
Sensitization and Habituation of Hyper-Excitation to Constant Presentation of Pattern-Glare Stimuli
 , ,
, ,
Abstract
1. Introduction
1.1. Change Through Time
1.2. Stationary Period and DC-Shift
1.3. Hyper-Excitation
1.4. Hypotheses
- (1)
- Hyper-excitation will build up through the course of a period of constant stimulation.
- (2)
- Participants experiencing discomfort to the aggravating stimulus, the Medium, will also exhibit sensitisation, i.e., hyper-excitation will increase across repeating presentation, over a relatively fine temporal period, of a few seconds. This is consistent with findings discussed previously that migraine and epilepsy sufferers do not habituate to the repeated presentation of aggravating stimuli.
- (3)
- For those same participants, who experience discomfort to the Medium stimulus, we also believe that they will exhibit a change in hyper-excitation over the coarser time resolution of the entire experiment. However, the direction of this effect was less clear before collecting the data. We may have also seen sensitisation through the course of the entire experiment because we are continuing to drive the system with an aggravating stimulus. However, we may have observed habituation across this longer time period because even our participants who are sensitive to the Medium stimulus have not been clinically diagnosed with migraine, headache, or epilepsy. Accordingly, their brains may successfully habituate to the Medium stimulus with sufficient repetitions.
- (4)
- Finally, it is also possible that we will observe an interaction between the two different time granularities, which might, for example, show that sensitisation to repeated presentations of the Medium