


Vestibular Testing—New Physiological Results for the Optimization of Clinical VEMP Stimuli


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Physiological Evidence—Overview
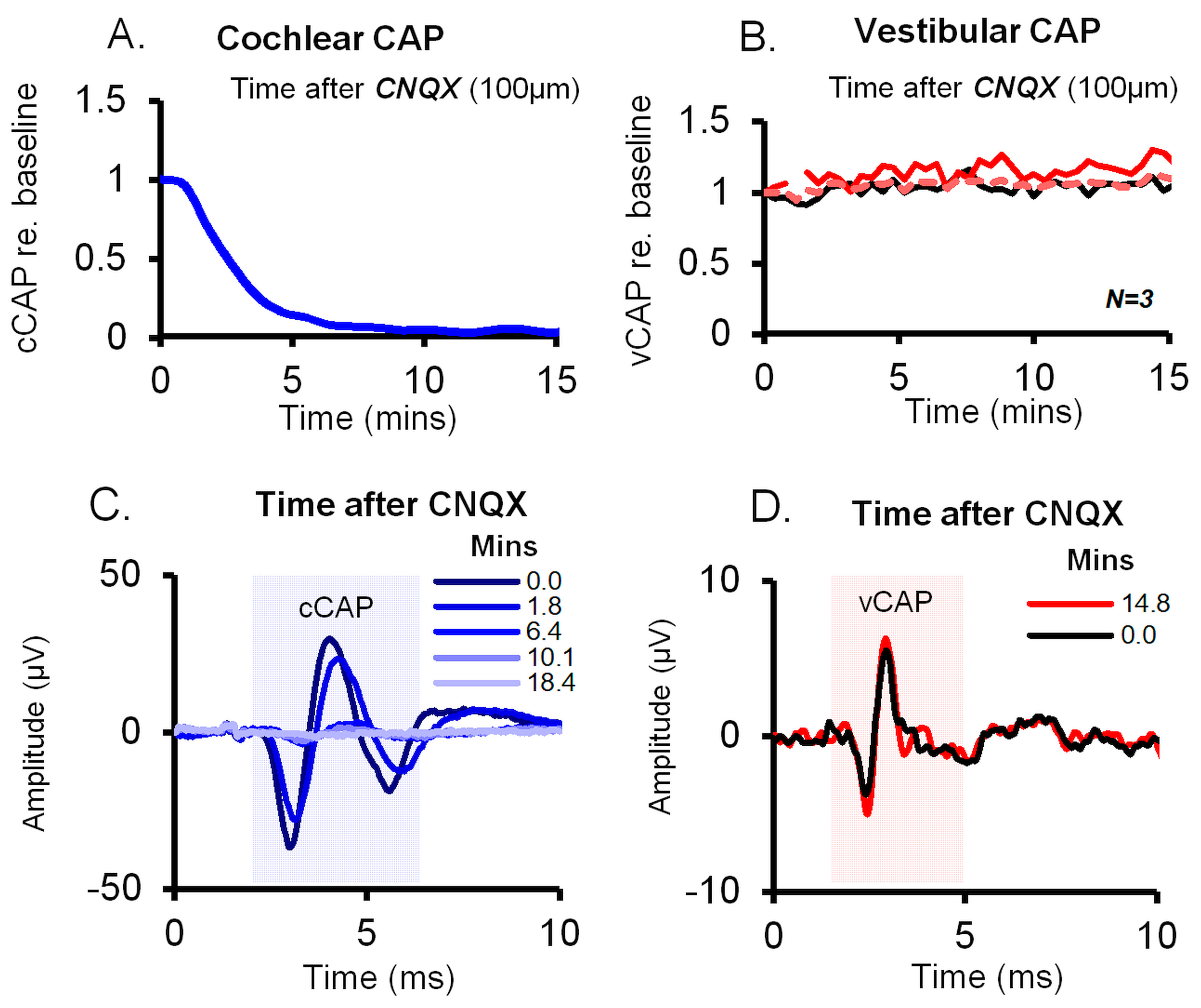
2.1. Independence of Cochlear and Vestibular Labyrinthine Divisions
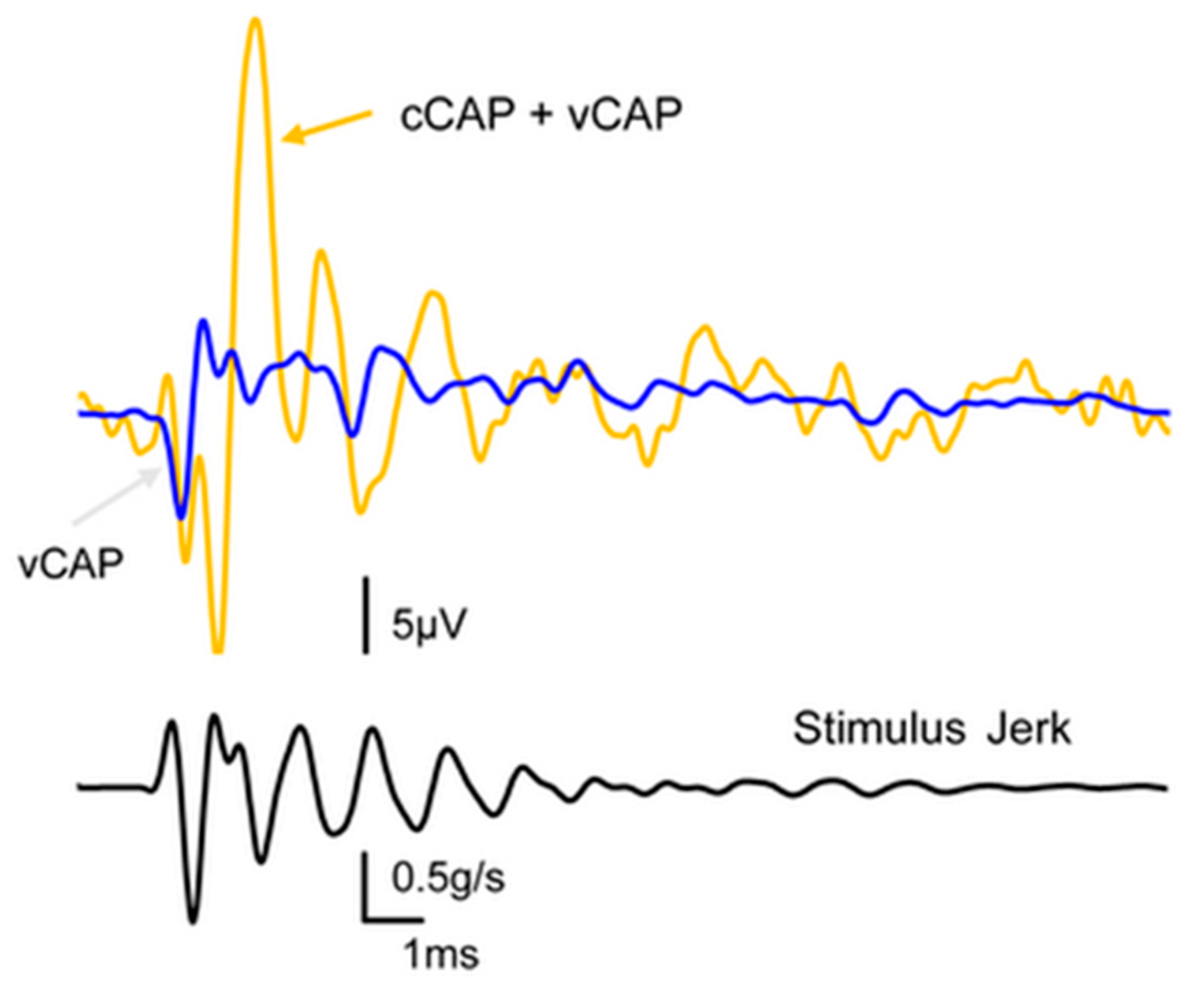
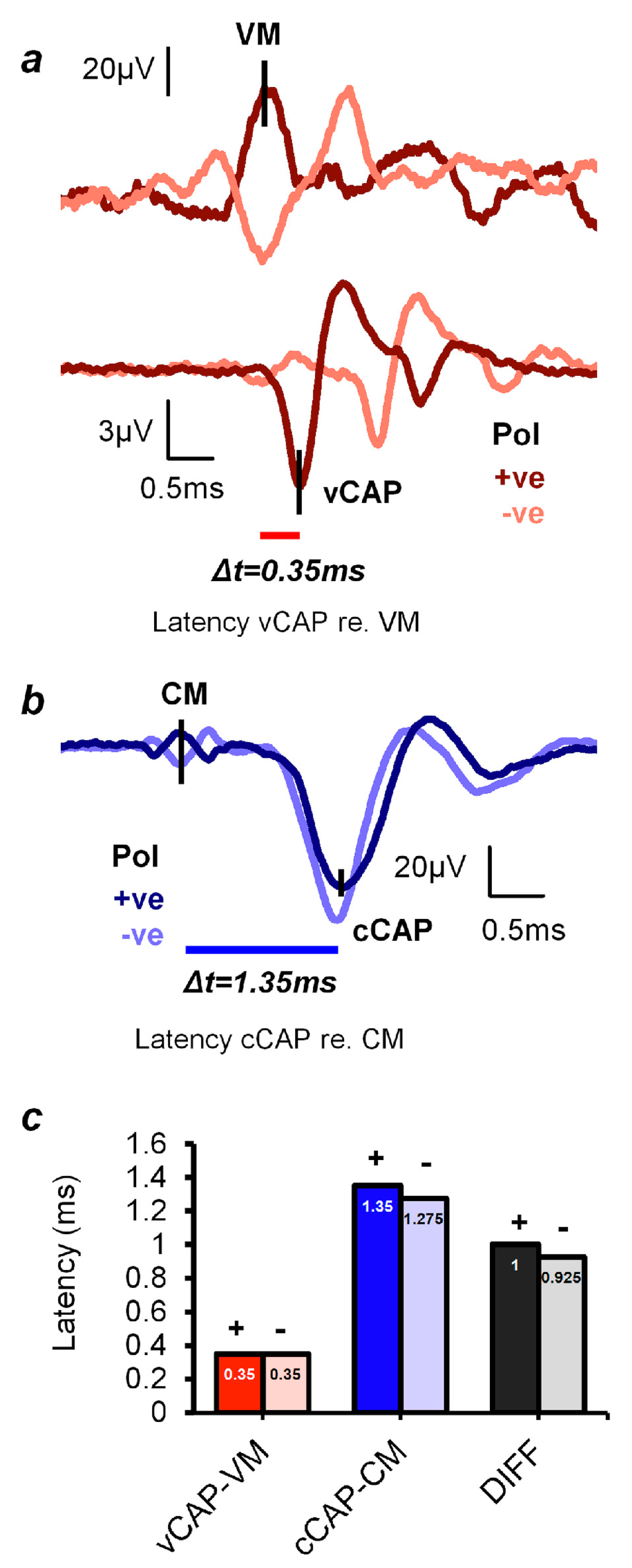
2.2. Differentiating Vestibular and Cochlear Compound Action Potentials
- (a)
- use stimuli that are accepted as otolithic—transient linear accelerations—and interpret the results (such as the VsEP) as being due to vestibular rather than cochlear activation [20]
- (b)
3. Anatomy
3.1. Cochlea
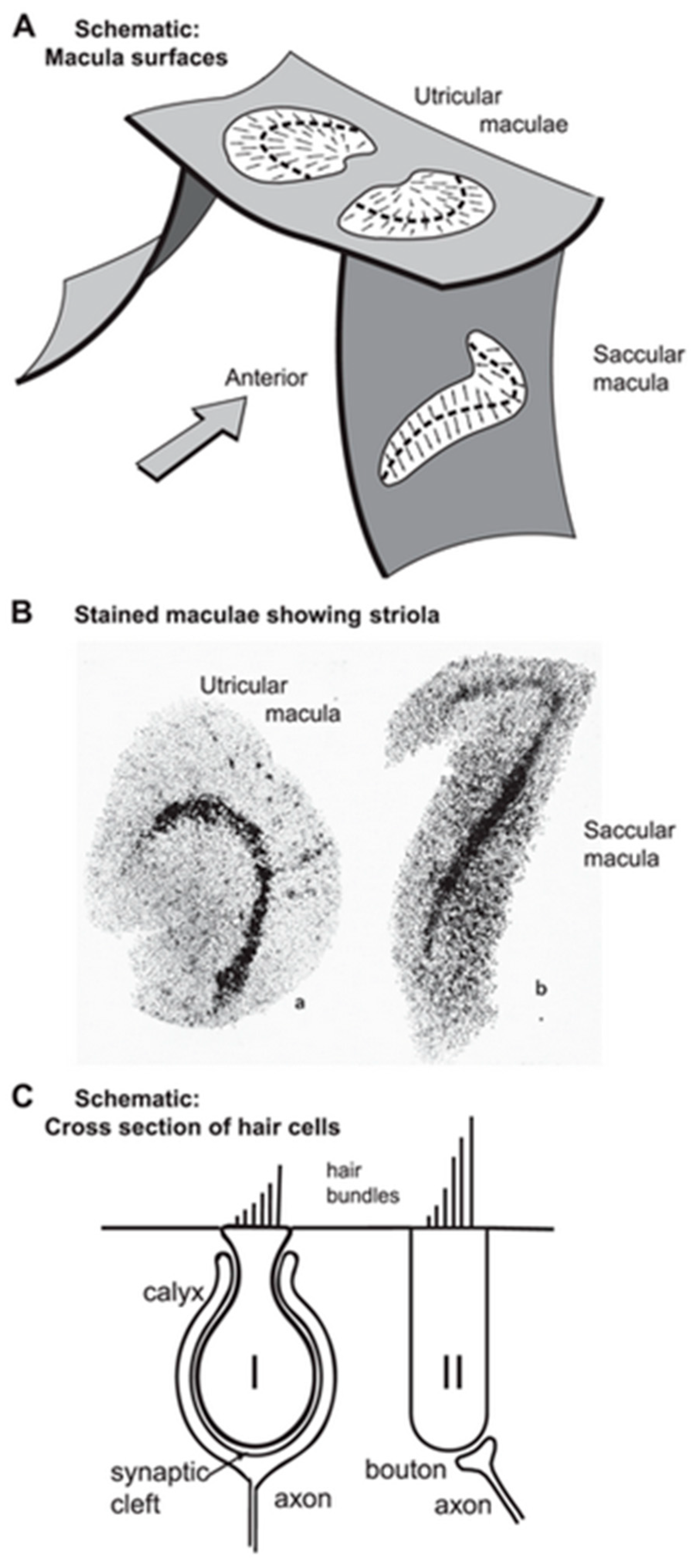
3.2. Vestibular


3.3. Specificity
3.4. Transmitter
- Glutamate release (quantal transmission);
- The level of potassium in the synaptic cleft;
- Resistive coupling between the type I receptor and the enveloping calyx (non-quantal transmission).
- (a)
- Glutamate release is apparently similar to that in the cochlea; vestibular type I receptors contain ribbon synapses, which release glutamate probably in response to relatively slow (sustained) stimuli. This is quantal glutamate transmission. The activation of the post-synaptic neuron by glutamatergic transmission is relatively slow [65].
- (b)
- Vestibular stimulation causes the deflection of the stereocilia and therefore opens the mechanoelectrical transduction (MET) channels on the stereocilia of type I receptors, so potassium enters the type I receptor from the potassium-rich endolymph and is released by the receptor into the narrow (femtolitre) synaptic cleft between the type I receptor and its enveloping calyx. Potassium levels in the narrow synaptic cleft modulate the membrane potential of the receptor and the calyx [64,67,68].
