
Evaluation of Antimicrobial Properties and Potential Applications of Pseudomonas gessardii M15 Rhamnolipids towards Multiresistant Staphylococcus aureus

 ,
,  ,
,  , ,
, ,  ,
,  ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Rhamnolipids Mixtures
2.2. Liquid Chromatography-High Resolution Tandem Mass Spectrometry Analysis of M15RL and Commercial RLs Mixtures
2.3. Antibacterial Assay
2.4. MIC and MBC
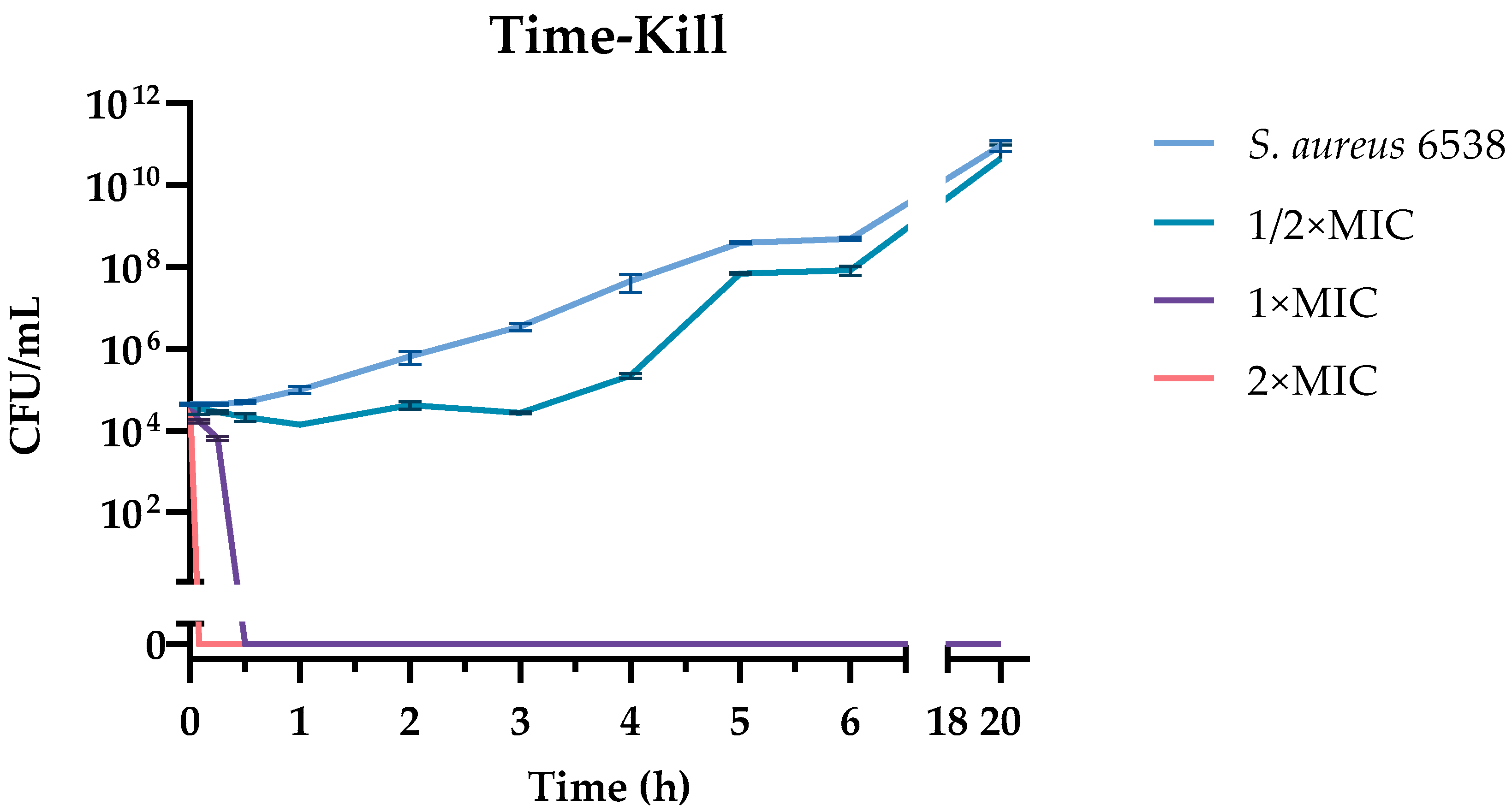
2.5. Time-Kill
2.6. Scanning Electron Microscopy
2.7. Antibiofilm Activity
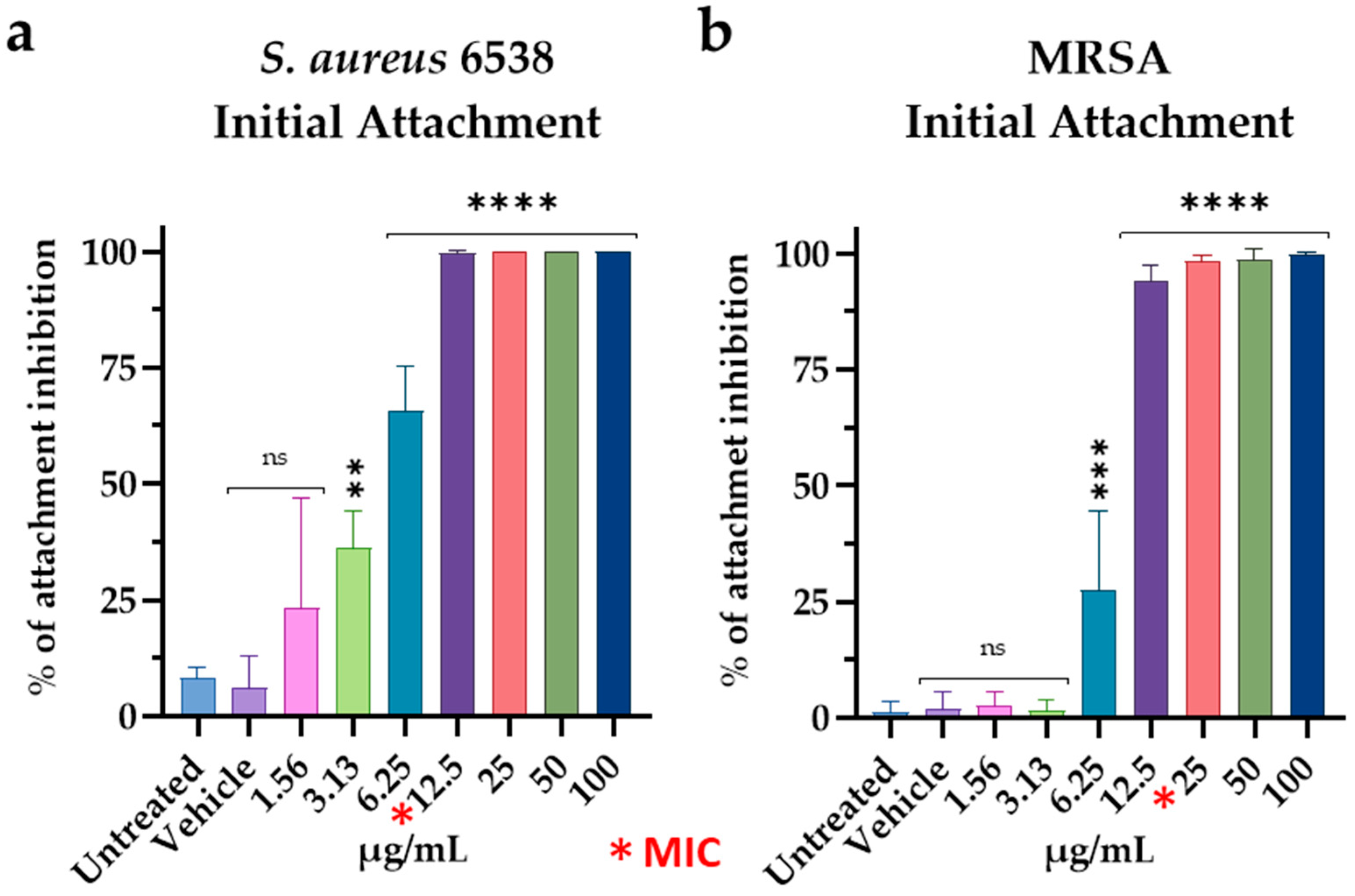
2.7.1. Initial Cell Attachment Assay
2.7.2. Biofilm Inhibition Assay
2.7.3. Biofilm Degradation
2.7.4. Biofilm Penetration
2.8. Cotton Swabs
2.9. Hemolytic Assay
2.10. Chemical and Physical Stress
2.11. Statistical Analysis
3. Results
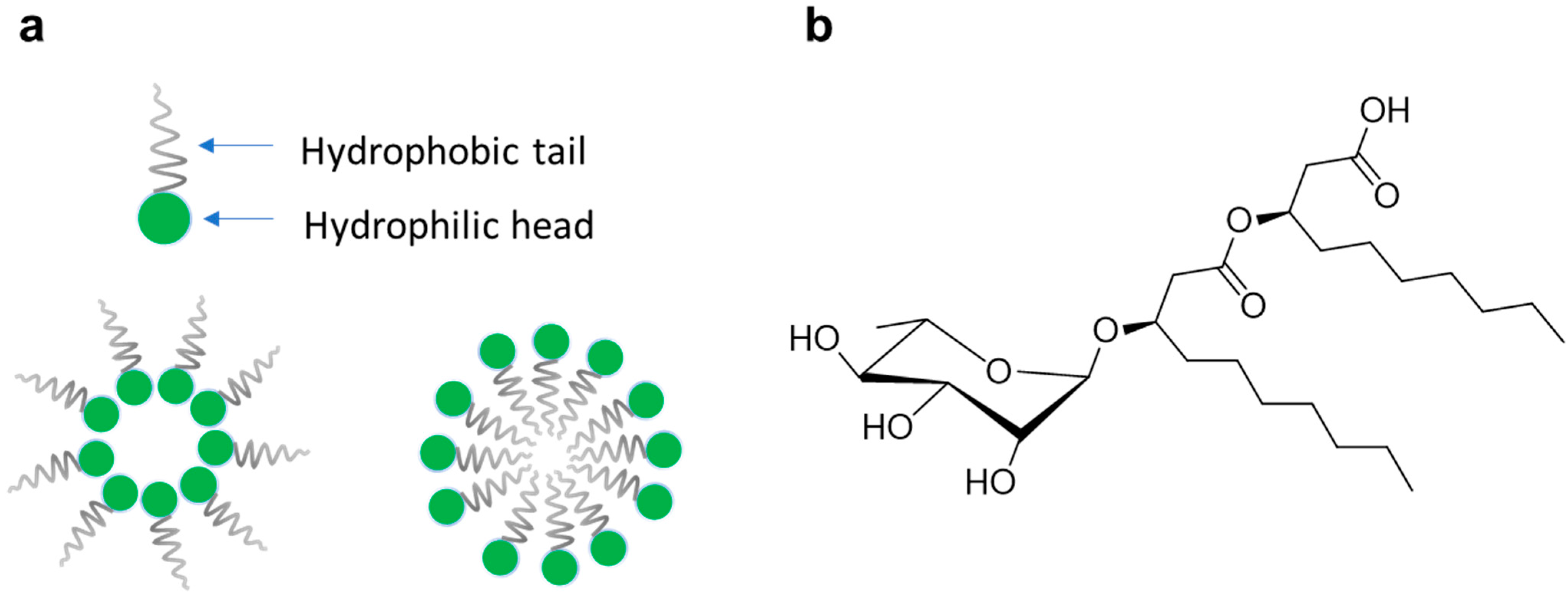
3.1. Chemical Characterization of M15RL and Commercial RLs Mixtures
3.2. Antimicrobial Activity of M15RL and Commercial RLs Mixtures
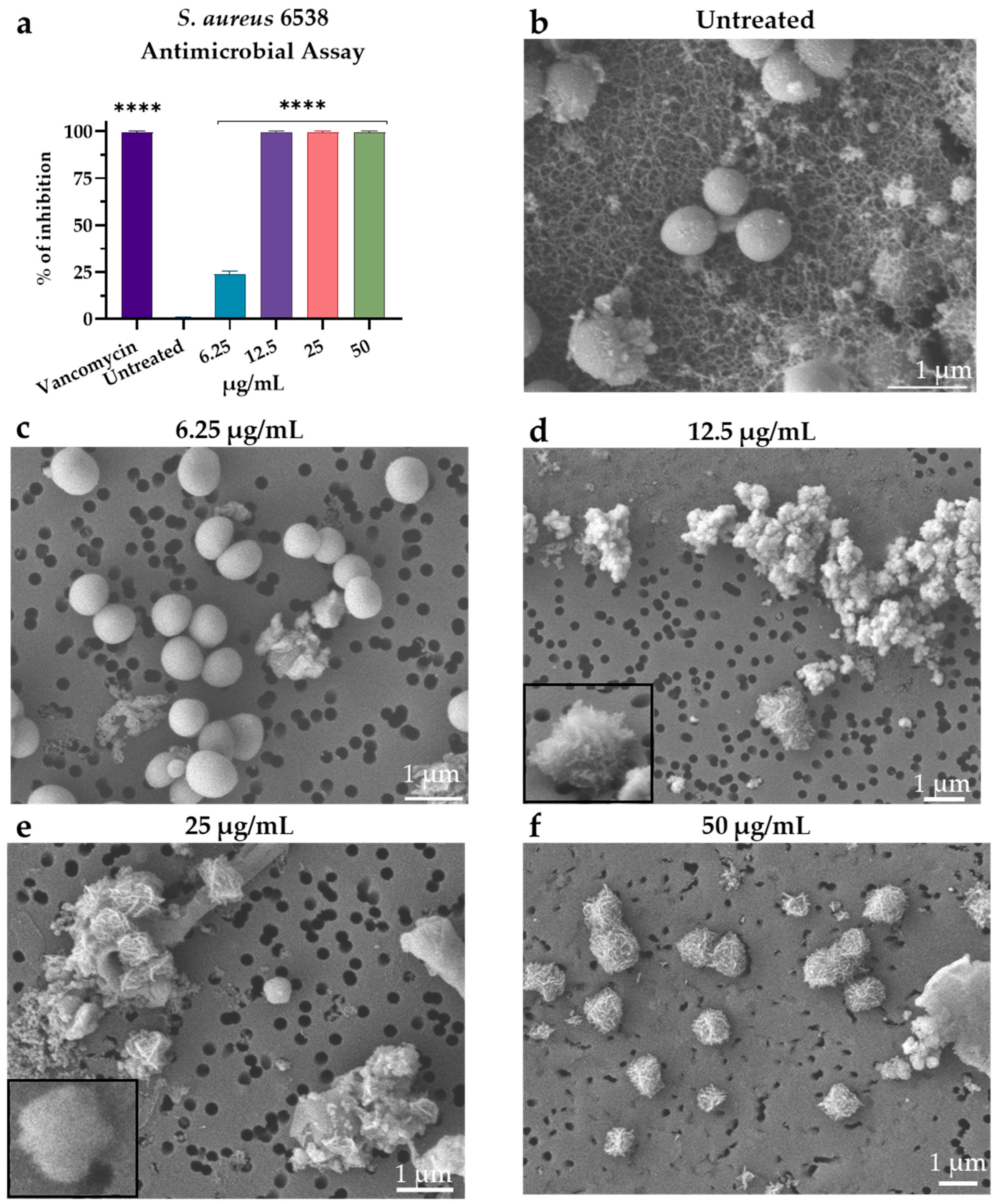
3.3. Visualization of M15RL Activity on S. aureus
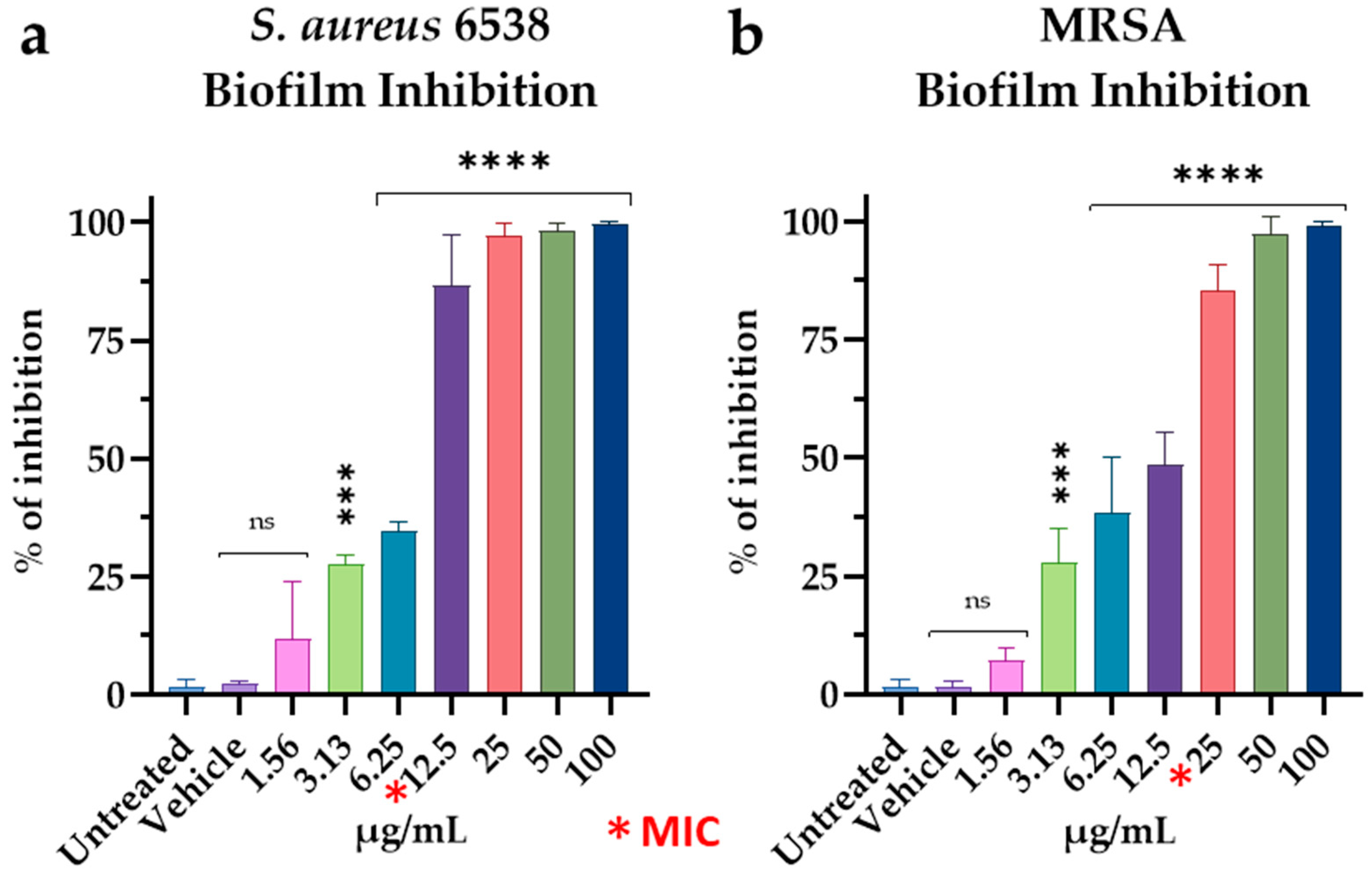
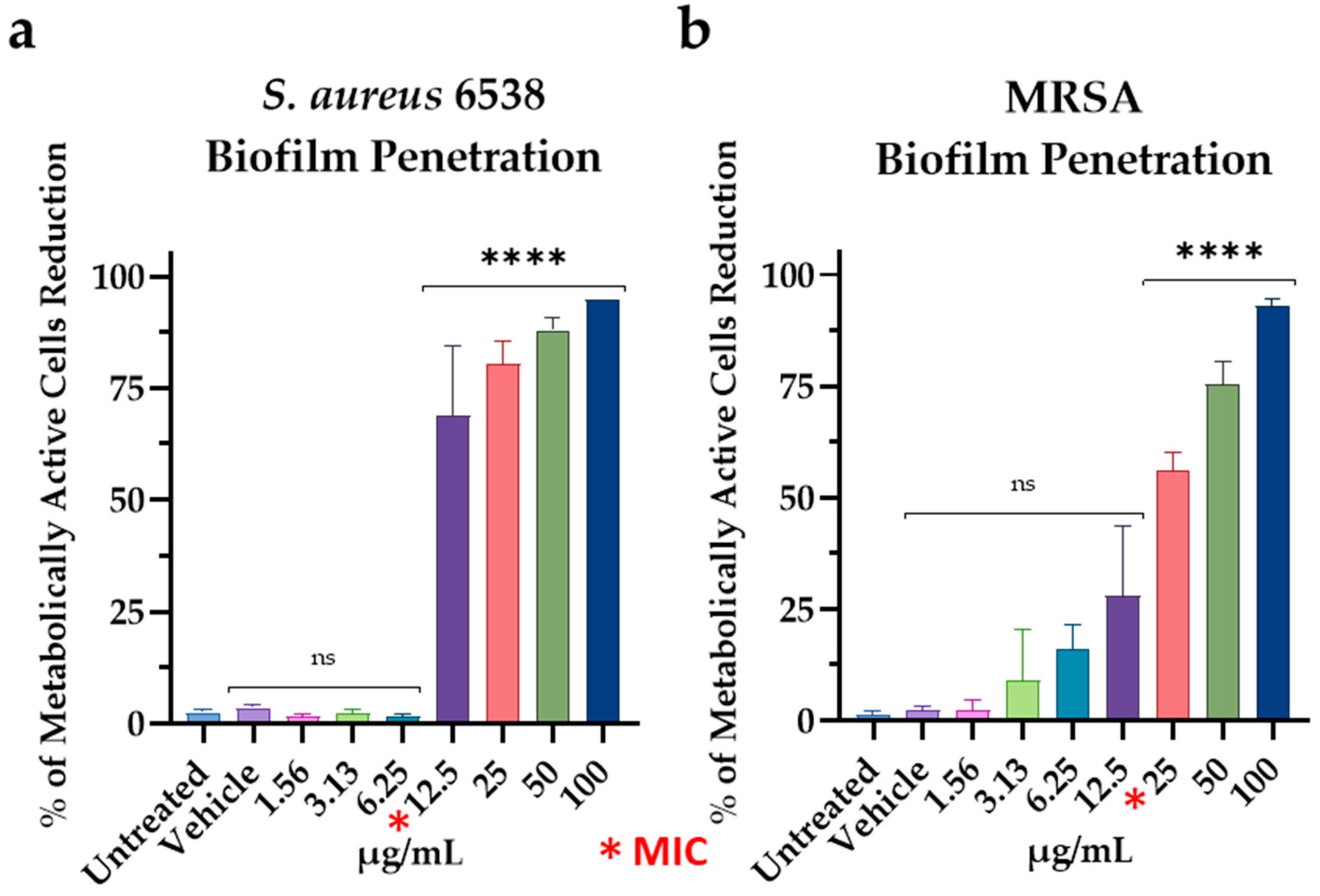
3.4. Effect of M15RL on S. aureus Biofilm
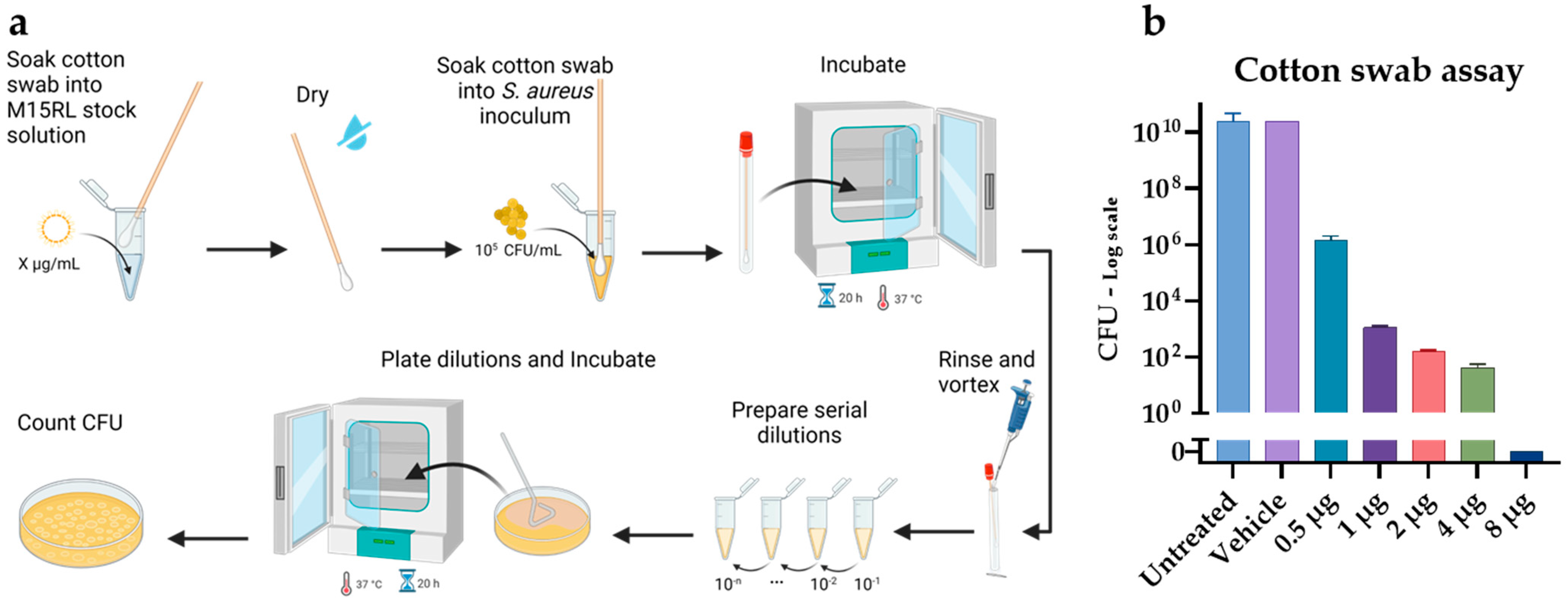
3.5. Antimicrobial Activity of Functionalized Textiles
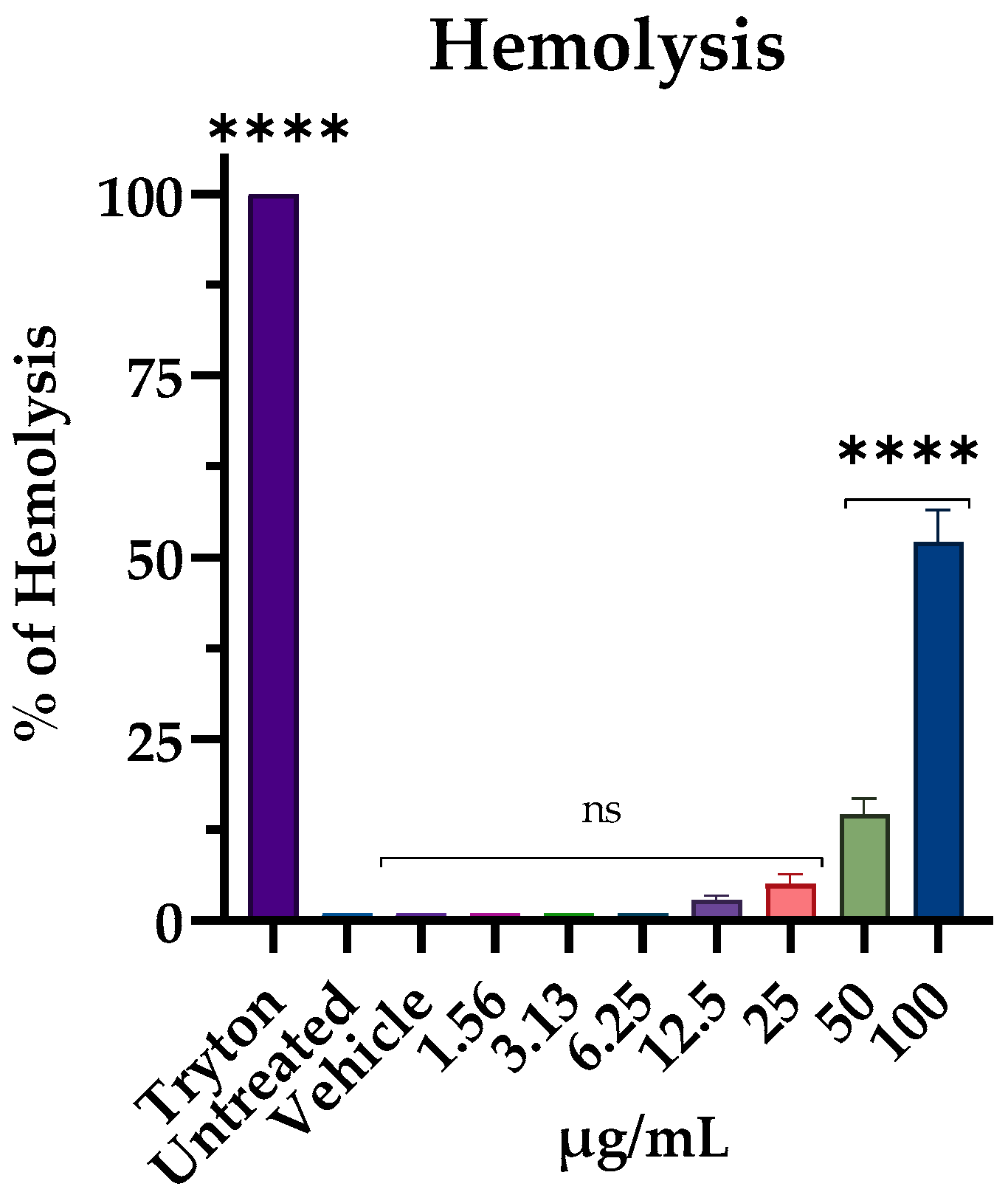
3.6. Hemolytic Activity
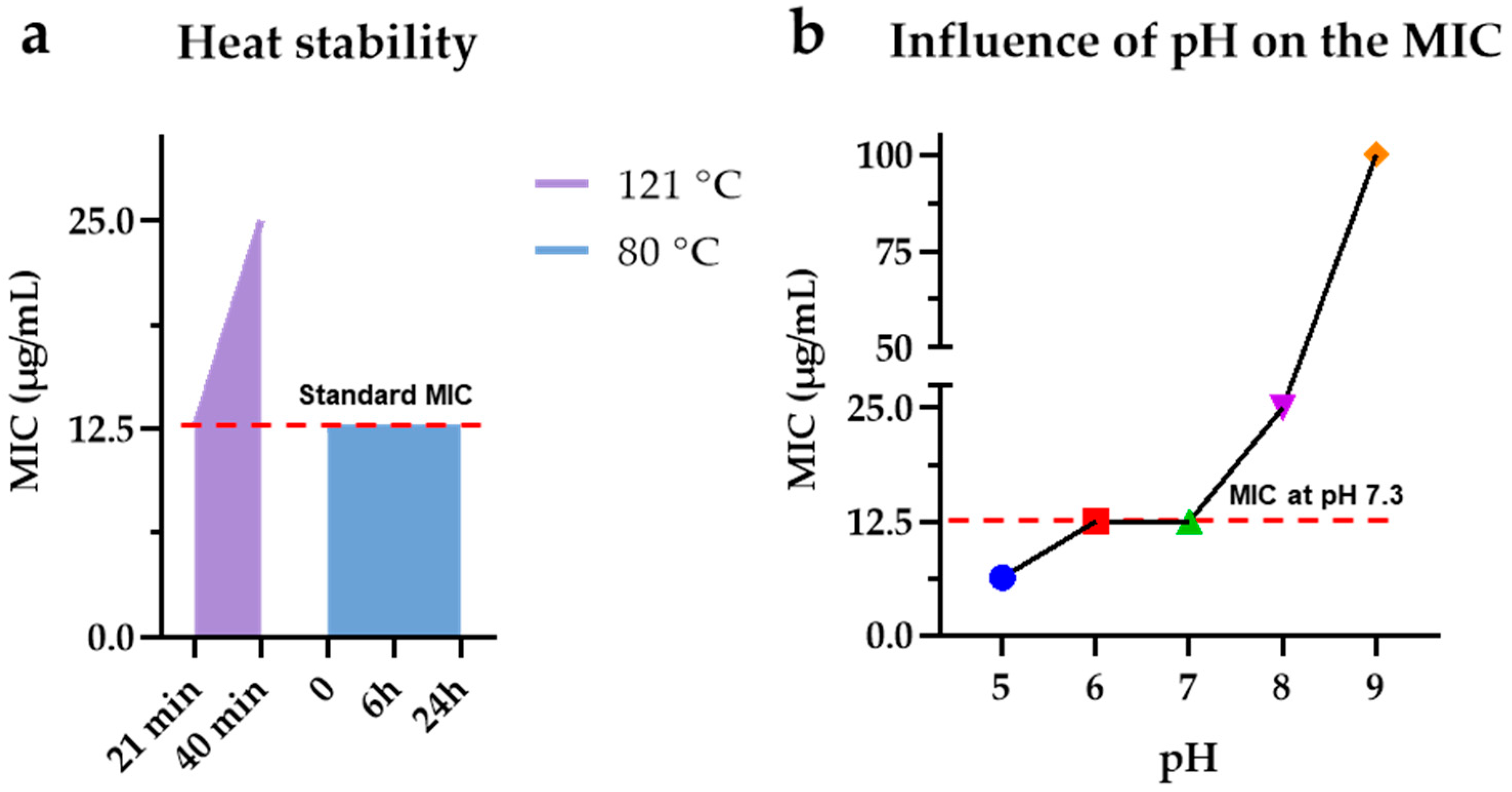
3.7. M15RL Stability to Harsh Conditions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De Oliveira, D.M.P.; Forde, B.M.; Kidd, T.J.; Harris, P.N.A.; Schembri, M.A.; Beatson, S.A.; Paterson, D.L.; Walker, M.J. Antimicrobial Resistance in ESKAPE Pathogens. Clin. Microbiol. Rev. 2020, 33, e00181-19. [Google Scholar] [CrossRef] [PubMed]
- WHO. WHO Publishes List of Bacteria for Which New Antibiotics Are Urgently Needed. Available online: https://www.who.int/news/item/27-02-2017-who-publishes-list-of-bacteria-for-which-new-antibiotics-are-urgently-needed (accessed on 20 September 2022).
- He, X.; Li, S.; Yin, Y.; Xu, J.; Gong, W.; Li, G.; Qian, L.; Yin, Y.; He, X.; Guo, T.; et al. Membrane Vesicles Are the Dominant Structural Components of Ceftazidime-Induced Biofilm Formation in an Oxacillin-Sensitive MRSA. Front. Microbiol. 2019, 10, 571. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.S.; de Lencastre, H.; Garau, J.; Kluytmans, J.; Malhotra-Kumar, S.; Peschel, A.; Harbarth, S. Methicillin-resistant Staphylococcus aureus. Nat. Rev. Dis. Prim. 2018, 4, 18033. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Zou, G.; Hari, T.P.A.; Wilt, I.K.; Zhu, W.; Galle, N.; Faizi, H.A.; Hendricks, G.L.; Tori, K.; Pan, W.; et al. A selective membrane-targeting repurposed antibiotic with activity against persistent methicillin-resistant Staphylococcus aureus. Proc. Natl. Acad. Sci. USA 2019, 116, 16529–16534. [Google Scholar] [CrossRef] [PubMed]
