


Assessing the Environmental Drivers of Lassa Fever in West Africa: A Systematic Review

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Literature Search and Eligibility Criteria
2.2. Selection
2.3. Extraction and Data Synthesis
3. Results
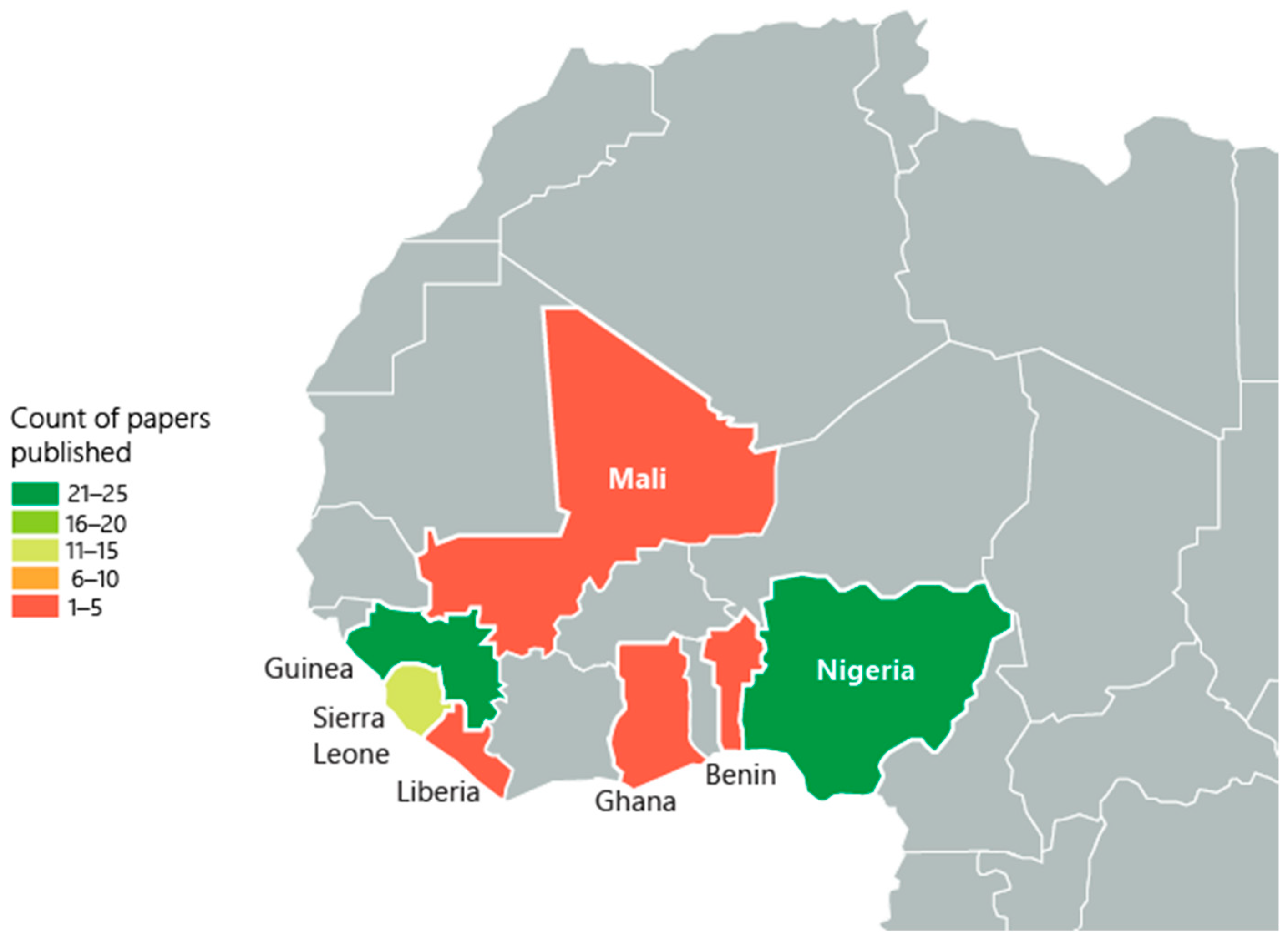
3.1. Search Results and Study Selection
3.2. Characteristics of the Included Studies
3.3. Seasonality
3.3.1. Seasonal Impacts on LASV Prevalence in Host Reservoirs
3.3.2. Seasonal Impacts on Host Reproduction
3.3.3. Rainfall and LF Incidence
3.4. Land Use
3.4.1. Impact of Households on LASV Transmission
3.4.2. Rodent Abundance Inside Houses
3.4.3. Ecological Niche of LASV-Positive Rodents
3.4.4. Habitat Fragmentation
3.4.5. Human LASV Differences in West Africa
3.5. Biodiversity Impacts on Lassa Virus
3.6. Other Hosts of Lassa Virus
3.6.1. Rodent Hosts
3.6.2. Non-Rodent Hosts
4. Discussion
4.1. Lassa Fever in a Changing Climate
4.1.1. Seasonality
4.1.2. Growth of Agriculture
4.2. LF Host Expansion
4.3. Call to Action
4.4. Study Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Monath, T.P.; Newhouse, V.F.; Kemp, G.E.; Setzer, H.W.; Cacciapuoti, A. Lassa Virus Isolation from Mastomys natalensis Rodents during an Epidemic in Sierra Leone. Science 1974, 185, 263–265. [Google Scholar] [PubMed]
- Basinski, A.J.; Fichet-Calvet, E.; Sjodin, A.R.; Varrelman, T.J.; Remien, C.H.; Layman, N.C.; Bird, B.H.; Wolking, D.J.; Monagin, C.; Ghersi, B.M.; et al. Bridging the gap: Using reservoir ecology and human serosurveys to estimate Lassa virus spillover in West Africa. PLoS Comput. Biol. 2021, 17, e1008811. [Google Scholar]
- Agbonlahor, D.E.; Akpede, G.O.; Happi, C.T.; Tomori, O. 52 Years of Lassa Fever Outbreaks in Nigeria, 1969–2020: An Epidemiologic Analysis of the Temporal and Spatial Trends. Am. J. Trop. Med. Hyg. 2021, 105, 974–985. [Google Scholar]
- Fichet-Calvet, E.; Lecompte, E.; Koivogui, L.; Soropogui, B.; Doré, A.; Kourouma, F.; Sylla, O.; Daffis, S.; Koulemou, K.; Ter Meulen, J. Fluctuation of Abundance and Lassa Virus Prevalence in Mastomys natalensis in Guinea, West Africa. Vector-Borne Zoonotic Dis. 2007, 7, 119–128. [Google Scholar] [PubMed]
- Frame, J.D.; Gocke, D.J.; Baldwin, J.M.; Troup, J.M. Lassa Fever, a New Virus Disease of Man from West Africa: I. Clinical Description and Pathological Findings. Am. J. Trop. Med. Hyg. 1970, 19, 670–676. [Google Scholar]
- Garry, R.F. Lassa fever—The road ahead. Nat. Rev. Microbiol. 2023, 21, 87–96. [Google Scholar]