- (c)
- Most importantly, the simultaneous dual patch clamp recording of a type I receptor and its enveloping calyx afferent conclusively demonstrated non-quantal transmission experimentally. This is a form of synaptic transmission that is not dependent on glutamate reception on the post-synaptic membrane, and one key component of this non-quantal transmission is called resistive coupling, which is essentially electrical transmission that is dependent on membrane potential. This is a form of ultrafast electrical coupling between the type I receptor and calyx afferent; channels on the type I receptor membrane and the facing membrane of its calyx afferent are both open near resting potential, enabling the ultrafast depolarization of the irregular primary afferent [63,66]. We contend that this ultrafast non-quantal resistive coupling explains the precision of vestibular phase locking, the very short latency of the vCAP in comparison to the cCAP, and the extremely short latency of some irregular primary afferents to ACS clicks (0.5 ms) [69], and so we think resistive coupling is likely the key neural event in the generation of vCAP and therefore the generation of VEMPs
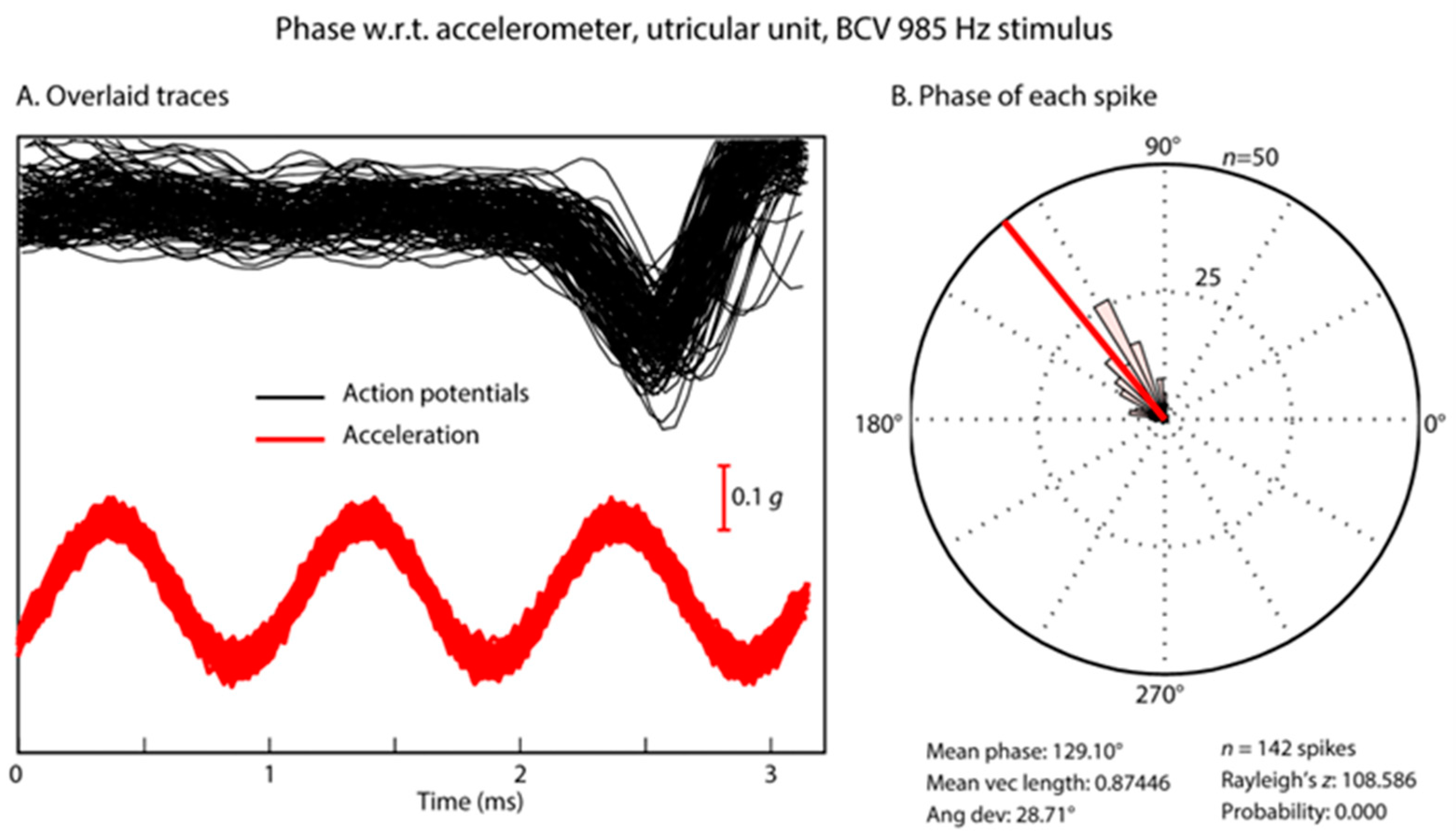
3.5. Temporal Precision of Irregular Vestibular Afferents—Latency
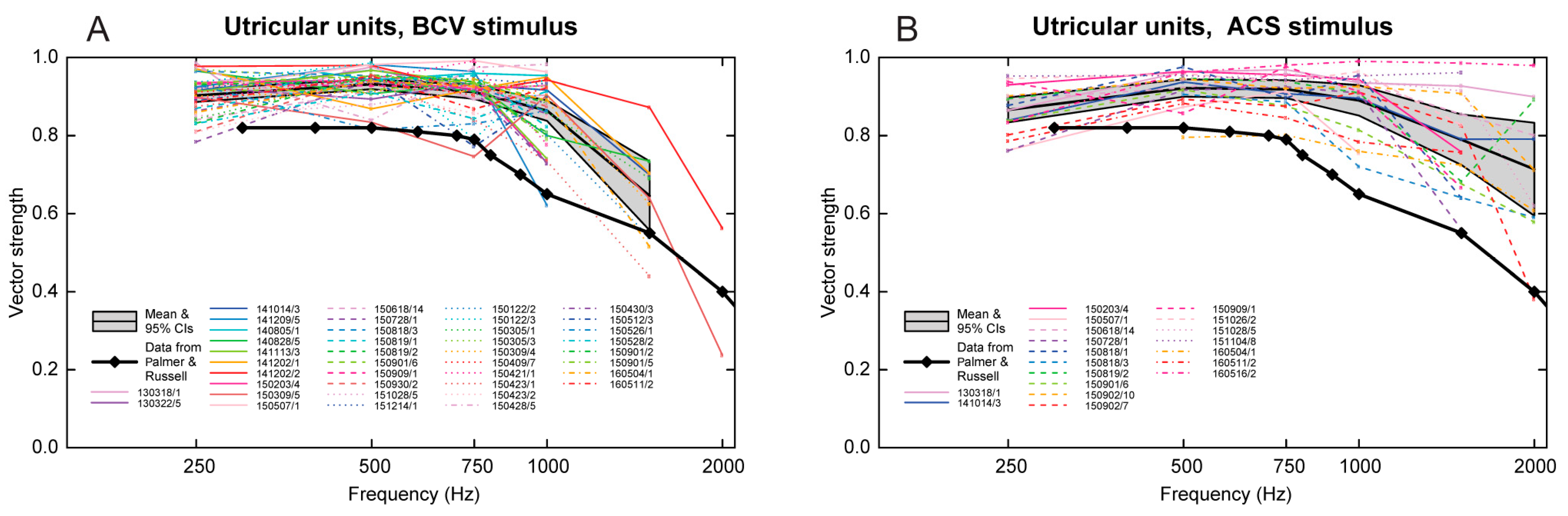
3.6. Temporal Precision of Irregular Vestibular Afferents—Phase Locking
4. Applications of Physiological Results to Clinical Vestibular (VEMP) Testing
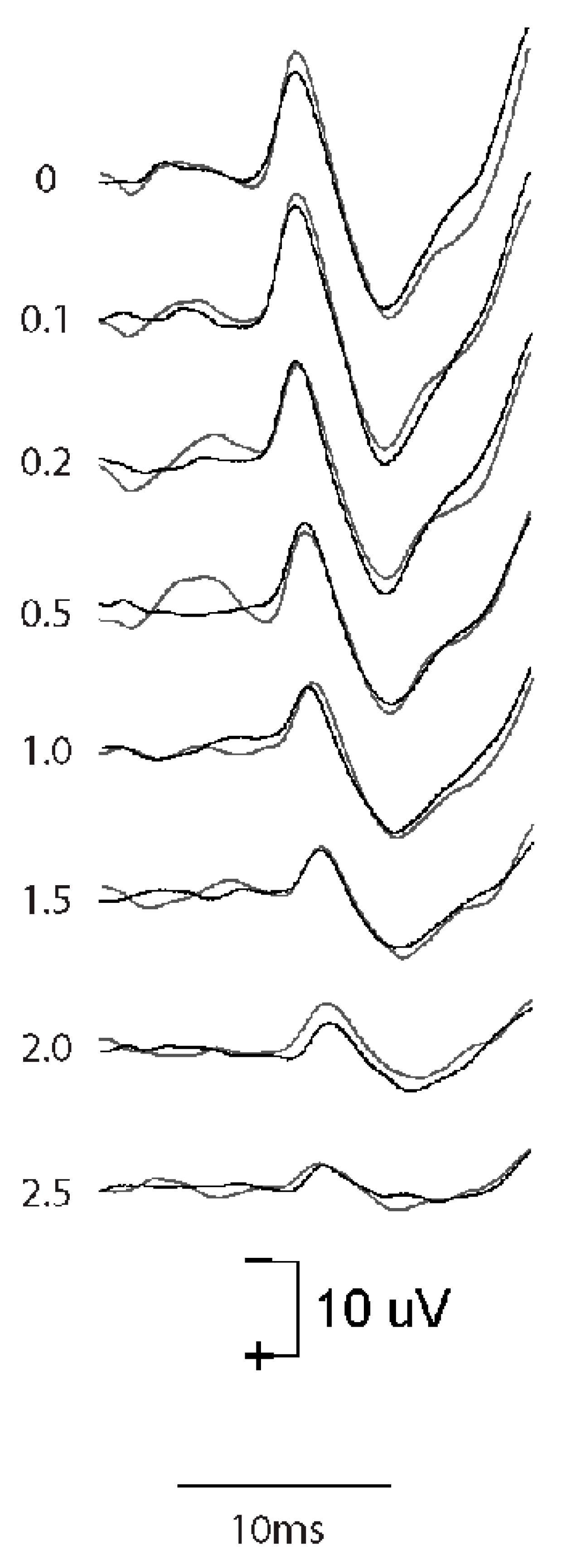
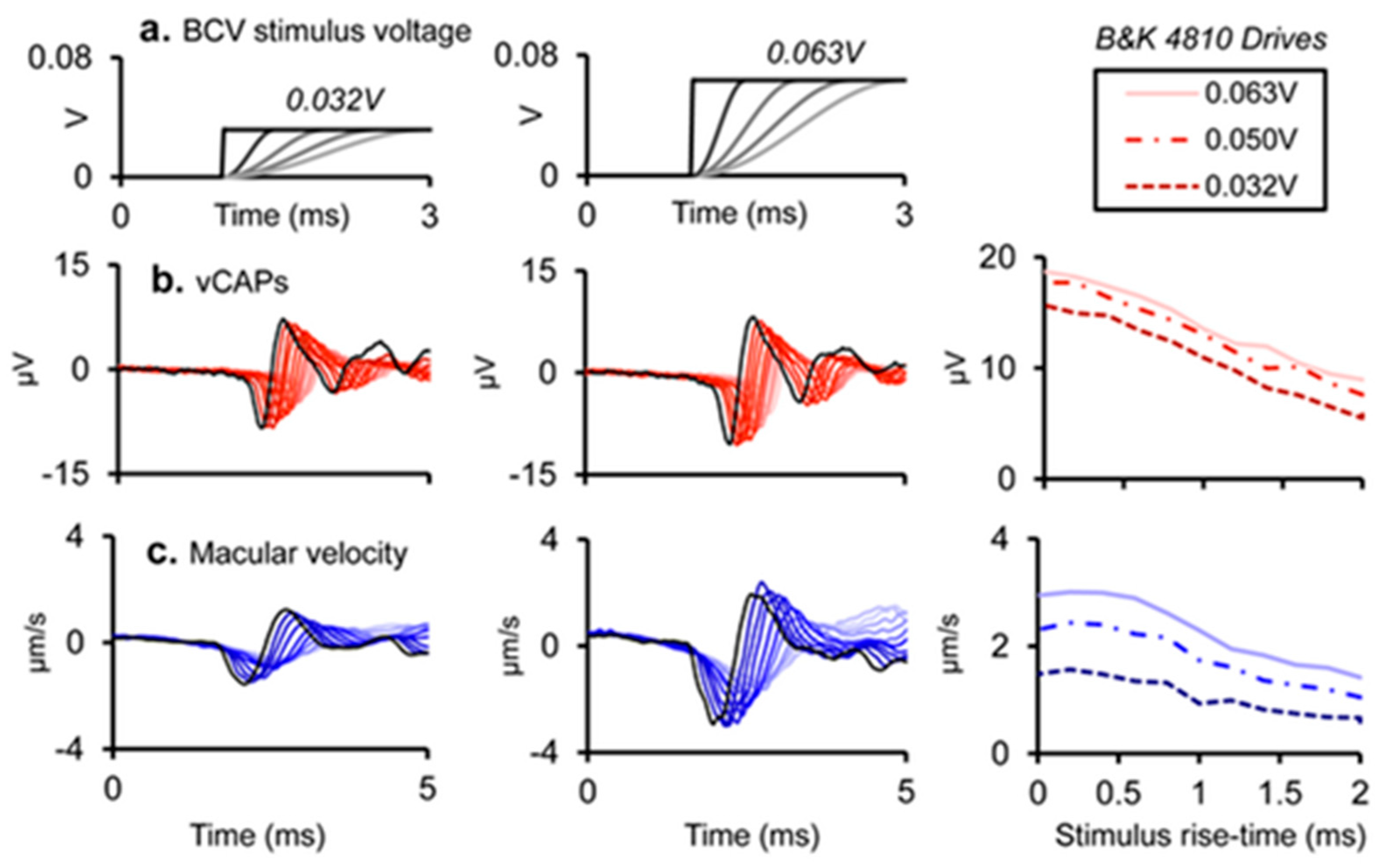
4.1. Effect of Rise Time
4.2. Masking
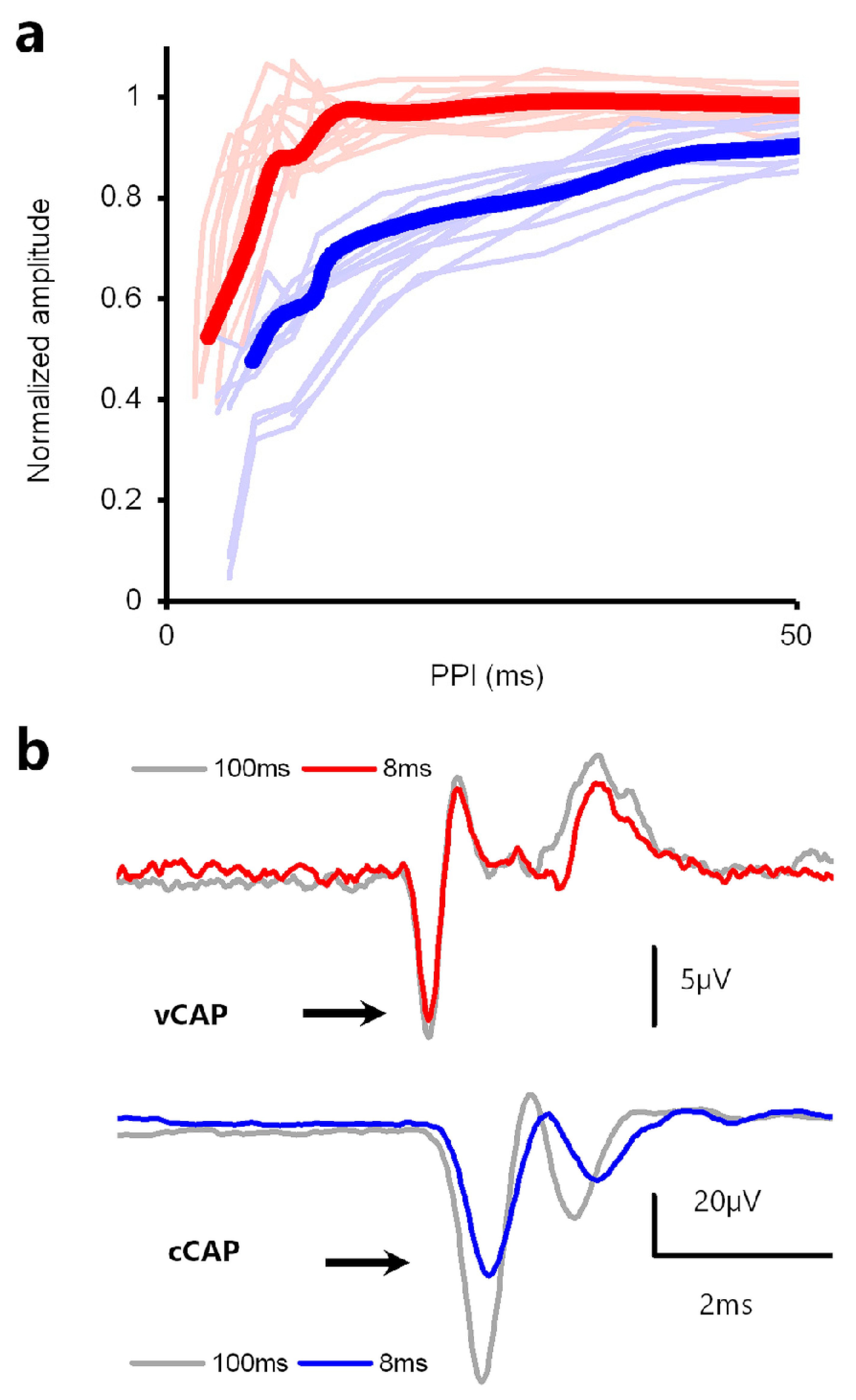
4.3. Paired Pulse Stimuli
5. Other Stimuli
6. Conclusions
7. Summary
- Vestibular receptors and afferents can function independently of the cochlea, both in humans and guinea pigs.
- It is possible to differentiate between the vestibular and cochlear responses to transient stimuli. Such differentiation provides further support to the basis of present vestibular testing, which usually involves using sound and vibration stimuli, and may allow new clinical tests of dynamic vestibular function. For future studies of putative vestibular responses to clinically realistic transient stimuli, it is advisable to have continuous broadband masking present simultaneously to minimize cochlear contributions in the response(s) [27].
- In clinical VEMP testing, the situation is exactly opposite to that in the audiometric testing of pure tone thresholds; effective VEMP stimuli have a very short rise time, synchronizing vestibular action potentials in primary afferent neurons. In contrast, for the audiometric testing of thresholds, a long rise time is mandatory. Unfortunately, some stimulus generators do not allow for rise times less than 2 ms, and some clinicians who measure VEMPs do not realize how important it is to minimize the rise time for VEMP testing as opposed to auditory threshold testing. Our recordings of the vCAP in response to chirp stimuli confirm that the chirp stimulus is particularly effective in generating vCAPs at the level of primary afferents in comparison to simple clicks.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Dedication
Abbreviations
| CAP | compound action potential |
| vCAP | vestibular compound action potential |
| cCAP | cochlear compound action potential |
| CM | cochlear microphonic |
| VM | vestibular microphonic |
| ANN | auditory nerve neurophonic |
| VNN | vestibular nerve neurophonic |
| ACS | air-conducted sound |
| BCV | bone-conducted vibration |
| SCD | semicircular canal dehiscence |
| VEMP | vestibular evoked myogenic potential |
| VsEP | vestibular evoked potential |
| oVEMP | ocular vestibular evoked myogenic potential |
| CNQX | 6-cyano-7-nitroquinoxaline-2,3-dione |
