Exploring Marine Environments for the Identification of Extremophiles and Their Enzymes for Sustainable and Green Bioprocesses


 ,
, 
 ,
, 
Abstract
1. Introduction
2. Extreme Ecosystems
2.1. Cold Environments
2.2. Hydrothermal Vent Habitats
2.3. Hypersaline Environments
3. Marine Extremozymes: Current and Potential Applications for Biofuels Production and Bioremediation Processes
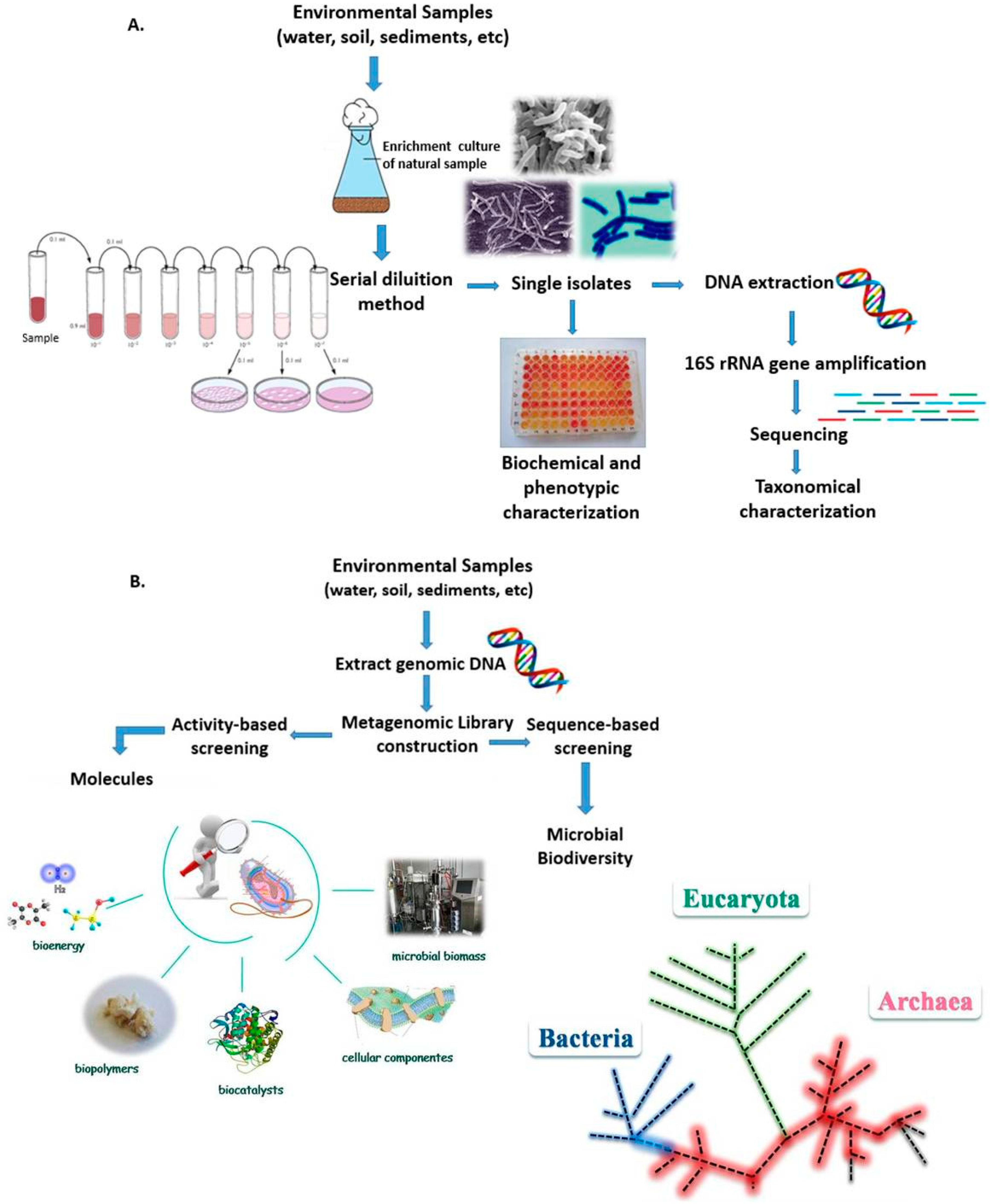
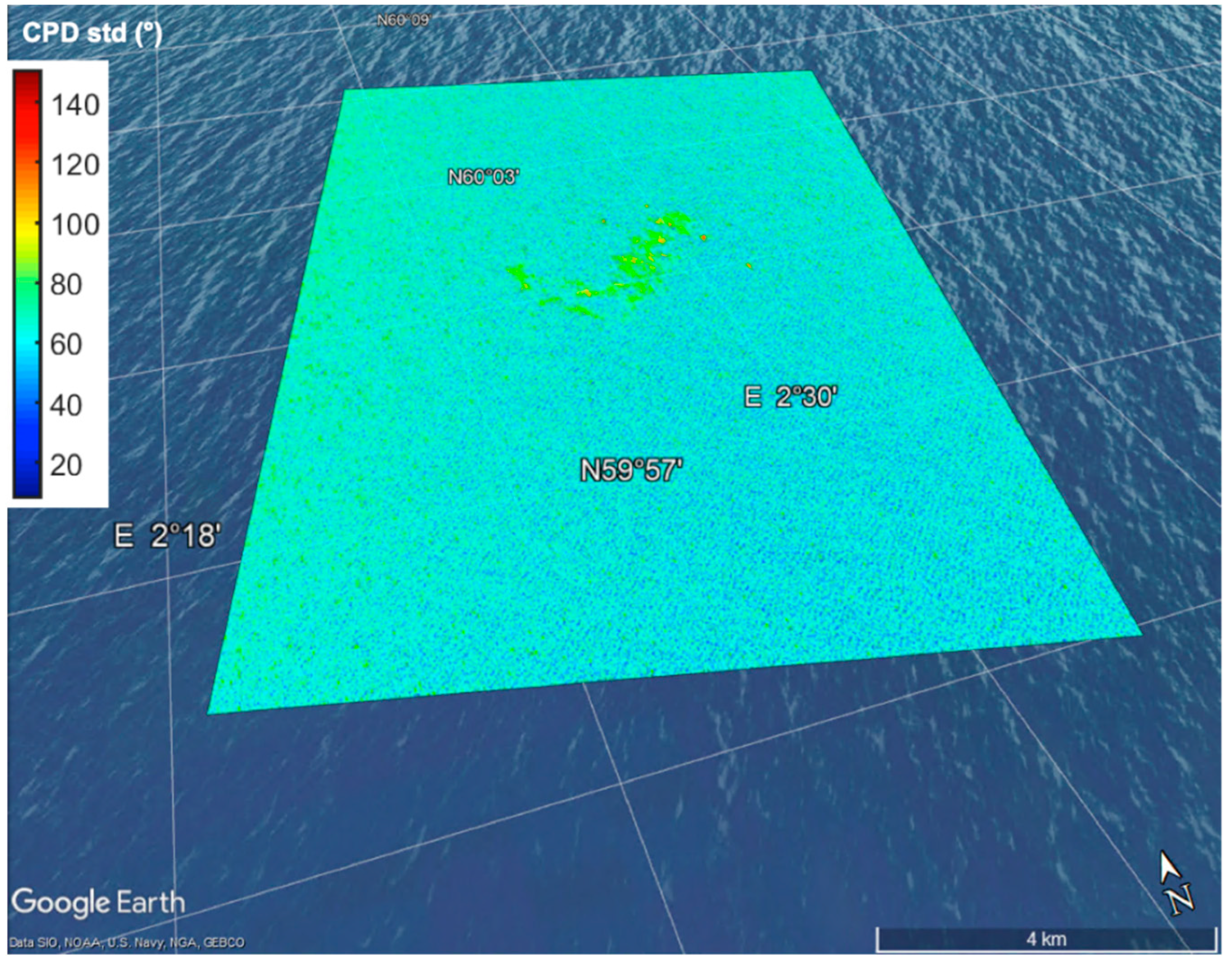
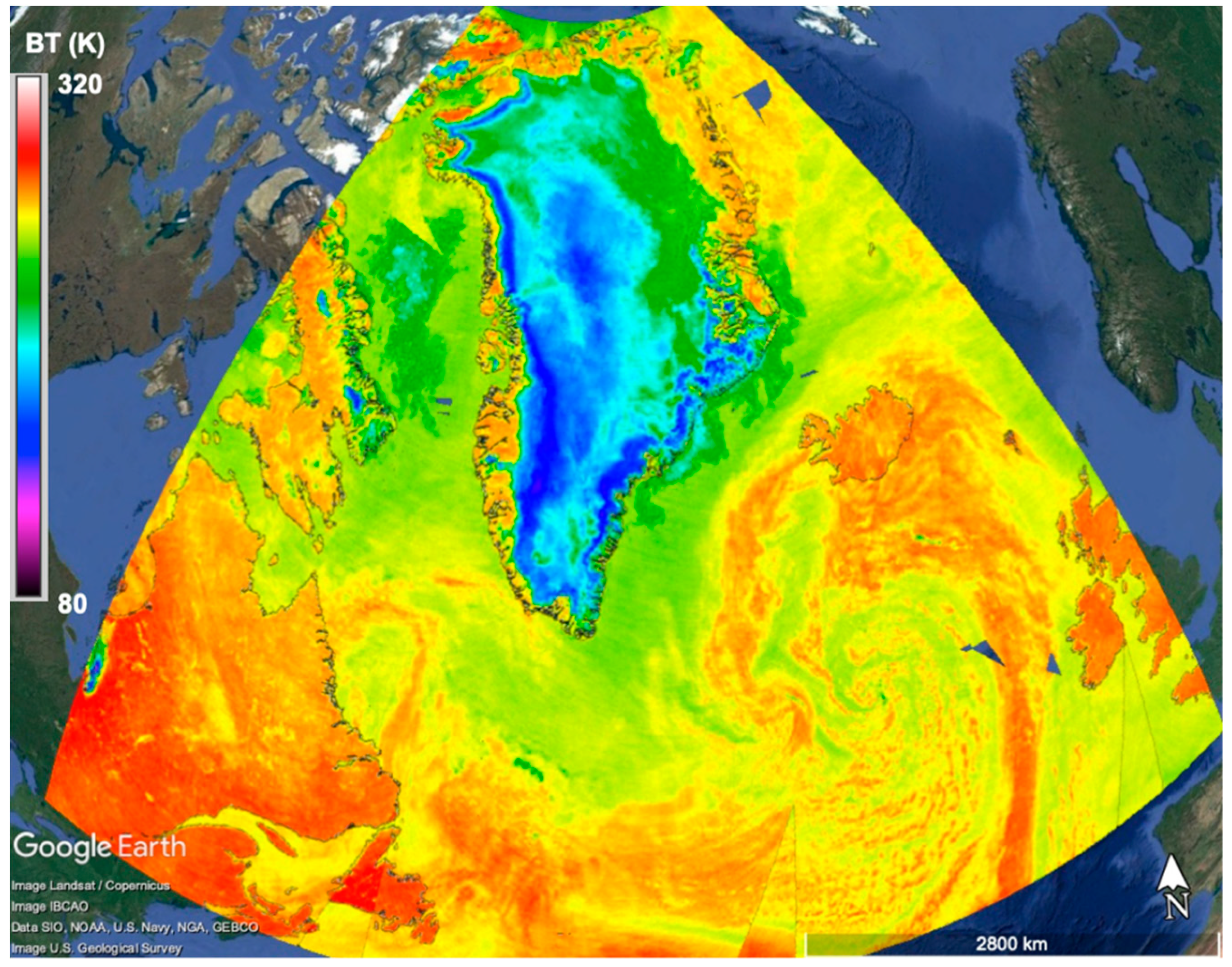
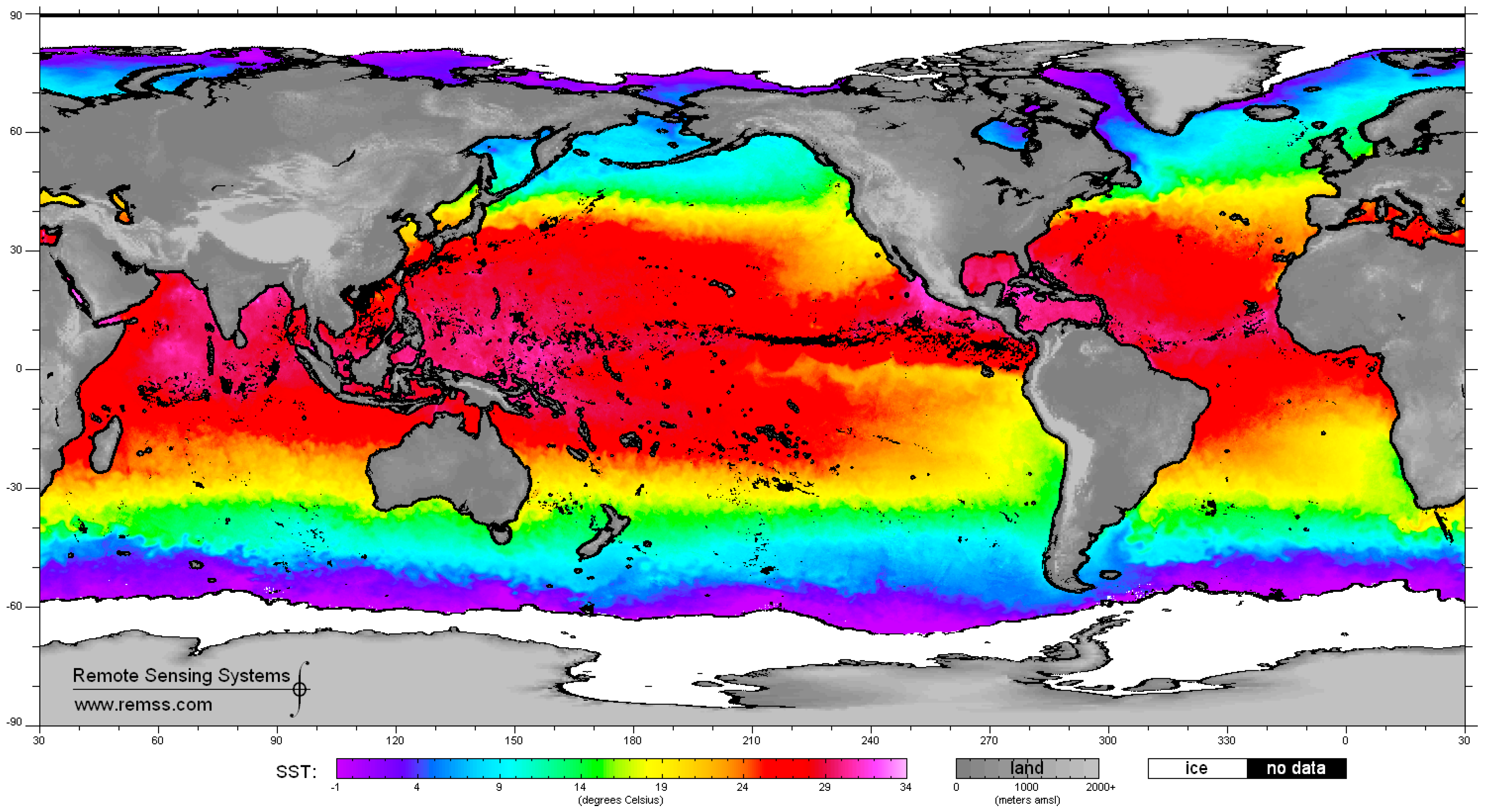
