


Seroprevalence, Genetic Characteristics, and Pathogenicity of Korean Porcine Sapeloviruses

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. RT-PCR and Sequencing of the PSV Genome
2.3. Cell Lines and Virus Isolation
2.4. Serum Neutralization
2.5. Virus Growth Kinetics
2.6. Phylogenetic and Recombination Analysis
2.7. Experimental Infection of Piglets and Fattening Pigs
2.8. Statistical Analysis
3. Results
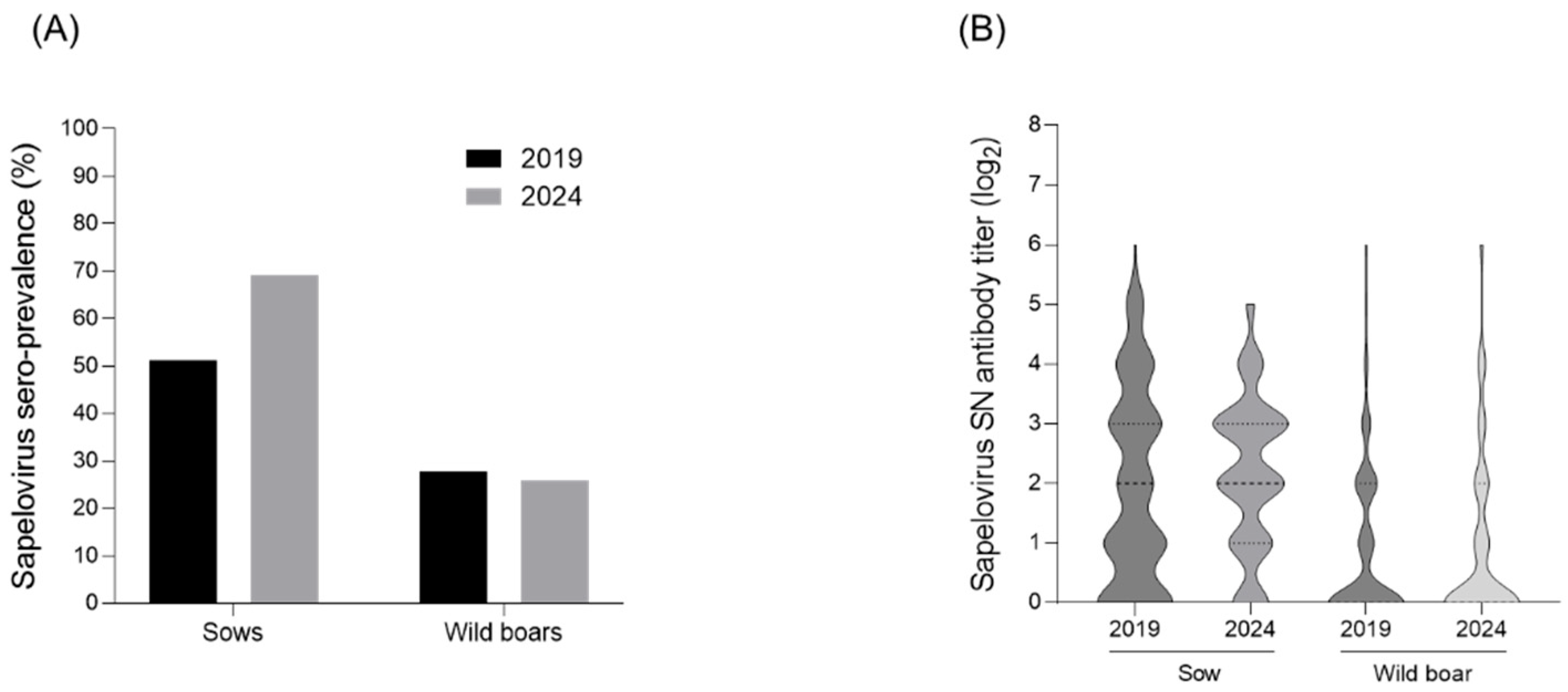
3.1. Seroprevalence of PSV
3.2. Single or Co-Infection Analysis of PSV
3.3. Isolation of PSV
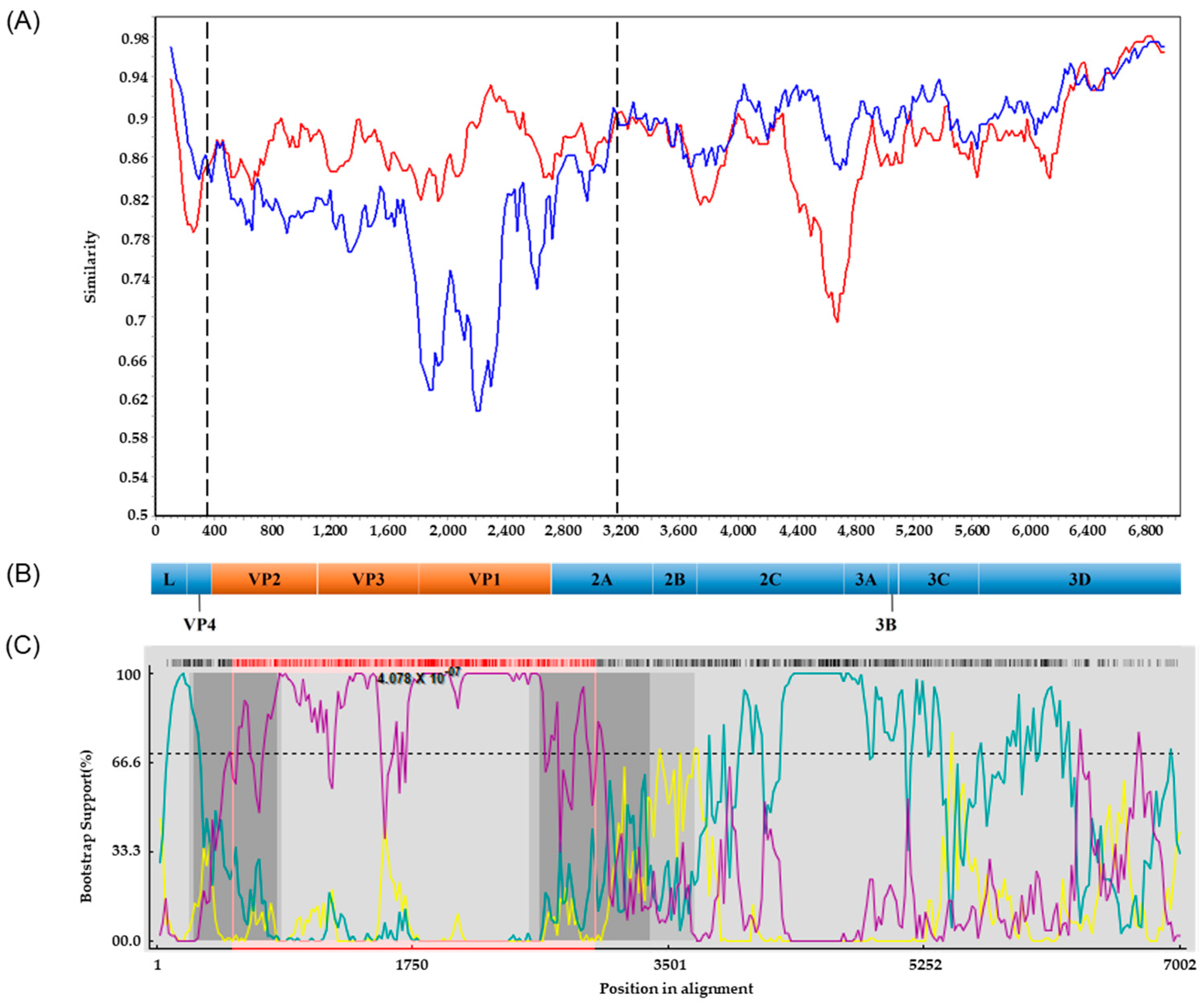
3.4. Phylogenetic Analysis of PSV
3.5. Recombination Analyses
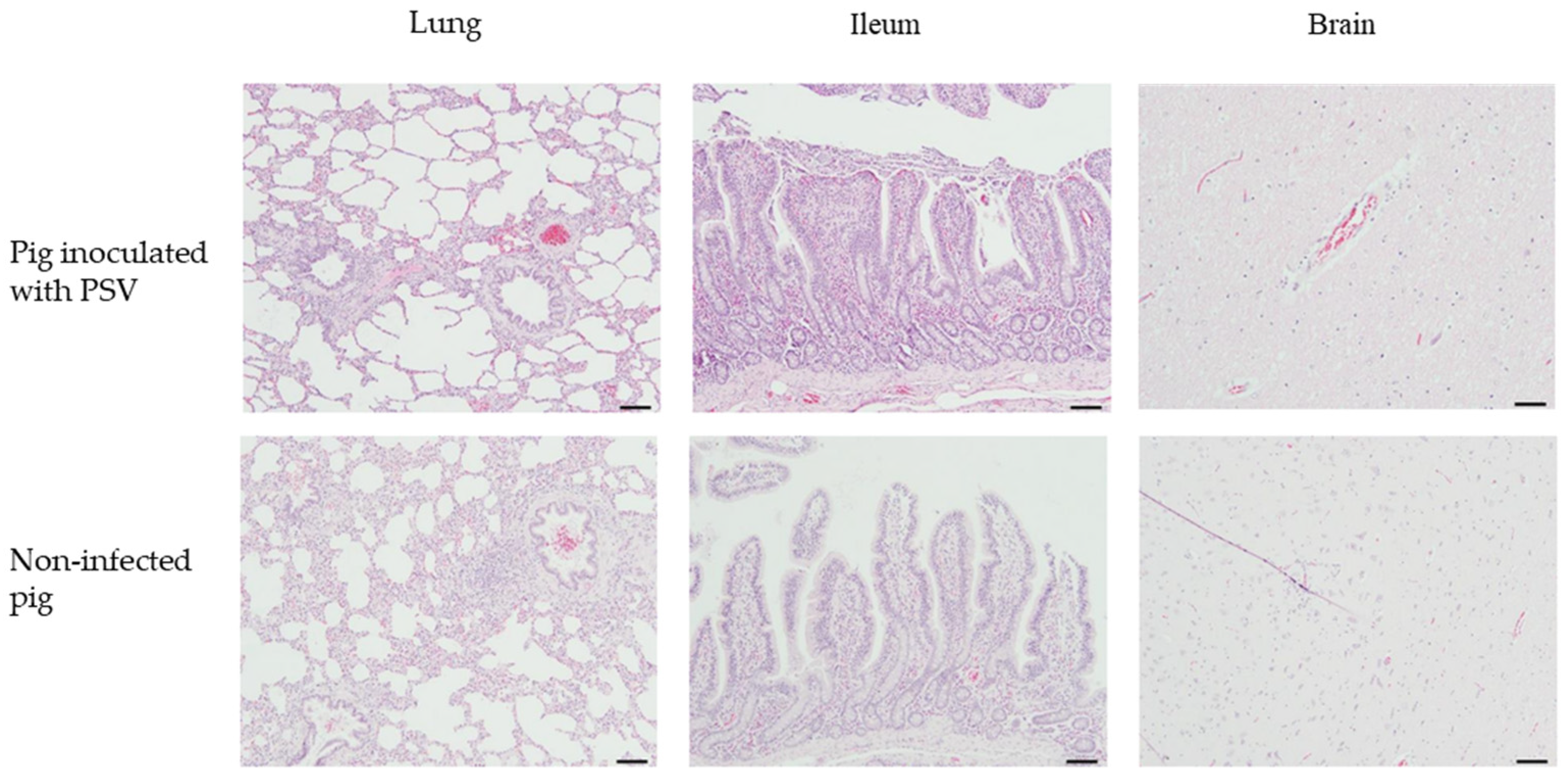
3.6. Clinical Signs and Histopathological Examination
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Krumbholz, A.; Dauber, M.; Henke, A.; Birch-Hirschfeld, E.; Knowles, N.J.; Stelzner, A.; Zell, R. Sequencing of porcine enterovirus groups II and III reveals unique features of both virus groups. J. Virol. 2002, 76, 5813–5821. [Google Scholar] [CrossRef] [PubMed]
- Zell, R.; Dauber, M.; Krumbholz, A.; Henke, A.; Birch-Hirschfeld, E.; Stelzner, A.; Prager, D.; Wurm, R. Porcine teschoviruses comprise at least eleven distinct serotypes: Molecular and evolutionary aspects. J. Virol. 2001, 75, 1620–1631. [Google Scholar] [CrossRef]
- Kaku, Y.; Sarai, A.; Murakami, Y. Genetic reclassification of porcine enteroviruses. J. Gen. Virol. 2001, 82, 417–424. [Google Scholar] [CrossRef]
- Zell, R.; Delwart, E.; Gorbalenya, A.; Hovi, T.; King, A.; Knowles, N.; Lindberg, A.M.; Pallansch, M.; Palmenberg, A.; Reuter, G.; et al. ICTV Virus TaxonomyProfile: Picornaviridae. J. Gen. Virol. 2017, 98, 2421–2422. [Google Scholar] [CrossRef] [PubMed]
