
Exploring Microbial Diversity in Forest Litter-Based Fermented Bioproducts and Their Effects on Tomato (Solanum lycopersicum L.) Growth in Senegal

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
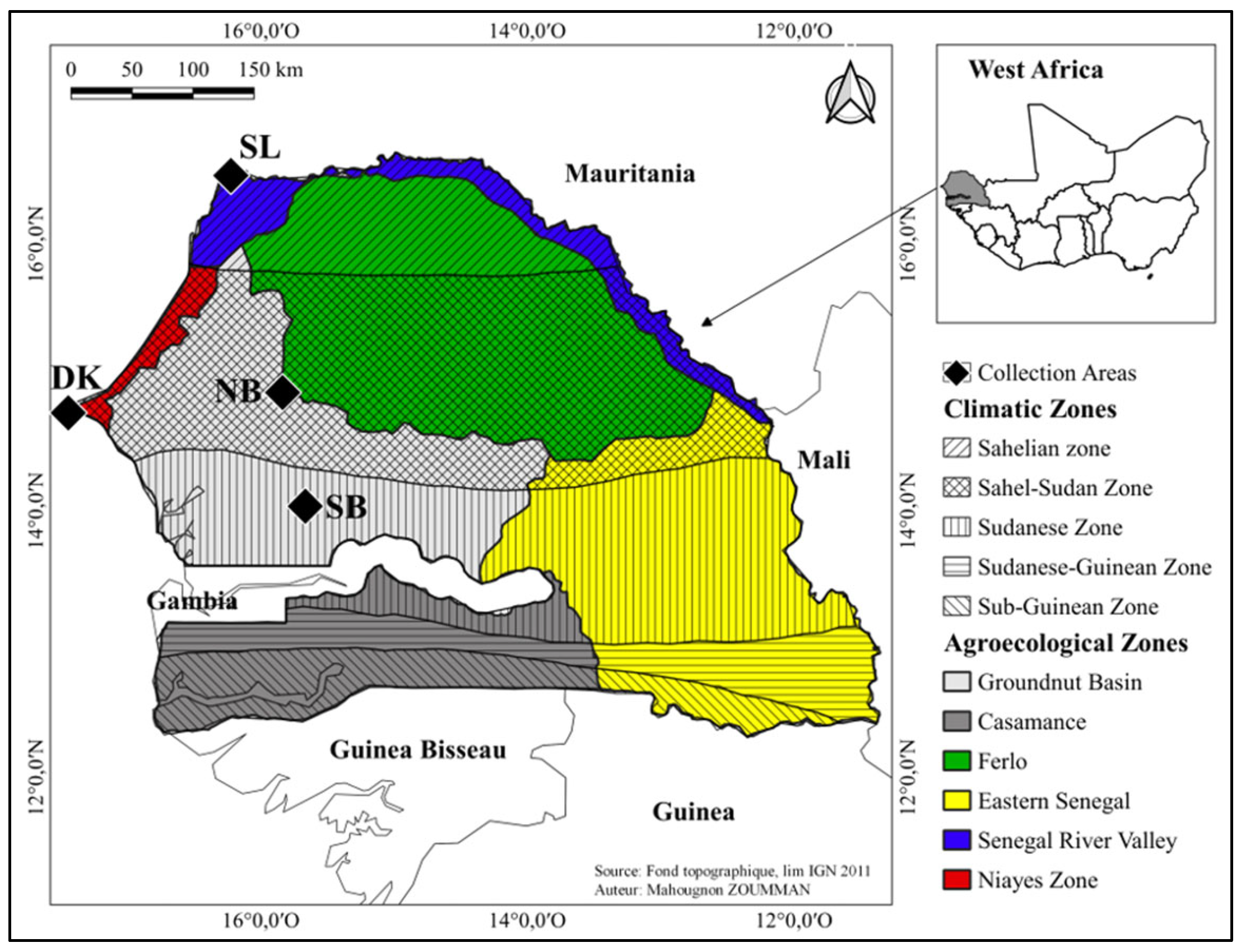
2.1. Litter—Collection and the Local Fermented Forest Litter Preparation
2.2. DNA Extraction and Sequencing
2.3. Effect of BMs Product on Tomato
3. Results
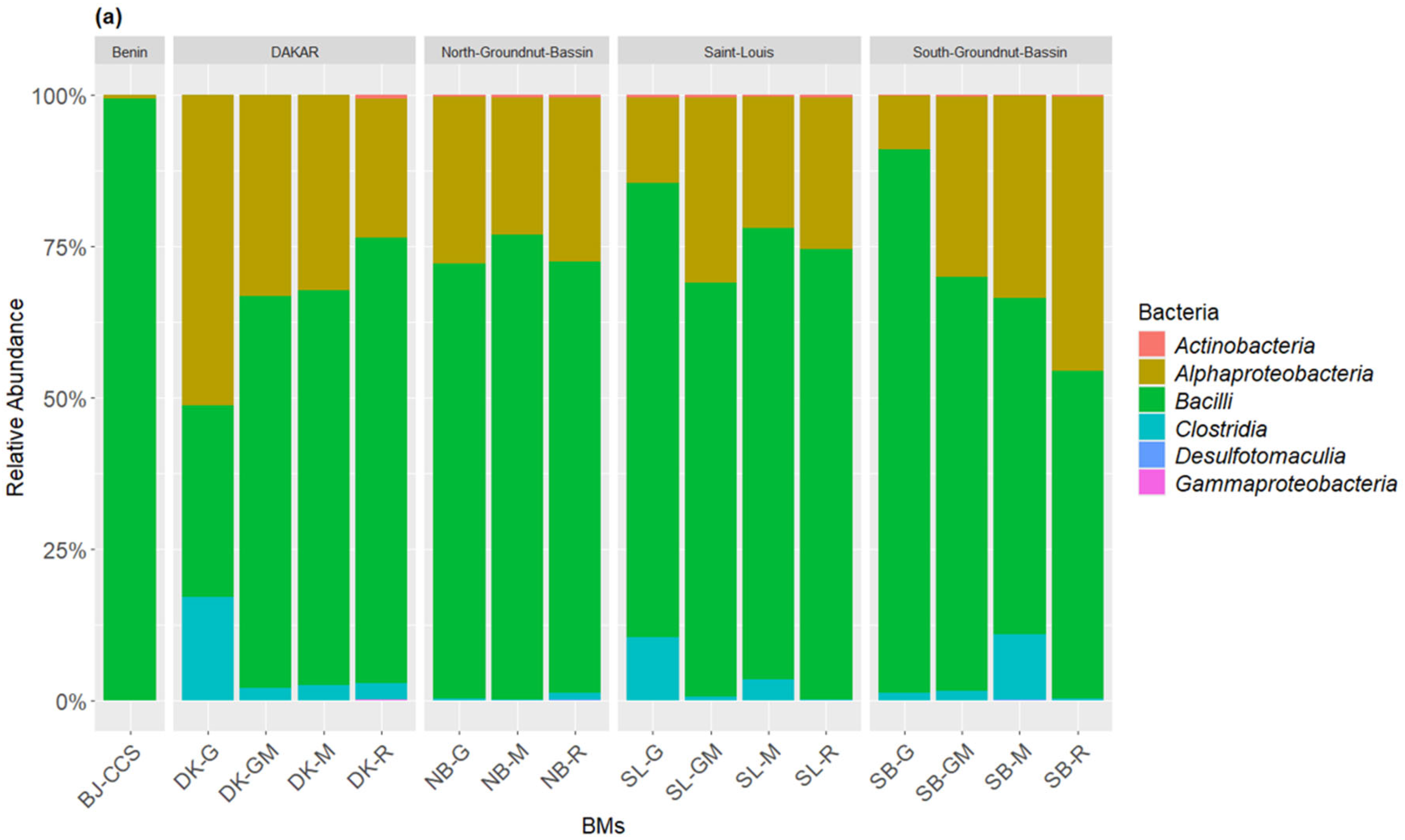
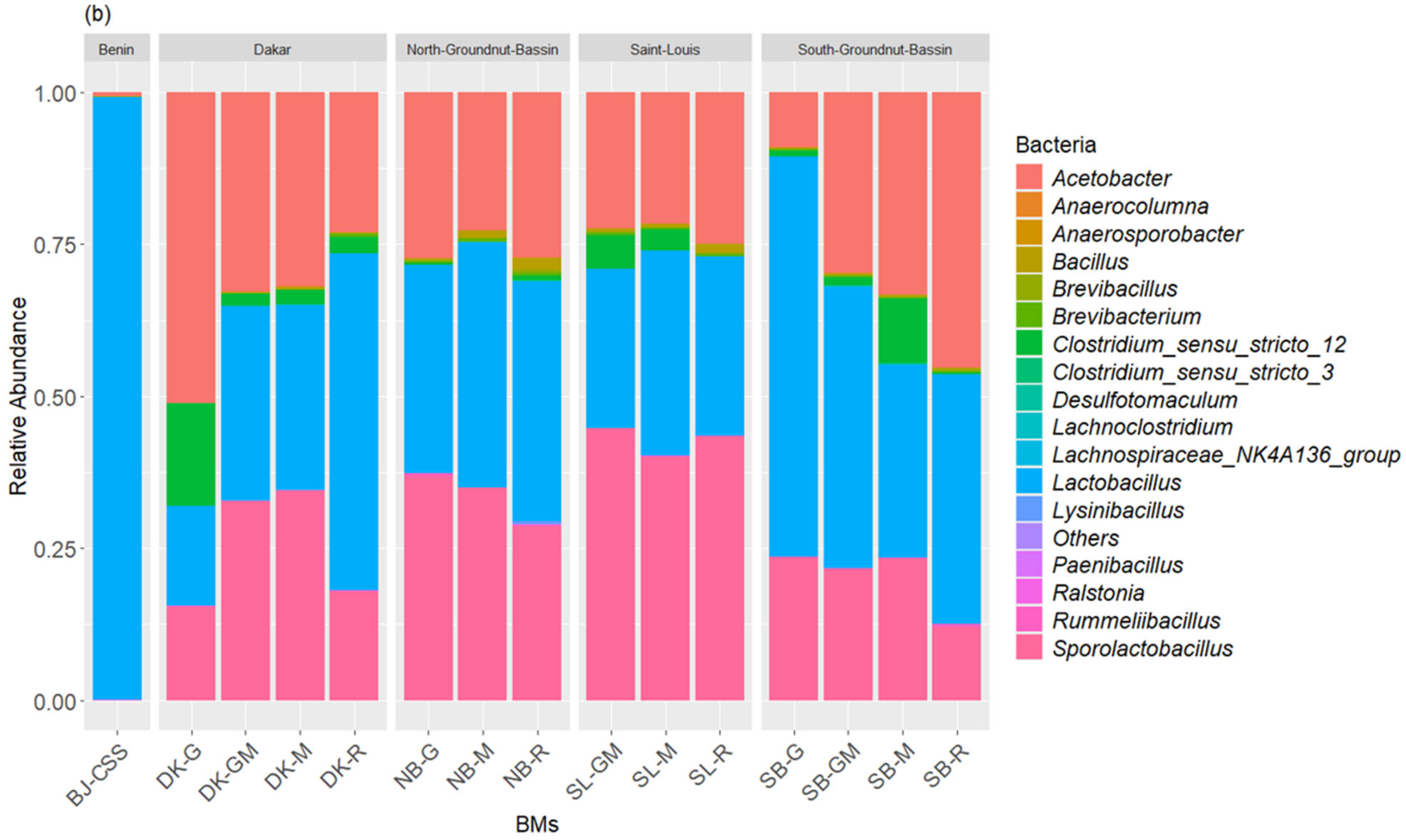
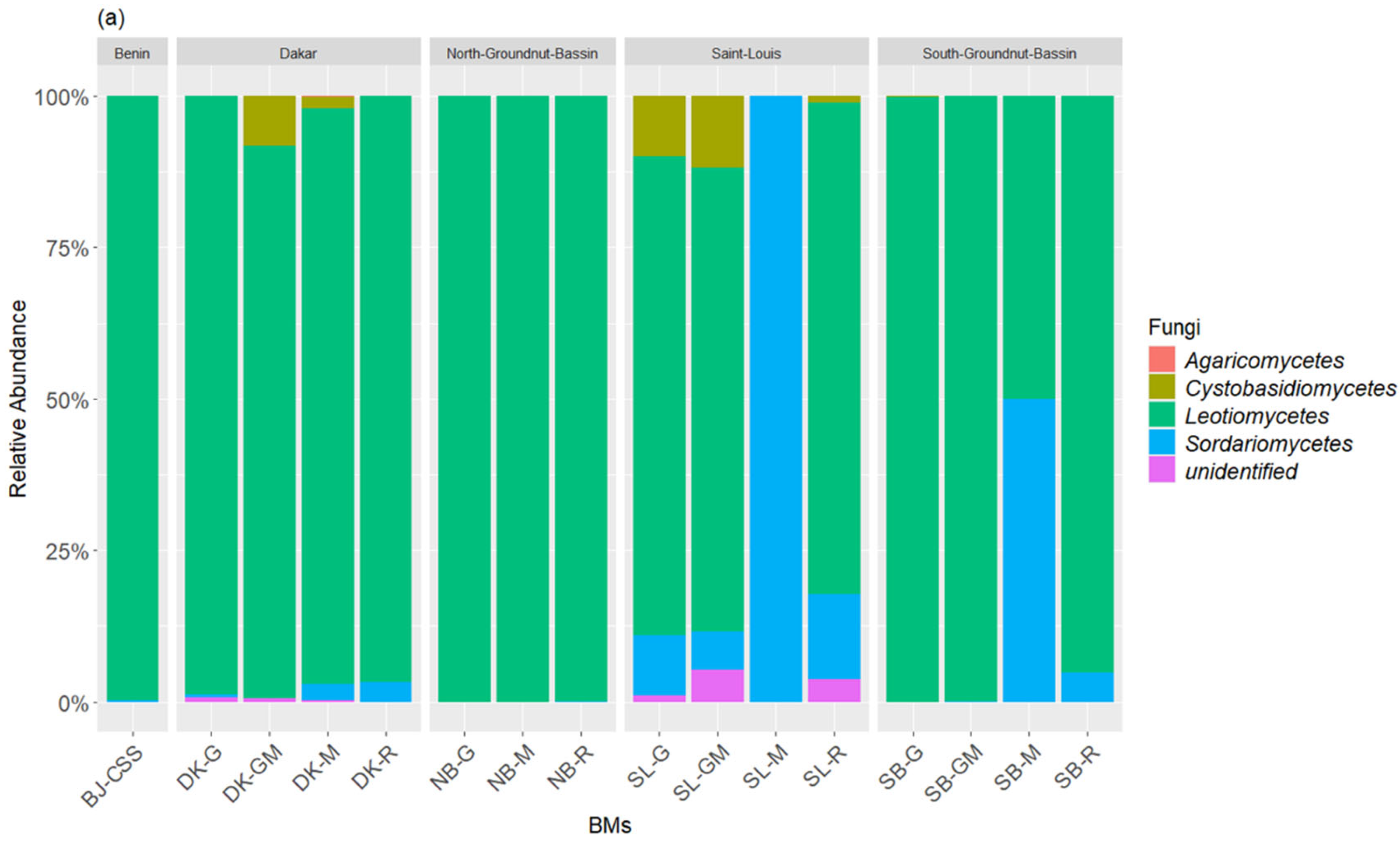
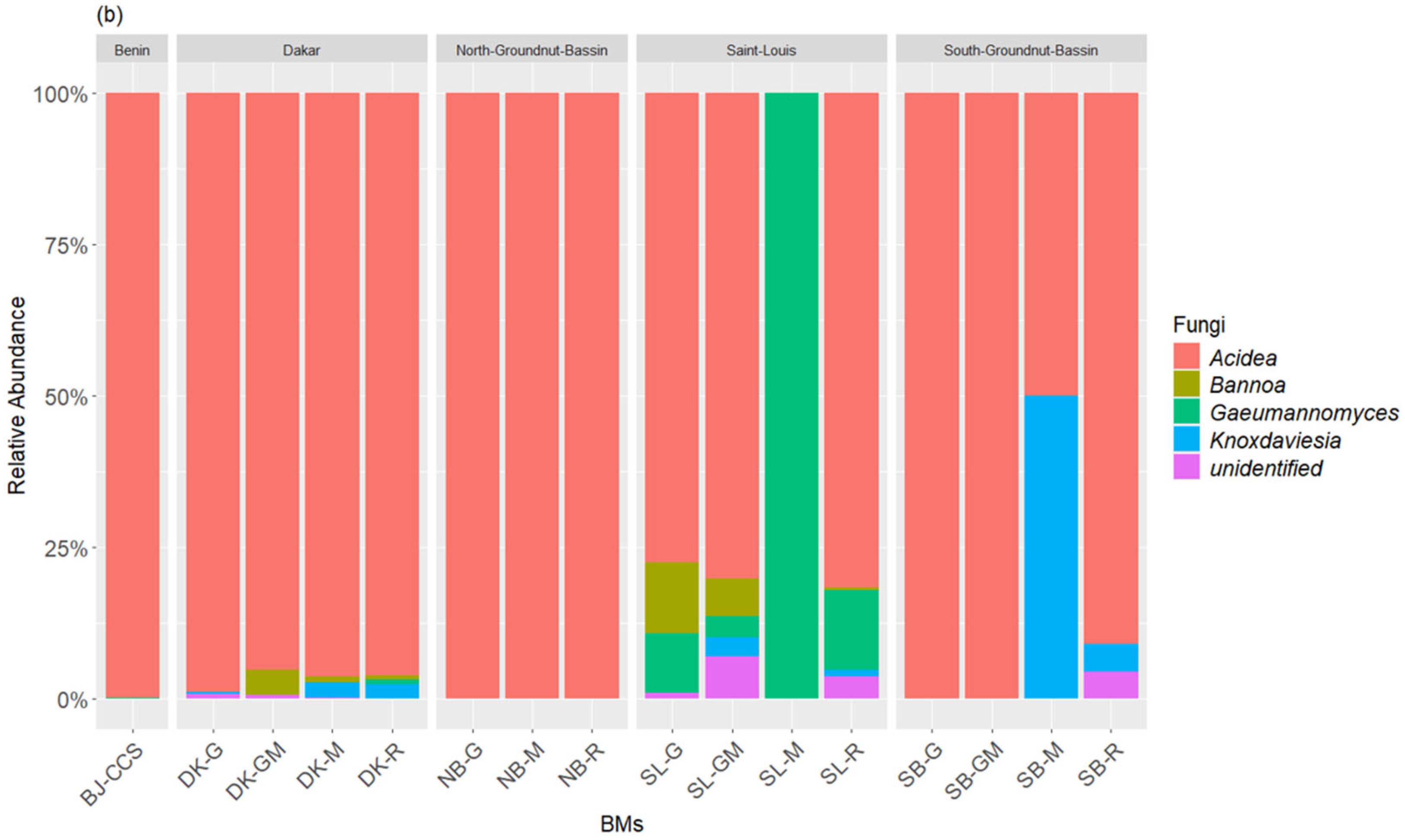
3.1. Composition of Bacterial and Fungal Communities of the Liquid BMs Samples
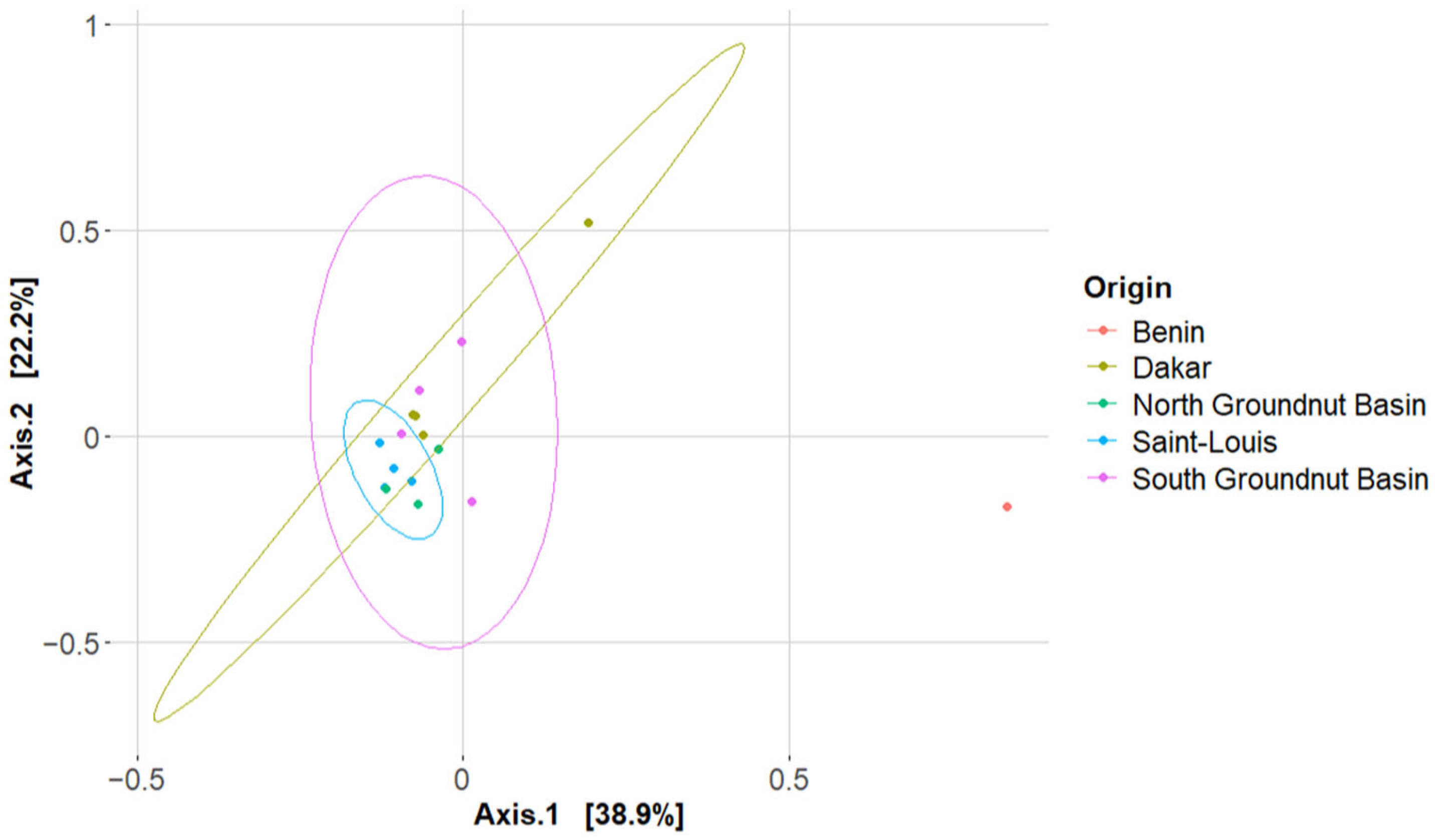
3.2. Bacterial α and β-Diversity
3.3. Effects of BMs on Aerial Growth of Tomato
3.4. Effect of BMs on Tomato Root Traits
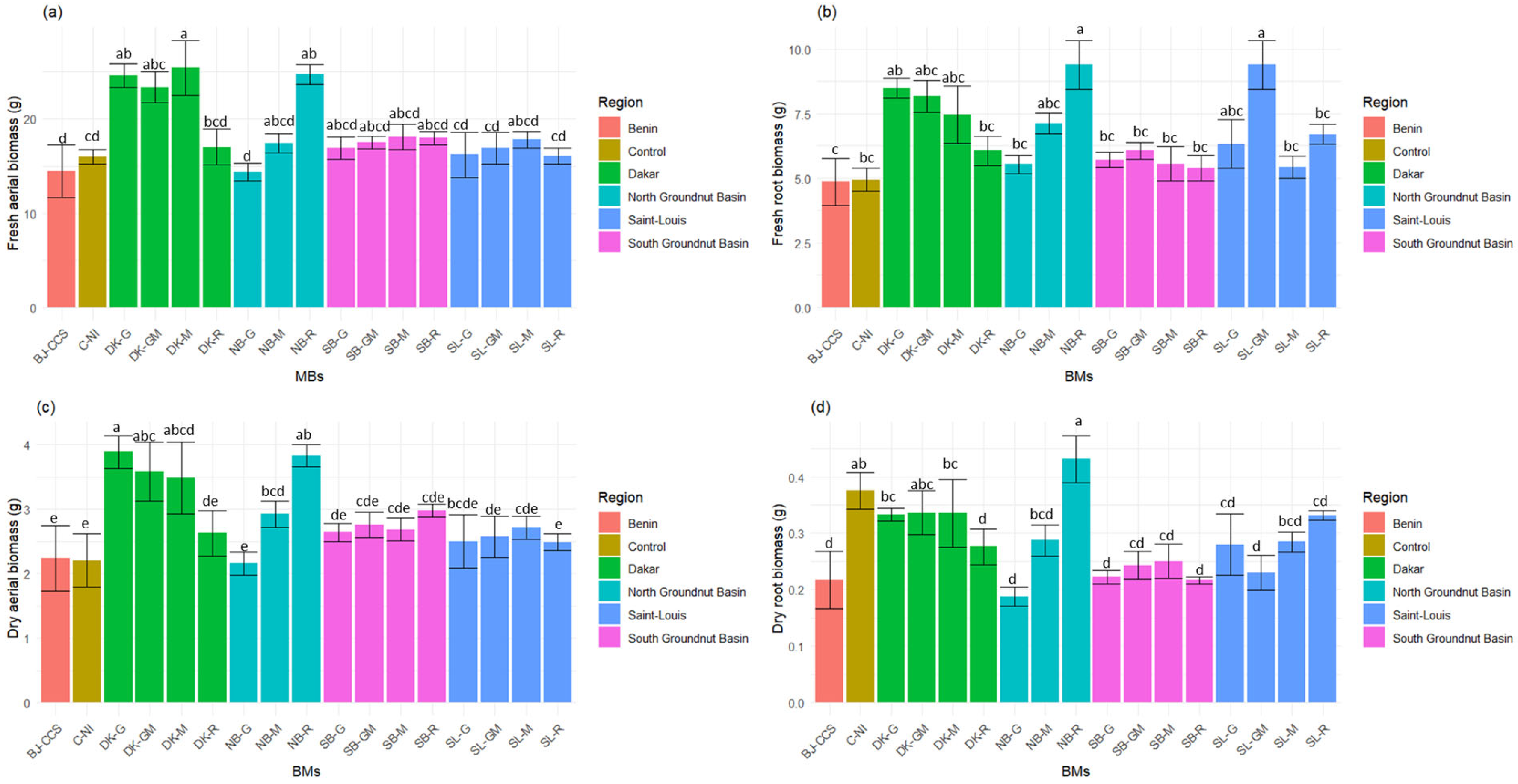
3.5. Effects of BMs on the Aerial and Root Biomass of Tomato Plants
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Diédhiou, S.O.; Ndiaye, T.M.N. Caractérisation des catégories d’espaces et contribution du maraîchage à la sécurité alimentaire dans la ville de Ziguinchor au Sénégal. Rev. Afr. D’environ. D’agric. 2021, 4, 10–22. [Google Scholar]