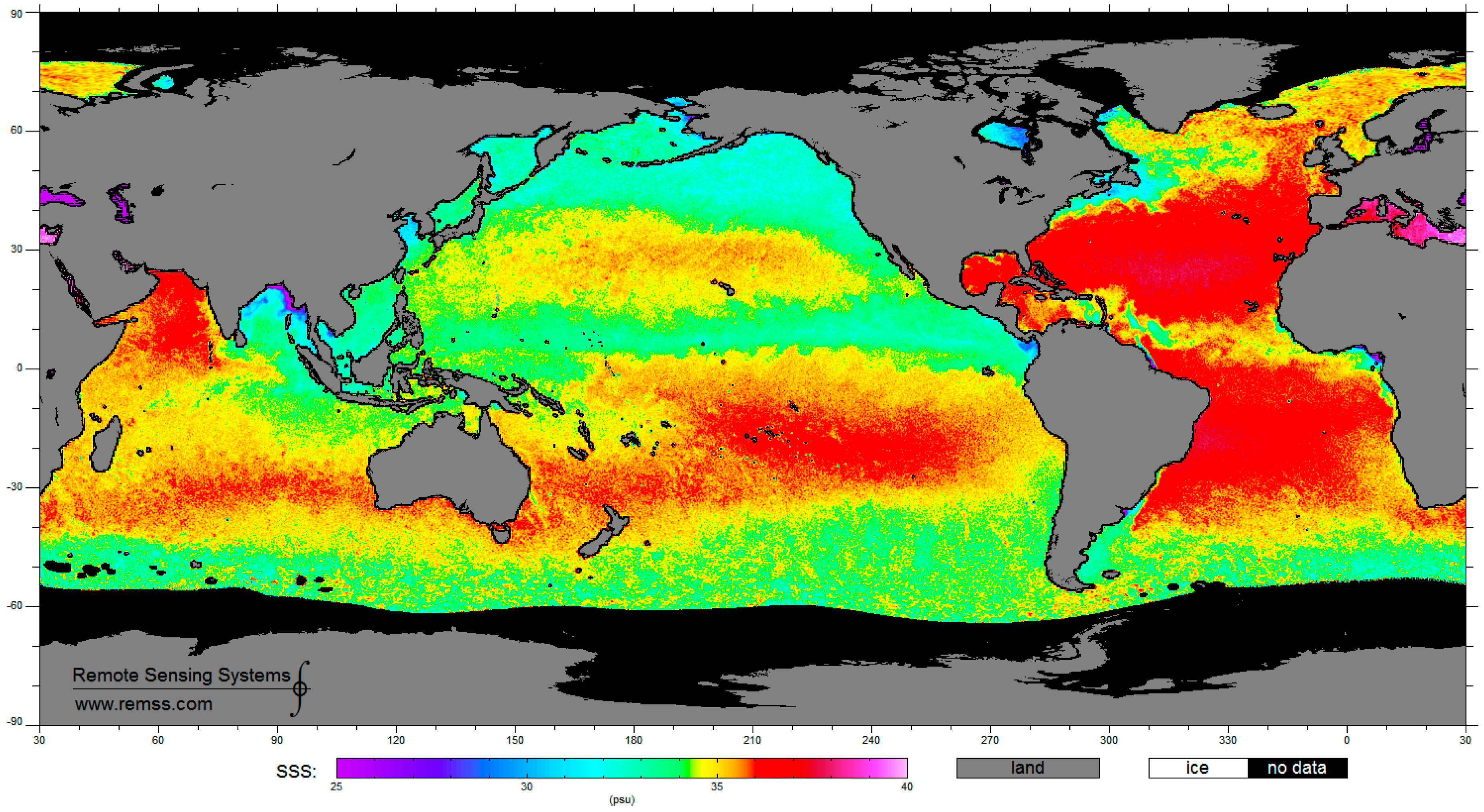
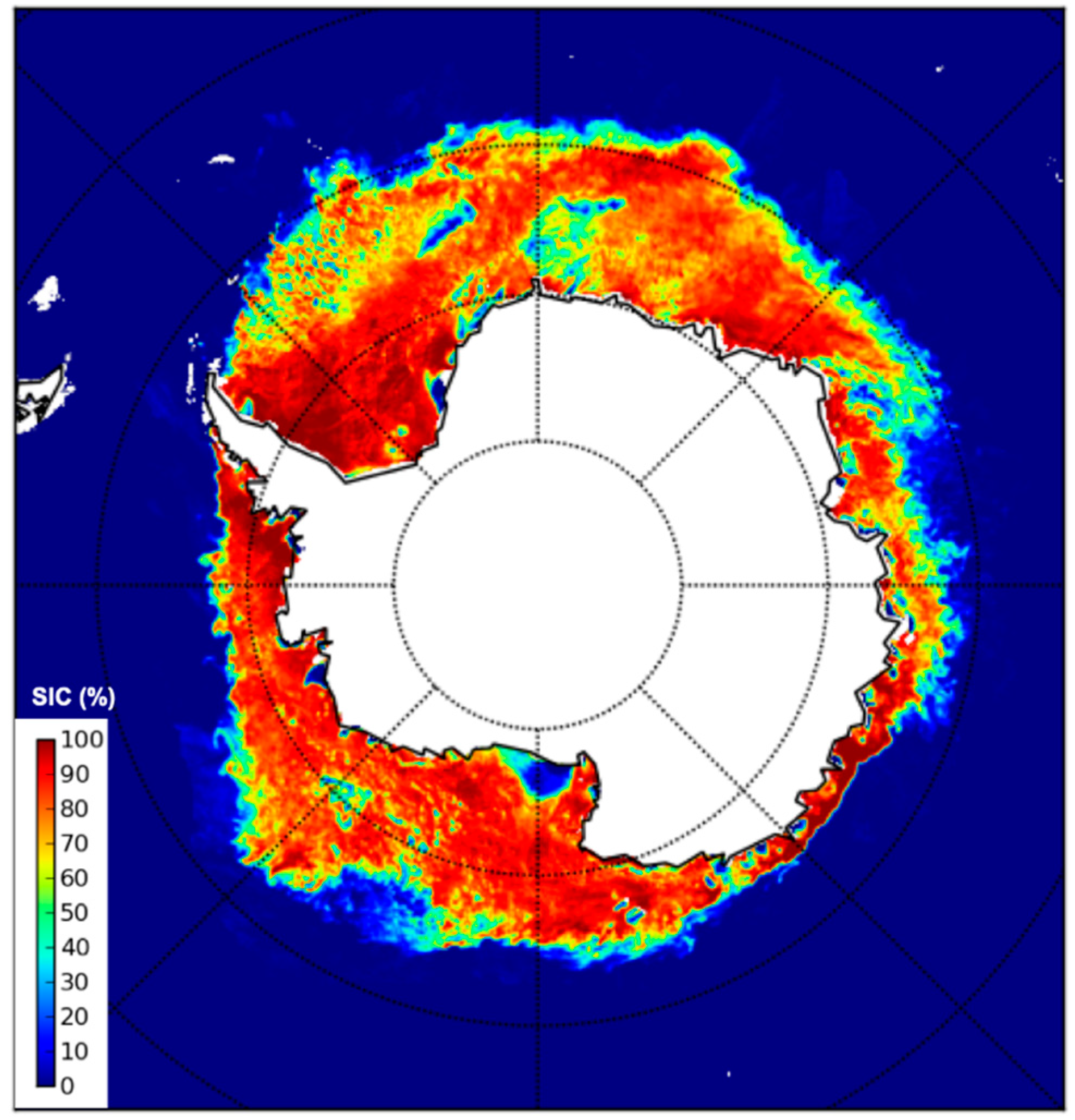
4. Satellite Microwave Remote Sensing to Support the Identification of Potential Sampling Sites
- Identification of oil-polluted sea surface areas by means of satellite polarimetric SARs.
- Identification of sea surface areas characterized by extreme temperature/salinity conditions by means of multi-frequency polarimetric radiometers.
- Identification of extreme ice-infested sea surface areas by means of single- and multi-frequency polarimetric radiometers.