- Tseng, C.H.; Tsai, H.J. Sequence analysis of a duck picornavirus isolate indicates that it together with porcine enterovirus type 8 and simian picornavirus type 2 should be assigned to a new picornavirus genus. Virus Res. 2007, 129, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Knowles, N.; Buckley, L.; Pereira, H. Classification of porcine enteroviruses by antigenic analysis and cytopathic effects in tissue culture: Description of 3 new serotypes. Arch. Virol. 1979, 62, 201–208. [Google Scholar] [CrossRef]
- Yang, T.; Yu, X.; Yan, M.; Luo, B.; Li, R.; Qu, T.; Luo, Z.; Ge, M.; Zhao, D. Molecular characterization of Porcine sapelovirus in Hunan, China. J. Gen. Virol. 2017, 98, 2738–2747. [Google Scholar] [CrossRef]
- Son, K.Y.; Kim, D.S.; Kwon, J.; Choi, J.S.; Kang, M.I.; Belsham, G.J.; Cho, K.O. Full-length genomic analysis of Korean porcine Sapelovirus strains. PLoS ONE 2014, 9, e107860. [Google Scholar] [CrossRef]
- Li, Y.; Du, L.; Jin, T.; Cheng, Y.; Zhang, X.; Jiao, S.; Huang, T.; Zhang, Y.; Yan, Y.; Gu, J. Characterization and epidemiological survey of porcine sapelovirus in China. Vet. Microbiol. 2019, 232, 13–21. [Google Scholar] [CrossRef]
- Lan, D.; Ji, W.; Yang, S.; Cui, L.; Yang, Z.; Yuan, C.; Hua, X. Isolation and characterization of the first Chinese porcine sapelovirus strain. Arch. Virol. 2011, 156, 1567–1574. [Google Scholar] [CrossRef]
- Arruda, P.; Arruda, B.; Schwartz, K.; Vannucci, F.; Resende, T.; Rovira, A.; Sundberg, P.; Nietfeld, J.; Hause, B. Detection of a novel sapelovirus in central nervous tissue of pigs with polioencephalomyelitis in the USA. Transbound. Emerg. Dis. 2017, 64, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Racaniello, V.R. Picornaviridae: The Viruses and Their Replication. In Fields Virology, 6th ed.; Knipe, D.M., Howley, P.M., Cohen, J.I., Griffin, D.E., Lamb, R.A., Martin, M.A., Racaniello, V.R., Roizman, B., Eds.; Wolters Kluwer/Lippincott Williams, & Winlkins: Philadelphia, PA, USA, 2013; Volume 2, pp. 453–489. [Google Scholar]
