.00 | 28.19 | 16.25 |
| Brachypodium sylvaticum (Huds.) P. Beauv. | 4.37 | S-P | E3 | 4.92 | 27.27 | 16.67 | - | 27.91 | 32.14 | 44.12 | 8.00 | 12.87 | 20.67 | - | 10.08 | 19.00 | 23.87 |
| Rubia peregrina subsp. peregrina L. | 4.32 | Q-I | E2 | 13.11 | 37.06 | 61.11 | - | 9.30 | 30.36 | 41.18 | 11.38 | 10.21 | 16.91 | - | 6.00 | 12.21 | 10.79 |
| Daphne laureola subsp. latifolia (Coss.) Rivas Mart. | 4.30 | Q-F | E2 | 14.75 | - | 44.44 | - | - | 53.57 | 41.18 | 37.67 | - | 14.63 | - | - | 17.57 | 16.71 |
| Arbutus unedo L. | 4.26 | Q-I | E2 | - | 50.35 | - | 90.00 | 16.28 | 2.68 | - | - | 20.39 | - | 9.33 | 13.14 | 8.00 | - |
| Helleborus foetidus L. | 4.10 | Q-F | E3 | 36.07 | 0.70 | 44.44 | - | - | 50.89 | 79.41 | 8.73 | 25.00 | 12.13 | - | - | 13.46 | 10.22 |
| Daphne gnidium L. | 3.64 | Q-I | E2 | 11.48 | 46.15 | 5.56 | 40.00 | 23.26 | 7.14 | 14.71 | 9.43 | 10.62 | 12.00 | 7.50 | 7.80 | 8.25 | 7.20 |
| Quercus faginea subsp. alpestris (Boiss.) Maire | 3.55 | Q-F | E1 | - | 0.70 | 83.33 | - | - | 5.36 | 2.94 | - | 45.00 | 26.00 | - | - | 43.00 | 25.00 |
| Tamus communis L. | 3.47 | Q-F | E2 | - | 37.76 | - | - | 32.56 | 15.18 | 23.53 | - | 11.04 | - | - | 10.36 | 11.00 | 9.00 |
| Paeonia broteri Boiss. & Reut. | 3.45 | Q-I | E3 | - | 32.87 | 11.11 | - | 53.49 | 16.96 | - | - | 11.99 | 9.00 | - | 13.87 | 7.42 | - |
| Hedera helix subsp. helix L. | 3.36 | Q-F | E2 | 8.20 | 2.10 | 50.00 | - | 6.98 | 29.46 | 67.65 | 6.00 | 16.33 | 12.89 | - | 8.00 | 11.42 | 16.35 |
| Quercus suber var. suber L. | 3.34 | Q-I | E1 | - | 35.66 | - | 60.00 | - | - | - | - | 18.95 | - | 10.00 | - | - | - |
| Myrtus communis L. | 3.04 | Q-I | E2 | - | 2.10 | - | - | - | - | - | - | 31.67 | - | - | - | - | - |
| Aristolochia paucinervis Pomel | 2.97 | S-P | E3 | 26.23 | 20.28 | 11.11 | - | 41.86 | 8.04 | - | 9.75 | 9.52 | 6.00 | - | 14.28 | 11.44 | - |
| Quercus broteroi (Cout.) Rivas Mart. | 2.88 | Q-I | E1 | - | 20.28 | - | - | 13.95 | - | - | - | 22.02 | - | - | 13.17 | - | - |
| Acer monspessulanum subsp. monspessulanum L. | 2.88 | Q-F | E2 | - | 15.38 | - | - | 20.93 | 20.54 | - | - | 14.10 | - | - | 15.11 | 15.78 | - |
| Primula acaulis (L.) L. | 2.80 | Q-F | E3 | - | 0.70 | 44.44 | - | 6.98 | 25.89 | 73.53 | - | 6.00 | 13.63 | - | 6.00 | 10.38 | 13.48 |
| Erica arborea L. | 2.79 | Q-I | E2 | - | 27.27 | - | 30.00 | 27.91 | - | - | - | 14.67 | - | 10.00 | 9.00 | - | - |
| Teucrium scorodonia subsp. scorodonia L. | 2.79 | Q-F | E3 | - | 33.57 | - | - | 44.19 | - | - | - | 11.04 | - | - | 10.58 | - | - |
| Ilex aquifolium L. | 2.76 | Q-F | E2 | - | 0.70 | 55.56 | 10.00 | 11.63 | 12.50 | 52.94 | - | 12.00 | 14.10 | 12.00 | 9.60 | 9.50 | 22.28 |
| Doronicum plantagineum L. | 2.74 | Q-I | E3 | 54.10 | 12.59 | - | - | 44.19 | - | - | 6.73 | 8.72 | - | - | 10.53 | - | - |
| Sorbus aria (L.) Crantz | 2.70 | Q-F | E1 | 24.59 | - | 22.22 | - | 13.95 | 30.36 | 8.82 | 16.13 | - | 13.75 | - | 10.00 | 10.53 | 8.00 |
| Quercus coccifera subsp. coccifera L. | 2.67 | Q-I | E2 | - | 4.20 | - | - | - | 5.36 | - | - | 28.33 | - | - | - | 19.67 | - |
| Luzula forsteri subsp. forsteri (Sm.) DC. in Lam. & DC. | 2.67 | Q-F | E3 | 21.31 | 13.29 | - | - | 72.09 | - | - | 8.85 | 9.84 | - | - | 12.16 | - | - |
| Pistacia terebinthus L. | 2.41 | Q-I | E2 | 1.64 | 6.29 | - | - | 6.98 | 17.86 | 11.76 | 12.00 | 11.57 | - | - | 14.33 | 17.60 | 9.00 |
| Ruscus aculeatus L. | 2.30 | Q-I | E3 | - | 25.17 | - | - | 11.63 | 5.36 | 2.94 | - | 11.48 | - | - | 6.00 | 15.33 | 12.00 |
| Holcus mollis subsp. mollis L. | 2.20 | Q-F | E3 | 3.28 | 11.19 | - | 20.00 | 16.28 | - | - | 6.00 | 16.50 | - | 6.00 | 19.71 | - | - |
| Carex distachya Desf. | 2.16 | Q-I | E3 | 3.28 | 13.99 | - | 50.00 | 39.53 | - | - | 6.00 | 9.35 | - | 10.80 | 12.53 | - | - |
| Piptatherum paradoxum (L.) P. Beauv. | 2.13 | Q-I | E3 | - | - | 5.56 | - | - | 14.29 | 17.65 | - | - | 6.00 | - | - | 19.31 | 11.00 |
| Pistacia lentiscus L. | 2.11 | Q-I | E2 | - | - | - | - | - | 3.57 | - | - | - | - | - | - | 21.75 | - |
| Phillyrea angustifolia L. | 2.09 | Q-I | E2 | - | 27.97 | - | - | - | 0.89 | - | - | 10.93 | - | - | - | 6.00 | - |
| Juniperus oxycedrus subsp. oxycedrus L. | 1.89 | Q-I | E2 | - | 8.39 | 5.56 | - | 9.30 | 16.07 | 2.94 | - | 11.58 | 12.00 | - | 6.00 | 8.33 | 12.00 |
| Paeonia officinalis subsp. microcarpa (Boiss. & Reut.) Nyman | 1.82 | Q-F | E3 | - | - | 33.33 | - | 6.98 | 17.86 | 11.76 | - | - | 9.00 | - | 10.00 | 10.55 | 6.00 |
| Rosa sicula Tratt. | 1.81 | Q-I | E2 | 1.64 | - | - | - | - | 12.50 | - | 12.00 | - | - | - | - | 14.50 | - |
| Smilax aspera L. | 1.79 | Q-I | E2 | - | 6.29 | - | 30.00 | 4.65 | 2.68 | - | - | 14.33 | - | 10.00 | 9.00 | 14.33 | - |
| Rhamnus alaternus subsp. alaternus L. | 1.76 | Q-I | E2 | - | 2.80 | - | - | - | 5.36 | - | - | 21.75 | - | - | - | 12.17 | - |
| Olea europaea var. sylvestris (Mill.) Lehr | 1.75 | Q-I | E2 | - | 4.90 | - | - | - | 0.89 | - | - | 18.57 | - | - | - | 6.00 | - |
| Euphorbia characias L. | 1.67 | Q-I | E3 | 14.75 | 0.70 | - | - | - | 2.68 | - | 12.00 | 25.00 | - | - | - | 10.00 | - |
| Fraxinus angustifolia subsp. angustifolia Vahl | 1.66 | S-P | E1 | 4.92 | 3.50 | - | - | - | 8.93 | 20.59 | 10.00 | 16.00 | - | - | - | 7.20 | 12.29 |
| Bunium macuca subsp. macuca Boiss. | 1.64 | Q-F | E3 | - | - | - | - | - | 3.57 | - | - | - | - | - | - | 17.00 | - |
| Sanicula europaea L. | 1.63 | Q-F | E3 | - | - | 16.67 | - | 11.63 | - | 5.88 | - | - | 12.00 | - | 12.20 | - | 18.50 |
| Melica uniflora Retz. | 1.59 | Q-F | E3 | - | - | - | - | 6.98 | - | - | - | - | - | - | 16.33 | - | - |
| Viola sylvestris subsp. sylvestris Lam. | 1.51 | Q-F | E3 | - | - | 16.67 | - | - | 9.82 | 8.82 | - | - | 14.33 | - | - | 10.36 | 16.33 |
| Asplenium onopteris L. | 1.48 | Q-I | E3 | - | 15.38 | - | - | - | - | - | - | 8.59 | - | - | - | - | - |
| Taxus baccata L. | 1.46 | Q-F | E1 | 1.64 | - | 16.67 | - | 6.98 | 8.04 | 14.71 | 6.00 | - | 8.00 | - | 10.00 | 9.11 | 8.40 |
| Vincetoxicum nigrum (L.) Moench | 1.46 | Q-I | E3 | - | 13.29 | 5.56 | - | 11.63 | 7.14 | - | - | 7.63 | 6.00 | - | 7.20 | 6.00 | - |
| Conopodium pyrenaeum (Loisel.) Miégev. | 1.44 | Q-F | E3 | 8.20 | - | 11.11 | - | 11.63 | 7.14 | - | 8.40 | - | 6.00 | - | 9.60 | 9.75 | - |
| Paeonia coriacea Boiss. | 1.39 | Q-I | E3 | 24.59 | - | - | - | - | 2.68 | - | 9.20 | - | - | - | - | 8.00 | - |
| Lathyrus linifolius (Reichard) Bässler | 1.37 | Q-F | E3 | - | 0.70 | - | - | 27.91 | - | - | - | 6.00 | - | - | 11.75 | - | - |
| Lonicera implexa Aiton | 1.36 | Q-I | E2 | - | 6.99 | - | - | - | 6.25 | - | - | 7.22 | - | - | - | 11.29 | - |
| Genista tournefortii Spach | 1.32 | Q-I | E3 | - | 9.79 | - | - | 2.33 | - | - | - | 9.50 | - | - | 6.00 | - | - |
| Sanguisorba hybrida (L.) Font Quer | 1.30 | Q-I | E3 | - | 5.59 | - | - | 4.65 | - | - | - | 13.00 | - | - | 6.00 | - | - |
| Genista falcata Brot. | 1.28 | Q-F | E3 | - | 2.10 | - | - | 30.23 | - | - | - | 10.00 | - | - | 7.38 | - | - |
| Hepatica nobilis Schreb. | 1.22 | Q-F | E3 | - | - | - | - | 9.30 | - | 8.82 | - | - | - | - | 6.00 | - | 18.67 |
| Pyrus bourgaeana Decne. | 1.17 | Q-I | E2 | - | 4.20 | - | - | - | - | - | - | 11.00 | - | - | - | - | - |
| Asparagus aphyllus subsp. aphyllus L. | 1.16 | Q-I | E2 | - | 2.80 | - | - | - | - | - | - | 12.00 | - | - | - | - | - |
| Potentilla sterilis (L.) Garcke | 1.14 | Q-F | E3 | - | - | - | - | 6.98 | - | - | - | - | - | - | 12.00 | - | - |
| Veronica officinalis L. | 1.09 | Q-F | E3 | - | - | 11.11 | - | 6.98 | - | 8.82 | - | - | 9.00 | - | 6.00 | - | 10.00 |