stimulus only obtains at the start of the experiment when those exhibiting discomfort are first subjected to the aggravating stimulus, with this effect waning through the course of the experiment, i.e., their brains habituating.
2. Methods
2.1. Data Collection
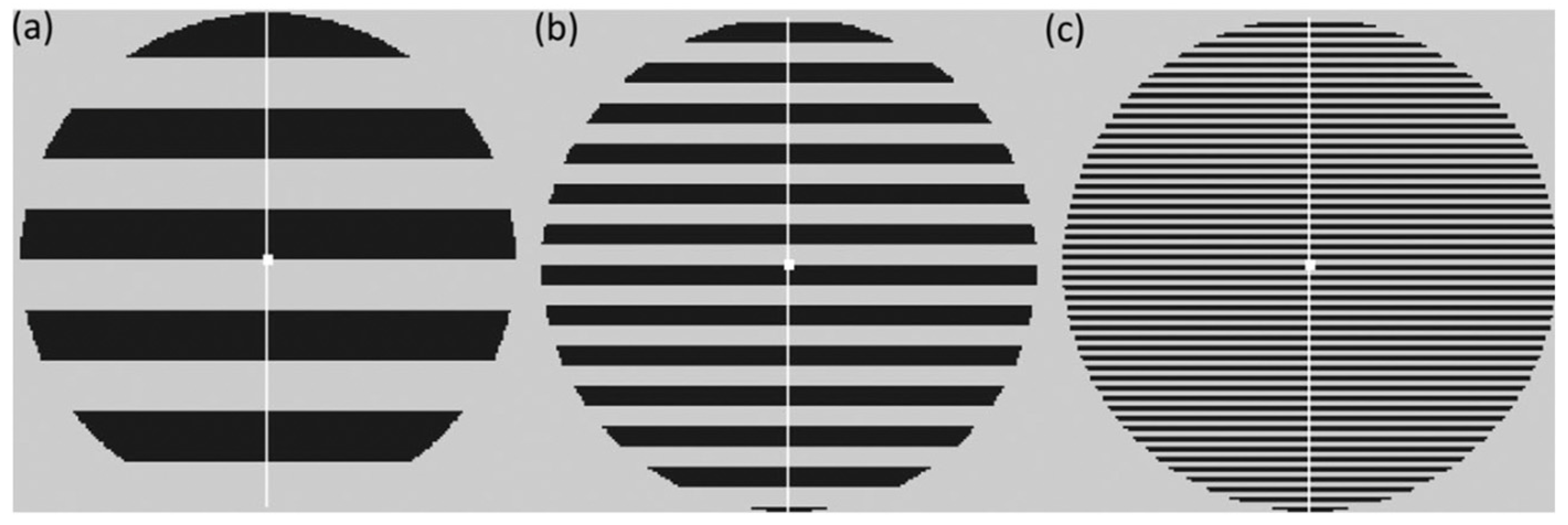
2.2. Questionnaires
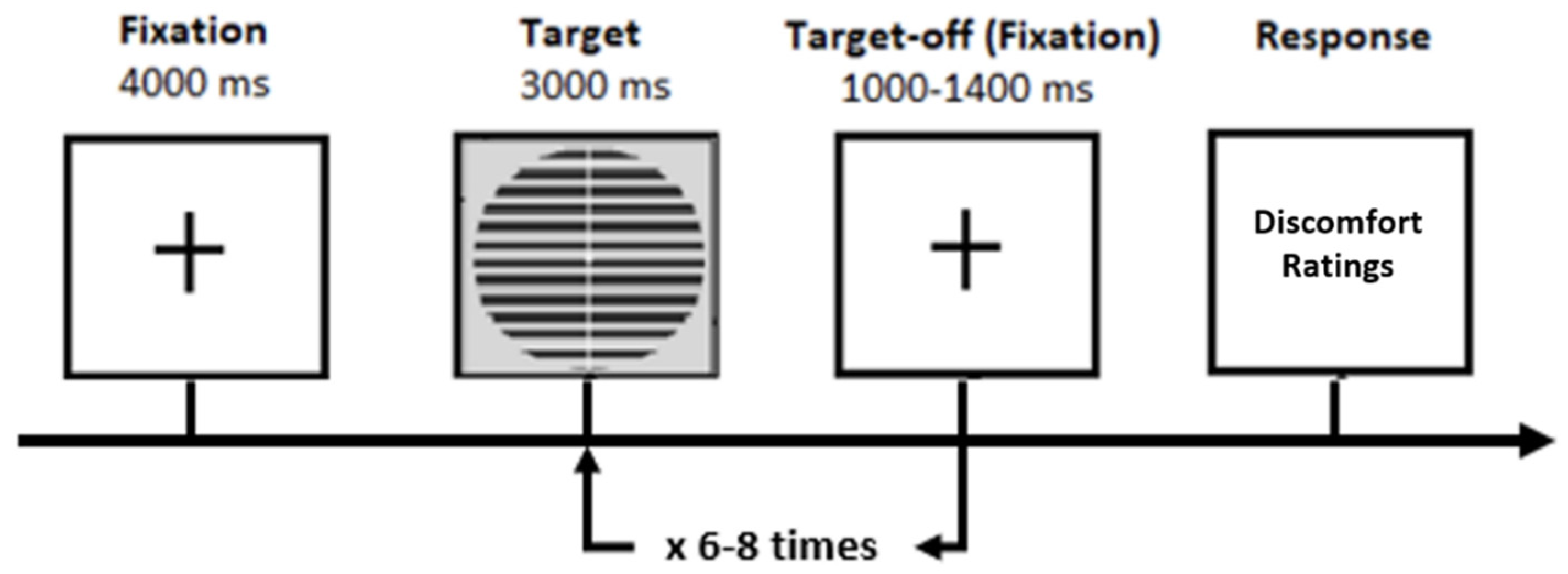
2.3. Procedure
2.4. Factor Analysis
2.5. Data Pre-Processing
2.6. Mass Univariate Analysis
2.6.1. Basic (Non-Temporal) Analyses
2.6.2. Change Through Time
2.6.3. Three-Way Interaction
2.7. Data Visualisation
3. Results
3.1. Average of Onsets 2–8
3.1.1. Mean/Intercept Effect
3.1.2. Factors
3.2. Partitions
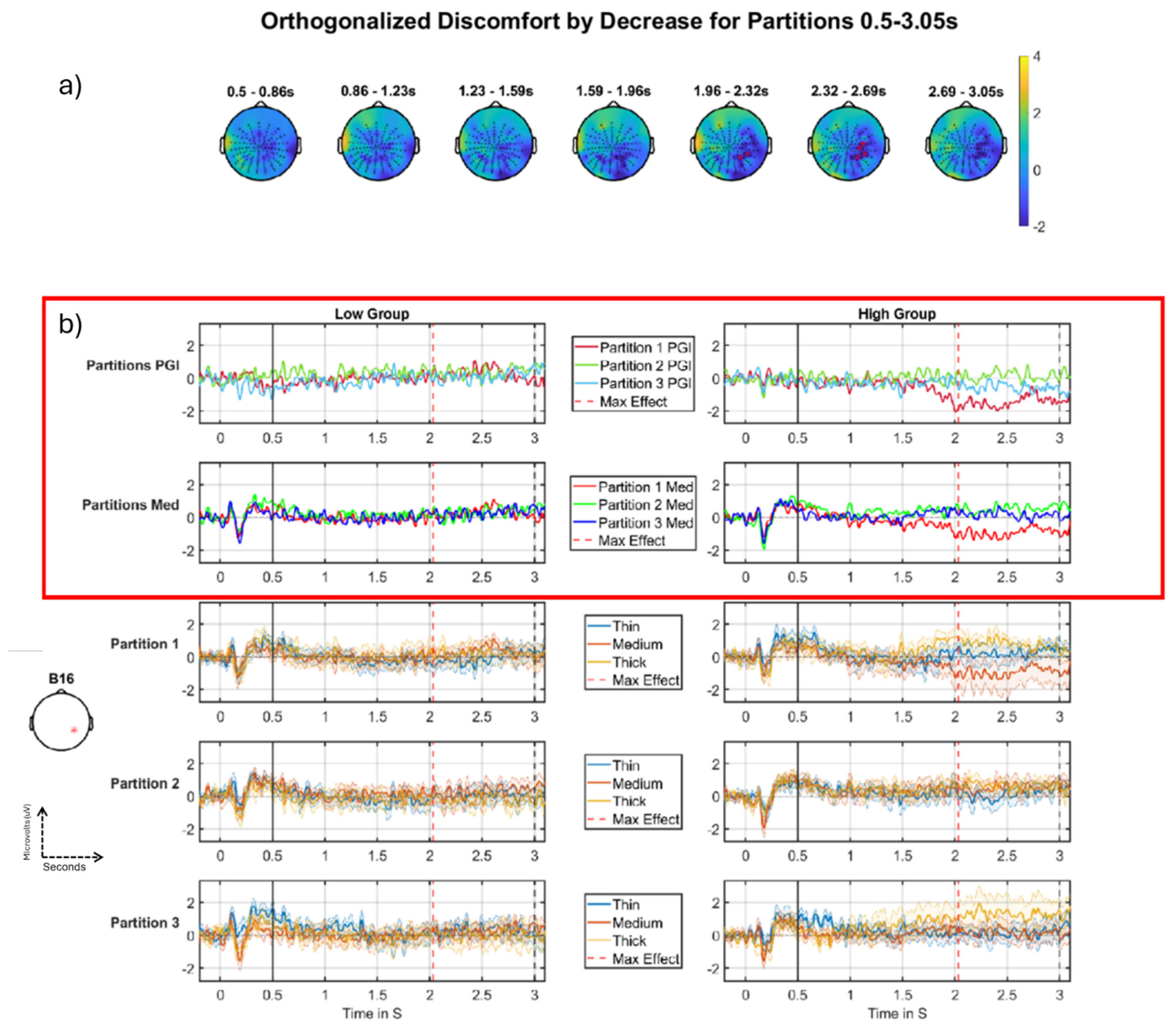
3.2.1. Discomfort-by-Decrease Across Partitions
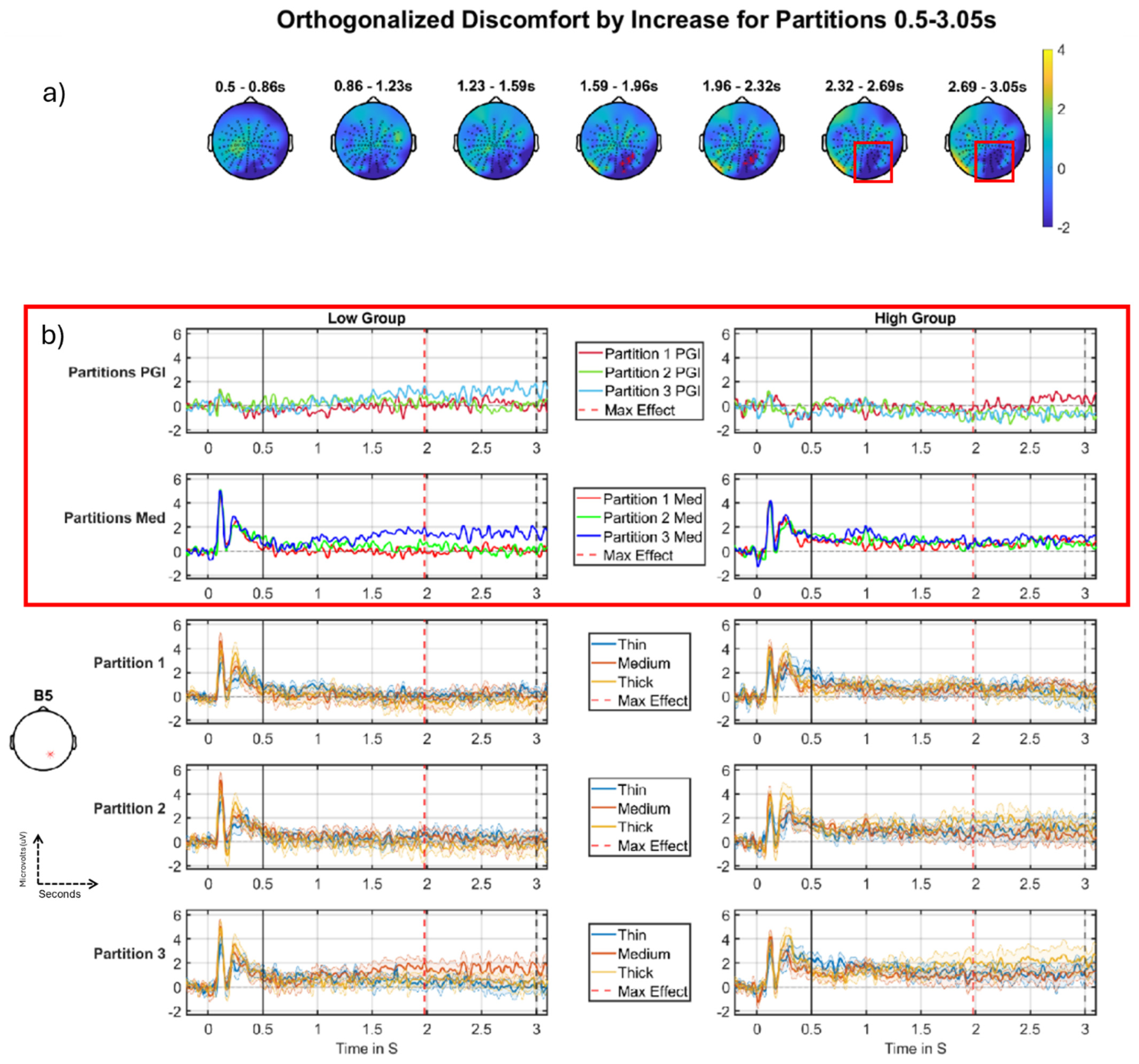
3.2.2. Discomfort-by-Increase Across Partitions