- Schock, A.; Gurrala, R.; Fuller, H.; Foyle, L.; Dauber, M.; Martelli, F.; Scholes, S.; Roberts, L.; Steinbach, F.; Dastjerdi, A. Investigation into an outbreak of encephalomyelitis caused by a neuroinvasive porcine sapelovirus in the United Kingdom. Vet. Microbiol. 2014, 172, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Zhu, P.; Li, Z.; Li, Z.; Meng, L.; Liu, P.; Sun, X.; Yang, Q.; Song, J. First Isolation and Characterization of Three Strains of Porcine Sapelovirus in Yunnan Province, China. Viruses 2025, 17, 505. [Google Scholar] [CrossRef]
- Chen, Q.Y.; Sun, Z.H.; Che, Y.L.; Chen, R.J.; Wu, X.M.; Wu, R.J.; Wang, L.B.; Zhou, L.J. High prevalence, genetic diversity, and recombination of porcine Sapelovirus in pig farms in Fujian, Southern China. Viruses 2023, 15, 1751. [Google Scholar] [CrossRef]
- Ibrahim, Y.M.; Zhang, W.; Werid, G.M.; Zhang, H.; Feng, Y.; Pan, Y.; Zhang, L.; Li, C.; Lin, H.; Chen, H. Isolation, characterization, and molecular detection of porcine sapelovirus. Viruses 2022, 14, 349. [Google Scholar] [CrossRef]
- Chen, J.; Suo, X.; Cao, L.; Yuan, C.; Shi, L.; Duan, Y.; Zheng, H.; Wang, Q. Virome analysis for identification of a novel porcine sapelovirus isolated in western China. Microbiol. Spectr. 2022, 10, e0180122. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Zhang, L.; Lu, Y.; Guo, M.; Zhang, Z.; Lin, A. Characterization of porcine sapelovirus prevalent in western Jiangxi, China. BMC Vet. Res. 2021, 17, 273. [Google Scholar] [CrossRef]
- Forman, A.; Pass, D.; Connaughton, I. The characterisation and pathogenicity of porcine enteroviruses isolated in Victoria. Aust. Vet. J. 1982, 58, 136–142. [Google Scholar] [CrossRef]
- Dunne, H.; Wang, J.; Ammerman, E.H. Classification of North American porcine enteroviruses: A comparison with European and Japanese strains. Infect. Immun. 1971, 4, 619–631. [Google Scholar] [CrossRef]
- Kumari, S.; Ray, P.; Singh, R.; Desingu, P.; Varshney, R.; Saikumar, G. Pathological and molecular investigation of porcine sapelovirus infection in naturally affected Indian pigs. Microb. Pathog. 2019, 127, 320–325. [Google Scholar] [CrossRef]