- Yadouleton, A.; Picard, C.; Rieger, T.; Loko, F.; Cadar, D.; Kouthon, E.C.; Job, E.O.; Bankolé, H.; Oestereich, L.; Gbaguidi, F.; et al. Lassa fever in Benin: Description of the 2014 and 2016 epidemics and genetic characterization of a new Lassa virus. Emerg. Microbes Infect. 2020, 9, 1761–1770. [Google Scholar]
- Gibb, R.; Moses, L.M.; Redding, D.W.; Jones, K.E. Understanding the cryptic nature of Lassa fever in West Africa. Pathog. Glob. Health 2017, 111, 276–288. [Google Scholar] [PubMed]
- Mylne, A.Q.N.; Pigott, D.M.; Longbottom, J.; Shearer, F.; Duda, K.A.; Messina, J.P.; Weiss, D.J.; Moyes, C.L.; Golding, N.; Hay, S.I. Mapping the zoonotic niche of Lassa fever in Africa. Trans. R. Soc. Trop. Med. Hyg. 2015, 109, 483–492. [Google Scholar]
- Klitting, R.; Kafetzopoulou, L.E.; Thiery, W.; Dudas, G.; Gryseels, S.; Kotamarthi, A.; Vrancken, B.; Gangavarapu, K.; Momoh, M.; Sandi, J.D.; et al. Predicting the evolution of the Lassa virus endemic area and population at risk over the next decades. Nat. Commun. 2022, 13, 5596. [Google Scholar]
- Redding, D.W.; Gibb, R.; Dan-Nwafor, C.C.; Ilori, E.A.; Yashe, R.U.; Oladele, S.H.; Amedu, M.O.; Iniobong, A.; Attfield, L.A.; Donnelly, C.A.; et al. Geographical drivers and climate-linked dynamics of Lassa fever in Nigeria. Nat. Commun. 2021, 12, 5759. [Google Scholar] [CrossRef]
- Nimo-Paintsil, S.C.; Fichet-Calvet, E.; Borremans, B.; Letizia, A.G.; Mohareb, E.; Bonney, J.H.K.; Obiri-Danso, K.; Ampofo, W.K.; Schoepp, R.J.; Kronmann, K.C. Rodent-borne infections in rural Ghanaian farming communities. PLoS ONE 2019, 14, e0215224. [Google Scholar] [CrossRef] [PubMed]
- Leach, M.; Bett, B.; Said, M.; Bukachi, S.; Sang, R.; Anderson, N.; Machila, N.; Kuleszo, J.; Schaten, K.; Dzingirai, V.; et al. Local disease–ecosystem–livelihood dynamics: Reflections from comparative case studies in Africa. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160163. [Google Scholar]
- Olayemi, A.; Cadar, D.; Magassouba, N.; Obadare, A.; Kourouma, F.; Oyeyiola, A.; Fasogbon, S.; Igbokwe, J.; Rieger, T.; Bockholt, S.; et al. New Hosts of The Lassa Virus. Sci. Rep. 2016, 6, 25280. [Google Scholar]
- Centers for Disease Control and Prevention. Bioterrorism Agents/Diseases [Internet]. CDC|Bioterrorism Agents/Diseases (by Category)|Emergency Preparedness & Response. 2018. Available online: https://emergency.cdc.gov/agent/agentlist-category.asp (accessed on 16 August 2024).