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bolhuis, H.; Cretoiu, M.S. What is so Special about Marine Microorganisms? Introduction to the Marine Microbiome—From Diversity to Biotechnological Potential. In The Marine Microbiome an Untapped Source of Biodiversity and Biotechnological; Stal, L.J., Cretoiu, M.S., Eds.; Springer: Cham, Switzerland, 2016; Chapter 1; pp. 3–20. [Google Scholar]
- MacElroy, R.D. Some comments on evolution of extremophiles. Biosystems 1974, 6, 74–75. [Google Scholar] [CrossRef]
- Rothschild, L.J.; Mancinelli, R.L. Life in extreme environments. Nature 2001, 409, 1092–1101. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekaran, M.; Kumar, S.R. Marine microbial enzymes. In Biotechnology; Werner, H., Roken, S., Eds.; EOLSS: Paris, France, 2010; Volume 9, pp. 47–49. [Google Scholar]
- Russo, R.; Giordano, D.; Riccio, A.; di Prisco, G.; Verde, C. Cold-adapted bacteria and the globin case study in the Antarctic bacterium Pseudoalteromonas haloplanktis TAC125. Mar. Genom. 2010, 3, 125–131. [Google Scholar] [CrossRef]
- Trincone, A. Potential biocatalysts originating from sea environments. J. Mol. Catal. B-Enzym. 2010, 66, 241–256. [Google Scholar] [CrossRef]
- Fulzele, R.; Desa, E.; Yadav, A.; Shouche, Y.; Bhadekar, R. Characterization of novel extracellular protease produced by marine bacterial isolate from the Indian Ocean. Braz. J. Microbiol. 2011, 42, 1364–1373. [Google Scholar] [CrossRef] [PubMed]
- Samuel, P.; Raja, A.; Prabakaran, P. Investigation and application of marine derived microbial enzymes: Status and prospects. Int. J. Oceanogr. Mar. Ecol. Syst. 2012, 1, 1–10. [Google Scholar] [CrossRef]
- Poli, A.; Finore, I.; Romano, I.; Gioiello, A.; Lama, L.; Nicolaus, B. Microbial Diversity in Extreme Marine Habitats and Their Biomolecules. Microorganisms 2017, 5, 25. [Google Scholar] [CrossRef]
- Mapelli, F.; Crotti, E.; Molinari, F.; Daffonchio, D.; Borin, S. Extreme Marine Environments (Brines, Seeps, and Smokers). In the Marine Microbiome; Stal, L.J., Cretoiu, M.S., Eds.; Springer: Cham, Switzerland, 2016; Chapter 9; pp. 251–282. ISBN 978-3-319-32998-7, 978-3-319-33000-6. [Google Scholar]
- Moyer, C.L.; Morita, R.Y. Psychrophiles and Psychrotrophs. In Encyclopedia of Life Sciences; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2007. [Google Scholar]
- Caruso, C.; Rizzo, C.; Mangano, S.; Rappazzo, A.C.; Poli, A.; Di Donato, P.; Nicolaus, B.; Di Marco, G.; Michaud, L.; Lo Giudice, A. Extracellular polymeric substances with metal adsorption capacity produced by Pseudoalteromonas sp. MER144 from Antarctic seawater. Environ. Sci. Pollut. Res. Int. 2018, 25, 4667–4677. [Google Scholar] [CrossRef]
- Breezee, J.; Cady, N.; Staley, J.T. Subfreezing growth of the sea ice bacterium ‘Psychromonas ingrahamii’. Microb. Ecol. 2004, 47, 300–304. [Google Scholar] [CrossRef]
- Franzmann, P.D.; Liu, Y.; Balkwill, D.L.; Aldrich, H.C.; De Macario, E.C.; Boone, D.R. Methanogenium frigidum sp. nov.; a psychrophilic, H2-using methanogen from Ace Lake, Antarctica. Int. J. Syst. Bacteriol. 1997, 47, 1068–1072. [Google Scholar] [CrossRef]
- Caruso, C.; Rizzo, C.; Mangano, S.; Poli, A.; Di Donato, P.; Finore, I.; Nicolaus, B.; Di Marco, G.; Michaud, L.; Lo Giudice, A. Production and biotechnological potentialities of extracellular polymeric substances from sponge-associated Antarctic bacteria. Appl. Environ. Microbiol. 2018, 84, e01624-17. [Google Scholar]
- Caruso, C.; Rizzo, C.; Mangano, S.; Poli, A.; Di Donato, P.; Nicolaus, B.; Finore, I.; Di Marco, G.; Michaud, L.; Lo Giudice, A. First evidence of extracellular polymeric substance production by a cold-adapted Marinobacter isolate from Antarctic seawater. Antarct. Sci. 2018, in press. [Google Scholar]
- Methé, B.A.; Nelson, K.E.; Deming, J.W.; Momen, B.; Melamud, E.; Zhang, X.J.; Moult, J.; Madupu, R.; Nelson, W.C.; Dodson, R.J.; et al. The psychrophilic lifestyle as revealed by the genome sequence of Colwellia psychrerythraea 34H through genomic and proteomic analyses. Proc. Natl. Acad. Sci. USA 2005, 102, 10913–10918. [Google Scholar] [CrossRef]
- Médigue, C.; Krin, E.; Pascal, G.; Barbe, V.; Bernsel, A.; Bertin, P.N.; Cheung, F.; Cruveiller, S.; D’Amico, S.; Duilio, A.; et al. Coping with cold: The genome of the versatile marine Antarctica bacterium Pseudoalteromonas haloplanktis TAC125. Genome Res. 2005, 15, 1325–1335. [Google Scholar] [CrossRef]
- Bowman, J.P. Genomics of Psychrophilic Bacteria and Archaea. In Psychrophiles: From Biodiversity to Biotechnology; Margesin, R., Ed.; Springer: Cham, Switzerland, 2017. [Google Scholar]
- Martin, W.; Baross, J.; Kelley, D.; Russell, M.J. Hydrothermal vents and the origin of life. Nat. Rev. Microb. 2008, 6, 805–814. [Google Scholar] [CrossRef]
- Tarasov, V.G.; Propp, M.V.; Propp, L.N. Functions of the coastal marine ecosystems in relation of hydrothermal venting. Ecol. Chim. 2002, 11, 1–15. [Google Scholar]