- Mlynarczyk-Bonikowska, B.; Kowalewski, C.; Krolak-Ulinska, A.; Marusza, W. Molecular Mechanisms of Drug Resistance in Staphylococcus aureus. Int. J. Mol. Sci. 2022, 23, 8088. [Google Scholar] [CrossRef] [PubMed]
- Miklasińska-Majdanik, M. Mechanisms of Resistance to Macrolide Antibiotics among Staphylococcus aureus. Antibiotics 2021, 10, 1406. [Google Scholar] [CrossRef]
- Simonetti, O.; Marasca, S.; Candelora, M.; Rizzetto, G.; Radi, G.; Molinelli, E.; Brescini, L.; Cirioni, O.; Offidani, A. Methicillin-resistant Staphylococcus aureus as a cause of chronic wound infections: Alternative strategies for management. AIMS Microbiol. 2022, 8, 125–137. [Google Scholar] [CrossRef]
- Tong, S.Y.C.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G., Jr. Staphylococcus aureus Infections: Epidemiology, Pathophysiology, Clinical Manifestations, and Management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef]
- Di Lodovico, S.; Bacchetti, T.; D’Ercole, S.; Covone, S.; Petrini, M.; Di Giulio, M.; Di Fermo, P.; Diban, F.; Ferretti, G.; Cellini, L. Complex Chronic Wound Biofilms Are Inhibited in vitro by the Natural Extract of Capparis spinose. Front. Microbiol. 2022, 13, 832919. [Google Scholar] [CrossRef]
- Hurdle, J.G.; O’neill, A.J.; Chopra, I.; Lee, R.E. Targeting bacterial membrane function: An underexploited mechanism for treating persistent infections. Nat. Rev. Microbiol. 2011, 9, 62–75. [Google Scholar] [CrossRef]
- Shireen, T.; Singh, M.; Dhawan, B.; Mukhopadhyay, K. Characterization of cell membrane parameters of clinical isolates of Staphylococcus aureus with varied susceptibility to alpha-melanocyte stimulating hormone. Peptides 2012, 37, 334–339. [Google Scholar] [CrossRef] [PubMed]
- Jahan, R.; Bodratti, A.M.; Tsianou, M.; Alexandridis, P. Biosurfactants, natural alternatives to synthetic surfactants: Physicochemical properties and applications. Adv. Colloid Interface Sci. 2019, 275, 102061. [Google Scholar] [CrossRef]