- Faye, C.; Sané, B.; Cissokho, D.; Diédhiou, S.O. Structure of peri-urban market gardening and degradation of resources (soils and water) in the zone of Boutoute in Ziguinchor (Senegal). J. D’econ. Manag. D’environ. Droit 2019, 2, 49–60. [Google Scholar]
- Higa, T.; Wididana, G.N. The Concept and Theories of Effective Microorganisms. In Proceedings of the First International Conference on Kyusei Nature Farming, Khon Kaen, Thailand, 17–21 October 1989; US Department of Agriculture: Washington, DC, USA, 1991; pp. 118–124. [Google Scholar]
- Higa, T. Effective Microorganisms: A New Dimension for Nature Farming. In Proceedings of the Second International Conference on Kyusei Nature Farming, Piracicaba, Brazil, 7–11 October 1991; US Department of Agriculture: Washington, DC, USA, 1994; pp. 20–22. [Google Scholar]
- Faltyn, U.; Miszkielo, T. The influence of effective microorganisms on germinability of dressed spring wheat seeds. Zesz. Nauk. Uniw. Przyr. We Wroclawiu Rolnictwo. Pol. 2008, 92, No. 568. 31–35 ref. 9. [Google Scholar]
- Ertekin, M. Effects of Microorganisms, Hormone Treatment and Stratification on Seed Germination of Goldenrain Tree (Koelreuteria paniculata). Int. J. Agric. Biol. 2011, 13, 38–42. [Google Scholar]
- Van Tonder, N.C.P.; Van der Westhuizen, C.; Van der Westhuizen, R.J. Interaction Effects of Effective Microorganisms and Prolonged Storage on Germination and Seedling Vigour of Maize, Sorghum and Sunflower. J. New Gener. Sci. 2014, 12, 147–161. Available online: https://journals.co.za/doi/abs/10.10520/EJC166483 (accessed on 5 May 2022).
- Hu, C.; Qi, Y. Long-Term Effective Microorganisms Application Promote Growth and Increase Yields and Nutrition of Wheat in China. Eur. J. Agron. 2013, 46, 63–67. [Google Scholar] [CrossRef]
- Abd El-Mageed, T.A.; Gyushi, M.A.H.; Hemida, K.A.; El-Saadony, M.T.; Abd El-Mageed, S.A.; Abdalla, H.; AbuQamar, S.F.; El-Tarabily, K.A.; Abdelkhalik, A. Coapplication of Effective Microorganisms and Nanomagnesium Boosts the Agronomic, Physio-Biochemical, Osmolytes, and Antioxidants Defenses Against Salt Stress in Ipomoea Batatas. Front. Plant Sci. 2022, 13, 883274. [Google Scholar] [CrossRef]