| MET | mechanoelectrical transduction |
| KCl | potassium chloride |
| PPI | paired pulse interval |
| LDV | laser Doppler vibrometer |
References
- Curthoys, I.S.; Grant, J.W.; Burgess, A.M.; Pastras, C.J.; Brown, D.J.; Manzari, L. Otolithic Receptor Mechanisms for Vestibular-Evoked Myogenic Potentials: A Review. Front. Neurol. 2018, 9, 366. [Google Scholar] [CrossRef] [PubMed]
- Rosengren, S.M.; Colebatch, J.G. The contributions of vestibular evoked myogenic potentials and acoustic vestibular stimulation to our understanding of the vestibular system. Front. Neurol. 2018, 9, 481. [Google Scholar] [CrossRef] [PubMed]
- Curthoys, I.S. The new vestibular stimuli: Sound and vibration-anatomical, physiological and clinical evidence. Exp. Brain Res. 2017, 235, 957–972. [Google Scholar] [CrossRef] [PubMed]
- Curthoys, I.S.; Vulovic, V.; Burgess, A.M.; Manzari, L.; Sokolic, L.; Pogson, J.; Robins, M.; Mezey, L.E.; Goonetilleke, S.; Cornell, E.D.; et al. Neural basis of new clinical vestibular tests: Otolithic neural responses to sound and vibration. Clin. Exp. Pharmacol. Physiol. 2014, 41, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Curthoys, I.S.; Grant, J.W.; Pastras, C.J.; Brown, D.J.; Burgess, A.M.; Brichta, A.M.; Lim, R. A review of mechanical and synaptic processes in otolith transduction of sound and vibration for clinical VEMP testing. J. Neurophysiol. (Bethesda) 2019, 122, 259–276. [Google Scholar] [CrossRef]
- Burgess, A.M.; Mezey, L.E.; Manzari, L.; Macdougall, H.G.; McGarvie, L.A.; Curthoys, I.S. Effect of Stimulus Rise-Time on the Ocular Vestibular-Evoked Myogenic Potential to Bone-Conducted Vibration. Ear Hear. 2013, 34, 799–805. [Google Scholar] [CrossRef]
- Curthoys, I.S. A critical review of the neurophysiological evidence underlying clinical vestibular testing using sound, vibration and galvanic stimuli. Clin. Neurophysiol. 2010, 121, 132–144. [Google Scholar] [CrossRef]
- Colebatch, J.G.; Rosengren, S.M. Investigating short latency subcortical vestibular projections in humans: What have we learned? J. Neurophysiol. 2019, 122, 2000–2015. [Google Scholar] [CrossRef]
- Rosengren, S.M.; Colebatch, J.G.; Young, A.S.; Govender, S.; Welgampola, M.S. Vestibular evoked myogenic potentials in practice: Methods, pitfalls and clinical applications. Clin. Neurophysiol. Pract. 2019, 4, 47–68. [Google Scholar] [CrossRef]
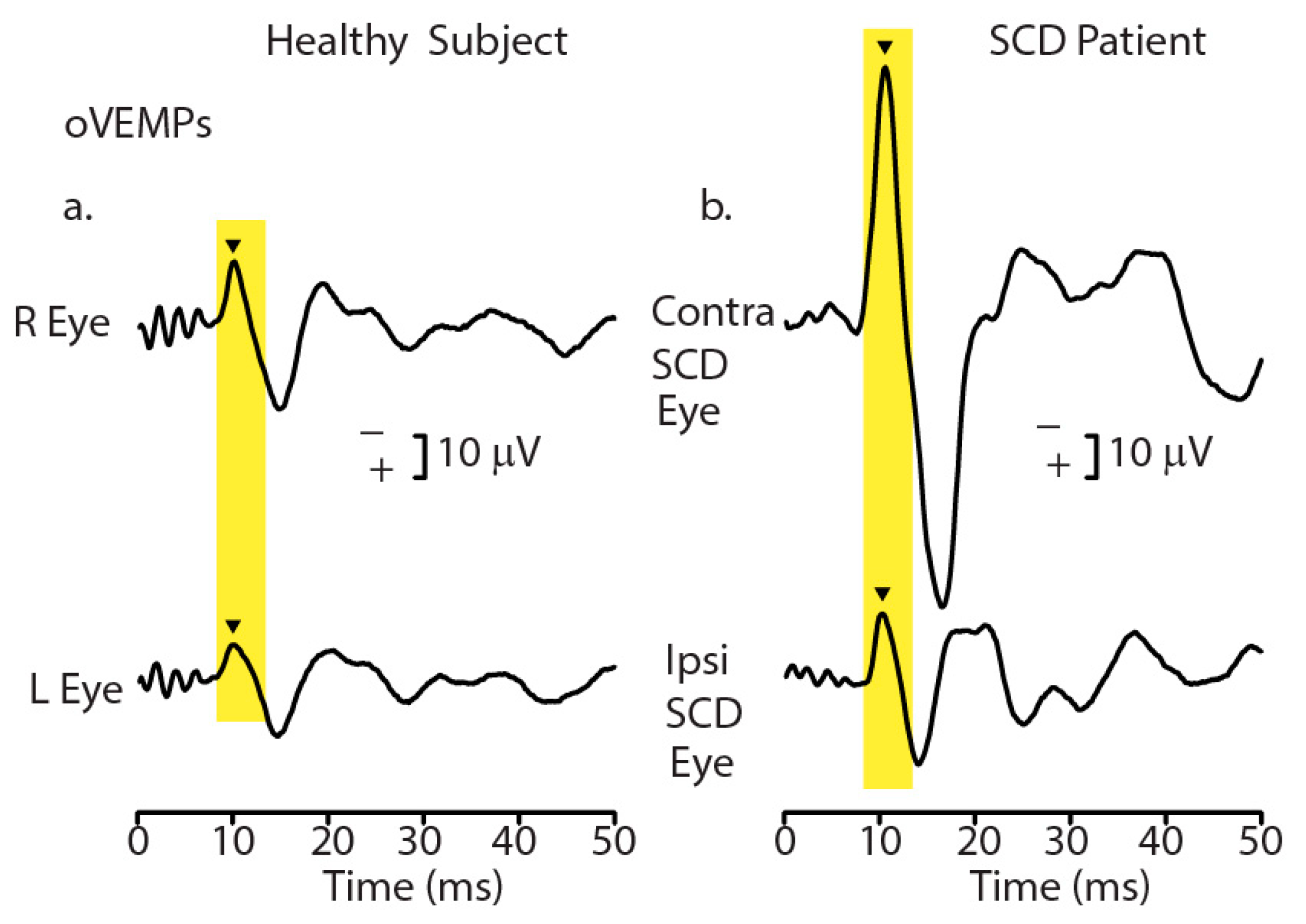
- Manzari, L.; Burgess, A.M.; McGarvie, L.A.; Curthoys, I.S. Ocular and cervical vestibular evoked myogenic potentials to 500 Hz fz bone-conducted vibration in superior semicircular canal dehiscence. Ear Hear. 2012, 33, 508–520. [Google Scholar] [CrossRef]
- Oh, S.Y.; Kim, H.J.; Kim, J.S. Vestibular-evoked myogenic potentials in central vestibular disorders. J. Neurol. 2016, 263, 210–220. [Google Scholar] [CrossRef]
- Pastras, C.J.; Curthoys, I.S.; Rabbitt, R.D.; Brown, D.J. Using macular velocity measurements to relate parameters of bone conduction to vestibular compound action potential responses. Sci. Rep. 2023, 13, 10204. [Google Scholar] [CrossRef]
- Pastras, C.J.; Gholami, N.; Jennings, S.; Zhu, H.; Zhou, W.; Brown, D.J.; Curthoys, I.S.; Rabbitt, R.D. A mathematical model for mechanical activation and compound action potential generation by the utricle in response to sound and vibration. Front. Neurol. 2023, 14, 1109506. [Google Scholar] [CrossRef]
- Pastras, C.J.; Curthoys, I.S.; Rabbitt, R.D.; Brown, D.J. Vestibular compound action potentials and macular velocity evoked by sound and vibration in the guinea pig. J. Neurosci. 2023, in press. [Google Scholar]
- Goldstein, M.H.; Kiang, N.Y.S. Synchrony of neural activity in electric responses evoked by transient acoustic stimuli. J. Acoust. Soc. Am. 1958, 30, 107–114. [Google Scholar] [CrossRef]