- World Health Organization. Prioritizing Diseases for Research and Development in Emergency Contexts [Internet]. 2019. Available online: https://www.who.int/activities/prioritizing-diseases-for-research-and-development-in-emergency-contexts (accessed on 24 August 2024).
- Fischer, W.A.; Wohl, D.A. Moving Lassa Fever Research and Care into the 21st Century. J. Infect. Dis. 2017, 215, 1779–1781. [Google Scholar] [PubMed]
- McKendrick, J.Q.; Tennant, W.S.D.; Tildesley, M.J. Modelling seasonality of Lassa fever incidences and vector dynamics in Nigeria. PLoS Negl. Trop. Dis. 2023, 17, e0011543. [Google Scholar]
- Redding, D.W.; Moses, L.M.; Cunningham, A.A.; Wood, J.; Jones, K.E. Environmental-mechanistic modelling of the impact of global change on human zoonotic disease emergence: A case study of Lassa fever. Methods Ecol. Evol. 2016, 7, 646–655. [Google Scholar]
- García-Peña, G.E.; Rubio, A.V.; Mendoza, H.; Fernández, M.; Milholland, M.T.; Aguirre, A.A.; Suzán, G.; Zambrana-Torrelio, C. Land-use change and rodent-borne diseases: Hazards on the shared socioeconomic pathways. Philos. Trans. R. Soc. B Biol. Sci. 2021, 376, 20200362. [Google Scholar] [CrossRef]
- Akhmetzhanov, A.R.; Asai, Y.; Nishiura, H. Quantifying the seasonal drivers of transmission for Lassa fever in Nigeria. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20180268. [Google Scholar]
- Musa, S.S.; Zhao, S.; Gao, D.; Lin, Q.; Chowell, G.; He, D. Mechanistic modelling of the large-scale Lassa fever epidemics in Nigeria from 2016 to 2019. J. Theor. Biol. 2020, 493, 110209. [Google Scholar]
- Olayemi, A.; Oyeyiola, A.; Obadare, A.; Igbokwe, J.; Adesina, A.S.; Onwe, F.; Ukwaja, K.N.; Ajayi, N.A.; Rieger, T.; Günther, S.; et al. Widespread arenavirus occurrence and seroprevalence in small mammals, Nigeria. Parasites Vectors 2018, 11, 416. [Google Scholar]
- Bangura, U.; Buanie, J.; Lamin, J.; Davis, C.; Bongo, G.N.; Dawson, M.; Ansumana, R.; Sondufu, D.; Thomson, E.C.; Sahr, F.; et al. Lassa Virus Circulation in Small Mammal Populations in Bo District, Sierra Leone. Biology 2021, 10, 28. [Google Scholar] [CrossRef] [PubMed]
- Fichet-Calvet, E.; LeCompte, E.; Koivogui, L.; Daffis, S.; Meulen, J.T. Reproductive Characteristics of Mastomys natalensis and Lassa Virus Prevalence in Guinea, West Africa. Vector-Borne Zoonotic Dis. 2008, 8, 41–48. [Google Scholar]
- Olayemi, A.; Obadare, A.; Oyeyiola, A.; Fasogbon, S.; Igbokwe, J.; Igbahenah, F.; Ortsega, D.; Günther, S.; Verheyen, E.; Fichet-Calvet, E. Small mammal diversity and dynamics within Nigeria, with emphasis on reservoirs of the lassa virus. Syst. Biodivers. 2018, 16, 118–127. [Google Scholar]
- Fichet-Calvet, E.; Lecompte, E.; Veyrunes, F.; Barrière, P.; Nicolas, V.; Koulémou, K. Diversity and dynamics in a community of small mammals in coastal Guinea, West Africa. Belg. J. Zool. 2009, 139, 93–102. [Google Scholar]
- Zhao, S.; Musa, S.S.; Fu, H.; He, D.; Qin, J. Large-scale Lassa fever outbreaks in Nigeria: Quantifying the association between disease reproduction number and local rainfall. Epidemiol. Infect. 2020, 148, e4. [Google Scholar] [PubMed]
- Ifejube, O.J.; Babalola, S.O.; Mukaila, I.O.; Badewa, A.O. A GIS-based approach to risk mapping of lassa fever outbreak in akure south local government area, Nigeria. Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 2022, 46, 147–153. [Google Scholar]
