Climate Change Sensitivity of Multi-Species Afforestation in Semi-Arid Benin

 ,
, 
 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
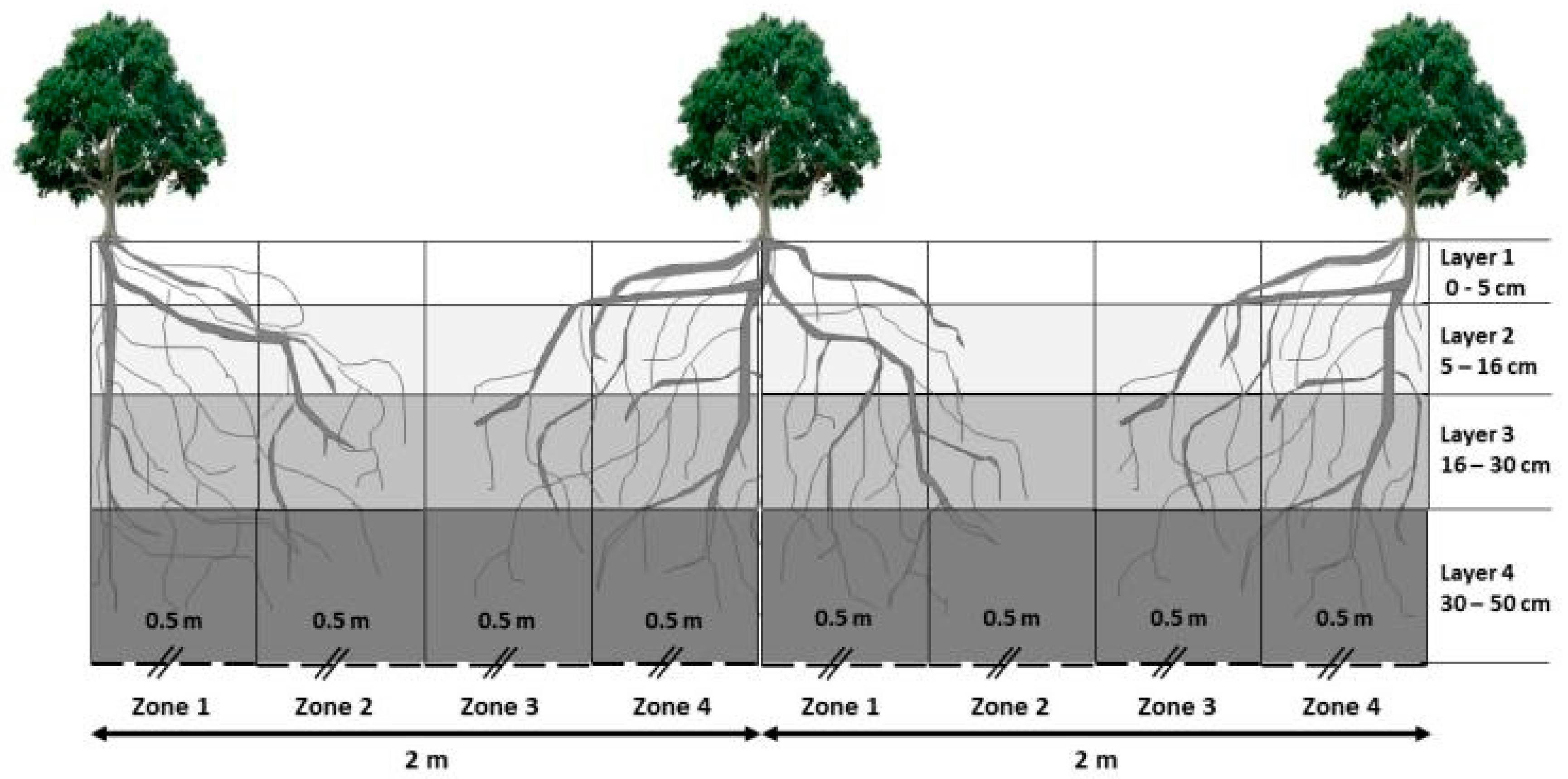
2.2. Experimental Design and Woody Species
2.3. Data Collection
2.4. Modelling Analysis
2.4.1. Description of WaNuLCAS
2.4.2. Input Data for WaNuLCAS
2.4.3. Model Calibration, Validation, and Evaluation
2.4.4. Climate–Growth Relationship Analysis
2.4.5. Scenario Description
3. Results
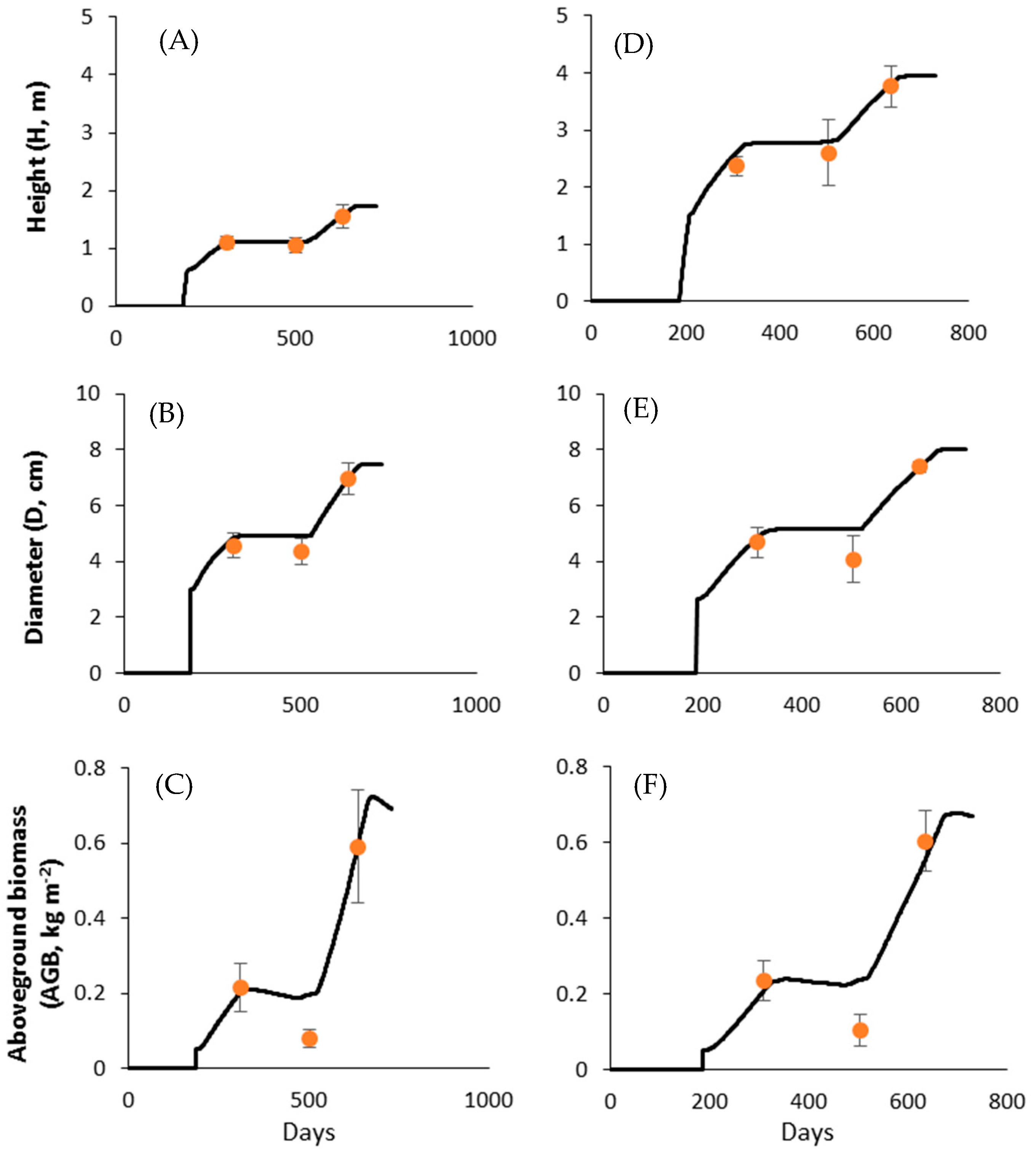
3.1. Simulation of Tree Height, Diameter, and Aboveground Biomass
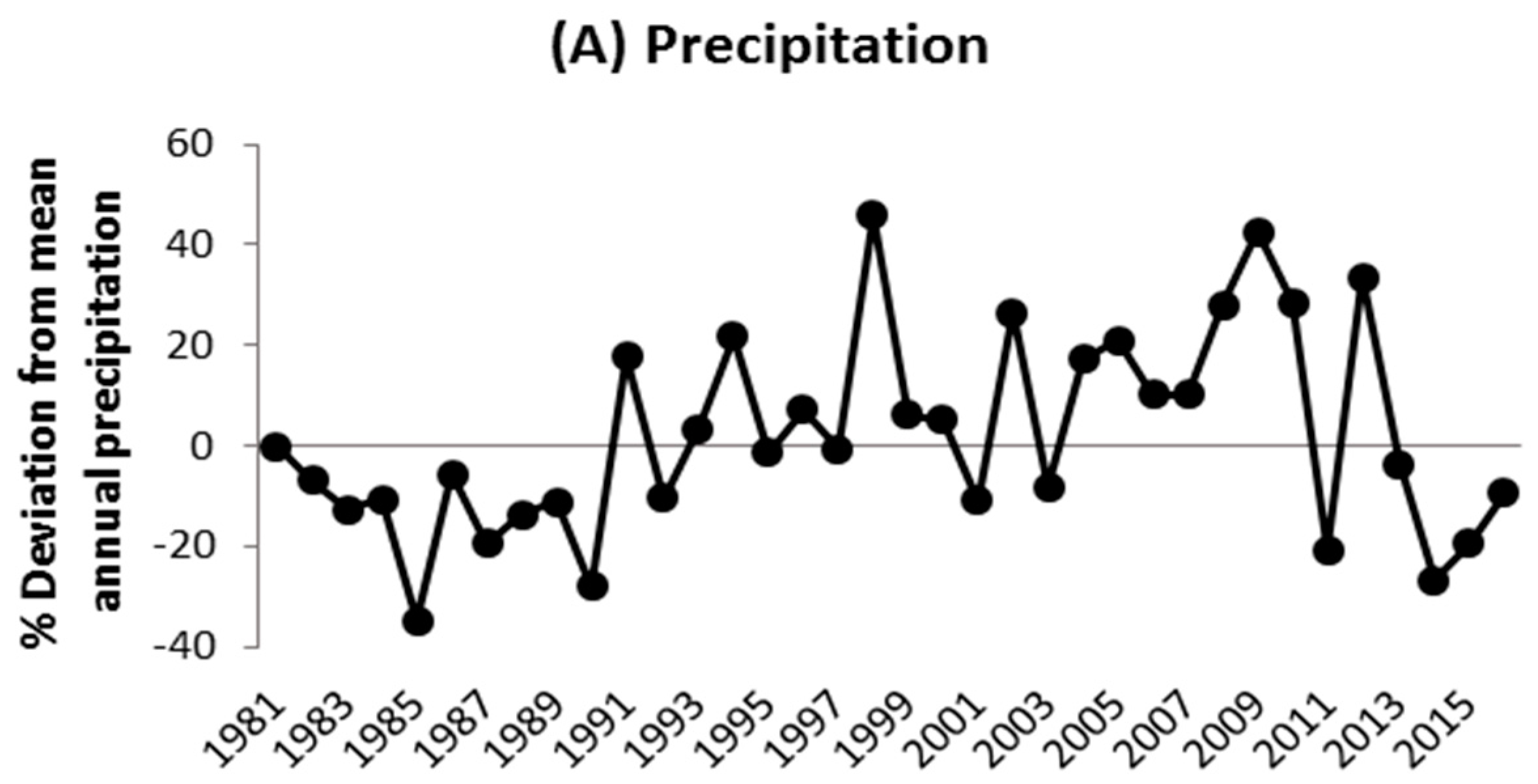
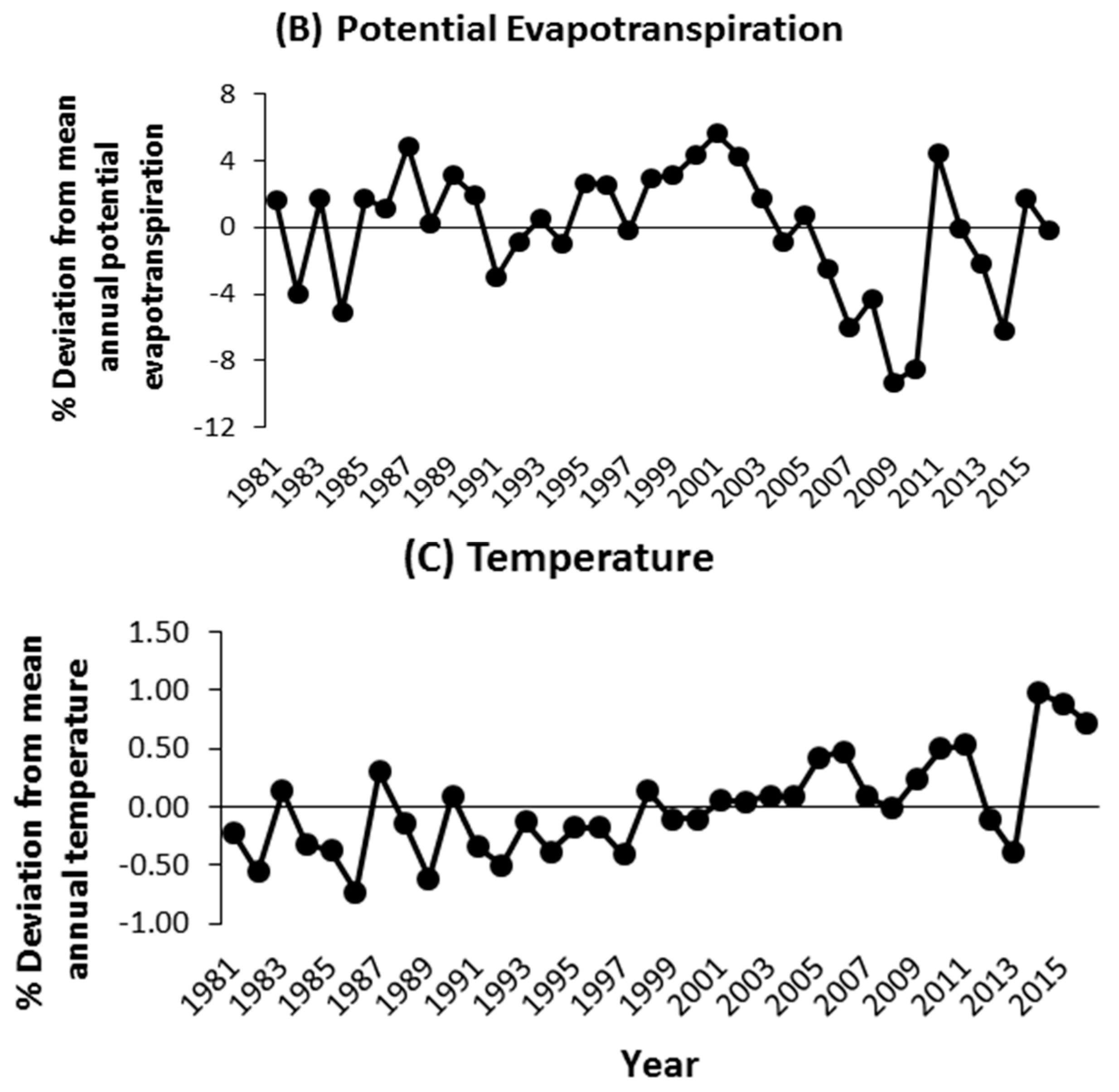
3.2. Climate–Growth Relationships of the Afforestation Species
3.3. Simulated Effect of Rooting Depth on Sapling Biomass under Contrasting Water Stress Conditions
4. Discussion
4.1. Model Performance
4.2. Climate Sensitivity of Afforestation Species
4.3. Effects of Rooting Depth on Biomass Growth under Extreme Dry Conditions
4.4. Replicability of the Approach
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Criteria | Symbol | Formula | Acceptable Range | Optimum Value |
|---|---|---|---|---|
| Model efficiency | EF | ≤1 | 1 | |
| Coefficient of determination | CD | 0.5–2 * | 1 | |
| Maximum error | ME | ≥0 | 0 | |
| Root mean square error | RMSE | ≥0 | 0 | |
| Coefficient of residual mass | CRM | ≤1 | 0 |



References
- UNFCCC. Kyoto Protocol to the United Nations Framework Convention on Climate Change; United Nations Framework Convention on Climate Change: Bonn, Germany, 1997. [Google Scholar]
- UNFCCC. Outcome of the work of the ad hoc working group on long-term cooperative action under the convention—Policy approaches and positive incentives on issues relating to reducing emissions from deforestation and forest degradation in developing countries, and the role of conservation, sustainable management of forests and enhancement of forest carbon stocks in developing countries. In Proceedings of the Ad Hoc Working Group on Long-Term Cooperative Actionunder the Convention, Copenhagen, Denmark, 7–15 December 2009. [Google Scholar]
- Garrity, D.P.; Akinnifesi, F.K.; Ajayi, O.C.; Weldesemayat, S.G.; Mowo, J.G.; Kalinganire, A.; Larwanou, M.; Bayala, J. Evergreen Agriculture: A robust approach to sustainable food security in Africa. Food Secur. 2010, 2, 197–214. [Google Scholar] [CrossRef]
- Sanderson, M.; Santini, M.; Valentini, R.; Pope, E. Relationships between Forests and Weather; EC Directorate General of the Environment: Brussels, Belgium, 2012. [Google Scholar]
- Betts, R.A.; Boucher, O.; Collins, M.; Cox, P.M.; Falloon, P.D.; Gedney, N.; Hemming, D.L.; Huntingford, C.; Jones, C.D.; Sexton, D.M.H.; et al. Projected increase in continental runoff due to plant responses to increasing carbon dioxide. Nature 2007, 448, 1037–1041. [Google Scholar] [CrossRef] [PubMed]