- Kim, D.-S.; Kang, M.-I.; Son, K.-Y.; Bak, G.-Y.; Park, J.-G.; Hosmillo, M.; Seo, J.-Y.; Kim, J.-Y.; Alfajaro, M.M.; Soliman, M. Pathogenesis of Korean Sapelovirus A in piglets and chicks. J. Gen. Virol. 2016, 97, 2566–2574. [Google Scholar] [CrossRef] [PubMed]
- Harima, H.; Kajihara, M.; Simulundu, E.; Bwalya, E.; Qiu, Y.; Isono, M.; Okuya, K.; Gonzalez, G.; Yamagishi, J.; Hang’ombe, B.M. Genetic and biological diversity of porcine Sapeloviruses prevailing in Zambia. Viruses 2020, 12, 180. [Google Scholar] [CrossRef]
- Vilar, M.; Peralta, B.; García-Bocanegra, I.; Simon-Grifé, M.; Bensaid, A.; Casal, J.; Segalés, J.; Pina-Pedrero, S. Distribution and genetic characterization of Enterovirus G and Sapelovirus A in six Spanish swine herds. Virus Res. 2016, 215, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Donin, D.G.; de Arruda Leme, R.; Alfieri, A.F.; Alberton, G.C.; Alfieri, A.A. First report of Porcine teschovirus (PTV), Porcine sapelovirus (PSV) and Enterovirus G (EV-G) in pig herds of Brazil. Trop. Anim. Health Prod. 2014, 46, 523–528. [Google Scholar] [CrossRef]
- Cano-Gómez, C.; García-Casado, M.A.; Soriguer, R.; Palero, F.; Jiménez-Clavero, M.A. Teschoviruses and sapeloviruses in faecal samples from wild boar in Spain. Vet. Microbiol. 2013, 165, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Prodělalová, J. The survey of porcine teschoviruses, sapeloviruses and enteroviruses B infecting domestic pigs and wild boars in the Czech Republic between 2005 and 2011. Infect. Genet. Evol. 2012, 12, 1447–1451. [Google Scholar] [CrossRef]
- Lamont, P.; Betts, A. Studies on enteroviruses of the Pig—IV: The isolation in tissue culture of a possible enteric cytopathogenic swine orphan (ECSO) virus (V 13) from the faeces of a pig. Res. Vet. Sci. 1960, 1, 152–161. [Google Scholar] [CrossRef]
- Ray, P.; Desingu, P.; Kumari, S.; John, J.; Sethi, M.; Sharma, G.; Pattnaik, B.; Singh, R.; Saikumar, G. Porcine sapelovirus among diarrhoeic piglets in India. Transbound. Emerg. Dis. 2018, 65, 261–263. [Google Scholar] [CrossRef]