- Maugeri, T.L.; Bianconi, G.; Canganella, F.; Danovaro, R.; Gugliandolo, C.; Italiano, F.; Lentini, V.; Manini, E.; Nicolaus, B. Shallow hydrothermal vents in the southern Tyrrhenian Sea. Chem. Ecol. 2010, 26, 285–298. [Google Scholar] [CrossRef]
- Hekinian, R.; Renard, V.; Cheminee, J.L. Hydrothermal deposits on the East Pacific Rise near 13°N: Geological setting and distribution of active sulfide chimneys. In Hydrothermal Processes at Seafloor Spreading Centers; Rona, P.A., Bostrom, K., Laubier, L., Smith, K.L., Jr., Eds.; Plenum Press: New York, NY, USA, 1984; pp. 571–594. [Google Scholar]
- Tarasov, V.G.; Gebruk, A.V.; Mironov, A.N.; Moskalev, L.I. Deep-sea and shallow-water hydrothermal vent communities: Two different phenomena? Chem. Geol. 2005, 224, 5–39. [Google Scholar] [CrossRef]
- Karl, D.M. The Microbiology of Deep-Sea Hydrothermal Vents; Karl, D.M., Ed.; CRC Press: Boca Raton, FL, USA, 1995. [Google Scholar]
- Takai, K.; Komatsu, T.; Inagaki, F.; Horikoshi, K. Distribution of Archaea in a black smoker chimney structure. Appl. Environ. Microbiol. 2001, 67, 618–629. [Google Scholar] [CrossRef]
- Oren, A. Industrial and environmental applications of halophilic microorganisms. Environ. Tech. 2010, 31, 825–834. [Google Scholar] [CrossRef]
- Oren, A. Life at high salt conditions. In the Prokaryotes. A Handbook on the Biology of Bacteria: Ecophysiology and Biochemistry; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; Volume 2, pp. 263–282. [Google Scholar]
- DasSarma, S.; DasSarma, P. Halophiles. In Encyclopedia of Life Sciences; John Wiley & Sons, Ltd.: Chichester, UK, 2012. [Google Scholar]
- Poli, A.; Kazak, H.; Gürleyendag, B.; Tommonaro, G.; Pieretti, G.; Toksoy Öner, E.; Nicolaus, B. High level synthesis of levan by a novel Halomonas species growing on defined media. Carbohydr. Polym. 2009, 78, 651–657. [Google Scholar] [CrossRef]
- Hasan, U.Ö.; Berna, S.A.; Burak, A.; Poli, A.; Denizci, A.A.; Utkan, G.; Nicolaus, B.; Kazan, D. Moderately Halophilic Bacterium Halomonas sp. AAD12: A promising candidate as a Hydroxyectoine Producer. J. Microbial. Biochem. Technol. 2015, 7, 262–268. [Google Scholar]
- Romano, I.; Poli, A.; Finore, I.; Huertas, F.J.; Gambacorta, A.; Pelliccione, S.; Nicolaus, G.; Lama, L.; Nicolaus, B. Haloterrigena hispanica sp. nov.; an extremely halophilic archaeon from Fuente de Piedra, Southern Spain. Int. J. Syst. Evol. Microbiol. 2007, 57, 1499–1503. [Google Scholar] [CrossRef] [PubMed]
- Romano, I.; Finore, I.; Nicolaus, G.; Huertas, F.J.; Lama, L.; Nicolaus, B.; Poli, A. Halobacillus alkaliphilus sp. nov.; a halophilic bacterium isolated from a salt lake in Fuente de Piedra, Southern Spain. Int. J. Syst. Evol. Microbiol. 2008, 58, 886–890. [Google Scholar] [CrossRef] [PubMed]
- Romano, I.; Orlando, P.; Gambacorta, A.; Nicolaus, B.; Dipasquale, L.; Pascual, J.; Giordano, A.; Lama, L. Salinivibrio sharmensis sp. nov, a novel haloalkaliphilic bacterium from a saline lake in Ras Mohammed Park (Egypt). Extremophiles 2011, 15, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Moreno, M.L.; Pérez, D.; García, M.T.; Mellado, E. Halophilic bacteria as a source of novel hydrolytic enzymes. Life 2013, 3, 38–51. [Google Scholar] [CrossRef] [PubMed]
- Dalmaso, G.Z.L.; Ferreira, D.; Vermelho, A.B. Marine extremophiles: A source of hydrolases for biotechnological applications. Mar. Drugs 2015, 13, 1925–1965. [Google Scholar] [CrossRef]
- Trincone, A. Enzymatic Processes in Marine Biotechnology. Mar. Drugs 2017, 15, 93. [Google Scholar] [CrossRef]
- Annamalai, N.; Rajeswari, M.; Balasubramanian, T. Enzymatic saccharification of pretreated rice straw by cellulase produced from Bacillus carboniphilus CAS 3 utilizing lignocellulosic wastes through statistical optimization. Biomass Bioenergy 2014, 68, 151–160. [Google Scholar] [CrossRef]
- Kim, B.; Lee, B.; Lee, Y.; Jin, I.; Chung, C.; Lee, J. Purification and characterization of carboxymethylcellulase isolated from a marine bacterium, Bacillus subtilis subsp. subtilis A-53. Enzym. Microb. Technol. 2009, 44, 411–416. [Google Scholar] [CrossRef]
- Annamalai, N.; Rajeswari, M.V.; Elayaraja, S.; Thavasi, R.; Vijayalakshmi, S.; Balasubramanian, T. Purification and Characterization of Thermostable Alkaline Cellulase from Marine Bacterium Bacillus licheniformis AU01 by Utilizing Cellulosic Wastes Waste Biomass. Valorization 2012, 3, 305–310. [Google Scholar] [CrossRef]
- Purwani, N.N.; Darmokoesoemo, H.; Tri Puspaningsih, N.N. Hydrolysis of Corncob Xylan using -xylosidase GbtXyl43B from Geobacillus thermoleovorans IT-08 Containing Carbohydrate Binding Module (CBM). Procedia Chem. 2016, 18, 75–81. [Google Scholar] [CrossRef]
- Harshvardhan, K.; Mishra, A.; Jha, B. Purification and characterization of cellulase from a marine Bacillus sp. H1666: A potential agent for single step saccharification of seaweed biomass. J. Mol. Catal. B Enzym. 2013, 93, 51–56. [Google Scholar] [CrossRef]