- Fichet-Calvet, E.; Rogers, D.J. Risk Maps of Lassa Fever in West Africa. PLoS Negl. Trop. Dis. 2009, 3, e388. [Google Scholar]
- Min, K.D.; Hwang, J.; Schneider, M.C.; So, Y.; Lee, J.Y.; Cho, S.-I. An exploration of the protective effect of rodent species richness on the geographical expansion of Lassa fever in West Africa. PLoS Negl. Trop. Dis. 2021, 15, e0009108. [Google Scholar]
- Cadmus, S.; Taiwo, O.J.; Akinseye, V.; Cadmus, E.; Famokun, G.; Fagbemi, S.; Ansumana, R.; Omoluabi, A.; Ayinmode, A.; Oluwayelu, D.; et al. Ecological correlates and predictors of Lassa fever incidence in Ondo State, Nigeria 2017–2021: An emerging urban trend. Sci. Rep. 2023, 13, 20855. [Google Scholar]
- Shaffer, J.G.; Grant, D.S.; Schieffelin, J.S.; Boisen, M.L.; Goba, A.; Hartnett, J.N.; Levy, D.C.; Yenni, R.E.; Moses, L.M.; Fullah, M.; et al. Lassa Fever in Post-Conflict Sierra Leone. PLoS Negl. Trop. Dis. 2014, 8, e2748. [Google Scholar]
- Barua, S.; Dénes, A.; Ibrahim, M.A. A seasonal model to assess intervention strategies for preventing periodic recurrence of Lassa fever. Heliyon 2021, 7, e07760. [Google Scholar] [PubMed]
- Nchom, J.I.; Abubakar, A.S.; Arimoro, F.O.; Mohammed, B.Y. The Role of Weather in the Spread of Lassa Fever in Parts of Northern Nigeria. Int. J. Trop. Dis. Health 2021, 42, 33–40. [Google Scholar]
- OYaa, P.N.; Oluseyi, O.B.; Chukwuyem, A.; Muyiwa, O.J. Mapping of Lassa Fever Epidemics in Owo, Ondo State, Nigeria, 2018–2020: A Descriptive and Categorical Analysis of Age, Gender and Seasonal Pattern. J. Public Allied Health Sci. 2021, 3, 22. [Google Scholar]
- Ibrahim, M.A.; Dénes, A. A mathematical model for Lassa fever transmission dynamics in a seasonal environment with a view to the 2017–20 epidemic in Nigeria. Nonlinear Anal. Real. World Appl. 2021, 60, 103310. [Google Scholar]
- Yaro, C.A.; Kogi, E.; Opara, K.N.; Batiha, G.E.-S.; Baty, R.S.; Albrakati, A.; Altalbawy, F.M.A.; Etuh, I.U.; Oni, J.P. Infection pattern, case fatality rate and spread of Lassa virus in Nigeria. BMC Infect. Dis. 2021, 21, 149. [Google Scholar]
- Shehu, N.Y.; Gomerep, S.S.; Isa, S.E.; Iraoyah, K.O.; Mafuka, J.; Bitrus, N.; Dachom, M.; Ogwuche, J.; Onukak, A.; Onyedibe, K.; et al. Lassa Fever 2016 Outbreak in Plateau State, Nigeria—The Changing Epidemiology and Clinical Presentation. Front. Public Health 2018, 6, 232. [Google Scholar]
- Fasipe, O.J.; Osho, P.O.; Osho, E.S.; Adu, B.S.; Akinrotimi, O.J.; Folayan, W.A.; Adebimpe, W.O. The observed seasonal variation pattern and changing epidemiology of lassa viral hemorrhagic fever disease in Ondo State, Nigeria. Med. J. Dr. Patil. Vidyapeeth 2020, 13, 22. [Google Scholar]
- Okoro, O.A.; Bamgboye, E.; Dan-Nwafor, C.; Umeokonkwo, C.; Ilori, E.; Yashe, R.; Balogun, M.; Nguku, P.; Ihekweazu, C. Descriptive epidemiology of Lassa fever in Nigeria, 2012–2017. Pan Afr. Med. J. 2020, 37, 15. [Google Scholar]
- Happi, A.N.; Olumade, T.J.; Ogunsanya, O.A.; Sijuwola, A.E.; Ogunleye, S.C.; Oguzie, J.U.; Nwofoke, C.; Ugwu, C.A.; Okoro, S.J.; Otuh, P.I.; et al. Increased Prevalence of Lassa Fever Virus-Positive Rodents and Diversity of Infected Species Found during Human Lassa Fever Epidemics in Nigeria. Microbiol. Spectr. 2022, 10, e00366-22. [Google Scholar]
- Fichet-Calvet, E.; Becker-Ziaja, B.; Koivogui, L.; Günther, S. Lassa Serology in Natural Populations of Rodents and Horizontal Transmission. Vector-Borne Zoonotic Dis. 2014, 14, 665–674. [Google Scholar] [PubMed]