- Hosonuma, N.; Herold, M.; De Sy, V.; De Fries, R.S.; Brockhaus, M.; Verchot, L.; Angelsen, A.; Romijn, E. An assessment of deforestation and forest degradation drivers in developing countries. Environ. Res. Lett. 2012, 7, 044009. [Google Scholar] [CrossRef]
- Pearson, T.R.H.; Brown, S.; Murray, L.; Sidman, G. Greenhouse gas emissions from tropical forest degradation: An underestimated source. Carbon Balance Manag. 2017, 12. [Google Scholar] [CrossRef] [PubMed]
- Pachauri, R.; Allen, M.; Barros, V.; Broome, J.; Cramer, W.; Christ, R.; Church, J.; Clarke, L.; Dahe, Q.; Dasgupta, P.; et al. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC, Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2014. [Google Scholar]
- Elliott, K.J.; Miniat, C.F.; Pederson, N.; Laseter, S.H. Forest tree growth response to hydroclimate variability in the southern Appalachians. Glob. Chang. Biol. 2015, 21, 4627–4641. [Google Scholar] [CrossRef] [PubMed]
- Thurm, E.A.; Uhl, E.; Pretzsch, H. Mixture reduces climate sensitivity of Douglas-fir stem growth. For. Ecol. Manag. 2016, 376, 205–220. [Google Scholar] [CrossRef]
- Poorter, L. Biodiversity of West African Forests: An Ecological Atlas of Woody Plant Species; CABI: Wallingford, UK, 2004. [Google Scholar]
- Sylla, M.B.; Giorgi, F.; Pal, J.S.; Gibba, P.; Kebe, I.; Nikiema, M. Projected Changes in the Annual Cycle of High-Intensity Precipitation Events over West Africa for the Late Twenty-First Century. J. Clim. 2015, 28, 6475–6488. [Google Scholar] [CrossRef]
- Sylla, M.B.; Nikiema, P.M.; Gibba, P.; Kebe, I.; Klutse, N.A.B. Climate Change over West Africa: Recent Trends and Future Projections. In Adaptation to Climate Change and Variability in Rural West Africa; Yaro, J.A., Hesselberg, J., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 25–40. ISBN 978-3-319-31497-6. [Google Scholar]
- Abdulai, I.; Vaast, P.; Hoffmann, M.P.; Asare, R.; Jassogne, L.; Van Asten, P.; Rötter, R.P.; Graefe, S. Cocoa agroforestry is less resilient to sub-optimal and extreme climate than cocoa in full sun. Glob. Chang. Biol. 2018, 24, 273–286. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Huang, J.; He, Y.; Guan, Y. Combined effects of the Pacific Decadal Oscillation and El Niño-Southern Oscillation on Global Land Dry–Wet Changes. Sci. Rep. 2015, 4. [Google Scholar] [CrossRef] [PubMed]
- Castro, J.; Zamora, R.; Hódar, J.A.; Gómez, J.M. Alleviation of Summer Drought Boosts Establishment Success of Pinus sylvestris in a Mediterranean Mountain: An Experimental Approach. Plant Ecol. 2005, 181, 191–202. [Google Scholar] [CrossRef]
- Comita, L.S.; Engelbrecht, B.M.J. Drought as a driver of tropical tree species regeneration dynamics and distribution patterns. In Forests and Global Change; Coomes, D.A., Burslem, D.F.R.P., Simonson, W.D., Eds.; Cambridge University Press: Cambridge, UK, 2014; pp. 261–308. ISBN 978-1-107-32350-6. [Google Scholar]
- Noulèkoun, F.; Lamers, J.P.A.; Naab, J.; Khamzina, A. Shoot and root responses of woody species to silvicultural management for afforestation of degraded croplands in the Sudano-Sahelian zone of Benin. For. Ecol. Manag. 2017, 385, 254–263. [Google Scholar] [CrossRef]