- Brown, D.J.; Pastras, C.J.; Curthoys, I.S. Electrophysiological measurements of peripheral vestibular function-a review of electrovestibulography. Front. Syst. Neurosci. 2017, 11, 14. [Google Scholar] [CrossRef] [PubMed]
- Pastras, C.J.; Stefani, S.P.; Camp, A.J.; Curthoys, I.S.; Brown, D.J. Summating potentials from the utricular macula of anaesthetized guinea pigs. Hear. Res. 2021, 406, 108259. [Google Scholar] [CrossRef] [PubMed]
- Bohmer, A.; Hoffman, L.F.; Honrubia, V. Characterization of vestibular potentials-evoked by linear acceleration pulses in the chinchilla. Am. J. Otol. 1995, 16, 498–504. [Google Scholar]
- Jones, T.A. Vestibular short latency responses to pulsed linear acceleration in unanesthetized animals. Electroencephalogr. Clin. Neurophysiol. 1992, 82, 377–386. [Google Scholar] [CrossRef]
- Jones, T.A.; Jones, S.M. Short latency compound action potentials from mammalian gravity receptor organs. Hear. Res. 1999, 136, 75–85. [Google Scholar] [CrossRef]
- Nazareth, A.M.; Jones, T.A. Central and peripheral components of short latency vestibular responses in the chicken. J. Vestib. Res. -Equilib. Orientat. 1998, 8, 233–252. [Google Scholar] [CrossRef]
- Jones, T.A.; Pedersen, T.L. Short latency vestibular responses to pulsed linear acceleration. Am. J. Otolaryngol. 1989, 10, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Elidan, J.; Langhofer, L.; Honrubia, V. Recording of short-latency vestibular evoked-potentials induced by acceleration impulses in experimental-animals—Current status of the method and its applications. Electroencephalogr. Clin. Neurophysiol. 1987, 68, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Rutherford, M.A.; von Gersdorff, H.; Goutman, J.D. Encoding sound in the cochlea: From receptor potential to afferent discharge. J. Physiol. Lond. 2021, 599, 2527–2557. [Google Scholar] [CrossRef]
- Plontke, S.K.; Rahne, T.; Curthoys, I.S.; Håkansson, B.; Fröhlich, L. A case series shows independent vestibular labyrinthine function after major surgical trauma to the human cochlea. Commun. Med. (Lond) 2021, 1, 37. [Google Scholar] [CrossRef]
- Jones, T.A.; Jones, S.M.; Vijayakumar, S.; Brugeaud, A.; Bothwell, M.; Chabbert, C. The adequate stimulus for mammalian linear vestibular evoked potentials (VsEPs). Hear. Res. 2011, 280, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.M.; Robertson, N.G.; Given, S.; Giersch, A.B.S.; Liberman, M.C.; Morton, C.C. Hearing and vestibular deficits in the Coch(-/-) null mouse model: Comparison to the Coch(G88E/G88E) mouse and to DFNA9 hearing and balance disorder. Hear. Res. 2011, 272, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Martin, P.; Hudspeth, A.J. Mechanical Frequency Tuning by Sensory Hair Cells, the Receptors and Amplifiers of the Inner Ear. Annu. Rev. Condens. Matter Phys 2021, 12, 29–49. [Google Scholar]
- Caprara, G.A.; Peng, A.W. Mechanotransduction in mammalian sensory hair cells. Mol. Cell. Neurosci. 2022, 120, 103706. [Google Scholar] [CrossRef]
- Littman, T.; Bobbin, R.P.; Fallon, M.; Puel, J.L. The quinoxalinediones DNQX, CNQX and 2 related congeners suppress hair cell to auditory nerve transmission. Hear. Res. 1989, 40, 45–53. [Google Scholar] [CrossRef]
- Doi, K.; Mori, N.; Matsunaga, T.; Tsumoto, T. Blockade of synaptic transmission from hair-cells to auditory afferent nerves by 6-cyano-2,3-dihydroxy-7-nitroquinoxaline, a selective non-nmda receptor antagonist. Eur. Arch. Oto-Rhino-Laryngol. 1990, 248, 25–30. [Google Scholar] [CrossRef]
- Eatock, R.A.; Songer, J.E. Vestibular hair cells and afferents: Two channels for head motion signals. Annu. Rev. Neurosci. 2011, 34, 501–534. [Google Scholar] [CrossRef] [PubMed]
- Heil, P.; Peterson, A.J. Spike timing in auditory-nerve fibers during spontaneous activity and phase locking. Synapse 2017, 71, 5–36. [Google Scholar] [CrossRef] [PubMed]