- Mariën, J.; Lo Iacono, G.; Rieger, T.; Magassouba, N.; Günther, S.; Fichet-Calvet, E. Households as hotspots of Lassa fever? Assessing the spatial distribution of Lassa virus-infected rodents in rural villages of Guinea. Emerg. Microbes Infect. 2020, 9, 1055–1064. [Google Scholar] [PubMed]
- McCormick, J.B.; Webb, P.A.; Krebs, J.W.; Johnson, K.M.; Smith, E.S. A Prospective Study of the Epidemiology and Ecology of Lassa Fever. J. Infect. Dis. 1987, 155, 437–444. [Google Scholar]
- Bonwitt, J.; Saez, A.M.; Lamin, J.; Ansumana, R.; Dawson, M.; Buanie, J.; Lamin, J.; Sondufu, D.; Borchert, M.; Sahr, F.; et al. At Home with Mastomys and Rattus: Human–Rodent Interactions and Potential for Primary Transmission of Lassa Virus in Domestic Spaces. Am. J. Trop. Med. Hyg. 2017, 96, 935. [Google Scholar]
- Fichet-Calvet, E.; Ölschläger, S.; Strecker, T.; Koivogui, L.; Becker-Ziaja, B.; Camara, A.B.; Soropogui, B.; Magassouba, N.; Günther, S. Spatial and temporal evolution of Lassa virus in the natural host population in Upper Guinea. Sci. Rep. 2016, 6, 21977. [Google Scholar]
- Fair, J.; Jentes, E.; Inapogui, A.; Kourouma, K.; Goba, A.; Bah, A.; Tounkara, M.; Coulibaly, M.; Garry, R.F.; Bausch, D.G. Lassa Virus-Infected Rodents in Refugee Camps in Guinea: A Looming Threat to Public Health in a Politically Unstable Region. Vector-Borne Zoonotic Dis. 2007, 7, 167–171. [Google Scholar]
- Lalis, A.; Leblois, R.; Lecompte, E.; Denys, C.; Ter Meulen, J.; Wirth, T. The Impact of Human Conflict on the Genetics of Mastomys natalensis and Lassa Virus in West Africa. PLoS ONE 2012, 7, e37068. [Google Scholar]
- Adesina, A.S.; Oyeyiola, A.; Obadare, A.; Igbokwe, J.; Abejegah, C.; Akhilomen, P.; Bangura, U.; Asogun, D.; Tobin, E.; Ayodeji, O.; et al. Circulation of Lassa virus across the endemic Edo-Ondo axis, Nigeria, with cross-species transmission between multimammate mice. Emerg. Microbes Infect. 2023, 12, 2219350. [Google Scholar]
- Wood, R.; Bangura, U.; Mariën, J.; Douno, M.; Fichet-Calvet, E. Detection of Lassa virus in wild rodent feces: Implications for Lassa fever burden within households in the endemic region of Faranah, Guinea. One Health 2021, 13, 100317. [Google Scholar]
- Ter Meulen, J.; Lukashevich, I.; Sidibe, K.; Inapogui, A.; Marx, M.; Dorlemann, A.; Yansane, M.L.; Koulemou, K.; Chang-Claude, J.; Schmitz, H. Hunting of Peridomestic Rodents and Consumption of Their Meat as Possible Risk Factors for Rodent-to-Human Transmission of Lassa Virus in the Republic of Guinea. Am. J. Trop. Med. Hyg. 1996, 55, 661–666. [Google Scholar]
- Demby, A.H.; Inapogui, A.; Kargbo, K.; Koninga, J.; Kourouma, K.; Kanu, J.; Coulibaly, M.; Wagoner, K.D.; Ksiazek, T.G.; Peters, C.; et al. Lassa Fever in Guinea: II. Distribution and Prevalence of Lassa Virus Infection in Small Mammals. Vector-Borne Zoonotic Dis. 2001, 1, 283–297. [Google Scholar] [CrossRef] [PubMed]
- Kronmann, K.C.; Nimo-Paintsil, S.; Guirguis, F.; Kronmann, L.C.; Bonney, K.; Obiri-Danso, K.; Ampofo, W.; Fichet-Calvet, E. Two Novel Arenaviruses Detected in Pygmy Mice, Ghana. Emerg. Infect. Dis. 2013, 19, 1832–1835. [Google Scholar] [CrossRef]
- Johnson, K.M.; Webb, P.A.; Smith, E.; Keenlyside, R.A.; Elliott, L.; McCormick, J.B. Case-Control Study of Mastomys natalensis and Humans in Lassa Virus-Infected Households in Sierra Leone. Am. J. Trop. Med. Hyg. 1983, 32, 829–837. [Google Scholar]