- Yang, T.; Li, R.; Peng, W.; Ge, M.; Luo, B.; Qu, T.; Yu, X. First isolation and genetic characteristics of porcine sapeloviruses in Hunan, China. Arch. Virol. 2017, 162, 1589–1597. [Google Scholar] [CrossRef]
- Boros, Á.; László, Z.; Pankovics, P.; Marosi, A.; Albert, M.; Cságola, A.; Bíró, H.; Fahsbender, E.; Delwart, E.; Reuter, G. High prevalence, genetic diversity and a potentially novel genotype of Sapelovirus A (Picornaviridae) in enteric and respiratory samples in Hungarian swine farms. J. Gen. Virol. 2020, 101, 609–621. [Google Scholar] [CrossRef]
- Stäubli, T.; Rickli, C.I.; Torgerson, P.R.; Fraefel, C.; Lechmann, J. Porcine teschovirus, sapelovirus, and enterovirus in Swiss pigs: Multiplex RT-PCR investigation of viral frequencies and disease association. J. Vet. Diagn. Investig. 2021, 33, 864–874. [Google Scholar] [CrossRef] [PubMed]
- Yue, F.; Cui, S.; Zhang, C.; Yoon, K.J. A multiplex PCR for rapid and simultaneous detection of porcine circovirus type 2, porcine parvovirus, porcine pseudorabies virus, and porcine reproductive and respiratory syndrome virus in clinical specimens. Virus Genes 2009, 38, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, H.; Taira, O.; Hirai, T.; Takeuchi, H.; Nagao, A.; Ishikawa, Y.; Tuchiya, K.; Nunoya, T.; Ueda, S. Multiplex PCR and multiplex RT-PCR for inclusive detection of major swine DNA and RNA viruses in pigs with multiple infections. J. Virol. Methods 2009, 160, 210–214. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Q.; Si, L.; Gao, J.; Yuan, J.; Xia, L.; Hu, H. Isolation and Characterization of Porcine Sapelovirus from the PDCoV-Positive Sample and Its Molecular Epidemiology in Henan Province, China. Transbound. Emerg. Dis. 2023, 2023, 9943040. [Google Scholar] [CrossRef]