| Bupleurum rigidum subsp. paniculatum (Brot.) H. Wolff. | 1.08 | Q-I | E3 | - | 4.20 | - | - | 2.33 | - | - | - | 8.00 | - | - | 12.00 | - | - |
| Dryopteris affinis subsp. affinis (Lowe) Fraser-Jenk. | 1.07 | Q-F | E3 | - | - | - | - | 11.63 | - | - | - | - | - | - | 9.60 | - | - |
| Ulmus glabra Huds. | 1.07 | Q-F | E1 | - | - | - | - | - | 2.68 | 8.82 | - | - | - | - | - | 8.00 | 10.00 |
| Euphorbia hyberna L. | 1.04 | Q-F | E3 | - | 0.70 | - | - | 4.65 | - | - | - | 12.00 | - | - | 9.00 | - | - |
| Carex pendula Huds. | 1.02 | S-P | E3 | - | - | - | - | - | 0.89 | 8.82 | - | - | - | - | - | 6.00 | 10.00 |
| Senecio lopezii Boiss. | 1.02 | Q-I | E3 | - | - | - | 40.00 | - | - | - | - | - | - | 9.00 | - | - | - |
| Ajuga x_rotundifolia Willk. & Cutanda ex Willk. | 0.97 | Q-F | E3 | - | - | - | - | 4.65 | - | - | - | - | - | - | 9.00 | - | - |
| Carex sylvatica Huds. | 0.95 | Q-F | E3 | - | - | - | - | - | - | 11.76 | - | - | - | - | - | - | 7.50 |
| Moehringia pentandra J. Gay | 0.92 | Q-I | E4 | - | 6.99 | - | - | 11.63 | - | - | - | 5.70 | - | - | 6.00 | - | - |
| Asparagus acutifolius L. | 0.91 | Q-I | E2 | - | 9.09 | - | - | - | 4.46 | - | - | 6.69 | - | - | - | 13.40 | - |
| Phillyrea latifolia subsp. latifolia L. | 0.86 | Q-I | E2 | - | 4.20 | - | - | - | 2.68 | 8.82 | - | 3.55 | - | - | - | 10.00 | 8.00 |
| Juniperus phoenicea subsp. phoenicea L. | 0.78 | Q-I | E2 | - | - | - | - | - | 6.25 | 2.94 | - | - | - | - | - | 6.86 | 6.00 |
| Clematis campaniflora Brot. | 0.72 | S-P | E2 | - | 3.50 | - | - | - | - | - | - | 7.20 | - | - | - | - | - |
| Epipactis microphylla (Ehrh.) Sw. | 0.66 | Q-F | E3 | - | - | - | - | - | 0.89 | 11.76 | - | - | - | - | - | 6.00 | 6.00 |
| Anemone palmata L. | 0.63 | Q-I | E3 | - | 2.80 | - | - | - | - | - | - | 6.00 | - | - | - | - | - |
| Viola suavis M. Bieb. | 0.63 | Q-F | E3 | - | - | - | - | - | 3.57 | - | - | - | - | - | - | 6.00 | - |
| Iris foetidissima L. | 0.61 | S-P | E3 | - | - | - | - | - | - | 8.82 | - | - | - | - | - | - | 6.00 |
References
- Piñar Fuentes, J.C.; Cano-Ortiz, A.; Musarella, C.; Pinto-Gomes, C.; Spampinato, G.; Cano, E. Rupicolous habitats of interest for conservation in the central-southern Iberian peninsula. Plant. Sociol. 2017, 54, 29–42. [Google Scholar]
- Molero, J.; Marfil, J.M. The bioclimates of Sierra Nevada National Park. Int. J. Geobot. Res. 2015, 5, 1–11. [Google Scholar]
- Rivas-Martínez, S. Bioclimatoloy of the Iberian Peninsula. Itinera Geobot. 1999, 13, 41–47. [Google Scholar]
- Cano, E.; Cano-Ortiz, A.; Musarella, C.M.; Piñar Fuentes, J.C.; Ighbareyeh, J.M.H.; Gea, F.L.; del Río, S. Mitigating Climate Change Through Bioclimatic Applications and Cultivation Techniques in Agriculture (Andalusia, Spain). In Sustainable Agriculture, Forest and Environmental Management; Springer: Singapore, 2019; pp. 31–69. [Google Scholar]
- Rivas-Martínez, S.; Rivas-Saenz, S.; Penas, Á. Worldwide bioclimatic classification system. Glob. Geobot. 2011, 1, 1–634. [Google Scholar]
- Valladares, F.; Pearcy, R.W. Drought can be more critical in the shade than in the sun: A field study of carbon gain and photo-inhibition in a Californian shrub during a dry El Niño year. Plant Cell Environ. 2002, 25, 749–759. [Google Scholar] [CrossRef]
- Stanhill, G.; Cohen, S. Global dimming: A review of the evidence for a widespread and significant reduction in global radiation with discussion of its probable causes and possible agricultural consequences. Agr. For. Meteorol. 2001, 107, 255–278. [Google Scholar] [CrossRef]
- Valladares, F. Global change and radiation in Mediterranean forest ecosystems: A meeting point for ecology and management. In Ecology, Conservation and Sustainable Management of Mediterranean Type Ecosystems of the World; Millpress: Rotterdam, The Netherlands, 2004; pp. 1–4. [Google Scholar]
- Gómez-Aparicio, L.; Gómez, J.M.; Zamora, R. Microhabitats shift rank in suitability for seedling establishment depending on habitat type and climate. J. Ecol. 2005, 93, 1194–1202. [Google Scholar] [CrossRef]
- Kitzberger, T.; Steinaker, D.; Veblen, T. Effects of climatic variability on facilitation of tree establishment in northern Patagonia. Ecology 2000, 81, 1914–1921. [Google Scholar] [CrossRef]
- Schenk, H.J.; Mahal, B.E. Positive and negative plant interactions contribute to north–south–patterned association between two desert shrub species. Oecologia 2002, 132, 402–410. [Google Scholar] [CrossRef]
- Evans, R.; Young, J. Plant litter and establishment of alien annual weed species in rangeland communities. Weed Sci. 1970, 18, 697–703. [Google Scholar] [CrossRef]
- Fowler, N. What is a safe site? Neighbour, litter, germination date, and patch effects. Ecology 1988, 69, 947–961. [Google Scholar] [CrossRef]
- Verdú, M.; García-Fayos, P. Nucleation processes in a Mediterranean bird-dispersed plant. Funct. Ecol. 1996, 10, 275–280. [Google Scholar] [CrossRef]
- Callaway, R.N. Facilitation and interference of Quercus douglasii on understorey productivity in central California. Ecology 1991, 72, 1484–1499. [Google Scholar] [CrossRef]
- Joffre, R. How tree cover influences the water balance of Mediterranean rangelands. Ecology 1993, 74, 570–882. [Google Scholar] [CrossRef]
- Quinto-Canas, R.; Mendes, P.; Cano-Ortiz, A.; Musarella, C.M.; Pinto-Gomes, C. Comunidades de orla forestal en el suroeste de la península ibérica. Rev. Chapingo Ser. Cienc. For. Y Ambiente 2018, XXIV, 415–434. [Google Scholar]
- Gómez, J.M.; Gómez, L.; Zamora, R.; Montes, J. Problemas de regeneración de especies forestales autóctonas en el espacio natural protegido de Sierra Nevada. In Congreso Forestal Español; Sociedad Española de Ciencias Forestales: Palenciana, Spain, 2001. [Google Scholar]
- Aparicio, L.G. Papel de la heterogeneidad en la regeneración del Acer opalus subsp. granatense en la montaña mediterránea: Impicaciones para la conservación y restauración de sus poblaciones. Ecosistemas 2004, 13, 123–128. [Google Scholar]
- Cano, E.; Musarella, C.M.; Cano-Ortiz, A.; Piñar Fuentes, J.C.; Rodríguez Torres, A.; Del Río González, S.; Pinto Gomes, C.P.; Quinto-Canas, R.; Spampinato, G. Geobotanical Study of the Microforests of Juniperus oxycedrus subsp. badia in the Central and Southern Iberian Peninsula. Sustainability 2019, 11, 1111. [Google Scholar] [CrossRef]
- Valle, F.; Mota, J.F.; Gomez Mercado, F. Los Avellanares del Macizo Segura-Cazorla: Relaciones Ombroclima-Vegetación. En A Pulido, El Agua en Andalucía; Departamento de Hidrogeología: Granada, Spain, 1986; Volume 2, pp. 567–580. [Google Scholar]
- Vila-Viçosa, C.; Vázquez, F.M.; Mendes, P.; Del Rio, S.; Musarella, C.; Cano-Ortiz, A.; Meireles, C. Syntaxonomic update on the relict groves of Mirbeck’s oak (Quercus canariensis Willd. and Q. marianica C. Vicioso) in southern Iberia. Plant Biosyst. 2015, 149, 512–526. [Google Scholar] [CrossRef]
- Reyes, O.; Casal, M. Seed germination of Quercus robur, Q. pyrenaica and Q. ilex and the effects of smoke, heat, ash and charcoal. Ann. For. Sci. 2006, 63, 205–212. [Google Scholar] [CrossRef]
- Itziar, R.U.; Pérez-Ramos, I.M.; Zavala, M.A.; Marañón, T.; Kobe, R.K. Soil water content and emergence time control seedling establishment in three co-occurring Mediterranean oak species. Can. J. For. Res. 2008, 38, 2382–2393. [Google Scholar]
- Bioclimatic Map of Europe. Available online: http://www.globalbioclimatics.org/form/bi_med.htm (accessed on 15 July 2019).