- Geßler, A.; Keitel, C.; Kreuzwieser, J.; Matyssek, R.; Seiler, W.; Rennenberg, H. Potential risks for European beech (Fagus sylvatica L.) in a changing climate. Trees 2007, 21, 1–11. [Google Scholar] [CrossRef]
- Noulèkoun, F.; Khamzina, A.; Naab, J.B.; Lamers, J.P.A. Biomass allocation in five semi-arid afforestation species is driven mainly by ontogeny rather than resource availability. Ann. For. Sci. 2017, 74. [Google Scholar] [CrossRef]
- Van Noordwijk, M.; van Lawson, G.; Hairiah, K.; Wilson, J. Root distribution of trees and crops: Competition and/or complementarity. In Tree-Crop Interactions: Agroforestry in a Changing Climate; Ong, C.K., Black, C.R., Wilson, J., Eds.; CABI: Wallingford, UK, 2015; pp. 221–257. ISBN 978-1-78064-511-7. [Google Scholar]
- Hartl-Meier, C.; Zang, C.; Buntgen, U.; Esper, J.; Rothe, A.; Gottlein, A.; Dirnbock, T.; Treydte, K. Uniform climate sensitivity in tree-ring stable isotopes across species and sites in a mid-latitude temperate forest. Tree Physiol. 2015, 35, 4–15. [Google Scholar] [CrossRef] [PubMed]
- Gebrekirstos, A.; Mitlöhner, R.; Teketay, D.; Worbes, M. Climate-growth relationships of the dominant tree species from semi-arid savanna woodland in Ethiopia. Trees 2008, 22, 631–641. [Google Scholar] [CrossRef]
- Teets, A.; Fraver, S.; Weiskittel, A.R.; Hollinger, D.Y. Quantifying climate-growth relationships at the stand level in a mature mixed-species conifer forest. Glob. Chang. Biol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Raz-Yaseef, N.; Yakir, D.; Schiller, G.; Cohen, S. Dynamics of evapotranspiration partitioning in a semi-arid forest as affected by temporal rainfall patterns. Agric. For. Meteorol. 2012, 157, 77–85. [Google Scholar] [CrossRef]
- Noulèkoun, F.; Naab, J.B.; Lamers, J.P.A.; Baumert, S.; Khamzina, A. Sapling biomass allometry and carbon content in five afforestation species on marginal farmland in semi-arid Benin. New For. 2018, 49, 363–382. [Google Scholar] [CrossRef]
- Schöngart, J.; Orthmann, B.; Hennenberg, K.J.; Porembski, S.; Worbes, M. Climate-growth relationships of tropical tree species in West Africa and their potential for climate reconstruction. Glob. Chang. Biol. 2006, 12, 1139–1150. [Google Scholar] [CrossRef]
- Myers-Smith, I.H.; Elmendorf, S.C.; Beck, P.S.A.; Wilmking, M.; Hallinger, M.; Blok, D.; Tape, K.D.; Rayback, S.A.; Macias-Fauria, M.; Forbes, B.C.; et al. Climate sensitivity of shrub growth across the tundra biome. Nat. Clim. Chang. 2015, 5, 887–891. [Google Scholar] [CrossRef]
- Brienen, R.J.W.; Zuidema, P.A. Lifetime growth patterns and ages of Bolivian rain forest trees obtained by tree ring analysis. J. Ecol. 2006, 94, 481–493. [Google Scholar] [CrossRef]