- Zhao, D.; Yang, B.; Yuan, X.; Shen, C.; Zhang, D.; Shi, X.; Zhang, T.; Cui, H.; Yang, J.; Chen, X. Advanced research in porcine reproductive and respiratory syndrome virus co-infection with other pathogens in swine. Front. Vet. Sci. 2021, 8, 699561. [Google Scholar] [CrossRef]
- Ouyang, T.; Zhang, X.; Liu, X.; Ren, L. Co-infection of swine with porcine circovirus type 2 and other swine viruses. Viruses 2019, 11, 185. [Google Scholar] [CrossRef]
- Van Dung, N.; Anh, P.H.; Van Cuong, N.; Hoa, N.T.; Carrique-Mas, J.; Hien, V.B.; Campbell, J.; Baker, S.; Farrar, J.; Woolhouse, M.E. Prevalence, genetic diversity and recombination of species G enteroviruses infecting pigs in Vietnam. J. Gen. Virol. 2014, 95, 549–556. [Google Scholar] [CrossRef]
- Holmblat, B.; Jégouic, S.; Muslin, C.; Blondel, B.; Joffret, M.L.; Delpeyroux, F. Nonhomologous recombination between defective poliovirus and coxsackievirus genomes suggests a new model of genetic plasticity for picornaviruses. mBio 2014, 5, e01119-14. [Google Scholar] [CrossRef]
- Lukashev, A.N. Recombination among picornaviruses. Rev. Med. Virol. 2010, 20, 327–337. [Google Scholar] [CrossRef]
- Li, Z.H.; Li, Z.R.; Zhu, P.; Zhang, Z.X.; Song, J.L. First identification and pathogenicity evaluation of an EV-G17 strain carrying a Torovirus papain-like cysteine protease (PLCP) gene in China. Viruses 2023, 15, 1747. [Google Scholar] [CrossRef] [PubMed]
- Nikolaidis, M.; Mimouli, K.; Kyriakopoulou, Z.; Tsimpidis, M.; Tsakogiannis, D.; Markoulatos, P.; Amoutzias, G.D. Large-scale genomic analysis reveals recurrent patterns of intertypic recombination in human enteroviruses. Virology 2019, 526, 72–80. [Google Scholar] [CrossRef]
- Yang, X.; Zhou, Y.S.; Wang, H.N.; Zhang, Y.; Wei, K.; Wang, T. Isolation, identification and complete genome sequence analysis of a strain of foot-and-mouth disease virus serotype Asia1 from pigs in southwest of China. Virol. J. 2011, 8, 175. [Google Scholar] [CrossRef] [PubMed]