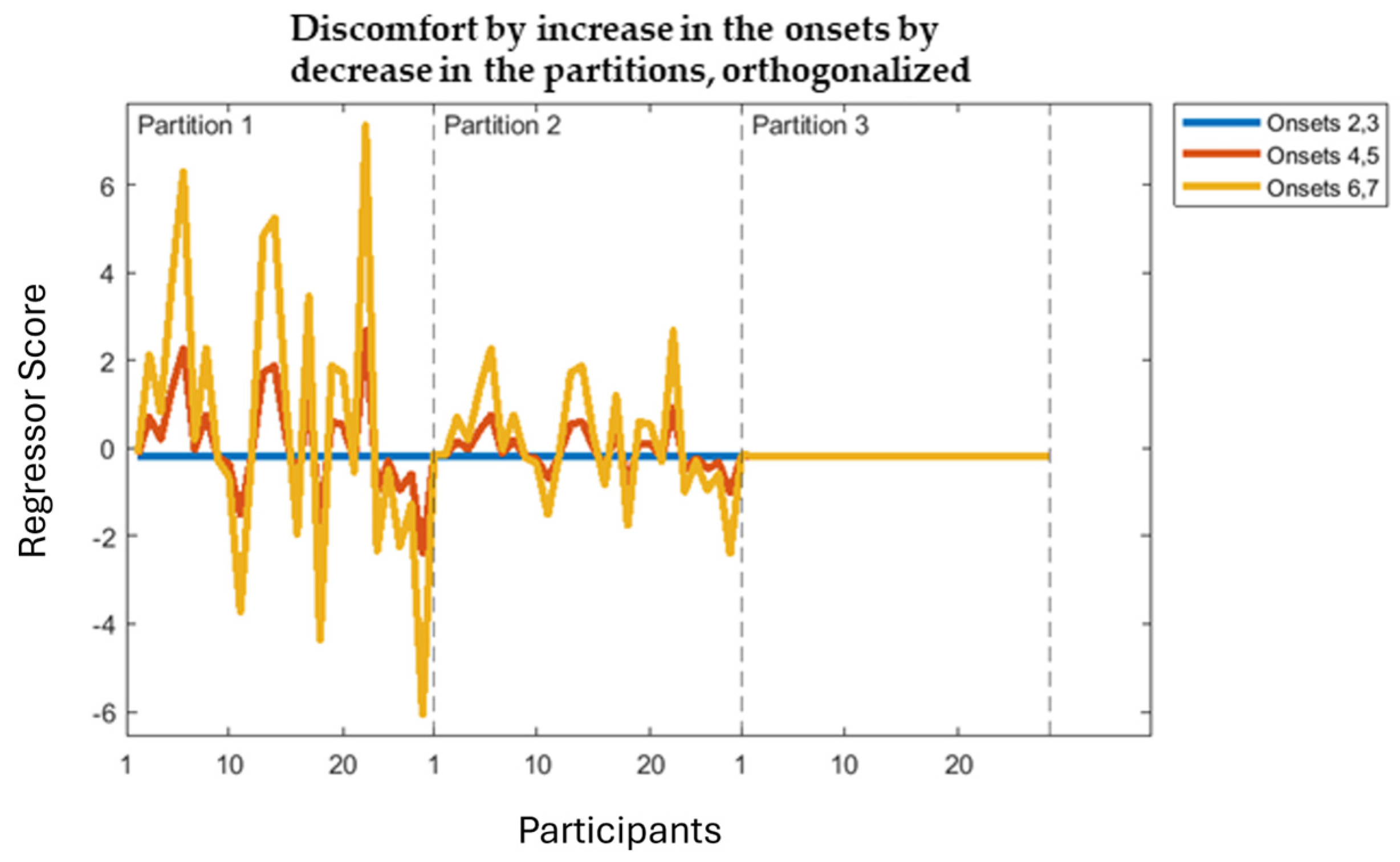
3.3. Onsets 2,3 vs. 4,5 vs. 6,7
Discomfort-by-Decrease
3.4. Three-Way Interaction
Discomfort-by-Increase Across the Partitions by Decrease Across the Onsets
4. Discussion
- Although we have made a number of comparisons, the number may not be unusual compared to higher-order ANOVAs that are routinely run. Additionally, the regressors we tested, e.g., the two-way interactions, are specifically formulated to test our prior temporal hypotheses, such as a factor interacting with habituation. Indeed, as indicated elsewhere, these two-way interactions are not the “standard” two-ways; they are tailored to our particular hypotheses. Having a strong hypothesis reduces vulnerability to type I errors.
- Also, we have been very conservative with regard to our analysis window. Specifically, we have undertaken a whole-volume analysis without systematically splitting the full analysis segment into sub windows, or indeed applying any sort of ROI. Additionally, that whole-volume is very large in comparison with what is typical in human-EEG cluster inference, viz. 2.5 s, when one would rarely see an analysis segment beyond 1 s. Such a large volume reduces statistical power with cluster inference; thus, our ability to find significant effects indicates the robustness of our findings.
- Additionally, some of our p-values are very small, e.g., the mean intercept p-value is 0.0002, and discomfort-by-decrease for onsets (a sensitisation effect) has a p-value of 0.0004. One would have had to perform an enormous number of comparisons before p-values of this size would become likely false positives. Although, of course, other of our effects are much weaker and should be viewed with more caution.