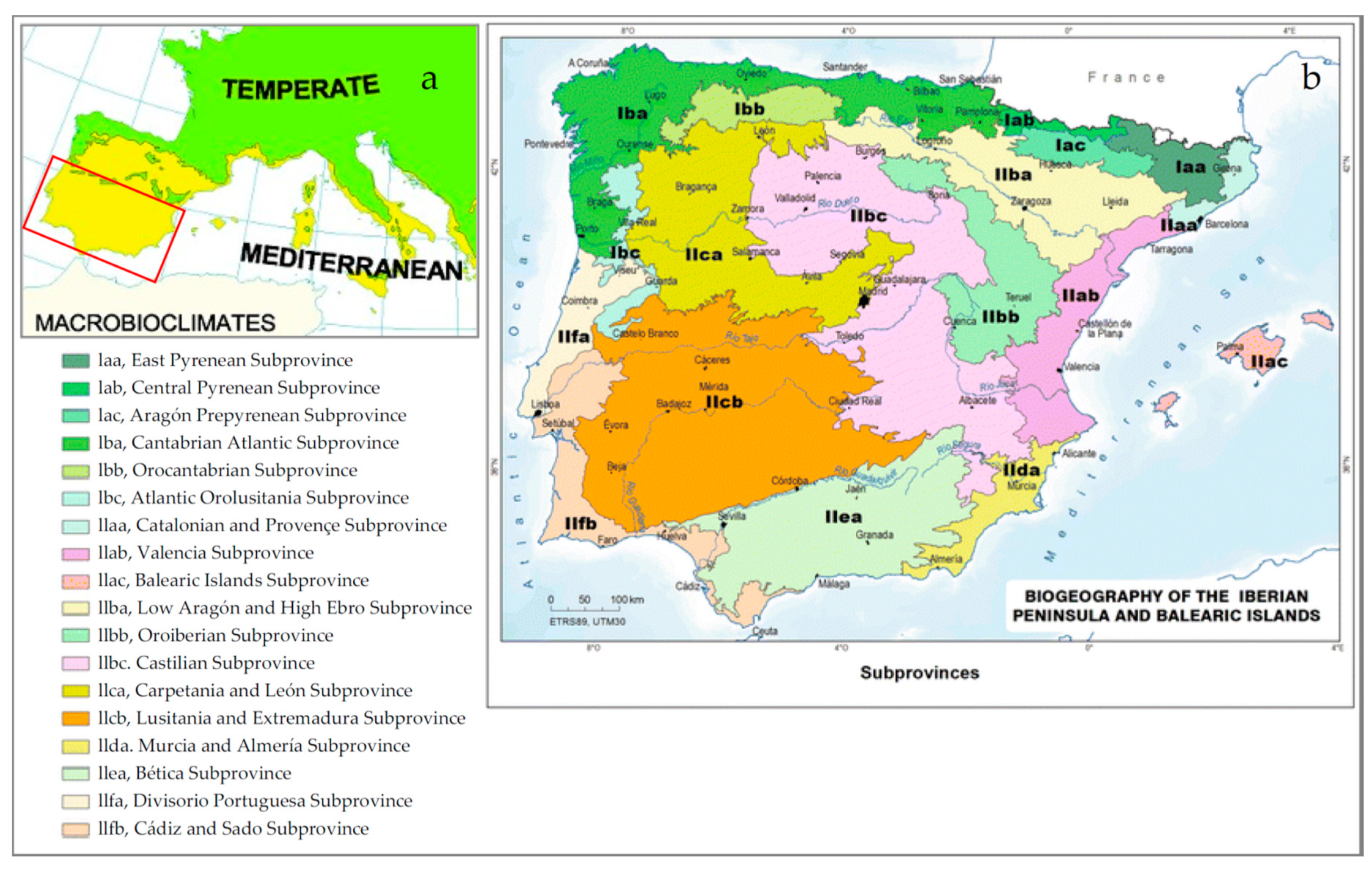
- Rivas-Martínez, S.; Penas, A.; Díaz González, T.E.; Cantó, P.; del Río, S.; Costa, J.C.; Herrero, L.; Molero Mesa, J. Biogeographic units of the iberian peninsula and baelaric islands to district level. A concise synopsis. In The Vegetation of the Iberian Peninsula Plant and Vegetation; Loidi, J., Ed.; Springer: Cham, Switzerland, 2017; Volume 12. [Google Scholar]
- Braun-Blanquet, J. Fitosociología: Bases Para el Estudio de Las Comunidades Vegetales; Ediciones Blume: Madrid, Spain, 1979. [Google Scholar]
- Martínez Parras, J.M.; Molero Mesa, J. Ecología y fitosociología de Quercus pyrenaica Willd. en la provincia Bética. Los melojares béticos y sus etapas de sustitución. Lazaroa 1982, 4, 91–104. [Google Scholar]
- Rivas Martinez, S. Memoria del Mapa de Las Series de Vegetación de España: 30 pp; ICONA Ministerio de Agricultura y Medio Ambiente: Madrid, Spain, 1987. [Google Scholar]
- Valle, F.; Mercado, F.G.; Poveda, J.F.M. Los robledales de la Sierra de Segura y otras comunidades relacionadas con ellos. Anal. Jardin. Bot. Madr. 1988, 45, 247–257. [Google Scholar]
- Rivas Martínez, S.; González, T.E.D.; González, F.F.; Arregui, J.J.L.; Lousã, M.F.; Merino, Á.P. Vascular plant communities of Spain and Portugal: Addenda to the syntaxonomical checklis of 2001. Part 1. Itinera Geobot. 2002, 15, 5–432. [Google Scholar]
- Rivas-Martínez, S. Esquema de la vegetación potencial y su correspondencia con los suelos en la España peninsular. Anal. Inst. Bot. Cavanilles 1964, 22, 341–405. [Google Scholar]
- Al Aallali, A.; Nieto, J.M.L.; Raya, F.A.P.; Mesa, J.M. Estudio de la vegetación forestal en la vertiente sur de Sierra Nevada (Alpujarra Alta granadina). Itinera Geobot. 1998, 11, 387–402. [Google Scholar]
- Losa, J.M.; Molero, J.; Casares, M.; Pérez-Raya, F. El paisaje vegetal de Sierra Nevada. La cuenca alta del río Genil. Granada. Serv. Publ. Univ. Granada 1986, 1, 1–285. [Google Scholar]
- Nieto, J.M.; Cabezudo, B. Series de vegetación climatófilas de las sierras Tejeda y Almijara (Málaga-Granada, España). Acta Bot. Malacit. 1988, 13, 229–260. [Google Scholar]
- Pérez Latorre, A.V.; Navas Fernández, D.; Gavira, O.; Caballero, G.; Cabezudo, B. Vegetación del Parque Natural de Las Sierras Tejeda, Almijara y Alhama (Málaga-Granada, España). Acta Bot. Malacit. 2004, 29, 117–190. [Google Scholar]
- Junta de Andalucía. Cartografía Y Evaluación de la Vegetación Y Flora de Los Ecosistemas Forestales de Andalucía a Escala de Detalle (1:10.000); Red de Información Ambiental de Andalucía (REDIAM): Andalucia, Spain, 1996–2006. [Google Scholar]
- Rivas Goday, S.; Borja Carbonell, J.; Esteve Chueca, F.; Fernández-Galiano, E.; Rigual Magallon, A.; Rivas Martínez, S. Contribución al estudio de la Quercetea ilicis hispánica. Anal. Inst. Bot. Cavanilles 1959, 17, 285–403. [Google Scholar]
- Cano Carmona, E.; Valle, F. Formaciones boscosas en Sierra Morena oriental. Acta Bot. Malacit. 1990, 15, 231–237. [Google Scholar]
- Melendo Luque, M. Cartografía Y Ordenación vegetal de Sierra Morena: Parque Natural de Las Sierras de Cardeña Y Montoro (Córdoba). Ph.D. Thesis, Universidad Jaén, Jaen, Spain, 1998. [Google Scholar]
- Vicente Orellana, J.A.; Galán De Mera, A. Nuevas aportaciones al Conocimiento de la vegetación Luso-Extremadurense. Estudio de las Sierras de Las Villuercas (Extremadura, España) y San Mamede (Alto Alentejo, Portugal). Acta Bot. Malacit. 2008, 33, 1–49. [Google Scholar]
- Amor, A. Flora Y Vegetación Vascular de la Comarca de la Vera Y Laderas Meridionales de la SIERRA de Tormantos (Cáceres). Ph.D. Thesis, Universidad de Salamanca, Salamanca, Spain, 1991. [Google Scholar]
- Belmonte, M.D. La Vegetación del Monfragüe y Su Área Socioeconómica de Influencia; Junta de Extremadura: Caceres, Spain, 2008. [Google Scholar]
- Monteiro-Henriques, T. Landscape and Phytosociology of the Paiva River’s Hidrographical Basin. Ph.D. Thesis, Universidade Técnica de Lisboa, Lisboa, Portuguesa, 2010. [Google Scholar]
- Pereira de Menezes Dias, M.C. A Flora e Vegetação da Serra de Monfurado (Alto Alentejo-Portugal). Guineana 2009, 15, 316. [Google Scholar]
- Sánchez-Pascual, N. Estudio fitosociológico y cartográfico de la comarca de Despeñaperros (Jaén). Ph.D. Thesis, Universidad de Granada, Granada, Spain, 1994. [Google Scholar]
- López Vélez, G. Flora y vegetación del macizo del Calar del Mundo y sierras adyacentes del sur de Albacete; Instituto de Estudios Albacetenses: Albacete, Spain, 1996; p. 520. [Google Scholar]
- Herranz, J.M.; Gómez Del Pozo, E.C. Contribución al conocimiento de la flora y vegetación de la comarca de Alcaraz; Caja de Ahorros de Albacete: Albacete, Spain, 1986; p. 279. [Google Scholar]
- Galán de Mera, A. Flora y Vegetación de los términos municipales de Alcalá de los Gazules y Medina Sidonia (Cádiz, España). Ph.D. Thesis, Universidad Complutense de Madrid, Madrid, Spain, 1993. [Google Scholar]
- Pérez Latorre, A.V.; Galán de Mera, A.; Deil, U.; Cabezudo, B. Fitogeografía y vegetación del sector aljíbico (Cádiz-Málaga, España). Acta Bot. Malacit. 1996, 21, 241–267. [Google Scholar]
- Asensi, A.; Díaz Garreta, B. The vegetation of Los Alcornocales Natural Park: Ronda-Faro (16 July). Itinera Geobot. 1999, 13, 129–135. [Google Scholar]
- Rivas Goday, S. Vegetación y flórula de la cuenca extremeña del Guadiana; Publicaciones de la Excma: Madrid, Spain, 1964; p. 777. [Google Scholar]
- Cantó, P. Estudio fitosociológico y biogeográfico de la sierra de San Vicente y tramo inferior del valle del Alberche (Toledo, España). Lazaroa 2004, 25, 187–250. [Google Scholar]
- Cabezudo, B.; Pérez-Latorre, A.; Navas, P.; Gil, Y.; Navas, D. Parque natural de la Sierra de las Nieves. Cartografía y evaluación de la flora y vegetación. Memoria; Departamento de Biología Vegetal, Universidad de Málaga: Málaga, Spain, 1998; p. 367. [Google Scholar]
- Giménez Luque, E. Bases Botánico-Ecológicas Para la Restauración de la Cubierta Vegetal de la Sierra de Gádor (Almería). Ph.D. Thesis, Universidad de Almería, Almería, Spain, 2004. [Google Scholar]
