

Development of Inhalable Bacteriophage Liposomes Against Pseudomonas aeruginosa
 , ,
, , 
Abstract

1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Phage Propagation and Determination of Phage Titer Using Plaque Assay
2.3. Phage Encapsulation in Liposomes
2.4. Phage Encapsulation Efficiency
2.5. Particle Characterization of Phage Liposomes
2.6. Release Studies
2.7. Nebulization Studies
2.7.1. Phage Viability upon Nebulization
2.7.2. In Vitro Aerosolization Studies
2.8. In Vitro Phage Cellular Uptake Studies
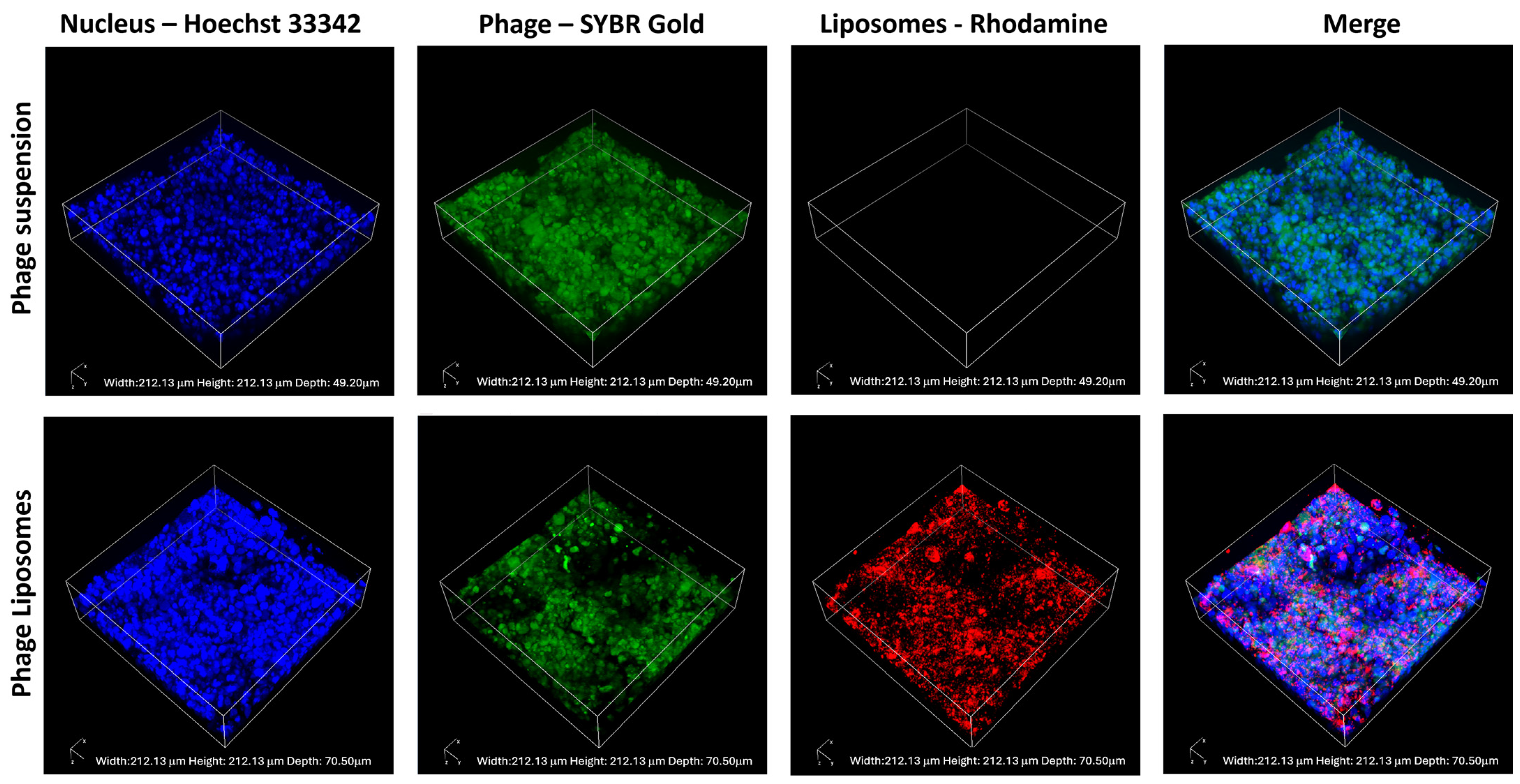
2.9. Phage Distribution Imaging Using Confocal Laser Scanning Microscopy
2.10. Statistical Analysis
3. Results
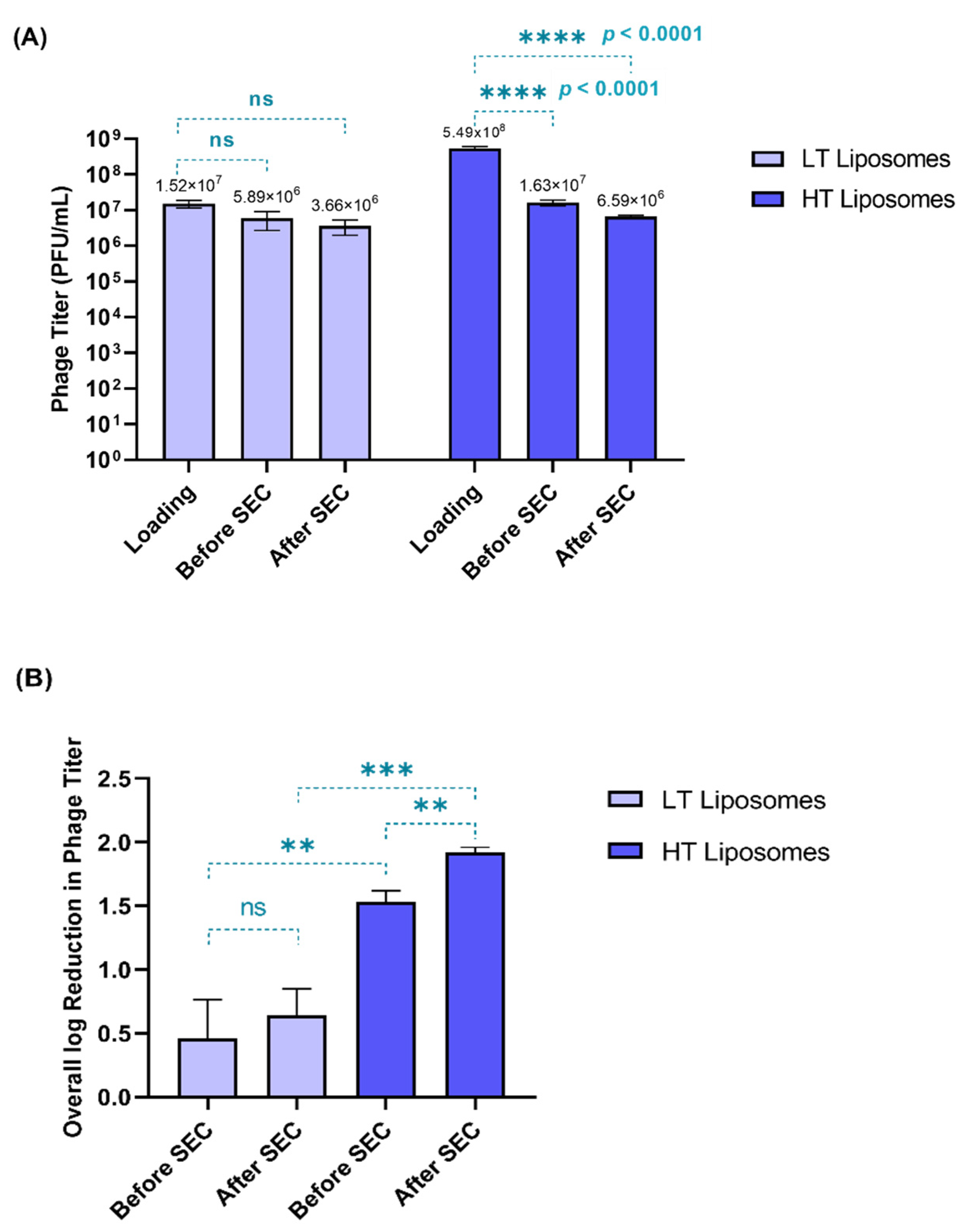
3.1. Development of Phage Liposomes—Effect of Process on Phage Viability