- Yagi, H.; Fujise, A.; Itabashi, N.; Ohshiro, T. Purification and characterization of a novel alginate lyase from the marine bacterium Cobetia sp. NAP1 isolated from brown algae. Biosci. Biotechnol. Biochem. 2016, 80, 2338–2346. [Google Scholar] [CrossRef]
- Mohapatra, B.R. Kinetic and thermodynamic properties of alginate lyase and cellulase co-produced by Exiguobacterium species Alg-S5. Int. J. Biol. Macromol. 2017, 98, 103–110. [Google Scholar] [CrossRef]
- Alkotaini, B.; Han, N.S.; Kim, B.S. Enhanced catalytic efficiency of endo-β-agarase I by fusion of carbohydrate-binding modules for agar prehydrolysis. Enzym. Microb. Technol. 2016, 93–94, 142–149. [Google Scholar] [CrossRef]
- Charoenpanich, J.; Suktanaraga, S.; Toobbucha, N. Production of a thermostable lipase by Aeromonas sp. EBB-1 isolated from marine sludge in Angsi1la. Thail. Sci. Asia 2011, 37, 105–114. [Google Scholar] [CrossRef]
- Mehrasbi, M.; Mohammadi, J.; Peyda, M.; Mohammadi, M. Covalent immobilization of Candida antarctica lipase on core-shell magnetic nanoparticles for production of biodiesel from waste cooking oil. Renew. Energy 2017, 101, 593–602. [Google Scholar] [CrossRef]
- Yang, K.; Sohn, J.; Kim, H. Catalytic properties of a lipase from Photobacterium lipolyticum for biodiesel production containing a high methanol concentration. J. Biosci. Bioeng. 2009, 107, 599–604. [Google Scholar] [CrossRef]
- Wu, Y.; Mao, A.; Sun, C.; Shanmugam, S.; Li, J.; Zhong, M. Catalytic hydrolysis of starch for biohydrogen production by using a newly identified amylase from a marine bacterium Catenovulum sp. X3. Int. J. Biol. Macromol. 2017, 104 Pt A, 716–723. [Google Scholar] [CrossRef]
- Sivaperumal, P.; Kamala, K.; Rajaram, R. Bioremediation of Industrial Waste through Enzyme Producing Marine Microorganisms. Adv. Food Nutr. Res. 2017, 80, 165–179. [Google Scholar] [PubMed]
- da Fonseca, F.; Angolini, C.; Zezzi Arruda, M.; Junior, C.; Santos, C.; Saraiva, A.; Pilau, E.; Souza, A.; Laborda, P.; de Oliveira, P.; et al. Identification of oxidoreductases from the petroleum Bacillus safensis strain. Biotechnol. Rep. 2015, 8, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.; Li, T.; Chang, F.; Zhou, P.; Fang, W.; Hong, Y.; Zhang, X.; Peng, H.; Xiao, Y. A new marine bacterial laccase with chloride-enhancing, alkaline-dependent activity and dye decolorization ability. Bioresour. Technol. 2012, 111, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, W.; Lai, Q.; Shao, Z. Gene diversity of CYP153A and AlkB alkane hydroxylases in oil-degrading bacteria isolated from the Atlantic Ocean. Environ. Microbiol. 2010, 12, 1230–1242. [Google Scholar] [CrossRef] [PubMed]
- Saito, A.; Iwabuchi, T.; Harayama, S. A novel phenanthrene dioxygenase from Nocardioides sp. Strain KP7: expression in Escherichia coli. J. Bacteriol. 2000, 182, 2134–2141. [Google Scholar] [CrossRef] [PubMed]
- Nikolaivits, E.; Dimarogona, M.; Fokialakis, N.; Topakas, E. Marine-Derived Biocatalysts: Importance, Accessing, and Application in Aromatic Pollutant Bioremediation. Front. Microbiol. 2017, 8, 265. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Cao, X.; Xin, Y.; Xue, S.; Zhang, W. Purification and characterization of a dehalogenase from Pseudomonas stutzeri DEH130 isolated from the marine sponge Hymeniacidon perlevis. World J. Microbiol. Biotechnol. 2013, 29, 1791–1799. [Google Scholar] [CrossRef]
- Zhang, J.; Xin, Y.; Cao, X.; Xue, S.; Zhang, W. Purification and characterization of 2-haloacid dehalogenase from marine bacterium Paracoccus sp. DEH99, isolated from marine sponge Hymeniacidon perlevis. J. Ocean Univ. China 2014, 13, 91–96. [Google Scholar] [CrossRef]
- Novak, H.R.; Sayer, C.; Panning, J.; Littlechild, J.A. Characterisation of an l-haloacid dehalogenase from the marine psychrophile Psychromonas ingrahamii with potential industrial application. Mar. Biotechnol. 2013, 15, 695–705. [Google Scholar] [CrossRef]
- Li, A.; Shao, Z. Biochemical characterization of a haloalkane dehalogenase DadB from Alcanivorax dieselolei B-5. PLoS ONE 2014, 9, e89144. [Google Scholar] [CrossRef]
- Ulaby, F.T.; Moore, R.K.; Fung, A.K. Microwave Remote Sensing: Active and Passive; Artech House: Norwood, MA, USA, 1981. [Google Scholar]
- Fung, K.; Chen, K.S. Microwave Scattering and Emission Models for Users; Artech House: Norwood, MA, USA, 1996. [Google Scholar]
- Kanevsky, M.B. Radar Imaging of Ocean Waves; Elsevier: Jordan Hill, UK, 2009. [Google Scholar]
- Donlon, C.; Berruti, B.; Buongiorno, A.; Ferreira, M.-H.; Femenias, P.; Frerick, J.; Goryl, P.; Klein, U.; Laur, H.; Mavrocordatos, C.; et al. The global monitoring for environment and security (GMES) Sentinel-3 mission. Remote Sens. Environ. 2012, 120, 37–57. [Google Scholar] [CrossRef]
- Bonekamp, H.; Montagner, F.; Santacesaria, V.; Noddo, C.N.; Wannop, S.; Tomazic, I.; O’Carroll, A.; Kwiatkowska, E.; Scharroo, R.; Wilson, H. Core operational Sentinel-3 marine data product services as part of the Copernicus space component. Ocean Sci. 2016, 12, 787–795. [Google Scholar] [CrossRef]