- Mbouobda, H.; Fotso; Astride Carole, D.; Kilovis, F.; Ndoumou, D. Impact of Effective and Indigenous Microorganisms Manures on Colocassia Esculenta and Enzymes Activities. Afr. J. Agric. Res. 2013, 8, 1086–1092. [Google Scholar] [CrossRef]
- Desfontaines, L.; Rotin, P.; Ozier-Lafontaine, H. Les Biostimulants: Qu’en savons-nous? Quelles alternatives pour l’agriculture Guyanaise? Innov. Agron. 2018, 64, 31–46. [Google Scholar] [CrossRef]
- Joshi, H.; Somduttand, C.P.; Mundra, S.L. Role of Effective Microorganisms (EM) in Sustainable Agriculture. Int. J. Curr. Microbiol. Appl. Sci. 2019, 8, 172–181. [Google Scholar] [CrossRef]
- Ahirwar, N.K.; Singh, R.; Chaurasia, S.; Chandra, R.; Prajapati, S.; Ramana, S. Effective Role of Beneficial Microbes in Achieving the Sustainable Agriculture and Eco-Friendly Environment Development Goals: A Review. Front. Environ. Microbiol. 2020, 5, 111–123. [Google Scholar] [CrossRef]
- Ezeagu, G.; Ijah, U.; Abioye, O.; Dauda, B. Efficacy of Organic Fertilizers Produced Using Locally Formulated Effective Microorganisms on the Growth and Yield Responses of Maize. Asian J. Biotechnol. Bioresour. Technol. 2017, 2, 1–9. [Google Scholar] [CrossRef]