3.2. Particle Characterization and Phage Encapsulation
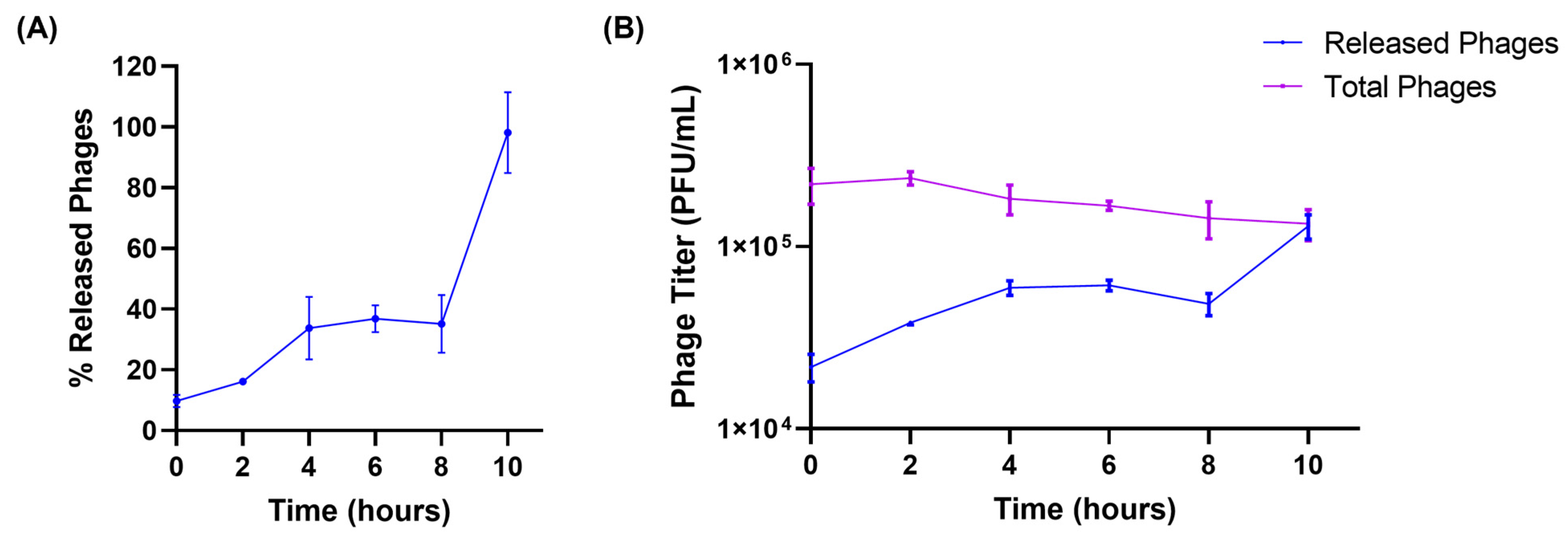
3.3. Release Studies
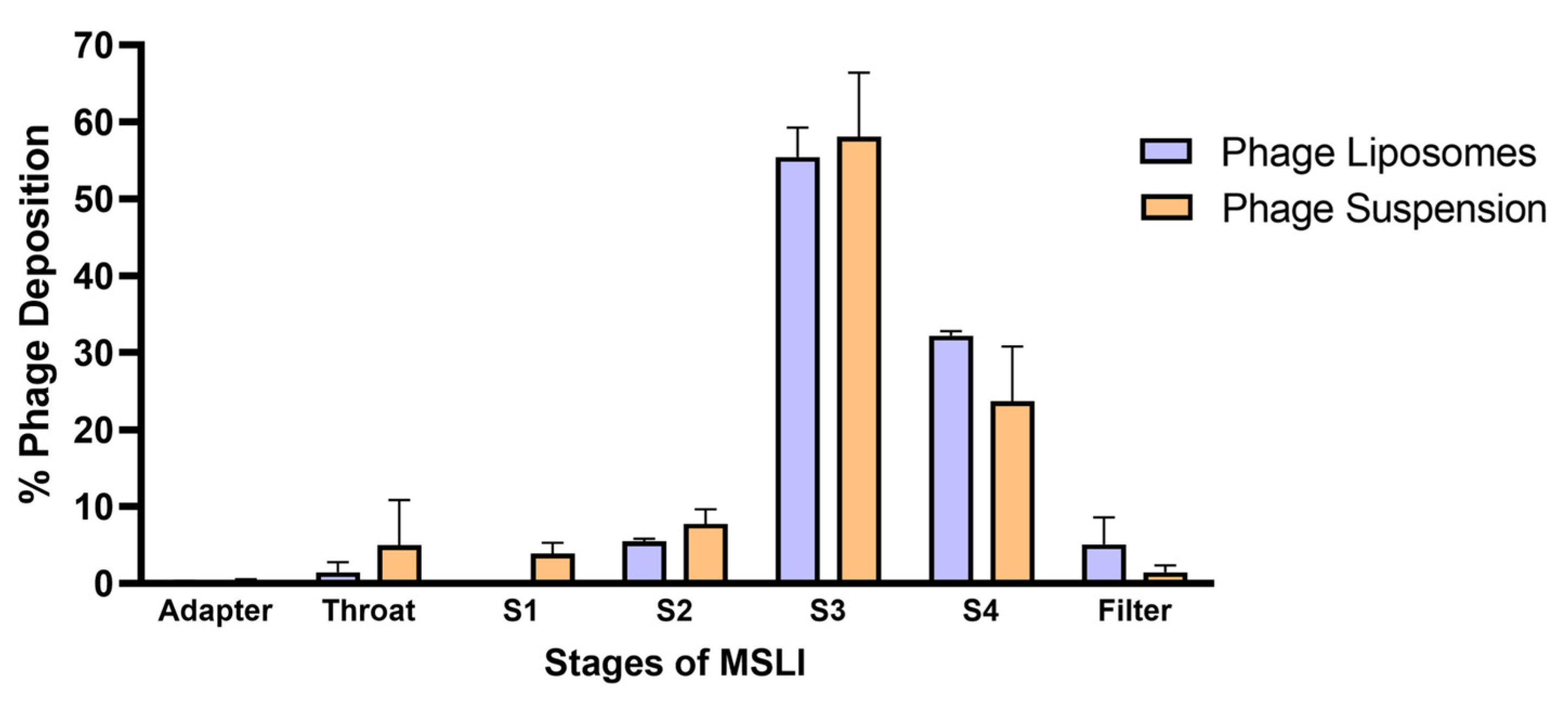
3.4. Nebulization Studies and In Vitro Lung Deposition
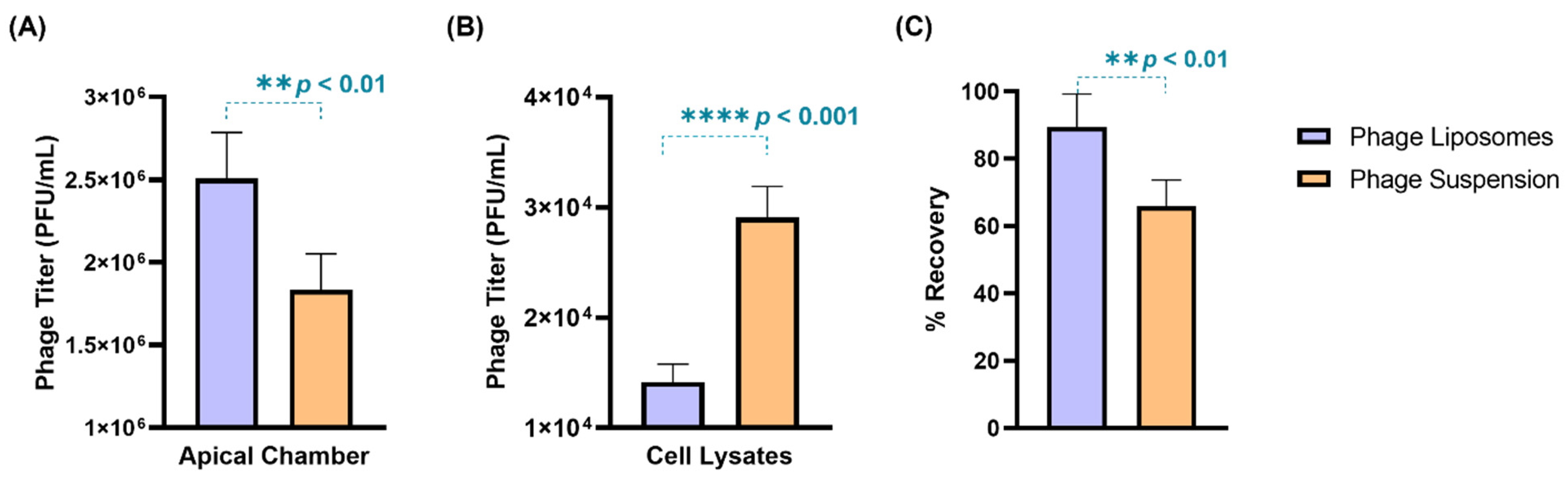
3.5. Phage Cellular Uptake—Quantitative Analysis
3.6. Phage Cellular Uptake—Confocal Laser Scanning Microscopy
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murray, C.J.L.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Aguilar, G.R.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global Burden of Bacterial Antimicrobial Resistance in 2019: A Systematic Analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations; Government of the United Kingdom: London, UK, 2016; Volume 2022, p. 10.