- Gonzalez, J.P.; McCormick, J.B.; Saluzzo, J.F.; Herve, J.P.; Georges, A.J.; Johnson, K.M. An arenavirus isolated from wild-caught rodents (Pramys species) in the Central African Republic. Intervirology 1983, 19, 105–112. [Google Scholar] [CrossRef]
- Mariën, J.; Kourouma, F.; Magassouba, N.; Leirs, H.; Fichet-Calvet, E. Movement Patterns of Small Rodents in Lassa Fever-Endemic Villages in Guinea. EcoHealth 2018, 15, 348–359. [Google Scholar] [CrossRef] [PubMed]
- Clark, J.; Yakob, L.; Douno, M.; Lamine, J.; Magassouba, N.; Fichet-Calvet, E.; Mari-Saez, A. Domestic risk factors for increased rodent abundance in a Lassa fever endemic region of rural Upper Guinea. Sci. Rep. 2021, 11, 20698. [Google Scholar] [CrossRef] [PubMed]
- Layman, N.C.; Basinski, A.J.; Zhang, B.; Eskew, E.A.; Bird, B.H.; Ghersi, B.M.; Bangura, J.; Fichet-Calvet, E.; Remien, C.H.; Vandi, M.; et al. Predicting the fine-scale spatial distribution of zoonotic reservoirs using computer vision. Ecol. Lett. 2023, 26, 1974–1986. [Google Scholar] [CrossRef]
- Fichet-Calvet, E.; Koulemou, K.; Koivogui, L.; Soropogui, B.; Sylla, O.; Lecompte, E.; Daffis, S.; Bernard, A.; Kouassi, K.; Akoua-Koffi, C.; et al. Spatial distribution of commensal rodents in regions with high and low Lassa fever prevalence in Guinea. Belg. J. Zool. 2005, 135, 63–67. [Google Scholar]
- Longet, S.; Leggio, C.; Bore, J.A.; Key, S.; Tipton, T.; Hall, Y.; Koundouno, F.R.; Bower, H.; Bhattacharyya, T.; Magassouba, N.; et al. Influence of Landscape Patterns on Exposure to Lassa Fever Virus, Guinea. Emerg. Infect. Dis. 2023, 29, 304–313. [Google Scholar] [CrossRef]
- Olugasa, B.O.; Dogba, J.B.; Ogunro, B.; Odigie, E.A.; Nykoi, J.; Ojo, J.F.; Taiwo, O.; Kamara, A.; Mulbah, C.K.; Fasunla, A.J. The rubber plantation environment and Lassa fever epidemics in Liberia, 2008–2012: A spatial regression. Spat. Spatio-Temporal Epidemiol. 2014, 11, 163–174. [Google Scholar] [CrossRef]
- Amifofum, O.S.; Fadugba, D.; Awosanya, E.; Icomiare, A.; Evbuomwan, K.; Balogun, M.S.; Nguku, P.; Ajisegiri, W.S.; Eugene, O.; Olugasa, B. Spatial analysis of confirmed Lassa fever cases in Edo State, Nigeria, 2008–2014. PAMJ—One Health 2021, 5, 11. [Google Scholar] [CrossRef]
- Kernéis, S.; Koivogui, L.; Magassouba, N.; Koulemou, K.; Lewis, R.; Aplogan, A.; Grais, R.F.; Guerin, P.J.; Fichet-Calvet, E. Prevalence and Risk Factors of Lassa Seropositivity in Inhabitants of the Forest Region of Guinea: A Cross-Sectional Study. PLoS Negl. Trop. Dis. 2009, 3, e548. [Google Scholar] [CrossRef] [PubMed]
- Safronetz, D.; Sogoba, N.; Lopez, J.E.; Maiga, O.; Dahlstrom, E.; Zivcec, M.; Feldmann, F.; Haddock, E.; Fischer, R.; Anderson, J.; et al. Geographic Distribution and Genetic Characterization of Lassa Virus in Sub-Saharan Mali. PLoS Negl. Trop. Dis. 2013, 7, e2582. [Google Scholar]
- Saez, A.M.; Haidara, M.C.; Camara, A.; Kourouma, F.; Sage, M.; Magassouba, N.; Fichet-Calvet, E. Rodent control to fight Lassa fever: Evaluation and lessons learned from a 4-year study in Upper Guinea. PLoS Negl. Trop. Dis. 2018, 12, e0006829. [Google Scholar]
- Leski, T.A.; Stockelman, M.G.; Moses, L.M.; Park, M.; Stenger, D.A.; Ansumana, R.; Bausch, D.G.; Lin, B. Sequence Variability and Geographic Distribution of Lassa Virus, Sierra Leone. Emerg. Infect. Dis. 2015, 21, 609–618. [Google Scholar]
- Safronetz, D.; Lopez, J.E.; Sogoba, N.; Traore’, S.F.; Raffel, S.J.; Fischer, E.R.; Ebihara, H.; Branco, L.; Garry, R.F.; Schwan, T.G.; et al. Detection of Lassa Virus, Mali. Emerg. Infect. Dis. 2010, 16, 1123–1126. [Google Scholar] [CrossRef]