- Lee, J.-S.; Pottier, C. Polarimetric Radar Imaging: From Basics to Applications; CRC Press: Boca Raton, FL, USA, 2009. [Google Scholar]
- Van Zyl, J.; Kim, Y. Synthetic Aperture Radar Polarimetry; JPL Space Science and Technology Series; NASA: Washington, DC, USA, 2010. [Google Scholar]
- Wentz, F.J.; Gentemann, C.L.; Smith, D.K.; Chelton, D. Satellite measurements of sea surface temperature through clouds. Science 2000, 288, 847–850. [Google Scholar] [CrossRef]
- Hosoda, K. Review of satellite-based microwave observations of sea surface temperatures. J. Oceanogr. 2010, 66, 439–473. [Google Scholar] [CrossRef]
- Migliaccio, M.; Nunziata, F.; Buono, A. SAR polari0metry for sea oil slick observation. Int. J. Remote Sens. 2015, 36, 3243–3273. [Google Scholar] [CrossRef]
- Nunziata, F.; Buono, A.; Migliaccio, M. COSMO-SkyMed Synthetic Aperture Radar data to observe the Deepwater Horizon oil spill. Sustainability 2018, 10, 39599. [Google Scholar] [CrossRef]
- Nunziata, F.; de Macedo, C.R.; Buono, A.; Velotto, D.; Migliaccio, M. On the analysis of a time series of X-band TerraSAR-X SAR imagery over oil seepages. Int. J. Remote Sens. 2018, in press. [CrossRef]
- Meissner, T.; Wentz, F.J.; Le Vine, D.M. The salinity retrieval algorithms for the NASA Aquarius version 5 and SMAP version 3 releases. Remote Sens. 2018, 10, 1121. [Google Scholar] [CrossRef]
- Lagerloef, G.S.E. Satellite Remote Sensing: Salinity Measurements, Encyclopedia of Ocean Sciences; Academic Press: Cambridge, MA, USA, 2009. [Google Scholar]
- Tikhonov, V.V.; Raev, M.D.; Sharkov, E.A.; Boyarskii, D.A.; Repina, I.A.; Komarova, N. Satellite microwave radiometry of sea ice of polar regions: A review. Atmos. Ocean. Phys. 2016, 52, 1012–1030. [Google Scholar] [CrossRef]
- Ivanova, N.; Pedersen, L.T.; Tonboe, R.T.; Kern, S.; Heygster, G.; Lavergne, T.; Sørensen, A.; Saldo, R.; Dybkjær, G.; Brucker, L.; et al. Inter-comparison and evaluation of sea ice algorithms: Towards further identification of challenges and optimal approach using passive microwave observations. Cryosphere 2015, 9, 1797–1817. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Enzyme | Application | Ref. |
|---|---|---|---|
| Thermococcus sp. | Amylase (Fuelzyme®) | 1G-Bioethanol production | [36] |
| Bacillus carboniphilus CAS 3 | Cellulase | 2G-Bioethanol production | [38] |
| B. subtilis subsp. subtilis A-53 | Carboxymethylcellulase | [39] | |
| Bacillus licheniformis AU01 | Cellulase | [40] | |
| G. thermoleovorans IT-08 | Cellulase | [41] | |
| Bacillus sp. H1666 | Cellulase | 3G-Bioethanol production | [42] |
| Cobetia sp. NAP1 | Alginate lyase | [43] | |
| Exiguobacterium sp. Alg-S5 | Alginate lyase and cellulase | [44] | |
| Microbulbifer thermotolerans JAMB-A94 | Agarase | [45] | |
| Aeromonas sp. EBB-1 | Lipase | Biodiesel production | [46] |
| Candida antarctica | [47] | ||
| Photobacterium lipolyticum | [48] | ||
| Catenovulum sp. X3 | Amylase | Biohydrogen production | [49] |
| Source | Enzyme | Application | Ref. |
|---|---|---|---|
| Bacillus safensis (CFA-06) | Oxidoreductase | Biodegradation of aromatic compounds | [51] |
| Marine metagenome | Laccase | Degradation of industrial dyes | [52] |
| Alcanivorax borkumensis SK2T Alcanivorax dieselolei B-5T Alcanivorax venustensis ISO4T | Alkane hydroxylases/ Cytochrome P450 | Degradation of alkanes | |
| Bacillus licheniformis ATCC 14580T Bacillus litoralis DSM 16303T Bacillus oshimensis JCM Halomonas ventosae Al12T Idiomarina baltica DSM 15154T | [53] | ||
| Nocardioides sp. strain KP7 | Dioxygenase | Degradation of PAH | [54] |
| Pseudomonas stutzeri DEH130 Paracoccus sp. DEH99 Psychromonas ingrahamii Alcanivorax dieselolei strain B-5 | Haloalkane dehalogenases | Degradation of halogenated pollutants | [55,56] [57] [55,58] [59] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Donato, P.; Buono, A.; Poli, A.; Finore, I.; Abbamondi, G.R.; Nicolaus, B.; Lama, L. Exploring Marine Environments for the Identification of Extremophiles and Their Enzymes for Sustainable and Green Bioprocesses. Sustainability 2019, 11, 149. https://doi.org/10.3390/su11010149
Di Donato P, Buono A, Poli A, Finore I, Abbamondi GR, Nicolaus B, Lama L. Exploring Marine Environments for the Identification of Extremophiles and Their Enzymes for Sustainable and Green Bioprocesses. Sustainability. 2019; 11(1):149. https://doi.org/10.3390/su11010149
Chicago/Turabian StyleDi Donato, Paola, Andrea Buono, Annarita Poli, Ilaria Finore, Gennaro Roberto Abbamondi, Barbara Nicolaus, and Licia Lama. 2019. "Exploring Marine Environments for the Identification of Extremophiles and Their Enzymes for Sustainable and Green Bioprocesses" Sustainability 11, no. 1: 149. https://doi.org/10.3390/su11010149
APA StyleDi Donato, P., Buono, A., Poli, A., Finore, I., Abbamondi, G. R., Nicolaus, B., & Lama, L. (2019). Exploring Marine Environments for the Identification of Extremophiles and Their Enzymes for Sustainable and Green Bioprocesses. Sustainability, 11(1), 149. https://doi.org/10.3390/su11010149