- Sunaga, F.; Masuda, T.; Ito, M.; Akagami, M.; Naoi, Y.; Sano, K.; Katayama, Y.; Omatsu, T.; Oba, M.; Sakaguchi, S.; et al. Complete genomic analysis and molecular characterization of Japanese porcine sapeloviruses. Virus Genes 2019, 55, 198–208. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.; Saif, L.J.; Wang, Q. Porcine epidemic diarrhea virus (PEDV): An update on etiology, transmission, pathogenesis, and prevention and control. Virus Res. 2020, 286, 198045. [Google Scholar] [CrossRef]
- Morimoto, T.; Watanabe, M. Serological identification of porcine enteroviruses isolated in Japan. Natl. Inst. Anim. Health Q. 1964, 4, 177–182. [Google Scholar]
- L’Ecuyer, C.; Greig, A. Serological and biological studies on porcine enteroviruses isolated in Canada. Can. Vet. J. 1966, 7, 148–154. [Google Scholar]
- Bak, G.-Y.; Kang, M.-I.; Son, K.-Y.; Park, J.-G.; Kim, D.-S.; Seo, J.-Y.; Kim, J.-Y.; Alfajaro, M.M.; Soliman, M.; Baek, Y.-B. Occurrence and molecular characterization of Sapelovirus A in diarrhea and non-diarrhea feces of different age group pigs in one Korean pig farm. J. Vet. Med. Sci. 2016, 78, 1911–1914. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Province | 2019 | 2024 | ||
|---|---|---|---|---|
| Sows | Wild Boars | Sows | Wild Boars | |
| Gangwon | 10 | 12 | 12 | 12 |
| Geonggi | 30 | 30 | 27 | 30 |
| Gyeongnam | 25 | 27 | 27 | 28 |
| Gyeongbuk | 5 | 6 | 6 | 6 |
| Jeonnam | 5 | 6 | 6 | 6 |
| Jeonbuk | 20 | 21 | 21 | 21 |
| Chungnam | 35 | 36 | 36 | 36 |
| Chungbuk | 10 | 12 | 12 | 12 |
| Jeju | 10 | 12 | 12 | 12 |
| Total | 150 | 162 | 159 | 163 |
| Primer Name | Oligonucleotide Sequence (5′-3′) | Primer Position | Product Size (bp) |
|---|---|---|---|
| PSV-1F | GGACTTGGACCTCTGGCAA | 1–19 | 1188 bp |
| PSV-1R | GCTGACCTGAGTTGGGGTTT | 1168–1187 | |
| PSV-2F | GACAGCCTGCAATGGACAAA | 972–991 | 1163 bp |
| PSV-2R | TGCACTGACAGTAACCACCA | 2115–2134 | |
| PSV-3F | TCAGATGGCCACCGGTAAAT | 1850–1869 | 1225 bp |
| PSV-3R | GGCCTAATTGTTCTGCAGGG | 3054–3074 | |
| PSV-4F | CGGAATGGTGCTTCTTATGGT | 2821–2842 | 1261 bp |