- Khan, B.M.; Hossain, M.K.; Mridha, M.A.U. Effect of Microbial Inoculants on Albizia Saman Germination and Seedling Growth. J. For. Res. 2006, 17, 99–102. [Google Scholar] [CrossRef]
- Marois, J.; Lerch, T.Z.; Dunant, U.; Silva, A.-M.F.D.; Christen, P. Chemical and Microbial Characterization of Fermented Forest Litters Used as Biofertilizers. Microorganisms 2023, 11, 306. [Google Scholar] [CrossRef] [PubMed]
- Ney, L.; Franklin, D.; Mahmud, K.; Cabrera, M.; Hancock, D.; Habteselassie, M.; Newcomer, Q.; Dahal, S. Impact of Inoculation with Local Effective Microorganisms on Soil Nitrogen Cycling and Legume Productivity Using Composted Broiler Litter. Appl. Soil Ecol. 2020, 154, 103567. [Google Scholar] [CrossRef]
- Yamada, K.; Xu, H.-L. Properties and Applications of an Organic Fertilizer Inoculated with Effective Microorganisms. J. Crop Prod. 2001, 3, 255–268. [Google Scholar] [CrossRef]
- FAOSTAT FAOSTAT. Available online: https://www.fao.org/faostat/fr/#data/QCL (accessed on 5 May 2022).