- Gómez Mercado, F.; Valle Tendero, F. Mapa de Vegetación de La Sierra de Baza; Servicio de Publicaciones: Madrid, Spain, 1988. [Google Scholar]
- Mercado, F.G. Vegetación y flora de la Sierra de Cazorla. Guineana 2011, 17, 1–481. [Google Scholar]
- Gómez Mercado, F.; Mota, J.F.; Peñas, J.; Cabello, J.; Valle, F. Vegetación. In Reconocimiento Biofísico de Espacios Naturales Protegidos. Parque Natural Sierras Subbéticas; Moreira, Ed.; Consejería de Medio Ambiente, Junta de Andalucía: Sevilla, Spain, 2000; pp. 195–311. [Google Scholar]
- Madrona, M.T. Cartografía de la vegetación actual y planificación de la restauración vegetal en las sierras de Lújar y la Contraviesa. Ph.D. Thesis, Universidad de Granada, Granada, Spain, 1994. [Google Scholar]
- Pavón Nuñez, M.; Pérez Latorre, A.V. Sobre las avellanedas subbéticas con Ulmus glabra Huds. en las sierras de Cazorla-Segura-Alcaráz, Jaen-Albacete (España). Lagascalia 2010, 30, 393–405. [Google Scholar]
- Pérez-Latorre, A.V.; Casimiro-Soriguer, F.; Gavira, O.; Cabezudo, B. Vegetación de la reserva de la biosfera Sierra de las Nieves: Río Grande y Sierras Prieta y Blanquilla (Málaga, España). Acta Bot. Malacit. 2012, 37, 103–140. [Google Scholar] [CrossRef]
- Inocencio, C.; Alcaraz, F.J.; Ríos, S. El paisaje vegetal de la Cuenca albacetense del Guadalmena. Inst. Estud. Albacet. Excma. Diput. Albacete Ser. I 1998, 100, 327. [Google Scholar]
- Olmedo, J.A. Análisis Biogeográfico y Cartografía de la Vegetación de la Sierra de Baza (Provincia de Granada). Ph.D. Thesis, Universidad de Granada, Granada, Spain, 2011. [Google Scholar]
- Gómez Navarro, J. Aportaciones al estudio de la flora y vegetación del extremo NE de la provincia de Albacete y zonas adyacentes de la provincia de Valencia (España). Fac. Ciènc. Biol. Univ. València. 2008, 2, 457–927. [Google Scholar]
- Molina Cantos, R.; Valdés Franzi, A.; Alcaraz Ariza, F.J. Flora y Vegetación del Tramo Medio del Valle del Río Júcar (Albacete); Instituto de Estudios Albacetenses ‘Don Juan Manuel’ de la Excma; Diputación de Albacete: Albacete, Spain, 2008; p. 663. [Google Scholar]
- Pavón Núñez, M.; Pérez Latorre, A.V.; Hidalgo Triana, N. Novedades fitosociológicas y florísticas en el Parque Natural de Tejeda-Almijara y Alhama (Málaga-Granada, España). Acta Bot. Malacit. 2015, 40, 199–205. [Google Scholar] [CrossRef][Green Version]
- Ríos Ruiz, S.; Alcaraz Ariza, F.; Valdés Franzi, A. Vegetación de Sotos Y Riberas de la Provincia de Albacete; Instituto de Estudios Albacetenses ‘Don Juan Manuel’: Albacetenses, Spain, 2003; p. 365. [Google Scholar]
- Vlieger, J. Aperçu sur les unités phytosociologiques supérieures des Pays-Bas. Ned. Kruidkd. Archief Ser. 3 1937, 47, 335–353. [Google Scholar]
- Bolòs, O.; de Bolòs Vayreda, A. Vegetación de Las Comarcas Barcelonesas: Descripción Geobotánica Y Catálogo Florístico Según Estudios Efectuados Por el Propio Autor y Por Oriol de Bolós Y Capdevilla; Instituto Español de Estudios Mediterráneos: Barcelona, Spain, 1950; p. 619. [Google Scholar]
- Rivas-Martínez, S.; Fernández-González, F.; Loidi, J.; Lousã, M.; Penas, A. Syntaxonomical checklist of vascular plant communities of Spain and Portugal to association level. Itinera Geobot. 2001, 14, 341. [Google Scholar]
- Rivas-Martínez, S. Vascular plant communities of Spain and Portugal (addenda to the syntaxonomical checklist of 2001, part II). Itinera Geobot. 2002, 15, 433–922. [Google Scholar]
- Del Pozo, A.; Brunel-Saldias, N.; Engler, A.; Ortega-Farias, S.; Acevedo-Opazo, C.; Lobos, G.A.; Jara-Rojas, R.; Molina-Montenegro, M.A. Climate Change Impacts and Adaptation Strategies of Agriculture in Mediterranean-Climate Regions (MCRs). Sustainability 2019, 11, 2769. [Google Scholar] [CrossRef]
- Del Río González, S. El cambio climático y su influencia en la vegetación de Castilla y León (España). Itinera Geobot. 2005, 16, 5–534. [Google Scholar]
- Del Río, S.; Penas, A.; Perez-Romero, R. Potential areas of deciduous forests in Spain (Castile and Leon) according to future climate change. Plant Biosyst. 2005, 139, 222–233. [Google Scholar] [CrossRef]
- Del Río, S.; Álvarez-Esteban, R.; Cano, E.; Pinto-Gomes, C.; Penas, Á. Potential impacts of climate change on habitat suitability of Fagus sylvatica L. forests in spain. Plant Biosyst. 2018, 152, 1205–1213. [Google Scholar] [CrossRef]
- Cano Ortiz, A.; Piñar Fuentes, J.C.; Pinto Gomes, C.J.; Musarella, C.M.; Cano, E. Expansion of the Juniperus Genus due to Anthropic Activity. In Old-Growth Forests and Coniferous Forests: Ecology, Habitat and Conservation; Serie Environmental Research Advances; Weber, R.P., Ed.; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2015; pp. 55–65. [Google Scholar]
- Torres, J.A.; Valle, F.; Pinto, C.; García-Fuentes, A.; Salazar, C.; Cano, E. Arbutus unedo L. communities in southern Iberian Peninsula mountains. Plant Ecol. 2002, 160, 207–223. [Google Scholar] [CrossRef]
- Torres, J.A.; Cano Carmona, E.; García Fuentes, A. Aportaciones al estudio fitosociológico de las comunidades de boj (Buxus sempervirens) en el sector subbético (Andalucía, España). Acta Bot. Malacit. 1996, 21, 319–321. [Google Scholar]
- Rivas-Martínez, S. La vegetación de la clase Quercetea ilicis en España y Portugal. Anal. Inst. Bot. Cavanilles 1974, 31, 205–259. [Google Scholar]
- Cano, E.; Rodríguez-Torres, A.; Pinto Gomes, C.J.; García-Fuentes, A.; Torres, J.A.; Salazar, C.; Ruiz-Valenzuela, L.; Cano-Ortiz, A.; Montilla, R.J. Analysis of the Juniperus oxycedrus L. communities in the centre and south of the Iberian peninsula (Spain and Portugal). Acta Bot. Gallica 2007, 154, 79–99. [Google Scholar] [CrossRef]
- Pavón Núñez, M.; Hidalgo Triana, N.; Pérez Latorre, A.V. Aportaciones al conocimiento de las comunidades de Pistacia terenbinthus L. y de Acer monspessulanum L. en el Sur de la Península Ibérica. Lagascalia 2013, 33, 299–311. [Google Scholar]
- Linares, J.C.; Carreira, J.A. El pinsapo, abeto endémico andaluz. O, ¿ Qué hace un tipo como tú en un sitio como éste? Rev. Ecosistemas 2006, 15, 171–191. [Google Scholar]