- Thakur, P.; Saini, N.K.; Thakur, V.K.; Gupta, V.K.; Saini, R.V.; Saini, A.K. Rhamnolipid the Glycolipid Biosurfactant: Emerging trends and promising strategies in the field of biotechnology and biomedicine. Microb. Cell Factories 2021, 20, 1. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Mawgoud, A.M.; Lépine, F.; Déziel, E. Rhamnolipids: Diversity of structures, microbial origins and roles. Appl. Microbiol. Biotechnol. 2010, 86, 1323–1336. [Google Scholar] [CrossRef] [PubMed]
- Christova, N.; Tuleva, B.; Kril, A.; Georgieva, M.; Konstantinov, S.; Terziyski, I.; Nikolova, B.; Stoineva, I. Chemical Structure and In Vitro Antitumor Activity of Rhamnolipids from Pseudomonas aeruginosa BN10. Appl. Biochem. Biotechnol. 2013, 170, 676–689. [Google Scholar] [CrossRef]
- VAN Gennip, M.; Christensen, L.D.; Alhede, M.; Phipps, R.; Jensen, P.; Christophersen, L.; Pamp, S.J.; Moser, C.; Mikkelsen, P.J.; Koh, A.Y.; et al. Inactivation of the rhlA gene in Pseudomonas aeruginosa prevents rhamnolipid production, disabling the protection against polymorphonuclear leukocytes. Apmis 2009, 117, 537–546. [Google Scholar] [CrossRef]
- Gomes, M.Z.D.V.; Nitschke, M. Evaluation of rhamnolipid and surfactin to reduce the adhesion and remove biofilms of individual and mixed cultures of food pathogenic bacteria. Food Control. 2012, 25, 441–447. [Google Scholar] [CrossRef]
- Haba, E.; Pinazo, A.; Jauregui, O.; Espuny, M.J.; Infante, M.R.; Manresa, A. Physicochemical characterization and antimicrobial properties of rhamnolipids produced byPseudomonas aeruginosa 47T2 NCBIM 40044. Biotechnol. Bioeng. 2002, 81, 316–322. [Google Scholar] [CrossRef]
- Yan, F.; Hu, H.; Lu, L.; Zheng, X. Rhamnolipids induce oxidative stress responses in cherry tomato fruit to Alternaria alternata. Pest Manag. Sci. 2015, 72, 1500–1507. [Google Scholar] [CrossRef]
- Skull, A. Evonik commercializes biosurfactants. Focus Surfactants 2016, 2016, 3–4. [Google Scholar] [CrossRef]
- Sekhon-Randhawa, K.K.; Rahman, P.K. Rhamnolipid biosurfactants—Past, present, and future scenario of global market. Front. Microbiol. 2014, 5, 454. [Google Scholar] [CrossRef]
- Shao, B.; Liu, Z.; Zhong, H.; Zeng, G.; Liu, G.; Yu, M.; Liu, Y.; Yang, X.; Li, Z.; Fang, Z.; et al. Effects of rhamnolipids on microorganism characteristics and applications in composting: A review. Microbiol. Res. 2017, 200, 33–44. [Google Scholar] [CrossRef]
- Buonocore, C.; Tedesco, P.; Vitale, G.A.; Esposito, F.P.; Giugliano, R.; Monti, M.C.; D’Auria, M.V.; De Pascale, D. Characterization of a New Mixture of Mono-Rhamnolipids Produced by Pseudomonas gessardii Isolated from Edmonson Point (Antarctica). Mar. Drugs 2020, 18, 269. [Google Scholar] [CrossRef] [PubMed]