- FAOSTAT FAOSTAT. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 11 October 2024).
- Manuel de la Litière Forestière Fermentée Une Préparation Simple et Économique Pour Des Cultures Vigoureuses-Broché–Terre Et Humanisme, Marc Dufumier—Achat Livre|Fnac. Available online: https://www.fnac.com/a15514025/Terre-Et-Humanisme-Manuel-de-la-litiere-forestiere-fermentee (accessed on 27 October 2024).
- Christen, P.; Marois, J.; Ammar, I.; Abecassis, V.; Lerch, T.; Criquet, S.; Greff, S.; Davidson, S.; Gries, A.; Martinez, M.; et al. In Proceedings of the Taller Científico Internacional Biochar en la Agricultura Sostenible, Matanzas, Cuba, 14–16 March 2023.
- Tournier, E.; Amenc, L.; Pablo, A.L.; Legname, E.; Blanchart, E.; Plassard, C.; Robin, A.; Bernard, L. Modification of a Commercial DNA Extraction Kit for Safe and Rapid Recovery of DNA and RNA Simultaneously from Soil, without the Use of Harmful Solvents. MethodsX 2015, 2, 182–191. [Google Scholar] [CrossRef]
- Javaid, A.; Bajwa, R. Field Evaluation of Effective Microorganisms (EM) Application for Growth, Nodulation, and Nutrition of Mung Bean. Turk. J. Agric. For. 2011, 35, 443–452. [Google Scholar] [CrossRef]
- Olle, M.; Williams, I.H. Effective Microorganisms and Their Influence on Vegetable Production—A Review. J. Hortic. Sci. Biotechnol. 2013, 88, 380–386. [Google Scholar] [CrossRef]
- Maheswari, N.U.; Abirami, R. A Review on: Effective Microorganisms and It Applications. Asian J. Multidimens. Res. 2019, 8, 121. [Google Scholar] [CrossRef]
- Denis, C. Effets de la Litière Forestière Fermentée sur la Croissance Végétative, la Santé et le Rendement de Lactuca sativa (Laitue) et Solanum lycopersicum (Tomate) en Conditions Expérimentales; Terre & Humanisme: Lablachère, France, 2021; p. 58+annexes. [Google Scholar]
- Higa, T. What Is EM Technology. EM World J. 2000, 1, 1–6. [Google Scholar]
- Ezeagu, G.G.; Omotosho, A.O.; Suleiman, K.O. Effective Microorganisms: A Review of Their Products and Uses. Nile J. Eng. Appl. Sci. 2023, 1. [Google Scholar] [CrossRef]
- Miché, L.; Dries, A.; Ammar, I.B.; Davidson, S.; Cagnacci, L.; Combet-Blanc, Y.; Abecassis, V.; Penton Fernandez, G.; Christen, P. Changes in Chemical Properties and Microbial Communities’ Composition of a Forest Litter–Based Biofertilizer Produced through Aerated Solid-State Culture under Different Oxygen Conditions. Environ. Sci. Pollut. Res. 2024, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.; Hou, Q.; Kwok, L.; Yu, Z.; Zheng, Y.; Sun, Z.; Menghe, B.; Zhang, H. Bacterial Microbiota Compositions of Naturally Fermented Milk Are Shaped by Both Geographic Origin and Sample Type. J. Dairy Sci. 2016, 99, 7832–7841. [Google Scholar] [CrossRef] [PubMed]
- Raman, J.; Kim, J.-S.; Choi, K.R.; Eun, H.; Yang, D.; Ko, Y.-J.; Kim, S.-J. Application of Lactic Acid Bacteria (LAB) in Sustainable Agriculture: Advantages and Limitations. Int. J. Mol. Sci. 2022, 23, 7784. [Google Scholar] [CrossRef] [PubMed]
- Aguirre-Garcia, Y.L.; Nery-Flores, S.D.; Campos-Muzquiz, L.G.; Flores-Gallegos, A.C.; Palomo-Ligas, L.; Ascacio-Valdés, J.A.; Sepúlveda-Torres, L.; Rodríguez-Herrera, R. Lactic Acid Fermentation in the Food Industry and Bio-Preservation of Food. Fermentation 2024, 10, 168. [Google Scholar] [CrossRef]
- Gashash, E.A.; Osman, N.A.; Alsahli, A.A.; Hewait, H.M.; Ashmawi, A.E.; Alshallash, K.S.; El-Taher, A.M.; Azab, E.S.; Abd El-Raouf, H.S.; Ibrahim, M.F. Effects of Plant-Growth-Promoting Rhizobacteria (PGPR) and Cyanobacteria on Botanical Characteristics of Tomato (Solanum lycopersicon L.) Plants. Plants 2022, 11, 2732. [Google Scholar] [CrossRef]
- Mukhopadhyay, M.; Mukherjee, A.; Ganguli, S.; Chakraborti, A.; Roy, S.; Choudhury, S.S.; Subramaniyan, V.; Kumarasamy, V.; Sayed, A.A.; El-Demerdash, F.M. Marvels of Bacilli in Soil Amendment for Plant-Growth Promotion toward Sustainable Development Having Futuristic Socio-Economic Implications. Front. Microbiol. 2023, 14, 1293302. [Google Scholar] [CrossRef]
- Samain, E.; Duclercq, J.; Ait Barka, E.; Eickermann, M.; Ernenwein, C.; Mazoyon, C.; Sarazin, V.; Dubois, F.; Aussenac, T.; Selim, S. PGPR-Soil Microbial Communities’ Interactions and Their Influence on Wheat Growth Promotion and Resistance Induction against Mycosphaerella graminicola. Biology 2023, 12, 1416. [Google Scholar] [CrossRef]
- Wang, D.; Wang, C.; Chen, Y.; Xie, Z. Developing Plant-Growth-Promoting Rhizobacteria: A Crucial Approach for Achieving Sustainable Agriculture. Agronomy 2023, 13, 1835. [Google Scholar] [CrossRef]
- Huang, W.; Hu, H.; Hu, T.; Chen, H.; Wang, Q.; Chen, G.; Tu, L. Impact of Aqueous Extracts of Cinnamomum septentrionale Leaf Litter on the Growth and Photosynthetic Characteristics of Eucalyptus Grandis Seedlings. New For. 2015, 46, 561–576. [Google Scholar] [CrossRef]