- Catalano, A.; Iacopetta, D.; Ceramella, J.; Pellegrino, M.; Giuzio, F.; Marra, M.; Rosano, C.; Saturnino, C.; Sinicropi, M.S.; Aquaro, S. Antibiotic-Resistant ESKAPE Pathogens and COVID-19: The Pandemic beyond the Pandemic. Viruses 2023, 15, 1843. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, A.; Khalid, N.A.; Zafar, S.; Majid, A.; Shahzadi, M.; Saleem, S.; Shah, A.A.; Badshah, M.; Khan, S. Phage Therapy as a Revolutionary Treatment for Multidrug-Resistant Pseudomonas aeruginosa Infections: A Narrative Review. Microbe 2024, 2, 100030. [Google Scholar] [CrossRef]
- Chauhan, G.; Shaik, A.A.; Sawant, S.S.; Diwan, R.; Mokashi, M.; Goyal, M.; Shukla, S.K.; Kunda, N.K.; Gupta, V. Continuously Producible Aztreonam-Loaded Inhalable Lipid Nanoparticles for Cystic Fibrosis-Associated Pseudomonas aeruginosa Infections—Development and in-Vitro Characterization. Biomater. Adv. 2025, 166, 214027. [Google Scholar] [CrossRef]
- Venturini, C.; Petrovic Fabijan, A.; Fajardo Lubian, A.; Barbirz, S.; Iredell, J. Biological Foundations of Successful Bacteriophage Therapy. EMBO Mol. Med. 2022, 14, e12435. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Le, S.; Zhu, T.; Wu, N. Regulations of Phage Therapy across the World. Front. Microbiol. 2023, 14, 1250848. [Google Scholar] [CrossRef]
- Yang, X.; Haque, A.; Matsuzaki, S.; Matsumoto, T.; Nakamura, S. The Efficacy of Phage Therapy in a Murine Model of Pseudomonas Aeruginosa Pneumonia and Sepsis. Front. Microbiol. 2021, 12, 682255. [Google Scholar] [CrossRef]
- Wienhold, S.-M.; Brack, M.C.; Nouailles, G.; Krishnamoorthy, G.; Korf, I.H.E.; Seitz, C.; Wienecke, S.; Dietert, K.; Gurtner, C.; Kershaw, O.; et al. Preclinical Assessment of Bacteriophage Therapy against Experimental Acinetobacter Baumannii Lung Infection. Viruses 2021, 14, 33. [Google Scholar] [CrossRef]
- Semler, D.D.; Goudie, A.D.; Finlay, W.H.; Dennis, J.J. Aerosol Phage Therapy Efficacy in Burkholderia Cepacia Complex Respiratory Infections. Antimicrob. Agents Chemother. 2014, 58, 4005–4013. [Google Scholar] [CrossRef]
- Guillon, A.; Pardessus, J.; L’Hostis, G.; Fevre, C.; Barc, C.; Dalloneau, E.; Jouan, Y.; Bodier-Montagutelli, E.; Perez, Y.; Thorey, C.; et al. Inhaled Bacteriophage Therapy in a Porcine Model of Pneumonia Caused by Pseudomonas Aeruginosa during Mechanical Ventilation. Br. J. Pharmacol. 2021, 178, 3829–3842. [Google Scholar] [CrossRef]
- Chow, M.Y.T.; Chang, R.Y.K.; Li, M.; Wang, Y.; Lin, Y.; Morales, S.; McLachlan, A.J.; Kutter, E.; Li, J.; Chan, H.-K. Pharmacokinetics and Time-Kill Study of Inhaled Antipseudomonal Bacteriophage Therapy in Mice. Antimicrob. Agents Chemother. 2020, 65, 10-1128. [Google Scholar] [CrossRef]
- Carmody, L.A.; Gill, J.J.; Summer, E.J.; Sajjan, U.S.; Gonzalez, C.F.; Young, R.F.; LiPuma, J.J. Efficacy of Bacteriophage Therapy in a Model of Burkholderia Cenocepacia Pulmonary Infection. J. Infect. Dis. 2010, 201, 264–271. [Google Scholar] [CrossRef]
- Flint, R.; Laucirica, D.R.; Chan, H.-K.; Chang, B.J.; Stick, S.M.; Kicic, A. Stability Considerations for Bacteriophages in Liquid Formulations Designed for Nebulization. Cells 2023, 12, 2057. [Google Scholar] [CrossRef]