- Herrera-Ramirez, D.; Andreu-Hayles, L.; del Valle, J.I.; Santos, G.M.; Gonzalez, P.L.M. Nonannual tree rings in a climate-sensitive Prioria copaifera chronology in the Atrato River, Colombia. Ecol. Evol. 2017, 7, 6334–6345. [Google Scholar] [CrossRef] [PubMed]
- Bouriaud, O.; Bréda, N.; Dupouey, J.-L.; Granier, A. Is ring width a reliable proxy for stem-biomass increment? A case study in European beech. Can. J. For. Res. 2005, 35, 2920–2933. [Google Scholar] [CrossRef]
- Foster, J.R.; D’Amato, A.W.; Bradford, J.B. Looking for age-related growth decline in natural forests: Unexpected biomass patterns from tree rings and simulated mortality. Oecologia 2014, 175, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Kimmins, J.P.; Mailly, D.; Seely, B. Modelling forest ecosystem net primary production: The hybrid simulation approach used in forecast. Ecol. Model. 1999, 122, 195–224. [Google Scholar] [CrossRef]
- Johnsen, K.; Samuelson, L.; Teskey, R.; McNulty, S.; Fox, T. Process Models as Tools in Forestry Research and Management. For. Sci. 2001, 47, 2–8. [Google Scholar]
- Van Noordwijk, M.; Lusiana, B. WaNuLCAS, a model of water, nutrient and light capture in agroforestry systems. In Agroforestry for Sustainable Land-Use Fundamental Research and Modelling with Emphasis on Temperate and Mediterranean Applications; Auclair, D., Dupraz, C., Eds.; Springer: Dordrecht, The Netherlands, 1999; Volume 60, pp. 217–242. ISBN 978-90-481-5258-2. [Google Scholar]
- Kang, H.; Seely, B.; Wang, G.; Cai, Y.; Innes, J.; Zheng, D.; Chen, P.; Wang, T. Simulating the impact of climate change on the growth of Chinese fir plantations in Fujian province, China. N. Z. J. For. Sci. 2017, 47. [Google Scholar] [CrossRef]
- Bayala, J.; Sanou, J.; Teklehaimanot, Z.; Ouedraogo, S.J.; Kalinganire, A.; Coe, R.; Van Noordwijk, M. Advances in knowledge of processes in soil–tree–crop interactions in parkland systems in the West African Sahel: A review. Agric. Ecosyst. Environ. 2015, 205, 25–35. [Google Scholar] [CrossRef]
- Luedeling, E.; Smethurst, P.J.; Baudron, F.; Bayala, J.; Huth, N.I.; van Noordwijk, M.; Ong, C.K.; Mulia, R.; Lusiana, B.; Muthuri, C.; et al. Field-scale modeling of tree–crop interactions: Challenges and development needs. Agric. Syst. 2016, 142, 51–69. [Google Scholar] [CrossRef]
- Bayala, J.; van Noordwijk, M.; Lusiana, B.; Ni’matul, K.; Teklehaimanot, Z.; Ouedraogo, S.J. Separating the Tree–Soil–Crop Interactions in Agroforestry Parkland Systems in Saponé (Burkina Faso) using WaNuLCAS. In Toward Agroforestry Design; Jose, S., Gordon, A.M., Eds.; Springer: Dordrecht, The Netherlands, 2008; Volume 4, pp. 285–297. ISBN 978-1-4020-6571-2. [Google Scholar]
- Bayala, J.; Heng, L.K.; van Noordwijk, M.; Ouedraogo, S.J. Hydraulic redistribution study in two native tree species of agroforestry parklands of West African dry savanna. Acta Oecol. 2008, 34, 370–378. [Google Scholar] [CrossRef]