- Carrión, J.S.; Errikarta, I.Y.; Walker, M.J.; Legaz, A.J.; Chaín§, C.; López, A. Glacial refugia of temperate, Mediterranean and Ibero-North African flora in south-eastern Spain: New evidence from cave pollen at two Neanderthal man sites. Glob. Ecol. Biogeogr. 2003, 12, 119. [Google Scholar] [CrossRef]
- Willis, K.; Niklas, K. The role of Quaternary environmental change in plant macroevolution: The exception or the rule? Philos. Trans. R. Soc. Lond. 2004, 359, 159–172. [Google Scholar] [CrossRef]
- Costa, J.C.; Neto, C.; Aguiar, C.; Capelo, J.; Espírito Santo, M.D.; Honrado, J.; Lousã, M. Vascular plant communities in Portugal (Continental, the Azores and Madeira). Glob. Geobot. 2012, 2, 1–180. [Google Scholar]
- Shorohova, E.; Kuuluvainen, T.; Kangur, A.; Jõgiste, K. Natural stand structures, disturbance regimes and successional dynamics in the Eurasian boreal forests: A review with special reference to Russian studies. Ann. For. Sci. 2009, 66, 1–20. [Google Scholar] [CrossRef]
- Strandberg, B.; Kristiansen, S.M.; Tybirk, K. Dynamic oak-scrub to forest succession: Effects of management on understorey vegetation, humus forms and soils. For. Ecol. Manag. 2005, 211, 318–328. [Google Scholar] [CrossRef]
- Cano-Ortiz, A.; Pinto-Gomes, C.; Quinto-Canas, R.; Vila-Viçosa, C.; Rodríguez-Torres, A.; Redondo, M.M. Current state of the Prunetalia spinosae communities in the centre and south of the Iberian Peninsula (Spain, Portugal). Acta Bot. Gallica 2012, 159, 211–221. [Google Scholar] [CrossRef]
- Castro, J.; Zamora, R.; Hódar, J.A. Restoring Quercus pyrenaica forests using pioneer shrubs as nurse plants. Appl. Veg. Sci. 2006, 9, 137–142. [Google Scholar] [CrossRef]
- Ajbilou, R.; Marañón, T.; Arroyo, J. Ecological and biogeographical analyses of Mediterranean forests of northern Morocco. Acta Oecol. 2006, 29, 104–113. [Google Scholar] [CrossRef]
- Aguirre, P.V. Estudio Integral de Los Bosques de Quercus Pyrenaica Willd. en la Península Ibérica: Características Bioclimáticas, Sintaxonómicas Y Especies Indicadoras Comprehensive Study of Quercus Pyrenaica Willd. Forests at Iberian Peninsula: Indicator Species, Bioclimatic, and Syntaxonomical. Ph.D. Thesis, Universidad Complutense de Madrid, Madrid, Spain, 2014. [Google Scholar]
- Aguilar Alba, M. Cambios y tendencias recientes en las precipitaciones de Andalucía. Cambio Clim. Andal. Evol. Y Consecuencias Medioambient. 2007, 99–116. [Google Scholar]
- Del Río, S.; Herrero, L.; Fraile, R.; Penas, A. Spatial distribution of recent rainfall trends in Spain (1961–2006). Int. J. Climatol. 2011, 31, 656–667. [Google Scholar]
- Del Río, S.; Cano-Ortiz, A.; Herrero, L.; Penas, A. Recent trends in mean maximum and minimum air temperatures over Spain (1961–2006). Theor. Appl. Clim. 2012, 109, 605–626. [Google Scholar] [CrossRef]
- Alexander, L.V.; Zhang, X.; Peterson, T.C.; Caesar, J.; Gleason, B.; Klein Tank, A.M.G.; Tagipour, A. Global observed changes in daily climate extremes of temperature and precipitation. J. Geophys. Res. Atmos. 2006, 111. [Google Scholar] [CrossRef]
- Shestakova, T.A.; Gutiérrez, E.; Kirdyanov, A.V.; Camarero, J.J.; Génova, M.; Knorre, A.A.; Voltas, J. Forests synchronize their growth in contrasting Eurasian regions in response to climate warming. Proc. Natl. Acad. Sci. USA 2016, 113, 662–667. [Google Scholar] [CrossRef]
- Linares, J.C.; Pazo Sarria, R.; Taïqui, L.; Camarero, J.J.; Ochoa, V.; Lechuga, V.; Seco, J.I.; Viñegla, B.; Sangüesa-Barreda, G.; Gilarte, P.; et al. Efectos de las tendencias climáticas y la degradación del hábitat sobre el decaimiento de los cedrales (Cedrus atlantica) del norte de Marruecos. Ecosistemas 2012, 21, 7–14. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Pachauri, R.K., Meyer, L.A., Eds.; IPCC: Geneva, Switzerland, 2014; p. 151. [Google Scholar]
- Loidi, J.; Biurrun, I.; Campos, J.A.; García-Baquero, G.; García-Mijangos, I.; Herrera, M. Climate change and its influence on plant species and terrestrial habitats. Int. J. Geobot. Res. 2012, 2, 13–19. [Google Scholar] [CrossRef]
- El Consejo, D.L.C.E. Directiva 92/43/CEE del Consejo, de 21 de mayo de 1992, relativa a la conservación de los hábitats naturales y de la fauna y flora silvestres. D. Of. Comunidades Eur. 1992, 7–50. [Google Scholar]
- Campos, J.A.; Liendo, D.; Horvat, V.; Villasante, J.; Biurrun, I.; García-Mijangos, I.; Herrera, M. Efectividad de la red Natura 2000 en la conservación de la flora y los hábitats amenazados. In Proceedings of the Conference: I Congreso de la Sociedad Española de Geobotánica. Conservación de Hábitats Naturales en Europa, Gijon, Spain, 8–11 July 2014. [Google Scholar]
- Nunes, L.J.; Meireles, C.I.; Pinto Gomes, C.J.; Almeida Ribeiro, N. The Evolution of Climate Changes in Portugal: Determination of Trend Series and Its Impact on Forest Development. Climate 2019, 7, 78. [Google Scholar] [CrossRef]
- Zhang, L.; Dawes, W.R.; Walker, G.R. Response of mean annual evapotranspiration to vegetation changes at catchment scale. Water Resour. Res. 2001, 37, 701–708. [Google Scholar] [CrossRef]
- Goulden, M.L.; Bales, R.C. Mountain runoff vulnerability to increased evapotranspiration with vegetation expansion. Proc. Natl. Acad. Sci. USA 2014, 111, 14071–14075. [Google Scholar] [CrossRef] [PubMed]
- Cooper, D.J.; Sanderson, J.S.; Stannard, D.I.; Groeneveld, D.P. Effects of long-term water table drawdown on evapotranspiration and vegetation in an arid region phreatophyte community. J. Hydrol. 2006, 325, 21–34. [Google Scholar] [CrossRef]
- Alcántara, J.M.; Rey, P.J.; Manzaneda, A.J. A model of plant community dynamics based on replacement networks. J. Veg. Sci. 2015, 26, 524–537. [Google Scholar] [CrossRef]




| Plant Community | Bibliography | Number of Relevés |
|---|---|---|
| AdQp | El Aallali et al. [33] | 12 |
| Losa et al. [34] | 1 | |
| Martínez Parras & Molero [28] | 15 | |
| Nieto & Cabezudo [35] | 3 | |
| Pérez Latorre et al. [36] | 5 | |
| Junta de Andalucía. [37] | 25 | |
| AuQp | Rivas Goday [38] | 14 |
| Cano E. & Valle F. [39] | 6 | |
| Melendo Luque [40] | 3 | |
| Vicente Orellana & Galán De Mera [41] | 11 | |
| Own unpublished relevés | 16 | |
| Amor, A.; [42] | 9 | |
| Belmonte M.D.; [43] | 12 | |
| Monteiro-Henriques, T.; [44] | 6 | |
| Pereira [45] | 10 | |
| Sánchez-Pascual, N. [46] | 8 | |