- Abedon, S.T. Phage Therapy of Pulmonary Infections. Bacteriophage 2015, 5, e1020260. [Google Scholar] [CrossRef]
- Bichet, M.C.; Chin, W.H.; Richards, W.; Lin, Y.-W.; Avellaneda-Franco, L.; Hernandez, C.A.; Oddo, A.; Chernyavskiy, O.; Hilsenstein, V.; Neild, A.; et al. Bacteriophage Uptake by Mammalian Cell Layers Represents a Potential Sink That May Impact Phage Therapy. iScience 2021, 24, 102287. [Google Scholar] [CrossRef]
- Colom, J.; Cano-Sarabia, M.; Otero, J.; Cortés, P.; Maspoch, D.; Llagostera, M. Liposome-Encapsulated Bacteriophages for Enhanced Oral Phage Therapy against Salmonella spp. Appl. Environ. Microbiol. 2015, 81, 4841–4849. [Google Scholar] [CrossRef]
- Singla, S.; Harjai, K.; Katare, O.P.; Chhibber, S. Encapsulation of Bacteriophage in Liposome Accentuates Its Entry in to Macrophage and Shields It from Neutralizing Antibodies. PLoS ONE 2016, 11, e0153777. [Google Scholar] [CrossRef]
- Li, J.; Zheng, H.; Leung, S.S.Y. Investigating the Effectiveness of Liposome-Bacteriophage Nanocomplex in Killing Staphylococcus Aureus Using Epithelial Cell Coculture Models. Int. J. Pharm. 2024, 657, 124146. [Google Scholar] [CrossRef]
- Nieth, A.; Verseux, C.; Barnert, S.; Süss, R.; Römer, W. A First Step toward Liposome-Mediated Intracellular Bacteriophage Therapy. Expert. Opin. Drug Deliv. 2015, 12, 1411–1424. [Google Scholar] [CrossRef] [PubMed]
- Bonilla, N.; Rojas, M.I.; Netto Flores Cruz, G.; Hung, S.-H.; Rohwer, F.; Barr, J.J. Phage on Tap-a Quick and Efficient Protocol for the Preparation of Bacteriophage Laboratory Stocks. PeerJ 2016, 4, e2261. [Google Scholar] [CrossRef] [PubMed]
- Bichet, M.C.; Patwa, R.; Barr, J.J. Protocols for Studying Bacteriophage Interactions with in Vitro Epithelial Cell Layers. STAR Protoc. 2021, 2, 100697. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Yuan, H.; Chai, G.; Peng, K.; Zou, P.; Li, X.; Li, J.; Zhou, F.; Chan, H.-K.; Zhou, Q.T. Optimization of Inhalable Liposomal Powder Formulations and Evaluation of Their in Vitro Drug Delivery Behavior in Calu-3 Human Lung Epithelial Cells. Int. J. Pharm. 2020, 586, 119570. [Google Scholar] [CrossRef]
- Sawant, S.S.; Patil, S.M.; Shukla, S.K.; Kulkarni, N.S.; Gupta, V.; Kunda, N.K. Pulmonary Delivery of Osimertinib Liposomes for Non-Small Cell Lung Cancer Treatment: Formulation Development and in Vitro Evaluation. Drug Deliv. Transl. Res. 2022, 12, 2474–2487. [Google Scholar] [CrossRef]
- Daram, A.; Sawant, S.S.; Mehta, D.A.; Sanhueza, C.A.; Kunda, N.K. Inhalable Anti-EGFR Antibody-Conjugated Osimertinib Liposomes for Non-Small Cell Lung Cancer. Pharmaceutics 2024, 16, 1444. [Google Scholar] [CrossRef] [PubMed]
- Otero, J.; García-Rodríguez, A.; Cano-Sarabia, M.; Maspoch, D.; Marcos, R.; Cortés, P.; Llagostera, M. Biodistribution of Liposome-Encapsulated Bacteriophages and Their Transcytosis During Oral Phage Therapy. Front. Microbiol. 2019, 10, 689. [Google Scholar] [CrossRef]
- Porat, S.B.; Gelman, D.; Yerushalmy, O.; Alkalay-Oren, S.; Coppenhagen-Glazer, S.; Cohen-Cymberknoh, M.; Kerem, E.; Amirav, I.; Nir-Paz, R.; Hazan, R. Expanding Clinical Phage Microbiology: Simulating Phage Inhalation for Respiratory Tract Infections. ERJ Open Res. 2021, 7, 00367-2021. [Google Scholar] [CrossRef]