- Li, G.L.; Cho, S.; von Gersdorff, H. Phase-Locking Precision Is Enhanced by Multiquantal Release at an Auditory Hair Cell Ribbon Synapse. Neuron 2014, 83, 1404–1417. [Google Scholar] [CrossRef] [PubMed]
- Rose, J.E.; Brugge, J.F.; Anderson, D.J.; Hind, J.E. Phase-locked response to low-frequency tones in single auditory nerve fibers of squirrel monkey. J. Neurophysiol. 1967, 30, 769. [Google Scholar] [CrossRef]
- Fettiplace, R. Hair Cell Transduction, Tuning, and Synaptic Transmission in the Mammalian Cochlea. Compr. Physiol. 2017, 7, 1197–1227. [Google Scholar] [CrossRef]
- Palmer, A.R.; Russell, I.J. Phase-locking in the cochlear nerve of the guinea-pig and its relation to the receptor potential of inner hair-cells. Hear. Res. 1986, 24, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, J.M. Afferent diversity and the organization of central vestibular pathways. Exp. Brain Res. 2000, 130, 277–297. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Xue, J.; Peterson, E.H. Architecture of the mouse utricle: Macular organization and hair bundle heights. J. Neurophysiol. 2008, 99, 718–733. [Google Scholar] [CrossRef] [PubMed]
- Nam, J.H.; Cotton, J.R.; Peterson, E.H.; Grant, W. Mechanical properties and consequences of stereocilia and extracellular links in vestibular hair bundles. Biophys. J. 2006, 90, 2786–2795. [Google Scholar] [CrossRef]
- Goldberg, J.M.; Desmadryl, G.; Baird, R.A.; Fernandez, C. The vestibular nerve of the chinchilla. V. Relation between afferent discharge properties and peripheral innervation patterns in the utricular macula. J. Neurophysiol. 1990, 63, 791–804. [Google Scholar] [CrossRef]
- Watanuki, K. Meyer zum Gottesberge. A. Morphological observations of sensory epithelium of macula sacculi and utriculi in guinea pig. Arch. Fur Klin. Und Exp. Ohren-Nasen-Und Kehlkopfheilkd. 1971, 200, 136. [Google Scholar] [CrossRef] [PubMed]
- Watanuki, K.; Schuknecht, H.F. Morphological-study of human vestibular sensory epithelia. Arch. Otolaryngol. Head. Neck Surg. 1976, 102, 583–588. [Google Scholar] [CrossRef]
- Eatock, R.A. Adaptation in hair cells. Annu. Rev. Neurosci. 2000, 23, 285–314. [Google Scholar] [CrossRef] [PubMed]
- Lysakowski, A. Synaptic organization of the crista ampullaris in vertebrates. In New Directions in Vestibular Research; Highstein, S.M., Cohen, B., ButtnerEnnever, J.A., Eds.; Annals of the New York Academy of Sciences: New York, NY, USA, 1996; Volume 781, pp. 164–182. [Google Scholar]
- Lysakowski, A.; Goldberg, J.M. A regional ultrastructural analysis of the cellular and synaptic architecture in the chinchilla cristae ampullares. J. Comp. Neurol. 1997, 389, 419–443. [Google Scholar] [CrossRef]
- Hudspeth, A.J. Micromechanics of Hearing. In Mechanics of Hearing: Protein to Perception; Karavitaki, K.D., Corey, D.P., Eds.; AIP Conference Proceedings, American Institute of Physics: Melville, NY, USA, 2015; Volume 1703. [Google Scholar]
- Pastras, C.J.; Curthoys, I.S.; Brown, D.J. In vivo recording of the vestibular microphonic in mammals. Hear. Res. 2017, 354, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Curthoys, I.S.; Vulovic, V.; Sokolic, L.; Pogson, J.; Burgess, A.M. Irregular primary otolith afferents from the guinea pig utricular and saccular maculae respond to both bone conducted vibration and to air conducted sound. Brain Res. Bull. 2012, 89, 16–21. [Google Scholar] [CrossRef]
- Zhu, H.; Tang, X.; Wei, W.; Mustain, W.; Xu, Y.; Zhou, W. Click-evoked responses in vestibular afferents in rats. J. Neurophysiol. 2011, 106, 754–763. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, J.M.; Desmadryl, G.; Baird, R.A.; Fernandez, C. The vestibular nerve of the chinchilla. IV. Discharge properties of utricular afferents. J. Neurophysiol. 1990, 63, 781–790. [Google Scholar] [CrossRef]