| Junta de Andalucía. [37] | 48 | |
| BhQp | Valle et al. [30] | 8 |
| López Vélez, G. [47] | 7 | |
| Herranz et al. [48] | 3 | |
| LbQp | Galán de Mera, A. [49] | 5 |
| Pérez Latorre et al. [50] | 4 | |
| Asensi, A.; Díaz Garreta, B.; [51] | 1 | |
| StQp | Rivas Goday, S. [52] | 13 |
| Cantó, Paloma. [53] | 20 | |
| Own unpublished relevés | 6 | |
| Sánchez-Pascual, N. [46] | 3 | |
| Junta de Andalucía. [37] | 1 | |
| DlAg | Cabezudo et al. [54] | 1 |
| Giménez Luque, Esther; [55] | 5 | |
| Gómez Mercado et Valle [56] | 5 | |
| Gómez-Mercado, F.; [57] | 22 | |
| Gómez Mercado et al. [58] | 4 | |
| Herranz et al. [49] | 14 | |
| López Vélez, G.; [48] | 10 | |
| Madrona, M.T.; [59] | 2 | |
| Pavón & Pérez Latorre [60] | 5 | |
| Pérez-Latorre et al. [61] | 1 | |
| Inocencio Pretel [62] | 1 | |
| Olmedo, J.A.; [63] | 3 | |
| Gómez Navarro, José; [64] | 3 | |
| Molina Cantos et al. [65] | 3 | |
| Pavón Núñez et al. [66] | 1 | |
| Junta de Andalucía. [37] | 32 | |
| GuCa | Gómez-Mercado, F.; [58] | 3 |
| López Vélez, G.; [47] | 8 | |
| Pavón & Pérez Latorre [60] | 5 | |
| Ríos Ruiz et al. [67] | 7 | |
| Valle et al. [21] | 8 | |
| Junta de Andalucía. [37] | 3 |
| Phytosociological Index | % of Average Cover |
|---|---|
| 5 | 85 |
| 4 | 65 |
| 3 | 45 |
| 2 | 25 |
| 1 | 12 |
| + | 6 |
| r | 3 |
| Ic Value | C Value |
|---|---|
| < = 8 | C = (Ic − 8) × 10 |
| 8 < Ic < = 18 | C = 0 |
| 18 < Ic < = 21 | C = (Ic − 18) × 5 |
| 21 < Ic < = 28 | C = (Ic − 21) × 15 |
| 28 < Ic < = 46 | C = (Ic − 28) × 25 |
| 46 < Ic < = 65 | C = (Ic − 46) × 60 |
| Axis | Tmax | Tmin | Tp | PEs | PE | Iar | Id | IH | Ioe | Ios1 | Ios2 | Ios3 | Ios4 | Ic | Io | Itc |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Axis1 | 0.60 | 0.55 | 0.63 | 0.56 | 0.65 | 0.44 | 0.21 | −0.44 | −0.44 | −0.36 | −0.30 | −0.22 | −0.33 | 0.01 | −0.50 | 0.63 |
| Axis2 | −0.36 | −0.57 | −0.46 | −0.43 | −0.43 | −0.37 | 0.15 | 0.37 | 0.37 | 0.54 | 0.56 | 0.58 | 0.55 | 0.29 | 0.42 | −0.45 |
| Species | Phytosociological Class | Stratum | IVI | Tmax | Tmin | Tp | PEs | PE | Iar | Id | IH | Ioe | Ios1 | Ios2 | Ios3 | Ios4 | Ic | Io | Itc |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Quercus pyrenaica Willd. | Q-F | E1 | 12.63 | 0.25 | 0.29 | 0.31 | 0.22 | 0.31 | 0.30 | −0.10 | −0.30 | −0.30 | −0.28 | −0.30 | −0.29 | −0.34 | −0.07 | −0.32 | 0.32 |
| Acer opalus subsp. granatense (Boiss.) Font Quer and Rothm. | Q-F | E2 | 6.31 | −0.38 | −0.42 | −0.45 | −0.37 | −0.45 | −0.13 | −0.09 | 0.13 | 0.13 | 0.27 | 0.24 | 0.19 | 0.20 | 0.11 | 0.19 | −0.45 |
| Corylus avellana L. | Q-F | E2 | 5.04 | −0.10 | −0.12 | −0.07 | −0.03 | −0.06 | −0.28 | 0.28 | 0.28 | 0.28 | 0.34 | 0.34 | 0.36 | 0.35 | 0.03 | 0.27 | −0.11 |
| Quercus faginea subsp. faginea Lam. | Q-F | E1 | 4.78 | 0.08 | 0.04 | 0.07 | 0.12 | 0.08 | −0.18 | 0.33 | 0.18 | 0.18 | 0.22 | 0.30 | 0.24 | 0.22 | 0.07 | 0.16 | 0.03 |
| Brachypodium sylvaticum (Huds.) P. Beauv. | S-P | E3 | 4.44 | 0.14 | 0.09 | 0.16 | 0.13 | 0.17 | −0.09 | 0.26 | 0.09 | 0.09 | 0.09 | 0.10 | 0.18 | 0.14 | 0.07 | 0.08 | 0.15 |
| Rubia peregrina subsp. peregrina L. | Q-I | E2 | 4.40 | 0.18 | 0.25 | 0.25 | 0.21 | 0.26 | −0.04 | 0.24 | 0.04 | 0.04 | 0.03 | 0.04 | 0.11 | 0.05 | −0.03 | 0.01 | 0.24 |
| Daphne laureola subsp. latifolia (Coss.) Rivas Mart. | Q-F | E3 | 4.24 | −0.25 | −0.33 | −0.29 | −0.23 | −0.29 | −0.42 | 0.19 | 0.42 | 0.42 | 0.38 | 0.41 | 0.37 | 0.40 | 0.22 | 0.43 | −0.32 |
| Helleborus foetidus L. | Q-I | E3 | 4.18 | −0.29 | −0.30 | −0.32 | −0.26 | −0.32 | −0.34 | 0.05 | 0.34 | 0.34 | 0.24 | 0.23 | 0.22 | 0.27 | 0.06 | 0.35 | −0.32 |
| Arbutus unedo L. | Q-F | E2 | 4.18 | 0.46 | 0.44 | 0.49 | 0.50 | 0.51 | 0.16 | 0.22 | −0.16 | −0.16 | −0.17 | −0.17 | −0.07 | −0.05 | 0.05 | −0.21 | 0.46 |
| Daphne gnidium L. | Q-I | E3 | 3.62 | 0.34 | 0.42 | 0.43 | 0.36 | 0.42 | 0.13 | 0.13 | −0.13 | −0.13 | −0.23 | −0.18 | −0.10 | −0.13 | −0.07 | −0.17 | 0.42 |
| Quercus faginea subsp. alpestris (Boiss.) Maire | Q-F | E1 | 3.60 | −0.13 | −0.14 | −0.11 | −0.10 | −0.11 | −0.24 | 0.07 | 0.24 | 0.24 | 0.20 | 0.22 | 0.20 | 0.23 | 0.04 | 0.24 | −0.13 |
| Paeonia broteri Boiss. & Reut. | Q-F | E3 | 3.52 | 0.23 | 0.01 | 0.13 | 0.27 | 0.17 | 0.18 | 0.21 | −0.18 | −0.18 | 0.10 | 0.17 | 0.18 | 0.07 | 0.31 | −0.19 | 0.08 |
| Tamus communis L. | Q-I | E2 | 3.51 | 0.29 | 0.33 | 0.37 | 0.33 | 0.38 | 0.06 | 0.22 | −0.06 | −0.06 | −0.12 | −0.06 | 0.01 | 0.00 | 0.00 | −0.11 | 0.34 |
| Hedera helix subsp. helix L. | Q-F | E2 | 3.25 | −0.09 | −0.11 | −0.08 | −0.03 | −0.06 | −0.28 | 0.24 | 0.28 | 0.28 | 0.29 | 0.31 | 0.31 | 0.31 | 0.06 | 0.27 | −0.10 |
| Quercus suber var. suber L. | Q-I | E1 | 3.23 | 0.35 | 0.49 | 0.50 | 0.37 | 0.48 | 0.11 | 0.13 | −0.11 | −0.11 | −0.23 | −0.31 | −0.20 | −0.18 | −0.18 | −0.17 | 0.49 |
| Myrtus communis L. | Q-I | E2 | 3.12 | 0.07 | 0.14 | 0.14 | 0.08 | 0.14 | 0.10 | 0.12 | −0.10 | −0.10 | −0.09 | −0.15 | −0.14 | −0.14 | −0.13 | −0.12 | 0.14 |
| Aristolochia paucinervis Pomel | S-P | E3 | 3.01 | 0.07 | 0.08 | 0.09 | 0.11 | 0.11 | 0.16 | −0.06 | −0.16 | −0.16 | −0.12 | −0.10 | −0.10 | −0.19 | 0.01 | −0.18 | 0.08 |
| Quercus broteroi (Cout.) Rivas Mart. | Q-I | E1 | 2.91 | 0.29 | 0.18 | 0.23 | 0.32 | 0.25 | 0.18 | 0.23 | −0.18 | −0.18 | −0.07 | −0.01 | 0.00 | −0.03 | 0.16 | −0.19 | 0.20 |
| Acer monspessulanum subsp. monspessulanum L. | Q-F | E2 | 2.89 | 0.20 | 0.10 | 0.14 | 0.22 | 0.16 | −0.03 | 0.17 | 0.03 | 0.03 | 0.02 | 0.11 | 0.10 | 0.12 | 0.14 | 0.01 | 0.12 |
| Primula acaulis (L.) L. | Q-F | E3 | 2.87 | −0.18 | −0.25 | −0.18 | −0.12 | −0.17 | −0.42 | 0.26 | 0.42 | 0.42 | 0.45 | 0.47 | 0.49 | 0.50 | 0.16 | 0.42 | −0.23 |
| Teucrium scorodonia subsp. scorodonia L. | Q-I | E3 | 2.83 | 0.27 | 0.24 | 0.32 | 0.25 | 0.34 | 0.16 | 0.20 | −0.16 | −0.16 | −0.09 | −0.08 | 0.08 | −0.02 | 0.06 | −0.19 | 0.32 |
| Sorbus aria (L.) Crantz | Q-F | E2 | 2.80 | −0.24 | −0.29 | −0.31 | −0.27 | −0.30 | −0.02 | −0.13 | 0.02 | 0.02 | 0.05 | 0.04 | 0.00 | 0.02 | 0.08 | 0.05 | −0.28 |
| Ilex aquifolium L. | Q-F | E2 | 2.76 | −0.11 | −0.19 | −0.11 | −0.10 | −0.10 | −0.29 | 0.23 | 0.29 | 0.29 | 0.37 | 0.38 | 0.40 | 0.40 | 0.09 | 0.30 | −0.13 |