- Lecompte, E.; Fichet-Calvet, E.; Daffis, S.; Koulemou, K.; Sylla, O.; Kourouma, F.; Dore, A.; Soropogui, B.; Aniskin, V.; Bernard, A.K.; et al. Mastomys natalensis and Lassa Fever, West Africa. Emerg. Infect. Dis. 2006, 12, 1971–1974. [Google Scholar] [PubMed]
- Simons, D.; Attfield, L.A.; Jones, K.E.; Watson-Jones, D.; Kock, R. Rodent trapping studies as an overlooked information source for understanding endemic and novel zoonotic spillover. PLoS Negl. Trop. Dis. 2023, 17, e0010772. [Google Scholar]
- Lalis, A.; Evin, A.; Janier, M.; Koivogui, L.; Denys, C. Host evolution in Mastomys natalensis (Rodentia: Muridae): An integrative approach using geometric morphometrics and genetics. Integr. Zool. 2015, 10, 505–514. [Google Scholar] [CrossRef]
- Demartini, J.C.; Green, D.E.; Monath, T.P. Lassa virus infection in Mastomys natalensis in Sierra Leone. Gross and microscopic findings in infected and uninfected animals. Bull. World Health Organ. 1975, 52, 651–663. [Google Scholar]
- Bangura, U.; Davis, C.; Lamin, J.; Bangura, J.; Soropogui, B.; Davison, A.J.; Nichols, J.; Vucak, M.; Dawson, M.; Ansumana, R.; et al. Spatio-temporal spread of Lassa virus and a new rodent host in the Mano River Union area, West Africa. Emerg. Microbes Infect. 2024, 13, 2290834. [Google Scholar] [CrossRef]
- Olayemi, A.; Adesina, A.S.; Strecker, T.; Magassouba, N.; Fichet-Calvet, E. Determining Ancestry between Rodent- and Human-Derived Virus Sequences in Endemic Foci: Towards a More Integral Molecular Epidemiology of Lassa Fever within West Africa. Biology 2020, 9, 26. [Google Scholar] [CrossRef]
- Mariën, J.; Borremans, B.; Gryseels, S.; Soropogui, B.; De Bruyn, L.; Bongo, G.N.; Becker-Ziaja, B.; de Bellocq, J.G.; Günther, S.; Magassouba, N.; et al. No measurable adverse effects of Lassa, Morogoro and Gairo arenaviruses on their rodent reservoir host in natural conditions. Parasit. Vectors 2017, 10, 210. [Google Scholar] [PubMed]
- Agbonlahor, D.; Erah, A.; Agba, I.; Oviasogie, F.; Ehiaghe, A.; Wankasi, M.; Eremwanarue, O.; Ehiaghe, I.; Ogbu, E.; Iyen, R.; et al. Prevalence of Lassa virus among rodents trapped in three South-South States of Nigeria. J. Vector Borne Dis. 2017, 54, 146. [Google Scholar] [CrossRef] [PubMed]
- Wulff, H.; Fabiyi, A.; Monath, T.P. Recent isolations of Lassa virus from Nigerian rodents. Bull. World Health Organ. 1975, 52, 609–613. [Google Scholar]
- Yadouleton, A.; Agolinou, A.; Kourouma, F.; Saizonou, R.; Pahlmann, M.; Bedié, S.K.; Bankolé, H.; Becker-Ziaja, B.; Gbaguidi, F.; Thielebein, A.; et al. Lassa Virus in Pygmy Mice, Benin, 2016–2017. Emerg. Infect. Dis. 2019, 25, 1977–1979. [Google Scholar] [CrossRef]
- Mariën, J.; Borremans, B.; Kourouma, F.; Baforday, J.; Rieger, T.; Günther, S.; Magassouba, N.; Leirs, H.; Fichet-Calvet, E. Evaluation of rodent control to fight Lassa fever based on field data and mathematical modelling. Emerg. Microbes Infect. 2019, 8, 640–649. [Google Scholar] [CrossRef]
- Nuismer, S.L.; Remien, C.H.; Basinski, A.J.; Varrelman, T.; Layman, N.; Rosenke, K.; Bird, B.; Jarvis, M.; Barry, P.; Hanley, P.W.; et al. Bayesian estimation of Lassa virus epidemiological parameters: Implications for spillover prevention using wildlife vaccination. PLoS Negl. Trop. Dis. 2020, 14, e0007920. [Google Scholar] [CrossRef]