- Fernandez, C.; Baird, R.A.; Goldberg, J.M. The vestibular nerve of the chinchilla. I. Peripheral innervation patterns in the horizontal and superior semicircular canals. J. Neurophysiol. 1988, 60, 167–181. [Google Scholar] [CrossRef]
- Curthoys, I.S.; Burgess, A.M.; Goonetilleke, S.C. Phase-locking of irregular guinea pig primary vestibular afferents to high frequency (> 250 Hz) sound and vibration. Hear. Res. 2019, 373, 59–70. [Google Scholar] [CrossRef]
- Curthoys, I.S.; Vulovic, V.; Burgess, A.M.; Sokolic, L.; Goonetilleke, S.C. The response of guinea pig primary utricular and saccular irregular neurons to bone-conducted vibration (BCV) and air-conducted, sound (ACS). Hear. Res. 2016, 331, 131–143. [Google Scholar] [CrossRef]
- Goldberg, J.M.; Lysakowski, A.; Fernandez, C. Structure and function of vestibular nerve fibers in the chinchilla and squirrel monkey. Ann. N. Y. Acad. Sci. 1992, 656, 92–107. [Google Scholar] [CrossRef]
- Mukhopadhyay, M.; Pangrsic, T. Synaptic transmission at the vestibular hair cells of amniotes. Mol. Cell. Neurosci. 2022, 121, 103749. [Google Scholar] [CrossRef]
- Pastras, C.J.; Curthoys, I.S.; Brown, D.J. Dynamic response to sound and vibration of the guinea pig utricular macula, measured in vivo using Laser Doppler Vibrometry. Hear. Res. 2018, 370, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Watanuki, K.; Meyer zum Gottesberge, A. Morphological study of sensory epithelium of vestibular organs. Tohoku J. Exp. Med. 1971, 104, 55–63. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pastras, C.J.; Curthoys, I.S.; Asadnia, M.; McAlpine, D.; Rabbitt, R.D.; Broen, D.J. Evidence that ultrafast non-quantal transmission underlies synchronized vestibular action potential generation. J. Neurosci. Press 2023, 43, 7149–7157. [Google Scholar] [CrossRef]
- Tasaki, I. Nerve impulses in individual auditory nerve fibers of guinea pig. J. Neurophysiol. 1954, 17, 97–122. [Google Scholar] [CrossRef]
- Rosowski, J.J.; Songer, J.E.; Nakajima, H.H.; Brinsko, K.M.; Merchant, S.N. Clinical, experimental, and theoretical investigations of the effect of superior semicircular canal dehiscence on hearing mechanisms. Otol. Neurotol. 2004, 25, 323–332. [Google Scholar] [CrossRef]
- Dlugaiczyk, J.; Burgess, A.M.; Curthoys, I.S. Activation of Guinea Pig Irregular Semicircular Canal Afferents by 100 Hz Vibration: Clinical Implications for Vibration-induced Nystagmus and Vestibular-evoked Myogenic Potentials. Otol. Neurotol. 2020, 41, E961–E970. [Google Scholar] [CrossRef]
- Contini, D.; Holstein, G.R.; Art, J.J. Synaptic cleft microenvironment influences potassium permeation and synaptic transmission in hair cells surrounded by calyx afferents in the turtle. J. Physiol. Lond. 2020, 598, 853–889. [Google Scholar] [CrossRef]
- Contini, D.; Holstein, G.R.; Art, J.J. Simultaneous Dual Recordings From Vestibular Hair Cells and Their Calyx Afferents Demonstrate Multiple Modes of Transmission at These Specialized Endings. Front. Neurol. 2022, 13, 891536. [Google Scholar] [CrossRef]
- Songer, J.E.; Eatock, R.A. Tuning and Timing in Mammalian Type I Hair Cells and Calyceal Synapses. J. Neurosci. 2013, 33, 3706–3724. [Google Scholar] [CrossRef] [PubMed]
- Govindaraju, A.C.; Quraishi, I.H.; Lysakowski, A.; Eatock, R.A.; Raphael, R.M. Nonquantal transmission at the vestibular hair cell-calyx synapse: KLV currents modulate fast electrical and slow K+ potentials. Proc. Natl. Acad. Sci. USA 2023, 120, e2207466120. [Google Scholar] [CrossRef] [PubMed]
- Lim, R.; Kindig, A.E.; Donne, S.W.; Callister, R.J.; Brichta, A.M. Potassium accumulation between type I hair cells and calyx terminals in mouse crista. Exp. Brain Res. 2011, 210, 607–621. [Google Scholar] [CrossRef] [PubMed]