- Huo, X.; Wang, D.; Bing, D.; Li, Y.; Kang, H.; Yang, H.; Wei, G.; Chao, Z. Appropriate Removal of Forest Litter Is Beneficial to Pinus tabuliformis Carr. Regeneration in a Pine and Oak Mixed Forest in the Qinling Mountains, China. Forests 2019, 10, 735. [Google Scholar] [CrossRef]
- Jarboui, R.; Dhouib, B.; Ammar, E. Effect of Food Waste Compost (FWC) and Its Non-Aerated Fermented Extract (NFCE) on Seeds Germination and Plant Growth. Open J. Soil Sci. 2021, 11, 122–138. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Cetner, M.D.; Samborska, I.A.; Lukasik, I.; Oukarroum, A.; Rusinowki, S.; Pietkiewicz, S.; Swiatek, M.; Dabrowski, P. Effective Microorganisms Impact on Photosynthetic Activity of Arabidopsis Plant Grown under Salinity Stress Conditions. Ann. Wars. Univ. Life Sci. SGGW. Land Reclam. 2016, 48, 153–163. [Google Scholar] [CrossRef]
- Domenico, P. Effective Microorganisms for Germination and Root Growth in Kalanchoe daigremontiana. World J. Adv. Res. Rev. 2019, 3, 47–53. [Google Scholar] [CrossRef]
- Cheng, H.; Yuan, M.; Tang, L.; Shen, Y.; Yu, Q.; Li, S. Integrated Microbiology and Metabolomics Analysis Reveal Responses of Soil Microorganisms and Metabolic Functions to Phosphorus Fertilizer on Semiarid Farm. Sci. Total Environ. 2022, 817, 152878. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Zhang, H.; Zhao, X.; Liu, P.; Wang, L.; Wang, W. A Functional Metagenomics Study of Soil Carbon and Nitrogen Degradation Networks and Limiting Factors on the Tibetan Plateau. Front. Microbiol. 2023, 14, 170806. [Google Scholar] [CrossRef]
- Morgan, J.A.W.; Bending, G.D.; White, P.J. Biological Costs and Benefits to Plant–Microbe Interactions in the Rhizosphere. J. Exp. Bot. 2005, 56, 1729–1739. [Google Scholar] [CrossRef]
- Ortíz-Castro, R.; Contreras-Cornejo, H.A.; Macías-Rodríguez, L.; López-Bucio, J. The Role of Microbial Signals in Plant Growth and Development. Plant Signal. Behav. 2009, 4, 701–712. [Google Scholar] [CrossRef]
- Lugtenberg, B.; Kamilova, F. Plant-Growth-Promoting Rhizobacteria. Annu. Rev. Microbiol. 2009, 63, 541–556. [Google Scholar] [CrossRef]
- Tian, H.; Lv, B.; Ding, T.; Bai, M.; Ding, Z. Auxin-BR Interaction Regulates Plant Growth and Development. Front. Plant Sci. 2018, 8, 2256. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Smith, S.M.; Li, J. Genetic Regulation of Shoot Architecture. Annu. Rev. Plant Biol. 2018, 69, 437–468. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Yu, J.; Sun, J.; Wang, C.; Chen, S.; Yang, C.; Yang, Y. Exogenous Inoculation of Microorganisms Effect on Root Exudates and Rhizosphere Microorganism of Tobaccos. Adv. Microbiol. 2021, 11, 510–528. [Google Scholar] [CrossRef]
- Youssef, M.A.; Yousef, A.F.; Ali, M.M.; Ahmed, A.I.; Lamlom, S.F.; Strobel, W.R.; Kalaji, H.M. Exogenously Applied Nitrogenous Fertilizers and Effective Microorganisms Improve Plant Growth of Stevia (Stevia rebaudiana Bertoni) and Soil Fertility. AMB Express 2021, 11, 133. [Google Scholar] [CrossRef]
- Pattnaik, S.; Mohapatra, B.; Kumar, U.; Pattnaik, M.; Samantaray, D. Microbe-Mediated Plant Growth Promotion: A Mechanistic Overview on Cultivable Plant Growth-Promoting Members. In Biofertilizers for Sustainable Agriculture and Environment; Giri, B., Prasad, R., Wu, Q.-S., Varma, A., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 435–463. ISBN 978-3-030-18933-4. [Google Scholar]
- Rêgo, M.C.F.; Ilkiu-Borges, F.; de Filippi, M.C.C.; Gonçalves, L.A.; Silva, G.B. da Morphoanatomical and Biochemical Changes in the Roots of Rice Plants Induced by Plant Growth-Promoting Microorganisms. J. Bot. 2014, 2014, 818797. [Google Scholar] [CrossRef]
- Niang, N.; Demaneche, S.; Ndoye, I.; Navarro, E. Genetic Diversity of Rhizobia and Plant Growth Promoting Rhizobacteria of Soil under the Influence of Piliostigma reticulatum (DC.) Hochst and Their Impact on Shrub Growth. Afr. J. Agric. Res. 2018, 13, 2668–2679. [Google Scholar] [CrossRef]
- Limanska, N.; Ivanytsia, T.; Basiul, O.; Krylova, K.; Biscola, V.; Chobert, J.-M.; Ivanytsia, V.; Haertlé, T. Effect of Lactobacillus plantarum on Germination and Growth of Tomato Seedlings. Acta Physiol. Plant. 2013, 35, 1587–1595. [Google Scholar] [CrossRef]
- Ngueuleu, D.A.; Muyang, R.F.; Tchiaze, I.A.V.; Wamba, F.O.S.; Dongho, D.F.F.; Tassong, D.S.; Tefouet, V.D.; Asseng, C.C.; Fotso, M.; Taffouo, V.D. Effects of Indigenous Microorganism Fertilizers (IMO), Effective Fertilizers (EM) and Mineral Fertilizers (NPK) on the Yield and Nutritional Value of Two Varieties of Arachis hypogaea Grown Locally in West Cameroon. Am. J. Plant Sci. 2025, 16, 216–231. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of BMs Products | Litter Collection Zones | Carbon Source Used | BMs Products Code * |
|---|---|---|---|
| 1 | Dakar | Groundnut shell | DK-G |
| 2 | Groundnut shell and Millet stover | DK-GM | |