4.1. Mean/Intercept Effects
4.2. Effects on Discomfort Factor
4.2.1. Discomfort Habituating Through Partitions
4.2.2. Sensitisation Through the Onsets for the Discomfort Factor
4.2.3. Three-Way Interaction
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Evans, B.J.W.; Stevenson, S.J. The Pattern Glare Test: A review and determination of normative values. Ophthalmic Physiol. Opt. 2008, 28, 295–309. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, D.; Wilkins, A.J. Uncomfortable Images in Art and Nature. Perception 2008, 37, 1098–1113. [Google Scholar] [CrossRef] [PubMed]
- Monger, L.J.; Wilkins, A.J.; Allen, P.M. Pattern glare: The effects of contrast and color. Front. Psychol. 2015, 6, 1651. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, A.J. Visual Stress; Oxford University Press: Oxford, UK, 1995. [Google Scholar]
- Harle, D.E.; Evans, B.J.W. The optometric correlates of migraine. Ophthalmic Physiol. Opt. 2004, 24, 369–383. [Google Scholar] [CrossRef]
- Conlon, E.; Lovegrove, W.; Hine, T.; Chekaluk, E.; Piatek, K.; Hayes-Williams, K. The effects of visual discomfort and pattern structure on visual search. Perception 1998, 27, 21–33. [Google Scholar] [CrossRef]
- Turville, A.E. Refraction and migraine. Optom. Vis. Sci. 1935, 12, 336–361. [Google Scholar] [CrossRef]
- Wilkins, A.J.; Evans, B.J.W. Pattern Glare Test Instructions; IOO Sales Ltd.: London, UK, 2001. [Google Scholar]
- Aurora, S.; Wilkinson, F. The Brain is Hyperexcitable in Migraine. Cephalalgia 2007, 27, 1442–1453. [Google Scholar] [CrossRef]
- Adjamian, P.; Holliday, I.E.; Barnes, G.R.; Hillebrand, A.; Hadjipapas, A.; Singh, K.D. Induced visual illusions and gamma oscillations in human primary visual cortex. Eur. J. Neurosci. 2004, 20, 587–592. [Google Scholar] [CrossRef]
- Fong, C.Y.; Law, W.H.C.; Braithwaite, J.J.; Mazaheri, A. Differences in early and late pattern-onset visual-evoked potentials between self- reported migraineurs and controls. NeuroImage Clin. 2020, 25, 102122. [Google Scholar] [CrossRef]
- Harle, D.E.; Shepherd, A.J.; Evans, B.J.W. Visual Stimuli Are Common Triggers of Migraine and Are Associated With Pattern Glare. Headache J. Head Face Pain 2006, 46, 1431–1440. [Google Scholar] [CrossRef]
- Tempesta, A.J.; Miller, C.E.; Litvak, V.; Bowman, H.; Schofield, A.J. The missing N1 or jittered P2: Electrophysiological correlates of pattern glare in the time and frequency domain. Eur. J. Neurosci. 2021, 54, 6168–6186. [Google Scholar] [CrossRef] [PubMed]
- Brazzo, D.; Di Lorenzo, G.; Bill, P.; Fasce, M.; Papalia, G.; Veggiotti, P.; Seri, S. Abnormal visual habituation in pediatric photosensitive epilepsy. Clin. Neurophysiol. 2011, 122, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Coppola, G.; Pierelli, F.; Schoenen, J. Habituation and migraine. Neurobiol. Learn. Mem. 2009, 92, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Luck, S.J. An Introduction to the Event-Related Potential Technique; A Bradford Book: Cambridge, MA, USA, 2014. [Google Scholar]
- Thompson, P.; Burr, D. Visual aftereffects. Curr. Biol. 2009, 19, R11–R14. [Google Scholar] [CrossRef] [PubMed]
- Brainard, D.H. The Psychophysics Toolbox. Spat. Vis. 1997, 10, 433–436. [Google Scholar] [CrossRef]
- Kleiner, M.; Brainard, D.; Pelli, D.; Ingling, A.; Murray, R.; Broussard, C. What’s new in psychtoolbox-3. Perception 2007, 36, 1–16. [Google Scholar]
- Pelli, D.G. The VideoToolbox software for visual psychophysics: Transforming numbers into movies. Spat. Vis. 1997, 10, 437–442. [Google Scholar] [CrossRef]