- Giugliano, R.; Buonocore, C.; Zannella, C.; Chianese, A.; Esposito, F.P.; Tedesco, P.; De Filippis, A.; Galdiero, M.; Franci, G.; de Pascale, D. Antiviral Activity of the Rhamnolipids Mixture from the Antarctic Bacterium Pseudomonas gessardii M15 against Herpes Simplex Viruses and Coronaviruses. Pharmaceutics 2021, 13, 2121. [Google Scholar] [CrossRef] [PubMed]
- Palma Esposito, F.; Giugliano, R.; Della Sala, G.; Vitale, G.A.; Buonocore, C.; Ausuri, J.; Galasso, C.; Coppola, D.; Franci, G.; Galdiero, M.; et al. Combining OSMAC Approach and Untargeted Metabolomics for the Identification of New Glycolipids with Potent Antiviral Activity Produced by a Marine Rhodococcus. Int. J. Mol. Sci. 2021, 22, 9055. [Google Scholar] [CrossRef]
- Caso, A.; Esposito, G.; Della Sala, G.; Pawlik, J.R.; Teta, R.; Mangoni, A.; Costantino, V. Fast Detection of Two Smenamide Family Members Using Molecular Networking. Mar. Drugs 2019, 17, 618. [Google Scholar] [CrossRef] [PubMed]
- Franci, G.; Folliero, V.; Cammarota, M.; Zannella, C.; Sarno, F.; Schiraldi, C.; de Lera, A.R.; Altucci, L.; Galdiero, M. Epigenetic modulator UVI5008 inhibits MRSA by interfering with bacterial gyrase. Sci. Rep. 2018, 8, 13117. [Google Scholar] [CrossRef] [PubMed]
- Dell’Annunziata, F.; Folliero, V.; Palma, F.; Crudele, V.; Finamore, E.; Sanna, G.; Manzin, A.; De Filippis, A.; Galdiero, M.; Franci, G. Anthraquinone Rhein Exhibits Antibacterial Activity against Staphylococcus aureus. Appl. Sci. 2022, 12, 8691. [Google Scholar] [CrossRef]
- Sotirova, A.; Avramova, T.; Stoitsova, S.; Lazarkevich, I.; Lubenets, V.; Karpenko, E.; Galabova, D. The Importance of Rhamnolipid-Biosurfactant-Induced Changes in Bacterial Membrane Lipids of Bacillus subtilis for the Antimicrobial Activity of Thiosulfonates. Curr. Microbiol. 2012, 65, 534–541. [Google Scholar] [CrossRef]
- Escobar, I.; White, A.; Kim, W.; Mylonakis, E. New Antimicrobial Bioactivity against Multidrug-Resistant Gram-Positive Bacteria of Kinase Inhibitor IMD0354. Antibiotics 2020, 9, 665. [Google Scholar] [CrossRef]
- Chang, A.W.; Dowd, S.E.; Brackee, G.; Fralick, J.A.; Vediyappan, G. Inhibition of Staphylococcus aureus biofilm formation by gurmarin, a plant-derived cyclic peptide. Front. Cell. Infect. Microbiol. 2022, 12, 1017545. [Google Scholar] [CrossRef]