- Coulibaly, Y.N.; Mulia, R.; Sanou, J.; Zombré, G.; Bayala, J.; Kalinganire, A.; van Noordwijk, M. Crop production under different rainfall and management conditions in agroforestry parkland systems in Burkina Faso: Observations and simulation with WaNuLCAS model. Agrofor. Syst. 2014, 88, 13–28. [Google Scholar] [CrossRef]
- Van Noordwijk, M.; Cadisch, G. Access and excess problems in plant nutrition. In Progress in Plant Nutrition: Plenary Lectures of the XIV International Plant Nutrition Colloquium; Horst, W.J., Bürkert, A., Claassen, N., Flessa, H., Frommer, W.B., Goldbach, H., Merbach, W., Olfs, H.-W., Römheld, V., Sattelmacher, B., et al., Eds.; Springer: Dordrecht, The Netherlands, 2002; pp. 25–40. ISBN 978-90-481-6191-1. [Google Scholar]
- Cao, K. Water relations and gas exchange of tropical saplings during a prolonged drought in a Bornean heath forest, with reference to root architecture. J. Trop. Ecol. 2000, 16, 101–116. [Google Scholar] [CrossRef]
- Gibbons, J.M.; Newbery, D.M. Drought avoidance and the effect of local topography on trees in the understorey of Bornean lowland rain forest. Plant Ecol. 2003, 164, 1–18. [Google Scholar] [CrossRef]
- Engelbrecht, B.M.J.; Kursar, T.A.; Tyree, M.T. Drought effects on seedling survival in a tropical moist forest. Trees 2005, 19, 312–321. [Google Scholar] [CrossRef]
- Adegbidi, A.; Burger, K.; Gandonou, E.; Mulder, I. Farmers’ Perceptions and Sustainable Land Use in the Atacora, Benin; CREED Working Paper 22; International Institute for Environment and Development: London, UK, 1999. [Google Scholar]
- Mulder, I. Soil Degradation in Benin: Farmers’ Perceptions and Responses. Ph.D. Thesis, Tinbergen Institute, Vrije Universiteit Amsterdam, Amsterdam, The Netherlands, 2000. [Google Scholar]
- Gandji, K.; Chadare, F.J.; Idohou, R.; Salako, V.K.; Assogbadjo, A.E.; Kakaï, R.L.G. Status and utilisation of Moringa oleifera Lam: A review. Afr. Crop Sci. J. 2018, 26, 137–156. [Google Scholar] [CrossRef]
- WRB (Ed.) World Reference Base for Soil Resources 2006: A Framework for International Classification, Correlation and Communication; World Soil Resources Reports; FAO: Rome, Italy, 2006; ISBN 978-92-5-105511-3. [Google Scholar]
- Walter, H.; Lieth, H. Climate Diagram World Atlas; VEB Gustav Fischer: Jena, Germany, 1967. [Google Scholar]
- Van Noordwijk, M.; Lusiana, B.; Khasanah, N.; Mulia, R. WaNuLCAS Version 4.0, Background on a Model of Water, Nutrient and Light Capture in Agrofoerestry Systems; World Agroforestry Center—ICRAF, SEA Regional Office: Bogor, Indonesia, 2011; ISBN 978-979-3198-59-0. [Google Scholar]
- De Willigen, P.; van Noordwijk, M. Roots for Plant Production and Nutrient Use Efficiency; Agricultural University Wageningen: Wageningen, The Netherlands, 1987. [Google Scholar]
- Emerman, S.H. The tipping bucket equations as a model for macropore flow. J. Hydrol. 1995, 171, 23–47. [Google Scholar] [CrossRef]