- Braithwaite, J.J.; Broglia, E.; Bagshaw, A.P.; Wilkins, A.J. Evidence for elevated cortical hyperexcitability and its association with out-of-body experiences in the non-clinical population: New findings from a pattern-glare task. Cortex 2013, 49, 793–805. [Google Scholar] [CrossRef]
- Wilkins, A. A physiological basis for visual discomfort: Application in lighting design. Light. Res. Technol. 2016, 48, 44–54. [Google Scholar] [CrossRef]
- Conlon, E.; Lovegrove, W.; Barker, S.; Chekaluk, E. Visual discomfort: The influence of spatial frequency. Perception 2001, 30, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Braithwaite, J.J.; Marchant, R.; Takahashi, C.; Dewe, H.; Watson, D.G. The Cortical Hyperexcitability Index (CHi): A new measure for quantifying correlates of visually driven cortical hyperexcitability. Cogn. Neuropsychiatry 2015, 20, 330–348. [Google Scholar] [CrossRef] [PubMed]
- Conlon, E.G.; Lovegrove, W.J.; Chekaluk, E.; Pattison, P.E. Measuring Visual Discomfort. Vis. Cogn. 1999, 6, 637–663. [Google Scholar] [CrossRef]
- Arnold, M. Headache Classification Committee of the International Headache Society (IHS) The International Classification of Headache Disorders, 3rd edition. Cephalalgia 2018, 38, 1–211. [Google Scholar] [CrossRef]
- Dogan, C.; Miller, C.E.; Jefferis, T.; Saranti, M.; Tempesta, A.; Scofield, A.; Palaniappan, R.; Bowman, H. Headache-specific Hyperexcitation Sensitises and Habituates on different Time Scales: An Event Related Potential study of Pattern-Glare. bioRxiv 2024. bioRxiv:2024.08.28.610154. [Google Scholar]
- Jefferis, T.; Dogan, C.; Miller, C.E.; Karathanou, M.; Tempesta, A.J.; Schofield, A.J.; Bowman, H. Beyond EEG Onset Transients: Sensitisation and Habituation of Hyper-excitation to Constant Presentation and Offset of Pattern-Glare Stimuli. bioRxiv 2024. bioRxiv:2024.09.13.612622. [Google Scholar]
- Arfken, G.B. Mathematical Methods For Physicists; Academic Press: Cambridge, MA, USA, 1985. [Google Scholar] [CrossRef]
- Delorme, A.; Makeig, S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef]
- Oostenveld, R.; Fries, P.; Maris, E.; Schoffelen, J.-M. FieldTrip: Open Source Software for Advanced Analysis of MEG, EEG, and Invasive Electrophysiological Data. Comput. Intell. Neurosci. 2010, 2011, e156869. [Google Scholar] [CrossRef]
- Chennu, S.; Noreika, V.; Gueorguiev, D.; Blenkmann, A.; Kochen, S.; Ibáñez, A.; Owen, A.M.; Bekinschtein, T.A. Expectation and Attention in Hierarchical Auditory Prediction. J. Neurosci. 2013, 33, 11194. [Google Scholar] [CrossRef]
- Obrig, H.; Israel, H.; Kohl-Bareis, M.; Uludag, K.; Wenzel, R.; Müller, B.; Arnold, G.; Villringer, A. Habituation of the Visually Evoked Potential and Its Vascular Response: Implications for Neurovascular Coupling in the Healthy Adult. NeuroImage 2002, 17, 1–18. [Google Scholar] [CrossRef]
- Wang, W.; Schoenen, J. Interictal potentiation of passive “oddball“ auditory event-related potentials in migraine. Cephalalgia 1998, 18, 261–265; discussion 241. [Google Scholar] [CrossRef]
- Judit, Á.; Sándor, P.; Schoenen, J. Habituation of Visual and Intensity Dependence of Auditory Evoked Cortical Potentials Tends to Normalize Just Before and During the Migraine Attack. Cephalalgia 2000, 20, 714–719. [Google Scholar] [CrossRef] [PubMed]
- Omland, P.M.; Nilsen, K.B.; Uglem, M.; Gravdahl, G.; Linde, M.; Hagen, K.; Sand, T. Visual Evoked Potentials in Interictal Migraine: No Confirmation of Abnormal Habituation. Headache J. Head Face Pain 2013, 53, 1071–1086. [Google Scholar] [CrossRef] [PubMed]