- Folliero, V.; Franci, G.; Dell’Annunziata, F.; Giugliano, R.; Foglia, F.; Sperlongano, R.; De Filippis, A.; Finamore, E.; Galdiero, M. Evaluation of Antibiotic Resistance and Biofilm Production among Clinical Strain Isolated from Medical Devices. Int. J. Microbiol. 2021, 2021, 9033278. [Google Scholar] [CrossRef]
- Zannella, C.; Chianese, A.; Greco, G.; Santella, B.; Squillaci, G.; Monti, A.; Doti, N.; Sanna, G.; Manzin, A.; Morana, A.; et al. Design of Three Residues Peptides against SARS-CoV-2 Infection. Viruses 2022, 14, 2103. [Google Scholar] [CrossRef] [PubMed]
- Lister, J.L.; Horswill, A.R. Staphylococcus aureus biofilms: Recent developments in biofilm dispersal. Front. Cell. Infect. Microbiol. 2014, 4, 178. [Google Scholar] [CrossRef]
- Boles, B.R.; Horswill, A.R. Staphylococcal biofilm disassembly. Trends Microbiol. 2011, 19, 449–455. [Google Scholar] [CrossRef] [PubMed]
- Allegrone, G.; Ceresa, C.; Rinaldi, M.; Fracchia, L. Diverse Effects of Natural and Synthetic Surfactants on the Inhibition of Staphylococcus aureus Biofilm. Pharmaceutics 2021, 13, 1172. [Google Scholar] [CrossRef] [PubMed]
- Samadi, N.; Abadian, N.; Ahmadkhaniha, R.; Amini, F.; Dalili, D.; Rastkari, N.; Safaripour, E.; Mohseni, F.A. Structural characterization and surface activities of biogenic rhamnolipid surfactants from Pseudomonas aeruginosa isolate MN1 and synergistic effects against methicillin-resistant Staphylococcus aureus. Folia Microbiol. 2012, 57, 501–508. [Google Scholar] [CrossRef]
- Ferreira, J.D.F.; Vieira, E.A.; Nitschke, M. The antibacterial activity of rhamnolipid biosurfactant is pH dependent. Food Res. Int. 2018, 116, 737–744. [Google Scholar] [CrossRef]
- Sana, S.; Datta, S.; Biswas, D.; Sengupta, D. Assessment of synergistic antibacterial activity of combined biosurfactants revealed by bacterial cell envelop damage. Biochim. Biophys. Acta (BBA)-Biomembr. 2018, 1860, 579–585. [Google Scholar] [CrossRef] [PubMed]
- Sotirova, A.V.; Spasova, D.I.; Galabova, D.N.; Karpenko, E.; Shulga, A. Rhamnolipid–Biosurfactant Permeabilizing Effects on Gram-Positive and Gram-Negative Bacterial Strains. Curr. Microbiol. 2008, 56, 639–644. [Google Scholar] [CrossRef]
- Rogiers, G.; Kebede, B.T.; Van Loey, A.; Michiels, C.W. Membrane fatty acid composition as a determinant of Listeria monocytogenes sensitivity to trans -cinnamaldehyde. Res. Microbiol. 2017, 168, 536–546. [Google Scholar] [CrossRef] [PubMed]
- Nikolic, P.; Mudgil, P.; Harman, D.G.; Whitehall, J. Untargeted lipidomic differences between clinical strains of methicillin-sensitive and methicillin-resistant Staphylococcus aureus. Infect. Dis. 2022, 54, 497–507. [Google Scholar] [CrossRef] [PubMed]