- De Willigen, P.; Nielsen, N.E.; Claassen, N.; Castrignanò, A.M. Modelling Water and Nutrient Uptake. In Root Methods; Smit, A.L., Bengough, A.G., Engels, C., van Noordwijk, M., Pellerin, S., van de Geijn, S.C., Eds.; Springer: Berlin/Heidelberg, Germany, 2000; ISBN 978-3-662-04188-8. [Google Scholar]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration—Guidelines for Computing Crop Water Requirements; FAO Irrigation and Drainage Paper 56; FAO: Rome, Italy, 1998; Volume 300. [Google Scholar]
- Hodnett, M.G.; Tomasella, J. Marked differences between van Genuchten soil water-retention parameters for temperate and tropical soils: A new water-retention pedo-transfer functions developed for tropical soils. Geoderma 2002, 108, 155–180. [Google Scholar] [CrossRef]
- Walker, A.P.; Mutuo, P.K.; Albrecht, A.; Cadisch, G. Modelling of planted legume fallows in Western Kenya using WaNuLCAS. (I) Model calibration and validation. Agrofor. Syst. 2007, 70, 197–209. [Google Scholar] [CrossRef]
- Khasanah, N.; Perdana, A.; Rahmanullah, A.; Manurung, G.; Roshetko, J.M.; van Noordwijk, M. Intercropping teak (Tectona grandis) and maize (Zea mays): Bioeconomic trade-off analysis of agroforestry management practices in Gunungkidul, West Java. Agrofor. Syst. 2015, 89, 1019–1033. [Google Scholar] [CrossRef]
- Loague, K.; Green, R.E. Statistical and graphical methods for evaluating solute transport models: Overview and application. J. Contam. Hydrol. 1991, 7, 51–73. [Google Scholar] [CrossRef]
- Gazol, A.; Camarero, J.J. Mediterranean dwarf shrubs and coexisting trees present different radial-growth synchronies and responses to climate. Plant Ecol. 2012, 213, 1687–1698. [Google Scholar] [CrossRef]
- Johnson, J.B.; Omland, K.S. Model selection in ecology and evolution. Trends Ecol. Evol. 2004, 19, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Burnham, K.P.; Anderson, D.R. Multimodel Inference: Understanding AIC and BIC in Model Selection. Sociol. Methods Res. 2004, 33, 261–304. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D. R Core Team nlme: Linear and Nonlinear Mixed Effects Models; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Duan, J.; Sikka, A.; Grant, G. A comparison of stochastic models for generating daily precipitation at the HJ Andrews Experimental Forest. Northwest Sci. 1995, 69, 318–329. [Google Scholar]
- Nakagawa, S.; Schielzeth, H.; O’Hara Robert, B. A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol. Evol. 2013, 4, 133–142. [Google Scholar] [CrossRef]
- Rodgers, V.L.; Smith, N.G.; Hoeppner, S.S.; Dukes, J.S. Warming increases the sensitivity of seedling growth capacity to rainfall in six temperate deciduous tree species. AoB PLANTS 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Friedrichs, D.A.; Trouet, V.; Büntgen, U.; Frank, D.C.; Esper, J.; Neuwirth, B.; Löffler, J. Species-specific climate sensitivity of tree growth in Central-West Germany. Trees 2009, 23, 729–739. [Google Scholar] [CrossRef]
- Rykiel, E.J. Testing ecological models: The meaning of validation. Ecol. Model. 1996, 90, 229–244. [Google Scholar] [CrossRef]