| Luzula forsteri subsp. forsteri (Sm.) DC. in Lam. & DC. | Q-I | E3 | 2.72 | 0.12 | −0.04 | 0.06 | 0.10 | 0.10 | 0.25 | −0.06 | −0.25 | −0.25 | −0.04 | −0.02 | 0.07 | −0.09 | 0.21 | −0.25 | 0.05 |
| Erica arborea L. | Q-F | E2 | 2.71 | 0.34 | 0.21 | 0.30 | 0.29 | 0.32 | 0.06 | 0.21 | −0.06 | −0.06 | −0.06 | −0.03 | 0.08 | 0.07 | 0.10 | −0.09 | 0.29 |
| Doronicum plantagineum L. | Q-I | E3 | 2.67 | 0.03 | −0.03 | −0.04 | 0.01 | −0.03 | 0.18 | −0.06 | −0.18 | −0.18 | −0.20 | −0.13 | −0.15 | −0.23 | 0.07 | −0.17 | −0.03 |
| Quercus coccifera subsp. coccifera L. | Q-F | E2 | 2.61 | 0.07 | 0.25 | 0.24 | 0.11 | 0.22 | 0.27 | 0.17 | −0.27 | −0.27 | −0.04 | −0.07 | −0.13 | −0.23 | −0.21 | −0.28 | 0.23 |
| Pistacia terebinthus L. | S-P | E2 | 2.39 | 0.20 | 0.16 | 0.17 | 0.22 | 0.18 | 0.02 | 0.16 | −0.02 | −0.02 | 0.00 | 0.08 | 0.05 | 0.02 | 0.06 | −0.04 | 0.16 |
| Ruscus aculeatus L. | Q-I | E3 | 2.38 | 0.23 | 0.26 | 0.27 | 0.26 | 0.28 | 0.03 | 0.17 | −0.03 | −0.03 | −0.06 | −0.04 | 0.07 | 0.04 | −0.01 | −0.05 | 0.26 |
| Holcus mollis subsp. mollis L. | Q-F | E3 | 2.23 | 0.00 | 0.15 | 0.16 | −0.04 | 0.16 | 0.16 | 0.05 | −0.16 | −0.16 | 0.04 | −0.05 | −0.05 | −0.11 | −0.20 | −0.17 | 0.19 |
| Pistacia lentiscus L. | Q-I | E2 | 2.20 | 0.12 | 0.12 | 0.12 | 0.13 | 0.13 | 0.09 | 0.05 | −0.09 | −0.09 | −0.04 | 0.04 | −0.02 | −0.09 | 0.01 | −0.10 | 0.12 |
| Piptatherum paradoxum (L.) P. Beauv. | Q-I | E3 | 2.20 | −0.04 | −0.06 | −0.05 | −0.04 | −0.04 | −0.17 | 0.14 | 0.17 | 0.17 | 0.15 | 0.20 | 0.17 | 0.17 | 0.05 | 0.17 | −0.06 |
| Carex distachya Desf. | Q-I | E3 | 2.17 | 0.26 | 0.14 | 0.20 | 0.28 | 0.22 | 0.17 | 0.02 | −0.17 | −0.17 | −0.06 | −0.08 | 0.01 | −0.08 | 0.19 | −0.18 | 0.17 |
| Phillyrea angustifolia L. | Q-F | E2 | 2.13 | 0.41 | 0.34 | 0.38 | 0.43 | 0.40 | 0.13 | 0.21 | −0.13 | −0.13 | −0.17 | −0.15 | 0.01 | −0.05 | 0.11 | −0.17 | 0.36 |
| Olea europaea var. sylvestris (Mill.) Lehr | Q-I | E2 | 1.85 | 0.09 | 0.22 | 0.22 | 0.11 | 0.21 | 0.16 | 0.15 | −0.16 | −0.16 | −0.12 | −0.19 | −0.18 | −0.20 | −0.15 | −0.18 | 0.22 |
| Juniperus oxycedrus subsp. oxycedrus L. | S-P | E2 | 1.84 | 0.18 | 0.10 | 0.15 | 0.22 | 0.17 | 0.05 | 0.16 | −0.05 | −0.05 | 0.08 | 0.15 | 0.10 | 0.09 | 0.14 | −0.06 | 0.12 |
| Paeonia officinalis subsp. microcarpa (Boiss. & Reut.) Nyman | Q-I | E3 | 1.81 | −0.23 | −0.33 | −0.27 | −0.20 | −0.26 | −0.23 | 0.07 | 0.23 | 0.23 | 0.34 | 0.36 | 0.35 | 0.35 | 0.21 | 0.26 | −0.31 |
| Rhamnus alaternus subsp. alaternus L. | Q-I | E2 | 1.79 | 0.09 | 0.15 | 0.15 | 0.11 | 0.15 | 0.18 | 0.18 | −0.18 | −0.18 | 0.01 | 0.01 | −0.05 | −0.13 | −0.11 | −0.19 | 0.15 |
| Rosa sicula Tratt. | Q-I | E3 | 1.76 | −0.19 | −0.16 | −0.23 | −0.21 | −0.23 | −0.02 | −0.14 | 0.02 | 0.02 | −0.03 | −0.07 | −0.04 | −0.04 | −0.02 | 0.05 | −0.20 |
| Bunium macuca subsp. macuca Boiss. | S-P | E3 | 1.74 | −0.10 | −0.04 | −0.10 | −0.10 | −0.11 | 0.09 | −0.13 | −0.09 | −0.09 | −0.09 | −0.13 | −0.10 | −0.12 | −0.07 | −0.07 | −0.08 |
| Fraxinus angustifolia subsp. angustifolia Vahl | Q-I | E1 | 1.71 | 0.00 | 0.00 | −0.01 | 0.01 | 0.01 | −0.11 | 0.13 | 0.11 | 0.11 | 0.09 | 0.13 | 0.13 | 0.11 | 0.05 | 0.10 | −0.01 |
| Smilax aspera L. | S-P | E2 | 1.69 | 0.12 | 0.23 | 0.22 | 0.13 | 0.21 | 0.02 | 0.12 | −0.02 | −0.02 | −0.13 | −0.14 | −0.14 | −0.10 | −0.17 | −0.05 | 0.22 |
| Melica uniflora Retz. | Q-F | E3 | 1.65 | 0.15 | −0.14 | 0.02 | −0.15 | 0.04 | 0.05 | 0.15 | −0.05 | −0.05 | 0.15 | 0.14 | 0.15 | 0.10 | 0.15 | −0.03 | 0.12 |
| Viola sylvestris subsp. sylvestris Lam. | Q-F | E3 | 1.62 | −0.09 | −0.10 | −0.08 | −0.05 | −0.07 | −0.21 | 0.12 | 0.21 | 0.21 | 0.22 | 0.22 | 0.22 | 0.23 | 0.04 | 0.21 | −0.09 |
| Vincetoxicum nigrum (L.) Moench | S-P | E3 | 1.57 | 0.18 | 0.08 | 0.13 | 0.20 | 0.14 | 0.01 | 0.17 | −0.01 | −0.01 | 0.01 | 0.06 | 0.09 | 0.08 | 0.16 | −0.03 | 0.10 |
| Euphorbia characias L. | Q-F | E3 | 1.56 | −0.10 | −0.09 | −0.16 | −0.14 | −0.17 | 0.04 | −0.19 | −0.04 | −0.04 | −0.17 | −0.21 | −0.19 | −0.18 | −0.03 | −0.02 | −0.13 |
| Sanicula europaea L. | Q-F | E3 | 1.55 | 0.05 | −0.16 | −0.03 | −0.09 | −0.02 | −0.01 | 0.14 | 0.01 | 0.01 | 0.18 | 0.20 | 0.19 | 0.14 | 0.17 | 0.02 | 0.00 |
| Lathyrus linifolius (Reichard) Bässler | S-P | E3 | 1.44 | 0.01 | −0.11 | −0.06 | 0.01 | −0.02 | 0.10 | 0.00 | −0.10 | −0.10 | 0.14 | 0.12 | 0.14 | −0.01 | 0.12 | −0.10 | −0.06 |
| Asparagus acutifolius L. | Q-I | E3 | 1.42 | 0.18 | 0.20 | 0.20 | 0.18 | 0.20 | 0.03 | 0.10 | −0.03 | −0.03 | −0.14 | −0.09 | −0.04 | −0.06 | 0.00 | −0.05 | 0.20 |
| Asplenium onopteris L. | Q-I | E3 | 1.41 | 0.22 | 0.22 | 0.24 | 0.24 | 0.25 | −0.02 | 0.04 | 0.02 | 0.02 | −0.15 | −0.11 | 0.06 | 0.06 | 0.05 | −0.01 | 0.23 |
| Conopodium pyrenaeum (Loisel.) Miégev. | Q-I | E3 | 1.40 | −0.06 | −0.18 | −0.13 | −0.03 | −0.11 | −0.01 | −0.06 | 0.01 | 0.01 | 0.14 | 0.13 | 0.12 | 0.05 | 0.22 | 0.04 | −0.16 |
| Taxus baccata L. | Q-F | E1 | 1.39 | −0.05 | −0.16 | −0.09 | −0.10 | −0.09 | −0.08 | 0.02 | 0.08 | 0.08 | 0.16 | 0.19 | 0.17 | 0.13 | 0.10 | 0.11 | −0.08 |
| Sanguisorba hybrida (L.) Font Quer | Q-F | E3 | 1.38 | 0.06 | 0.17 | 0.18 | 0.08 | 0.17 | 0.03 | 0.12 | −0.03 | −0.03 | −0.13 | −0.15 | −0.09 | −0.09 | −0.11 | −0.05 | 0.18 |
| Paeonia coriacea Boiss. | Q-F | E3 | 1.35 | −0.07 | −0.04 | −0.10 | −0.10 | −0.11 | 0.08 | −0.17 | −0.08 | −0.08 | −0.21 | −0.20 | −0.25 | −0.22 | −0.04 | −0.06 | −0.08 |
| Lonicera implexa Aiton | Q-I | E2 | 1.32 | 0.15 | 0.17 | 0.16 | 0.16 | 0.16 | 0.12 | 0.10 | −0.12 | −0.12 | −0.10 | −0.04 | −0.08 | −0.13 | −0.07 | −0.14 | 0.16 |
| Hepatica nobilis Schreb. | Q-I | E3 | 1.29 | 0.04 | −0.14 | −0.02 | −0.13 | −0.01 | −0.03 | 0.16 | 0.03 | 0.03 | 0.18 | 0.18 | 0.19 | 0.15 | 0.06 | 0.04 | 0.03 |
| Genista tournefortii Spach | Q-I | E3 | 1.28 | 0.25 | 0.17 | 0.20 | 0.24 | 0.22 | 0.08 | 0.12 | −0.08 | −0.08 | −0.13 | −0.10 | 0.02 | −0.04 | 0.12 | −0.10 | 0.19 |
| Asparagus aphyllus subsp. aphyllus L. | Q-F | E3 | 1.27 | 0.04 | 0.16 | 0.16 | 0.06 | 0.16 | 0.14 | 0.10 | −0.14 | −0.14 | −0.10 | −0.17 | −0.16 | −0.16 | −0.16 | −0.15 | 0.16 |
| Potentilla sterilis (L.) Garcke | Q-F | E3 | 1.24 | 0.08 | 0.13 | 0.11 | 0.05 | 0.09 | −0.13 | −0.09 | 0.13 | 0.13 | −0.06 | −0.12 | −0.11 | 0.01 | −0.10 | 0.12 | 0.12 |