- Schierle, C.F.; De La Garza, M.; Mustoe, T.A.; Galiano, R.D. Staphylococcal biofilms impair wound healing by delaying reepithelialization in a murine cutaneous wound model. Wound Repair Regen. 2009, 17, 354–359. [Google Scholar] [CrossRef] [PubMed]
- Doebbeling, B.N. The epidemiology of methicillin-resistant Staphylococcus aureus colonisation and infection. J. Chemother. 1995, 7 (Suppl. S3), 99–103. [Google Scholar] [PubMed]
- Aleksic, I.; Petkovic, M.; Jovanovic, M.; Milivojevic, D.; Vasiljevic, B.; Nikodinovic-Runic, J.; Senerovic, L. Anti-biofilm Properties of Bacterial Di-Rhamnolipids and Their Semi-Synthetic Amide Derivatives. Front. Microbiol. 2017, 8, 2454. [Google Scholar] [CrossRef]
- Zhong, H.; Jiang, Y.; Zeng, G.; Liu, Z.; Liu, L.; Liu, Y.; Yang, X.; Lai, M.; He, Y. Effect of low-concentration rhamnolipid on adsorption of Pseudomonas aeruginosa ATCC 9027 on hydrophilic and hydrophobic surfaces. J. Hazard. Mater. 2015, 285, 383–388. [Google Scholar] [CrossRef]
- Ceresa, C.; Tessarolo, F.; Maniglio, D.; Tambone, E.; Carmagnola, I.; Fedeli, E.; Caola, I.; Nollo, G.; Chiono, V.; Allegrone, G.; et al. Medical-Grade Silicone Coated with Rhamnolipid R89 Is Effective against Staphylococcus spp. Biofilms. Molecules 2019, 24, 3843. [Google Scholar] [CrossRef]
- Dardouri, M.; Bettencourt, A.; Martin, V.; Carvalho, F.A.; Colaço, B.; Gama, A.; Ramstedt, M.; Santos, N.C.; Fernandes, M.H.; Gomes, P.S.; et al. Assuring the Biofunctionalization of Silicone Covalently Bonded to Rhamnolipids: Antibiofilm Activity and Biocompatibility. Pharmaceutics 2022, 14, 1836. [Google Scholar] [CrossRef]
- Dardouri, M.; Bettencourt, A.; Martin, V.; Carvalho, F.A.; Santos, C.; Monge, N.; Santos, N.C.; Fernandes, M.H.; Gomes, P.S.; Ribeiro, I.A. Using plasma-mediated covalent functionalization of rhamnolipids on polydimethylsiloxane towards the antimicrobial improvement of catheter surfaces. Biomater. Adv. 2021, 134, 112563. [Google Scholar] [CrossRef]
- Dardouri, M.; Aljnadi, I.M.; Deuermeier, J.; Santos, C.; Costa, F.; Martin, V.; Fernandes, M.H.; Gonçalves, L.; Bettencourt, A.; Gomes, P.S.; et al. Bonding antimicrobial rhamnolipids onto medical grade PDMS: A strategy to overcome multispecies vascular catheter-related infections. Colloids Surf. B Biointerfaces 2022, 217, 112679. [Google Scholar] [CrossRef]