- Hussain, K.; Wongleecharoen, C.; Hilger, T.; Ahmad, A.; Kongkaew, T.; Cadisch, G. Modelling resource competition and its mitigation at the crop-soil-hedge interface using WaNuLCAS. Agrofor. Syst. 2016, 90, 1025–1044. [Google Scholar] [CrossRef]
- Kozlowski, T.T.; Winget, C.H. Diurnal and Seasonal Variation in Radii of Tree Stems. Ecology 1964, 45, 149–155. [Google Scholar] [CrossRef]
- Muthuri, C.W.; Ong, C.K.; Black, C.R.; Mati, B.M.; Ngumi, V.W.; van Noordwijk, M. Modelling the effects of leafing phenology on growth and water use by selected agroforestry tree species in semi-arid Kenya. Land Use Water Resour. Res. 2004, 4, 1–11. [Google Scholar]
- Ettinger, A.K.; Ford, K.R.; HilleRisLambers, J. Climate determines upper, but not lower, altitudinal range limits of Pacific Northwest conifers. Ecology 2011, 92, 1323–1331. [Google Scholar] [CrossRef] [PubMed]
- Weber, P.; Bugmann, H.; Rigling, A. Radial growth responses to drought of Pinus sylvestris and Quercus pubescens in an inner-Alpine dry valley. J. Veg. Sci. 2007, 18, 777–792. [Google Scholar] [CrossRef]
- Ministère de l’Environnement, de l’Habitat et de l’Urbanisme (MEHU). Deuxième Communication Nationale de la République du Bénin sur les Changements Climatiques; MEHU: Cotonou, Republic of Benin, 2011. [Google Scholar]






| Parameter | Soil Layer (cm) | |||
|---|---|---|---|---|
| 0–5 | 5–16 | 16–30 | 30–50 | |
| Clay (%) | 11.8 | 15.7 | 25.5 | 29.4 |
| Silt (%) | 31.4 | 35.3 | 33.3 | 37.3 |
| Sand (%) | 56.9 | 49.0 | 41.2 | 33.3 |
| BD (g cm−3) | 1.5 | 1.5 | 1.6 | 1.6 |
| pH-H2O | 5.3 | 5.2 | 5.7 | 5.8 |
| TOC (%) | 0.7 | 0.6 | 0.6 | 0.6 |
| Ntot (%) | 0.1 | 0.1 | 0.1 | 0.1 |
| P (mg kg−1) | 15.7 | 15.3 | 14.5 | 14.8 |
| CEC (cmol kg−1) | 4.7 | 6.0 | 5.2 | 5.3 |
| Ksat (cm d−1) | 73.2 | 18.4 | 207.7 | 232.4 |
| Species | Biomass Fractions | a | b |
|---|---|---|---|
| Jatropha curcas | Total | 0.004 | 2.93 |
| Stem | 0.004 | 2.74 | |
| Leaf + Twig | 0.003 | 2.48 | |
| Litterfall * | 0.0002 | 3.29 | |
| Moringa oleifera | Total | 0.010 | 2.36 |
| Stem | 0.008 | 2.38 | |
| Leaf + Twig | 0.010 | 1.49 | |
| Litterfall * | 0.0002 | 3.29 |
| Parameters | Unit | Description | Definition | Range | 2014 | Rank ** | 2015 | Rank ** |
|---|---|---|---|---|---|---|---|---|
| AWD | mm | Annual water deficit | Difference between annual potential evapotranspiration (ET0) and precipitation | 155–1153 | 931 | 29/36 | 978 | 31/36 |
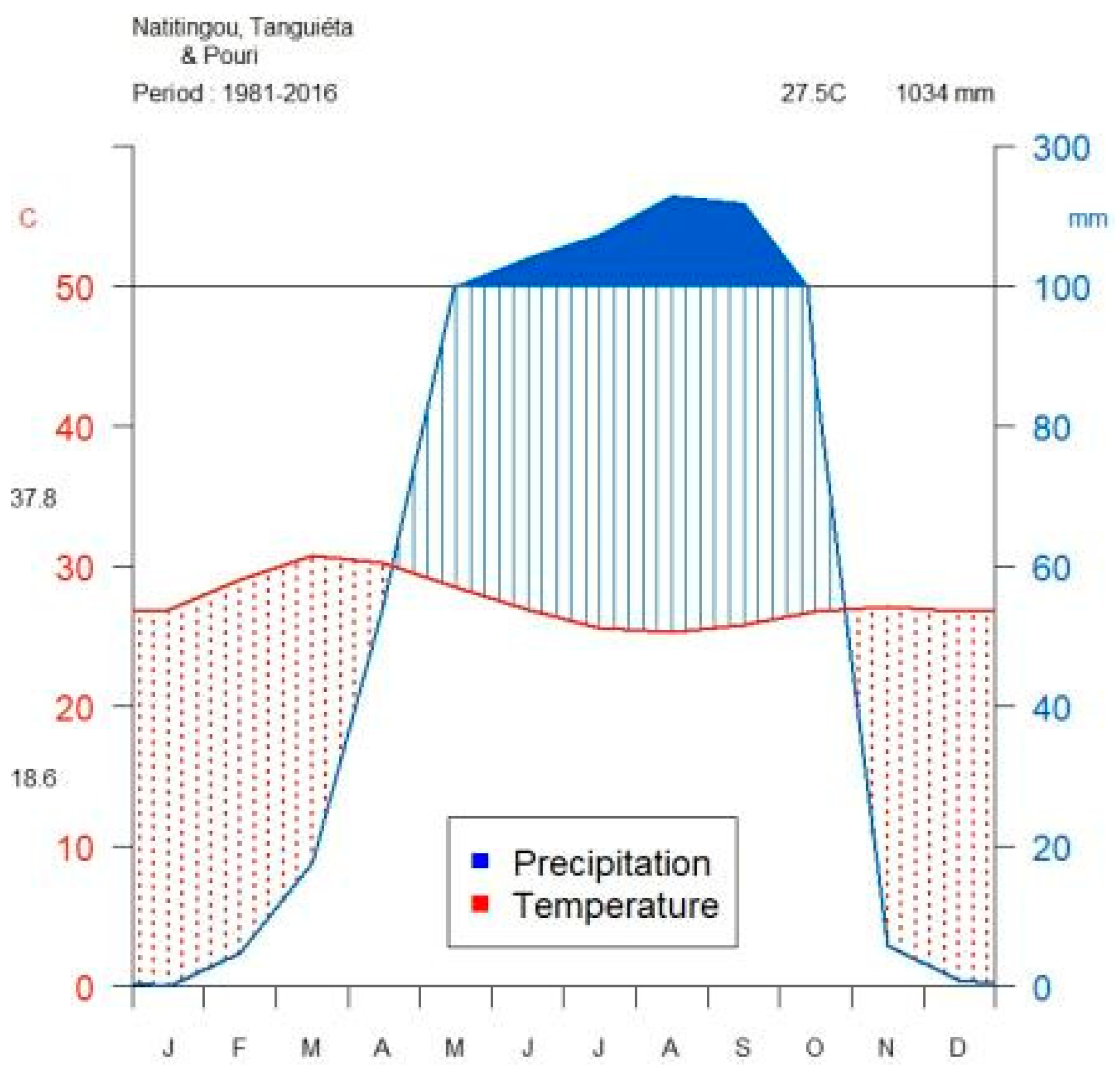
| LDSP | Days | Length of the longest dry spell | Maximum number of consecutive days of no precipitation within the rainy season (April–October; Figure 1) | 3–40 | 24 | 32/36 | 19 | 29/36 |
| LEDP * | Days | Length of extremely dry period | Number of days within a year when daily maximum temperature >99th percentile (40° C) and no precipitation (Zhang et al. 2011) | 0–42 | 8 | 30/36 | 42 | 36/36 |
| LDS | Months | Length of dry season | Number of consecutive months within a year with less than 100 mm of rainfall (Wood et al. 2001) | 5–10 | 10 | 36/36 | 9 | 35/36 |