| Pyrus bourgaeana Decne. | Q-F | E2 | 1.21 | 0.13 | 0.16 | 0.16 | 0.14 | 0.16 | 0.10 | 0.09 | −0.10 | −0.10 | −0.09 | −0.08 | −0.05 | −0.11 | −0.05 | −0.11 | 0.16 |
| Genista falcata Brot. | Q-F | E3 | 1.18 | −0.06 | −0.11 | −0.07 | −0.02 | −0.05 | 0.12 | −0.08 | −0.12 | −0.12 | 0.10 | 0.12 | 0.14 | 0.03 | 0.07 | −0.12 | −0.10 |
| Dryopteris affinis subsp. affinis (Lowe) Fraser-Jenk. | Q-F | E3 | 1.06 | 0.03 | −0.13 | −0.06 | −0.13 | −0.05 | −0.03 | 0.11 | 0.03 | 0.03 | 0.12 | 0.14 | 0.15 | 0.11 | 0.04 | 0.04 | 0.00 |
| Ulmus glabra Huds. | Q-F | E1 | 1.03 | −0.04 | −0.08 | −0.06 | −0.03 | −0.05 | −0.10 | 0.07 | 0.10 | 0.10 | 0.13 | 0.16 | 0.14 | 0.13 | 0.08 | 0.10 | −0.07 |
| Euphorbia hyberna L. | Q-F | E3 | 1.03 | −0.09 | −0.05 | −0.07 | −0.08 | −0.06 | −0.14 | 0.01 | 0.14 | 0.14 | 0.04 | 0.05 | 0.11 | 0.14 | −0.10 | 0.14 | −0.06 |
| Veronica officinalis L. | Q-F | E3 | 1.02 | 0.02 | −0.16 | −0.05 | −0.14 | −0.04 | −0.08 | 0.16 | 0.08 | 0.08 | 0.21 | 0.20 | 0.21 | 0.18 | 0.10 | 0.09 | 0.00 |
| Bupleurum rigidum subsp. paniculatum (Brot.) H. Wolff. | Q-F | E3 | 1.01 | 0.13 | 0.11 | 0.11 | 0.11 | 0.11 | 0.02 | 0.14 | −0.02 | −0.02 | −0.09 | −0.03 | −0.03 | −0.05 | 0.00 | −0.03 | 0.11 |
| Senecio lopezii Boiss. | S-P | E3 | 0.98 | 0.10 | 0.15 | 0.13 | 0.07 | 0.11 | −0.12 | −0.11 | 0.12 | 0.12 | −0.06 | −0.13 | −0.13 | −0.03 | −0.12 | 0.10 | 0.14 |
| Carex pendula Huds. | Q-I | E3 | 0.98 | −0.05 | −0.06 | −0.05 | −0.05 | −0.05 | −0.11 | 0.11 | 0.11 | 0.11 | 0.10 | 0.11 | 0.12 | 0.12 | 0.02 | 0.11 | −0.06 |
| Moehringia pentandra J. Gay | S-P | E4 | 0.97 | 0.16 | 0.08 | 0.12 | 0.19 | 0.14 | 0.16 | 0.12 | −0.16 | −0.16 | −0.04 | 0.01 | 0.01 | −0.04 | 0.12 | −0.16 | 0.10 |
| Ajuga x_rotundifolia Willk. & Cutanda ex Willk. | S-P | E3 | 0.92 | 0.12 | −0.12 | 0.06 | −0.12 | 0.10 | 0.09 | 0.12 | −0.09 | −0.09 | 0.12 | 0.12 | 0.12 | 0.07 | 0.12 | −0.07 | 0.12 |
| Phillyrea latifolia subsp. latifolia L. | Q-F | E2 | 0.91 | 0.15 | 0.10 | 0.12 | 0.16 | 0.13 | 0.02 | 0.11 | −0.02 | −0.02 | −0.04 | 0.03 | 0.03 | 0.03 | 0.06 | −0.04 | 0.11 |
| Juniperus phoenicea subsp. phoenicea L. | Q-I | E2 | 0.86 | 0.05 | 0.13 | 0.11 | 0.06 | 0.10 | 0.14 | 0.06 | −0.14 | −0.14 | 0.07 | 0.07 | −0.02 | −0.10 | −0.11 | −0.14 | 0.11 |
| Clematis campaniflora Brot. | Q-I | E2 | 0.83 | 0.12 | 0.11 | 0.11 | 0.10 | 0.11 | 0.01 | 0.10 | −0.01 | −0.01 | −0.11 | −0.03 | −0.04 | −0.05 | −0.03 | −0.02 | 0.11 |
| Carex sylvatica Huds. | Q-F | E3 | 0.82 | −0.06 | −0.09 | −0.08 | −0.06 | −0.08 | −0.11 | 0.11 | 0.11 | 0.11 | 0.12 | 0.13 | 0.14 | 0.14 | 0.06 | 0.12 | −0.09 |
| Epipactis microphylla (Ehrh.) Sw. | Q-F | E3 | 0.71 | −0.05 | −0.09 | −0.07 | −0.04 | −0.06 | −0.11 | 0.10 | 0.11 | 0.11 | 0.13 | 0.15 | 0.14 | 0.14 | 0.08 | 0.11 | −0.08 |
| Anemone palmata L. | Q-I | E3 | 0.69 | 0.13 | 0.09 | 0.10 | 0.14 | 0.11 | 0.10 | 0.12 | −0.10 | −0.10 | −0.04 | −0.02 | −0.03 | −0.06 | 0.06 | −0.12 | 0.10 |
| Viola suavis M. Bieb. | Q-F | E3 | 0.69 | −0.06 | −0.12 | −0.09 | −0.05 | −0.09 | −0.09 | 0.02 | 0.09 | 0.09 | 0.13 | 0.15 | 0.13 | 0.12 | 0.13 | 0.10 | −0.11 |
| Iris foetidissima L. | S-P | E3 | 0.66 | −0.08 | −0.12 | −0.10 | −0.08 | −0.10 | −0.12 | 0.10 | 0.12 | 0.12 | 0.12 | 0.12 | 0.13 | 0.14 | 0.07 | 0.13 | −0.11 |
| Climate Variable | R² | F | Pr > F (p-Value) | Climate Variable | R² | F | Pr > F (p-Value) |
|---|---|---|---|---|---|---|---|
| Tmax | 0.0018 | 0.1569 | 0.6930 | Ioe | 0.1983 | 21.2673 | <0.0001 |
| Tmin | 0.5960 | 126.8766 | <0.0001 | Ios1 | 0.2591 | 30.0685 | <0.0001 |
| Tp | 0.5550 | 107.2388 | <0.0001 | Ios2 | 0.3395 | 44.1971 | <0.0001 |
| PEs | 0.4093 | 59.5860 | <0.0001 | Ios3 | 0.4922 | 83.3669 | <0.0001 |
| PE | 0.5089 | 89.1102 | <0.0001 | Ios4 | 0.4068 | 58.9869 | <0.0001 |
| Iar | 0.1842 | 19.4198 | <0.0001 | Ic | 0.0668 | 6.1594 | 0.0150 |
| Id | 0.0052 | 0.4524 | 0.5030 | Io | 0.2589 | 30.0473 | <0.0001 |
| IH | 0.1983 | 21.2673 | <0.0001 | Itc | 0.0329 | 2.9248 | 0.0908 |
| Phytosociological Class | AdQp | AuQp | BhQp | LbQp | StQp | DlAg | GuCa |
|---|---|---|---|---|---|---|---|
| Querco roboris-Fagetea sylvaticae | 54.84 | 49.41 | 69.23 | 27.78 | 60.92 | 40.98 | 60.42 |
| Quercetea ilicis | 32.26 | 40 | 23.08 | 66.67 | 26.44 | 40.98 | 22.92 |
| Salici purpureae-Populetea nigrae | 12.9 | 10.59 | 7.692 | 5.556 | 12.64 | 18.03 | 16.67 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piñar Fuentes, J.C.; Cano-Ortiz, A.; Musarella, C.M.; Quinto Canas, R.; Pinto Gomes, C.J.; Spampinato, G.; del Río, S.; Cano, E. Bioclimatology, Structure, and Conservation Perspectives of Quercus pyrenaica, Acer opalus subsp. Granatensis, and Corylus avellana Deciduous Forests on Mediterranean Bioclimate in the South-Central Part of the Iberian Peninsula. Sustainability 2019, 11, 6500. https://doi.org/10.3390/su11226500
Piñar Fuentes JC, Cano-Ortiz A, Musarella CM, Quinto Canas R, Pinto Gomes CJ, Spampinato G, del Río S, Cano E. Bioclimatology, Structure, and Conservation Perspectives of Quercus pyrenaica, Acer opalus subsp. Granatensis, and Corylus avellana Deciduous Forests on Mediterranean Bioclimate in the South-Central Part of the Iberian Peninsula. Sustainability. 2019; 11(22):6500. https://doi.org/10.3390/su11226500
Chicago/Turabian StylePiñar Fuentes, J. C., A. Cano-Ortiz, C. M. Musarella, R. Quinto Canas, C. J. Pinto Gomes, G. Spampinato, S. del Río, and E. Cano. 2019. "Bioclimatology, Structure, and Conservation Perspectives of Quercus pyrenaica, Acer opalus subsp. Granatensis, and Corylus avellana Deciduous Forests on Mediterranean Bioclimate in the South-Central Part of the Iberian Peninsula" Sustainability 11, no. 22: 6500. https://doi.org/10.3390/su11226500
APA StylePiñar Fuentes, J. C., Cano-Ortiz, A., Musarella, C. M., Quinto Canas, R., Pinto Gomes, C. J., Spampinato, G., del Río, S., & Cano, E. (2019). Bioclimatology, Structure, and Conservation Perspectives of Quercus pyrenaica, Acer opalus subsp. Granatensis, and Corylus avellana Deciduous Forests on Mediterranean Bioclimate in the South-Central Part of the Iberian Peninsula. Sustainability, 11(22), 6500. https://doi.org/10.3390/su11226500