- Kong, S.; Shen, C.; Li, Y.; Meng, Q. Rhamnolipids Sustain Unchanged Surface Activities during Decomposition in Alkaline Solutions. ACS Omega 2021, 6, 15750–15755. [Google Scholar] [CrossRef] [PubMed]
- Tyeb, S.; Kumar, N.; Kumar, A.; Verma, V. pH modulating agar dressing for chronic wounds. Soft Mater. 2022, 20, 379–393. [Google Scholar] [CrossRef]
- Lovaglio, R.B.; dos Santos, F.J.; Jafelicci, M.; Contiero, J. Rhamnolipid emulsifying activity and emulsion stability: pH rules. Colloids Surf. B Biointerfaces 2011, 85, 301–305. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rt (min.) | Rhamnolipids 1 | M15RL (%) 2 | MDRL (%) 2 | DDRL (%) 2 | Notes |
|---|---|---|---|---|---|
| 6.1 | Rha-Rha-C10 | - | 0.6 | 1 | |
| 7.8 | Rha-C10 | - | 1.6 | 1.2 | |
| 12.5 | Rha-Rha-C11 | - | 0.5 | 0.2 | |
| 14.5 | Rha-C11 | - | 1.3 | 0.2 | |
| 14.8 | Rha-C12:1 | 0.2 | - | - | |
| 17.7 | Rha-C12 | 0.2 | - | - | |
| 23.3 | Rha-Rha-C8-C9 Rha-Rha-C9-C8 | - | - | 0.1 | |
| 25.0 | Rha-Rha-C12:1-C10;O Rha-Rha-C10-C12:1;O | - | - | 0.5 | |
| 25.5 | Rha-C14 | 0.3 | - | - | |
| 26.7 | Rha-Rha-C8-C10 Rha-Rha-C10-C8 | - | 1.8 | 1.2 | |
| 29.5 | Rha-Rha-C9-C10 Rha-Rha-C10-C9 Rha-Rha-C8-C11 | - | 0.1 | 0.9 | |
| 30.1 | Rha-C8-C10 Rha-C10-C8 | 4.2 | 6.9 | 1.3 | in M15RL, only Rha-C8-C10 is present |
| 31.4 | Rha-C10;O-C13;O | - | - | 0.7 | |
| 33.1 | Rha-Rha-C10-C10 Rha-Rha-C8-C12 | - | 15.2 | 19.6 | Rha-Rha-C8-C12 present only in DDRL |
| 33.7 | Rha-C10-C9 Rha-C8-C11 | 0.7 | 0.3 | 0.2 | in M15RL, different isomers are present, i.e., Rha-C8-C11 and Rha-C9-C10 |
| 34.1 | Rha-C12:1-C8 Rha-C10:1-C10 Rha-C10-C10:1 Rha-C8-C12:1 | 0.5 | 3.4 | 0.5 | in M15RL, Rha-C10-C10:1 is missing |
| 35.8 | Rha-Rha-C10-C11 Rha-Rha-C11-C10 | - | 0.4 | 5.1 | |
| 36.3 | Rha-Rha-C12:1-C10 Rha-Rha-C10-C12:1 | - | 3.3 | 6.9 | |
| 37.5 | Rha-C10-C10 Rha-C8-C12 | 19.3 | 35.0 | 20.8 | in M15RL, there is also Rha-C12-C8 |
| 39.1 | Rha-Rha-C12-C10 Rha-Rha-C10-C12 | - | 7.7 | 18.2 | |
| 40.8 | Rha-C10-C11 Rha-C11-C10 | 3.8 | 0.4 | 1.1 | in M15RL, there is also Rha-C12-C9 |
| 41.6 | Rha-C12:1-C10 Rha-C10-C12:1 Rha-C8-C14:1 | 29.8 | 8.7 | 4.7 | in M15RL, Rha-C8-C14:1 is missing |
| 42.2 | Rha-Rha-C12-C11 | - | - | 0.7 | |
| 41.5 42.9 | Rha-Rha-C14:1-C10 Rha-Rha-C10-C14:1 Rha-Rha-C12:1-C12 Rha-Rha-C12-C12:1 | - | 0.6 | 2.9 | |
| 44.6 | Rha-C12-C10 Rha-C10-C12 | 29.8 | 10.0 | 8.3 | |
| 45.6 | Rha-C13:1-C10 Rha-C12:1-C11 | 0.7 | - | - | |
| 45.7 | Rha-Rha-C14-C10 Rha-Rha-C12-C12 Rha-Rha-C10-C14 | - | 0.9 | 3.3 | |
| 46.1 | Rha-C12:1-C12:1 | 1.2 | - | - | |