| LWS | Day | Length of the longest wet spell | Maximum number of consecutive rainy days within the rainy season (April–October; see Figure 1) | 2–21 | 10 | 30/36 | 21 | 36/36 |
| ATWP | mm | Annual total wet-day precipitation | Annual total precipitation from wet days (i.e., days with precipitation ≥1 mm) (Zhang et al. 2011) | 670–1501 | 749 | 3/36 | 825 | 5/36 |
| R95p * | mm | Very wet days | Annual total precipitation from days with precipitation >95th percentile (19 mm) (Zhang et al. 2011) | 344–1096 | 395 | 4/36 | 424 | 7/36 |
| R95pot | - | Total precipitation with respect to the R95p | Fraction of precipitation accounted for by the very wet days (R95p) (Zhang et al. 2011) | 0.42–0.80 | 0.52 | 10/36 | 0.51 | 7/36 |
| Species | Growth Parameters | EF (1) | CD (1) | RMSE (0) | CRM (0) | ME (0) | R2 (1) |
|---|---|---|---|---|---|---|---|
| Jatropha curcas | Height (H, m) | 0.72 | 0.71 | 6.02 | 0.00 | 0.16 | 0.80 |
| Diameter (D, cm) | 0.72 | 2.48 | 11.12 | −0.03 | 1.17 | 0.81 | |
| Aboveground biomass (AGB, kg m−2) | 0.77 | 2.21 | 33.78 | −0.04 | 0.19 | 0.83 | |
| Moringa oleifera | Height (H, m) | 0.82 | 2.59 | 9.42 | −0.03 | 0.40 | 0.97 |
| Diameter (D, cm) | 0.70 | 1.76 | 12.94 | −0.08 | 1.32 | 0.88 | |
| Aboveground biomass (AGB, kg m−2) | 0.78 | 2.23 | 28.55 | −0.09 | 0.13 | 0.88 |
| Variables | Jatropha curcas | Moringa oleifera | ||||
|---|---|---|---|---|---|---|
| ΔAICc | Slope | R² | ΔAICc | Slope | R² | |
| LDSP1 | 12.22 | −0.63 | 0.39 | 9.33 | −0.58 | 0.33 |
| LDSP2 | 9.91 | −0.59 | 0.35 | 8.31 | −0.57 | 0.31 |
| LDS1 | 2.27 | −0.43 | 0.18 | 2.16 | −0.42 | 0.18 |
| LDS2 | 12.16 | −0.63 | 0.39 | 9.13 | −0.58 | 0.33 |
| AWD1 | 6.02 | −0.53 | 0.27 | 5.05 | −0.5 | 0.24 |
| AWD2 | 14.92 | −0.67 | 0.44 | 11.98 | −0.63 | 0.38 |
| ATWP1 | 3.60 | 0.46 | 0.21 | 2.96 | 0.45 | 0.20 |
| ATWP2 | 10.44 | 0.60 | 0.36 | 8.32 | 0.57 | 0.31 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Noulèkoun, F.; Khamzina, A.; Naab, J.B.; Khasanah, N.; Van Noordwijk, M.; Lamers, J.P.A. Climate Change Sensitivity of Multi-Species Afforestation in Semi-Arid Benin. Sustainability 2018, 10, 1931. https://doi.org/10.3390/su10061931
Noulèkoun F, Khamzina A, Naab JB, Khasanah N, Van Noordwijk M, Lamers JPA. Climate Change Sensitivity of Multi-Species Afforestation in Semi-Arid Benin. Sustainability. 2018; 10(6):1931. https://doi.org/10.3390/su10061931
Chicago/Turabian StyleNoulèkoun, Florent, Asia Khamzina, Jesse B. Naab, Ni’matul Khasanah, Meine Van Noordwijk, and John P. A. Lamers. 2018. "Climate Change Sensitivity of Multi-Species Afforestation in Semi-Arid Benin" Sustainability 10, no. 6: 1931. https://doi.org/10.3390/su10061931
APA StyleNoulèkoun, F., Khamzina, A., Naab, J. B., Khasanah, N., Van Noordwijk, M., & Lamers, J. P. A. (2018). Climate Change Sensitivity of Multi-Species Afforestation in Semi-Arid Benin. Sustainability, 10(6), 1931. https://doi.org/10.3390/su10061931