| 3 | Millet stover | DK-M | |
| 4 | Rice bran | DK-R | |
| 5 | North Groundnut Basin | Groundnut shell | NB-G |
| 6 | Millet stover | NB-M | |
| 7 | Rice bran | NB-R | |
| 8 | South Groundnut Basin | Groundnut shell | SB-G |
| 9 | Groundnut shell and Millet stover | SB-GM | |
| 10 | Millet stover | SB-M | |
| 11 | Rice bran | SB-R | |
| 12 | Saint-Louis | Groundnut shell | SL-G |
| 13 | Groundnut shell and millet stover | SL-GM | |
| 14 | Millet stover | SL-M | |
| 15 | Rice bran | SL-R | |
| 16 | Commercial product Songhai Center Benin | Unknown | BJ-CCS |
| Bacterial α-Diversity | Fungal α-Diversity | |||||
|---|---|---|---|---|---|---|
| OTU Observed | Chao1 | Shannon Indices | OTU Observed | Chao1 | Shannon Indices | |
| BJ-CCS | 105 | 191.13 | 1.48 | 19 | 19.17 | 1.31 |
| SL-R | 69 | 104.10 | 1.92 | 14 | 15.00 | 1.53 |
| SL-M | 74 | 83.10 | 2.07 | 1 | 1.00 | 0.00 |
| SL-G | 87 | 108.08 | 2.17 | 8 | 8.25 | 0.86 |
| SL-GM | 100 | 117.10 | 1.86 | 13 | 16.00 | 1.54 |
| NB-R | 106 | 144.15 | 2.29 | 6 | 6.50 | 0.00 |
| NB-M | 87 | 94.77 | 2.03 | 2 | 2.00 | 0.01 |
| NB-G | 96 | 114.07 | 2.16 | 2 | 3.00 | 0.69 |
| SB-R | 88 | 121.46 | 2.02 | 4 | 7.00 | 0.55 |
| SB-M | 121 | 146.83 | 2.38 | 2 | 3.00 | 0.69 |
| SB-G | 80 | 101.08 | 2.11 | 3 | 3.00 | 0.00 |
| SL-GM | 96 | 153.00 | 2.30 | 3 | 4.00 | 0.01 |
| DK-R | 94 | 117.21 | 2.29 | 7 | 7.00 | 0.22 |
| DK-M | 126 | 138.50 | 2.12 | 17 | 17.00 | 0.69 |
| DK-G | 49 | 57.75 | 1.81 | 7 | 8.00 | 0.23 |
| DK-GM | 92 | 132.07 | 2.16 | 4 | 4.00 | 0.28 |
| Bacteria β-Diversity | Fungi β-Diversity | |||
|---|---|---|---|---|
| Structure | Dispersion | Structure | Dispersion | |
| R2 | F | R2 | F | |
| Origin | 0.59132 *** | 4.2700 | 0.33199 | 1.3518 |
| Carbon_Sources | 0.13171 | 1.2682 | 0.17683 | 0.9600 |
| BMs | Chlorophyll Content in Tomato Leaves | Plant Height (cm) |
|---|---|---|
| Untreated control | 38.182 cde | 44.318 def |
| BJ-CCS | 38.183 cde | 43.500 efg |
| DK-G | 45.033 a *** | 49.083 ab *** |
| DK-GM | 45.136 a *** | 49.182 ab *** |
| DK-M | 44.050 a *** | 50.125 a *** |
| DK-R | 37.600 de | 43.818 defg |
| NB-G | 39.864 bcd | 42.909 efg |
| NB-M | 41.318 b * | 42.909 efg |
| NB-R | 40.467 bc | 48.500 abc ** |
| SB-G | 36.492 e | 41.625 fg |
| SB-GM | 37.942 de | 44.333 def |
| SB-M | 36.658 e | 41.083 g |
| SUB-R | 39.845 bcd | 43.818 defg |
| SL-G | 38.000 de | 46.583 bcd |
| SL-GM | 36.045 e | 44.636 def |
| SL-M | 38.067 de | 45.667 cde |
| SL-R | 36.845 e | 44.955 de |
| BMs | Length (cm) | Surface (cm2) | Diameter (mm) | Volume (cm3) |
|---|---|---|---|---|
| C-NI | 11 921.47 | 99.05 | 0.57 | 2.68 |
| BJ-CCS | 12 743.54 | 98.28 | 0.54 | 2.54 |
| DK-G | 18 697.76 * | 151.71 ** | 0.67 | 4.94 *** |
| DK-GM | 19 208.93 | 150.17 * | 0.67 | 4.97 *** |
| DK-M | 16 479.96 | 133.92 | 0.64 | 4.267 * |
| DK-R | 17 740.28 | 122.10 | 0.54 | 3.21 |
| NB-G | 92 92.78 | 76.02 | 0.50 | 1.67 |
| NB-M | 20 959.97 ** | 141.52 | 0.58 | 3.971 * |
| NB-R | 19 336.82 * | 155.24 ** | 0.69 | 5.289 *** |
| SB-G | 15 106.81 | 108.89 | 0.54 | 2.73 |
| SB-GM | 18 328.67 | 118.15 | 0.52 | 2.87 |
| SB-M | 10 632.62 | 91.97 | 0.56 | 2.39 |
| SB-R | 14 974.29 | 97.89 | 0.48 | 2.15 |
| SL-G | 18 078.49 | 113.17 | 0.49 | 2.70 |
| SL-GM | 14 198.09 | 96.86 | 0.49 | 2.18 |
| SL-M | 14 782.35 | 114.65 | 0.62 | 3.60 |
| SL-R | 14 901.43 | 118.29 | 0.58 | 3.09 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zoumman, A.M.A.; Fernandes, P.; Gueye, M.; Chaintreuil, C.; Cournac, L.; Kane, A.; Assigbetse, K. Exploring Microbial Diversity in Forest Litter-Based Fermented Bioproducts and Their Effects on Tomato (Solanum lycopersicum L.) Growth in Senegal. Int. J. Plant Biol. 2025, 16, 55. https://doi.org/10.3390/ijpb16020055
Zoumman AMA, Fernandes P, Gueye M, Chaintreuil C, Cournac L, Kane A, Assigbetse K. Exploring Microbial Diversity in Forest Litter-Based Fermented Bioproducts and Their Effects on Tomato (Solanum lycopersicum L.) Growth in Senegal. International Journal of Plant Biology. 2025; 16(2):55. https://doi.org/10.3390/ijpb16020055
Chicago/Turabian StyleZoumman, Alexandre Mahougnon Aurel, Paula Fernandes, Mariama Gueye, Clémence Chaintreuil, Laurent Cournac, Aboubacry Kane, and Komi Assigbetse. 2025. "Exploring Microbial Diversity in Forest Litter-Based Fermented Bioproducts and Their Effects on Tomato (Solanum lycopersicum L.) Growth in Senegal" International Journal of Plant Biology 16, no. 2: 55. https://doi.org/10.3390/ijpb16020055
APA StyleZoumman, A. M. A., Fernandes, P., Gueye, M., Chaintreuil, C., Cournac, L., Kane, A., & Assigbetse, K. (2025). Exploring Microbial Diversity in Forest Litter-Based Fermented Bioproducts and Their Effects on Tomato (Solanum lycopersicum L.) Growth in Senegal. International Journal of Plant Biology, 16(2), 55. https://doi.org/10.3390/ijpb16020055