| 47.4 49.0 | Rha-C14:1-C10 Rha-C10-C14:1 Rha-C12:1-C12 Rha-C12-C12:1 | 5.6 | 0.9 | 0.2 | in M15RL, Rha-C10-C14:1 is missing |
| 48.2 | Rha-C13-C10 Rha-C12-C11 Rha-C11-C12 | 0.9 | - | - | |
| 52.4 | Rha-C14-C10 Rha-C10-C14 Rha-C12-C12 | 2.4 | 0.5 | 0.1 | in M15RL, Rha-C10-C14 is missing |
| 54.2 | Rha-C16:1-C10 Rha-C14:1-C12 | 0.4 | - | - |
| M15RL | MDRL | DDRL | |||||
|---|---|---|---|---|---|---|---|
| S. aureus Strains | MIC (µg/mL) | MBC (µg/mL) | MIC (µg/mL) | MBC (µg/mL) | MIC (µg/mL) | MBC (µg/mL) | |
| S. aureus 6538 | 12.5 | 12.5 | 50 | 100 | 50 | 50 | |
| S. aureus 6538p | 25 | 25 | 100 | - | 100 | - | |
| Clinical isolates | MRSA | 25 | 25 | >100 | - | >100 | - |
| MSSA | 25 | 50 | 100 | - | 100 | - | |
| β-LPSA | 12.5 | 12.5 | 50 | 100 | >100 | - | |
| QRSA | 25 | 50 | 100 | - | >100 | - | |
| VRSA | 50 | 50 | >100 | - | >100 | - | |
| MLSB | 12.5 | 12.5 | 50 | 100 | 100 | - | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buonocore, C.; Giugliano, R.; Della Sala, G.; Palma Esposito, F.; Tedesco, P.; Folliero, V.; Galdiero, M.; Franci, G.; de Pascale, D. Evaluation of Antimicrobial Properties and Potential Applications of Pseudomonas gessardii M15 Rhamnolipids towards Multiresistant Staphylococcus aureus. Pharmaceutics 2023, 15, 700. https://doi.org/10.3390/pharmaceutics15020700
Buonocore C, Giugliano R, Della Sala G, Palma Esposito F, Tedesco P, Folliero V, Galdiero M, Franci G, de Pascale D. Evaluation of Antimicrobial Properties and Potential Applications of Pseudomonas gessardii M15 Rhamnolipids towards Multiresistant Staphylococcus aureus. Pharmaceutics. 2023; 15(2):700. https://doi.org/10.3390/pharmaceutics15020700
Chicago/Turabian StyleBuonocore, Carmine, Rosa Giugliano, Gerardo Della Sala, Fortunato Palma Esposito, Pietro Tedesco, Veronica Folliero, Massimiliano Galdiero, Gianluigi Franci, and Donatella de Pascale. 2023. "Evaluation of Antimicrobial Properties and Potential Applications of Pseudomonas gessardii M15 Rhamnolipids towards Multiresistant Staphylococcus aureus" Pharmaceutics 15, no. 2: 700. https://doi.org/10.3390/pharmaceutics15020700
APA StyleBuonocore, C., Giugliano, R., Della Sala, G., Palma Esposito, F., Tedesco, P., Folliero, V., Galdiero, M., Franci, G., & de Pascale, D. (2023). Evaluation of Antimicrobial Properties and Potential Applications of Pseudomonas gessardii M15 Rhamnolipids towards Multiresistant Staphylococcus aureus. Pharmaceutics, 15(2), 700. https://doi.org/10.3390/pharmaceutics15020700