- Salomon, J.J.; Muchitsch, V.E.; Gausterer, J.C.; Schwagerus, E.; Huwer, H.; Daum, N.; Lehr, C.-M.; Ehrhardt, C. The Cell Line NCl-H441 Is a Useful in Vitro Model for Transport Studies of Human Distal Lung Epithelial Barrier. Mol. Pharm. 2014, 11, 995–1006. [Google Scholar] [CrossRef]
- Ren, H.; Birch, N.P.; Suresh, V. An Optimised Human Cell Culture Model for Alveolar Epithelial Transport. PLoS ONE 2016, 11, e0165225. [Google Scholar] [CrossRef]
- Ahmed, M.U.; Li, J.; Zhou, Q. Tobramycin Reduces Pulmonary Toxicity of Polymyxin B via Inhibiting the Megalin-Mediated Drug Uptake in the Human Lung Epithelial Cells. Pharmaceutics 2024, 16, 389. [Google Scholar] [CrossRef]
- Loh, B.; Gondil, V.S.; Manohar, P.; Khan, F.M.; Yang, H.; Leptihn, S. Encapsulation and Delivery of Therapeutic Phages. Appl. Environ. Microbiol. 2021, 87, e01979-20. [Google Scholar] [CrossRef]
- Cipolla, D.; Blanchard, J.; Gonda, I. Development of Liposomal Ciprofloxacin to Treat Lung Infections. Pharmaceutics 2016, 8, 6. [Google Scholar] [CrossRef] [PubMed]
- Mehta, P.P.; Ghoshal, D.; Pawar, A.P.; Kadam, S.S.; Dhapte-Pawar, V.S. Recent Advances in Inhalable Liposomes for Treatment of Pulmonary Diseases: Concept to Clinical Stance. J. Drug Deliv. Sci. Technol. 2020, 56, 101509. [Google Scholar] [CrossRef]
- Elhissi, A. Liposomes for Pulmonary Drug Delivery: The Role of Formulation and Inhalation Device Design. Curr. Pharm. Des. 2017, 23, 362–372. [Google Scholar] [CrossRef]
- Cinquerrui, S.; Mancuso, F.; Vladisavljević, G.T.; Bakker, S.E.; Malik, D.J. Nanoencapsulation of Bacteriophages in Liposomes Prepared Using Microfluidic Hydrodynamic Flow Focusing. Front. Microbiol. 2018, 9, 2172. [Google Scholar] [CrossRef] [PubMed]
- Kotta, S.; Aldawsari, H.M.; Badr-Eldin, S.M.; Binmahfouz, L.S.; Bakhaidar, R.B.; Sreeharsha, N.; Nair, A.B.; Ramnarayanan, C. Aerosol Delivery of Surfactant Liposomes for Management of Pulmonary Fibrosis: An Approach Supporting Pulmonary Mechanics. Pharmaceutics 2021, 13, 1851. [Google Scholar] [CrossRef]
- Fukunaga, Y.; Zandieh, M.; Liu, Y.; Liu, J. Salt-Induced Adsorption and Rupture of Liposomes on Microplastics. Langmuir 2023, 39, 16395–16403. [Google Scholar] [CrossRef]
- Cipolla, D.; Gonda, I.; Chan, H.-K. Liposomal Formulations for Inhalation. Ther. Deliv. 2013, 4, 1047–1072. [Google Scholar] [CrossRef]
- Wang, X.; Xie, Z.; Zhao, J.; Zhu, Z.; Yang, C.; Liu, Y. Prospects of Inhaled Phage Therapy for Combatting Pulmonary Infections. Front. Cell. Infect. Microbiol. 2021, 11, 758392. [Google Scholar] [CrossRef]
- Mirhadi, E.; Askarizadeh, A.; Farhoudi, L.; Mashreghi, M.; Behboodifar, S.; Alavizadeh, S.H.; Arabi, L.; Jaafari, M.R. The Impact of Phospholipids with High Transition Temperature to Enhance Redox-Sensitive Liposomal Doxorubicin Efficacy in Colon Carcinoma Model. Chem. Phys. Lipids 2024, 261, 105396. [Google Scholar] [CrossRef]
- Astudillo, A.; Leung, S.S.Y.; Kutter, E.; Morales, S.; Chan, H.-K. Nebulization Effects on Structural Stability of Bacteriophage PEV 44. Eur. J. Pharm. Biopharm. 2018, 125, 124–130. [Google Scholar] [CrossRef]