- Schoenen, J.; Wang, W.; Albert, A.; Delwaide, P. Potentiation instead of habituation characterizes visual evoked potentials in migraine patients between attacks. Eur. J. Neurol. 1995, 2, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Fischer, H.; Furmark, T.; Wik, G.; Fredrikson, M. Brain representation of habituation to repeated complex visual stimulation studied with, P.E.T. NeuroReport 2000, 11, 123–126. [Google Scholar] [CrossRef]
- Mathewson, K.E.; Lleras, A.; Beck, D.M.; Fabiani, M.; Ro, T.; Gratton, G. Pulsed out of awareness: EEG alpha oscillations represent a pulsed-inhibition of ongoing cortical processing. Front. Psychol. 2011, 2, 99. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Effect | Tail (+1) | Tail (−1) |
|---|---|---|
| Mean/ intercept 0.5–3.0 s | 1st cluster p-value: 0.0002 1st electrode: A25 1st peak time: 0.9414 1st r correlation effect size: 0.79 1st Cohens d effect size: 2.59 | 1st cluster p-value: 0.0318 1st electrode: C13 1st peak time: 2.6914 1st r correlation effect size: −0.65 1st Cohens d effect size: −1.69 |
| Effect | Tail (+1) | Tail (−1) |
|---|---|---|
| Discomfort by decrease 0.5–3.0 s | No significant Cluster | 1st cluster p-value: 0.0192 1st electrode: B16 1st peak time: 2.0332 1st r correlation effect size: −0.48 1st Cohens d effect size: −1.1 |
| Effect | Tail (+1) | Tail (−1) |
|---|---|---|
| Discomfort by increase 0.5–3.0 s | No significant Cluster | 1st cluster p-value: 0.0108 1st electrode: B5 1st peak time: 1.9766 1st r correlation effect size: −0.45 1st Cohens d effect size: −1.0 2nd cluster p-value: 0.0116 2nd electrode: A31 2nd peak time: 2.8809 2nd r correlation effect size: −0.40 2nd Cohens d effect size: −0.88 |
| Effect | Tail (+1) | Tail (−1) |
|---|---|---|
| Discomfort by decrease 0.5–3.0 s | No significant Cluster | 1st cluster p-value: 0.0004 1st electrode: B16 1st peak time: 2.8037 1st r correlation effect size: −0.53 1st Cohens d effect size: −1.24 |
| Effect | Tail (+1) | Tail (−1) |
|---|---|---|
| Discomfort by increase in partitions by decrease in onsets 0.5–3.0 s | No significant Cluster | 1st cluster p-value: 0.0686 1st electrode: B5 1st peak time: 1.7578 1st r correlation effect size: −0.28 1st Cohens d effect size: −0.58 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jefferis, T.; Dogan, C.; Miller, C.E.; Karathanou, M.; Tempesta, A.; Schofield, A.J.; Bowman, H. Sensitization and Habituation of Hyper-Excitation to Constant Presentation of Pattern-Glare Stimuli. Neurol. Int. 2024, 16, 1585-1610. https://doi.org/10.3390/neurolint16060116
Jefferis T, Dogan C, Miller CE, Karathanou M, Tempesta A, Schofield AJ, Bowman H. Sensitization and Habituation of Hyper-Excitation to Constant Presentation of Pattern-Glare Stimuli. Neurology International. 2024; 16(6):1585-1610. https://doi.org/10.3390/neurolint16060116
Chicago/Turabian StyleJefferis, Thomas, Cihan Dogan, Claire E. Miller, Maria Karathanou, Austyn Tempesta, Andrew J. Schofield, and Howard Bowman. 2024. "Sensitization and Habituation of Hyper-Excitation to Constant Presentation of Pattern-Glare Stimuli" Neurology International 16, no. 6: 1585-1610. https://doi.org/10.3390/neurolint16060116
APA StyleJefferis, T., Dogan, C., Miller, C. E., Karathanou, M., Tempesta, A., Schofield, A. J., & Bowman, H. (2024). Sensitization and Habituation of Hyper-Excitation to Constant Presentation of Pattern-Glare Stimuli. Neurology International, 16(6), 1585-1610. https://doi.org/10.3390/neurolint16060116