- Contini, D.; Price, S.D.; Art, J.J. Accumulation of K+ in the synaptic cleft modulates activity by influencing both vestibular hair cell and calyx afferent in the turtle. J. Physiol. Lond. 2017, 595, 777–803. [Google Scholar] [CrossRef]
- Murofushi, T.; Curthoys, I.S.; Topple, A.N.; Colebatch, J.G.; Halmagyi, G.M. Responses of guinea pig primary vestibular neurons to clicks. Exp. Brain Res. 1995, 103, 174–178. [Google Scholar] [CrossRef]
- Spaiardi, P.; Tavazzani, E.; Manca, M.; Russo, G.; Prigioni, I.; Biella, G.; Giunta, R.; Johnson, S.L.; Marcotti, W.; Masetto, S. K(+)Accumulation and Clearance in the Calyx Synaptic Cleft of Type I Mouse Vestibular Hair Cells. Neuroscience 2020, 426, 69–86. [Google Scholar] [CrossRef]
- Young, E.D.; Fernandez, C.; Goldberg, J.M. Responses of squirrel monkey vestibular neurons to audio-frequency sound and head vibration. Acta Otolaryngol. 1977, 84, 352–360. [Google Scholar] [CrossRef]
- Peterson, A.J.; Heil, P. Phase Locking of Auditory Nerve Fibers: The Role of Lowpass Filtering by Hair Cells. J. Neurosci. 2020, 40, 4700–4714. [Google Scholar] [CrossRef]
- Lim, L.J.Z.; Dennis, D.L.; Govender, S.; Colebatch, J.G. Differential effects of duration for ocular and cervical vestibular evoked myogenic potentials evoked by air- and bone-conducted stimuli. Exp. Brain Res. 2013, 224, 437–445. [Google Scholar] [CrossRef]
- Curthoys, I.S.; Grant, J.W.; Pastras, C.J.; Frohlich, L.; Brown, D.J. Similarities and differences between vestibular and cochlear systems—A review of clinical and physiological evidence. Front. Neurosci. 2021, 15, 695179. [Google Scholar] [CrossRef] [PubMed]
- Curthoys, I.S.; Kim, J.; McPhedran, S.K.; Camp, A.J. Bone conducted vibration selectively activates irregular primary otolithic vestibular neurons in the guinea pig. Exp. Brain Res. 2006, 175, 256–267. [Google Scholar] [CrossRef]
- Parham, K.; Zhao, H.B.; Kim, D.O. Responses of auditory nerve fibers of the unanesthetized decerebrate cat to click pairs as simulated echoes. J. Neurophysiol. 1996, 76, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Wickesberg, R.E.; Stevens, H.E. Responses of auditory nerve fibers to trains of clicks. J. Acoust. Soc. Am. 1998, 103, 1990–1999. [Google Scholar] [CrossRef] [PubMed]
- Henry, K.R. Auditory-nerve neurophonic recorded from the round window of the mongolian gerbil. Hear. Res. 1995, 90, 176–184. [Google Scholar] [CrossRef]
- Walther, L.E.; Cebulla, M. Band limited chirp stimulation in vestibular evoked myogenic potentials. Eur. Arch. Oto-Rhino-Laryngol. 2016, 273, 2983–2991. [Google Scholar] [CrossRef]
- Reddy, T.M.; Heinze, B.; Biagio-de Jager, L.; Maes, L. Cervical and ocular vestibular evoked myogenic potential: A comparison of narrowband chirp, broadband chirp, tone burst and click stimulation. Int. J. Audiol. 2023, 62, 579–586. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pastras, C.J.; Curthoys, I.S. Vestibular Testing—New Physiological Results for the Optimization of Clinical VEMP Stimuli. Audiol. Res. 2023, 13, 910-928. https://doi.org/10.3390/audiolres13060079
Pastras CJ, Curthoys IS. Vestibular Testing—New Physiological Results for the Optimization of Clinical VEMP Stimuli. Audiology Research. 2023; 13(6):910-928. https://doi.org/10.3390/audiolres13060079
Chicago/Turabian StylePastras, Christopher J., and Ian S. Curthoys. 2023. "Vestibular Testing—New Physiological Results for the Optimization of Clinical VEMP Stimuli" Audiology Research 13, no. 6: 910-928. https://doi.org/10.3390/audiolres13060079
APA StylePastras, C. J., & Curthoys, I. S. (2023). Vestibular Testing—New Physiological Results for the Optimization of Clinical VEMP Stimuli. Audiology Research, 13(6), 910-928. https://doi.org/10.3390/audiolres13060079