- Sahota, J.S.; Smith, C.M.; Radhakrishnan, P.; Winstanley, C.; Goderdzishvili, M.; Chanishvili, N.; Kadioglu, A.; O’Callaghan, C.; Clokie, M.R.J. Bacteriophage Delivery by Nebulization and Efficacy Against Phenotypically Diverse Pseudomonas Aeruginosa from Cystic Fibrosis Patients. J. Aerosol Med. Pulm. Drug Deliv. 2015, 28, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Huh, H.; Wong, S.; Jean, J.S.; Slavcev, R. Bacteriophage Interactions with Mammalian Tissue: Therapeutic Applications. Adv. Drug Deliv. Rev. 2019, 145, 4–17. [Google Scholar] [CrossRef]
- Williams, B.J.; Dehnbostel, J.; Blackwell, T.S. Pseudomonas Aeruginosa: Host Defence in Lung Diseases. Respirology 2010, 15, 1037–1056. [Google Scholar] [CrossRef] [PubMed]
- Resko, Z.J.; Suhi, R.F.; Thota, A.V.; Kroken, A.R. Evidence for Intracellular Pseudomonas Aeruginosa. J. Bacteriol. 2024, 206, e00109-24. [Google Scholar] [CrossRef]
- Malet, K.; Faure, E.; Adam, D.; Donner, J.; Liu, L.; Pilon, S.-J.; Fraser, R.; Jorth, P.; Newman, D.K.; Brochiero, E.; et al. Intracellular Pseudomonas Aeruginosa within the Airway Epithelium of Cystic Fibrosis Lung Tissues. Am. J. Respir. Crit. Care Med. 2024, 209, 1453–1462. [Google Scholar] [CrossRef] [PubMed]
- Nunes, S.S.; de Oliveira Silva, J.; Fernandes, R.S.; Miranda, S.E.M.; Leite, E.A.; de Farias, M.A.; Portugal, R.V.; Cassali, G.D.; Townsend, D.M.; Oliveira, M.C.; et al. PEGylated versus Non-PEGylated pH-Sensitive Liposomes: New Insights from a Comparative Antitumor Activity Study. Pharmaceutics 2022, 14, 272. [Google Scholar] [CrossRef]
- Miernikiewicz, P.; Dąbrowska, K. Endocytosis of Bacteriophages. Curr. Opin. Virol. 2022, 52, 229–235. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Formulation | Particle Size (nm) | Polydispersity Index (PDI) | Zeta Potential (mV) | Phage Encapsulation Efficiency (%) |
|---|---|---|---|---|
| HT Liposomes | 175.36 ± 3.87 | 0.036 ± 0.010 | −55.47 ± 2.07 | 25.15 ± 5.67 |
| LT Liposomes | 171.13 ± 2.04 | 0.030 ± 0.004 | −50.36 ± 6.07 | 58.33 ± 6.02 |
| Sample | Log Phage Titer (PFU/mL) | |||
|---|---|---|---|---|
| Loaded in Nebulizer | Phages Aerosolized | Phage Recovered | Loss on Nebulization | |
| Phage Suspension | 6.18 ± 0.07 | 6.04 ± 0.01 | 4.49 ± 0.04 | 1.55 ± 0.04 |
| Phage Liposomes | 5.92 ± 0.04 | 5.48 ± 0.05 | 4.40 ± 0.10 | 1.08 ± 0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sawant, S.S.; Ahmed, M.U.; Gantala, N.-G.; Chiu, C.; Qu, L.; Zhou, Q. Development of Inhalable Bacteriophage Liposomes Against Pseudomonas aeruginosa. Pharmaceutics 2025, 17, 405. https://doi.org/10.3390/pharmaceutics17040405
Sawant SS, Ahmed MU, Gantala N-G, Chiu C, Qu L, Zhou Q. Development of Inhalable Bacteriophage Liposomes Against Pseudomonas aeruginosa. Pharmaceutics. 2025; 17(4):405. https://doi.org/10.3390/pharmaceutics17040405
Chicago/Turabian StyleSawant, Shruti S., Maizbha Uddin Ahmed, Nathan-Gautham Gantala, Caitlin Chiu, Li Qu, and Qi Zhou. 2025. "Development of Inhalable Bacteriophage Liposomes Against Pseudomonas aeruginosa" Pharmaceutics 17, no. 4: 405. https://doi.org/10.3390/pharmaceutics17040405
APA StyleSawant, S. S., Ahmed, M. U., Gantala, N.-G., Chiu, C., Qu, L., & Zhou, Q. (2025). Development of Inhalable Bacteriophage Liposomes Against Pseudomonas aeruginosa. Pharmaceutics, 17(4), 405. https://doi.org/10.3390/pharmaceutics17040405