| PSV-4R | TAAGCCACTCAGAAGGTCCC | 4062–4082 | |
| PSV-5F | GAATGCACGACTGGGTTCAA | 3728–3748 | 1212 bp |
| PSV-5R | TTGCTTTGCCACAAACCAGT | 4920–4940 | |
| PSV-6F | ACAACCACCTACATTCCACCT | 4660–4681 | 1209 bp |
| PSV-6R | ACCTCCACACTGTCCCATTT | 5849–5869 | |
| PSV-7F | TTTCACTGGACTGGGCATCT | 5496–5516 | 1200 bp |
| PSV-7R | GGTGACAGACTAGCATCCCA | 6676–6696 | |
| PSV-8F | TGAGCTGAGACCCAAAGAGA | 6444–6464 | 907 bp |
| PSV-8R | ATCCAACCAAGACCTACGCA | 7331–7351 |
| Virus | No. of Positive Samples /No. of Test Samples | Positive Rate (%) |
|---|---|---|
| PSV | 12/57 | 21.1 |
| PSV + PRRSV | 3/57 | 5.3 |
| PSV + PCV2 | 1/57 | 1.8 |
| PSV + PEDV | 1/57 | 1.8 |
| PSV + Rota | 3/57 | 5.3 |
| PSV + PRRSV + PCV2 | 4/57 | 7.0 |
| PRRSV | 7/57 | 12.3 |
| PCV2 | 1/57 | 1.8 |
| PEDV | 1/57 | 1.8 |
| Rota | 3/57 | 5.3 |
| PRRSV + PCV2 | 2/57 | 3.5 |
| PCV2 + PED | 1/57 | 1.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.-Y.; Park, C.-K.; Park, G.-N.; Choe, S.; Jang, M.-K.; Lee, Y.-H.; Cho, Y.S.; An, D.-J. Seroprevalence, Genetic Characteristics, and Pathogenicity of Korean Porcine Sapeloviruses. Viruses 2025, 17, 870. https://doi.org/10.3390/v17070870
Kim S-Y, Park C-K, Park G-N, Choe S, Jang M-K, Lee Y-H, Cho YS, An D-J. Seroprevalence, Genetic Characteristics, and Pathogenicity of Korean Porcine Sapeloviruses. Viruses. 2025; 17(7):870. https://doi.org/10.3390/v17070870
Chicago/Turabian StyleKim, Song-Yi, Choi-Kyu Park, Gyu-Nam Park, SeEun Choe, Min-Kyung Jang, Young-Hyeon Lee, Yun Sang Cho, and Dong-Jun An. 2025. "Seroprevalence, Genetic Characteristics, and Pathogenicity of Korean Porcine Sapeloviruses" Viruses 17, no. 7: 870. https://doi.org/10.3390/v17070870
APA StyleKim, S.-Y., Park, C.-K., Park, G.-N., Choe, S., Jang, M.-K., Lee, Y.-H., Cho, Y. S., & An, D.-J. (2025). Seroprevalence, Genetic Characteristics, and Pathogenicity of Korean Porcine Sapeloviruses. Viruses, 17(7), 870. https://doi.org/10.3390/v17070870