- Happi, A.N.; Ogunsanya, O.A.; Ayinla, A.O.; Sijuwola, A.E.; Saibu, F.M.; Akano, K.; Nwofoke, C.; Elias, O.T.; Achonduh-Atijegbe, O.; Daodu, R.O.; et al. Lassa virus in novel hosts: Insights into the epidemiology of lassa virus infections in southern Nigeria. Emerg. Microbes Infect. 2024, 13, 2294859. [Google Scholar] [CrossRef]
- Ogunro, B.N.; Olugasa, B.O.; Kayode, A.; Ishola, O.O.; Kolawole, O.N.; Odigie, E.A.; Happi, C. Detection of Antibody and Antigen for Lassa Virus Nucleoprotein in Monkeys from Southern Nigeria. J. Epidemiol. Glob. Health 2019, 9, 125–127. [Google Scholar] [CrossRef]
- Sylla, M.B.; Nikiema, P.M.; Gibba, P.; Kebe, I.; Klutse, N.A.B. Climate Change over West Africa: Recent Trends and Future Projections. In Adaptation to Climate Change and Variability in Rural West Africa [Internet]; Yaro, J.A., Hesselberg, J., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 25–40. Available online: http://link.springer.com/10.1007/978-3-319-31499-0_3 (accessed on 6 August 2024).
- Herrmann, S.M.; Brandt, M.; Rasmussen, K.; Fensholt, R. Accelerating land cover change in West Africa over four decades as population pressure increased. Commun. Earth Environ. 2020, 1, 53. [Google Scholar]
- Brandt, M.; Rasmussen, K.; Peñuelas, J.; Tian, F.; Schurgers, G.; Verger, A.; Mertz, O.; Palmer, J.R.B.; Fensholt, R. Human population growth offsets climate-driven increase in woody vegetation in sub-Saharan Africa. Nat. Ecol. Evol. 2017, 1, 0081. [Google Scholar]
- Granjon, L.; Artige, E.; Bâ, K.; Brouat, C.; Dalecky, A.; Diagne, C.; Diallo, M.; Fossati-Gaschignard, O.; Gauthier, P.; Kane, M.; et al. Sharing space between native and invasive small mammals: Study of commensal communities in Senegal. Ecol. Evol. 2023, 13, e10539. [Google Scholar]
- Fichet-Calvet, E.; Audenaert, L.; Barrière, P.; Verheyen, E. Diversity, dynamics and reproduction in a community of small mammals in Upper Guinea, with emphasis on pygmy mice ecology. Afr. J. Ecol. 2010, 48, 600–614. [Google Scholar]
- Grange, Z.L.; Goldstein, T.; Johnson, C.K.; Anthony, S.; Gilardi, K.; Daszak, P.; Olival, K.J.; O’rourke, T.; Murray, S.; Olson, S.H.; et al. Ranking the risk of animal-to-human spillover for newly discovered viruses. Proc. Natl. Acad. Sci. USA 2021, 118, e2002324118. [Google Scholar]
- Kenmoe, S.; Tchatchouang, S.; Ebogo-Belobo, J.T.; Ka’E, A.C.; Mahamat, G.; Simo, R.E.G.; Bowo-Ngandji, A.; Emoh, C.P.D.; Che, E.; Ngongang, D.T.; et al. Systematic review and meta-analysis of the epidemiology of Lassa virus in humans, rodents and other mammals in sub-Saharan Africa. PLoS Negl. Trop. Dis. 2020, 14, e0008589. [Google Scholar]
- USAID Environment—Biodiversity and Climate Change [Internet]. United States Agency for International Development. Available online: https://www.usaid.gov/west-africa-regional/environment-biodiversty-and-climate-change (accessed on 14 August 2024).
- Keesing, F.; Belden, L.K.; Daszak, P.; Dobson, A.; Harvell, C.D.; Holt, R.D.; Hudson, P.; Jolles, A.E.; Jones, K.E.; Mitchell, C.E.; et al. Impacts of biodiversity on the emergence and transmission of infectious diseases. Nature 2010, 468, 647–652. [Google Scholar]
- Khalil, H.; Ecke, F.; Evander, M.; Magnusson, M.; Hörnfeldt, B. Declining ecosystem health and the dilution effect. Sci. Rep. 2016, 6, 31314. [Google Scholar] [CrossRef]
- Ostfeld, R.S.; Keesing, F. Effects of Host Diversity on Infectious Disease. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 157–182. [Google Scholar]
- Eskew, E.A.; Bird, B.H.; Ghersi, B.M.; Bangura, J.; Basinski, A.J.; Amara, E.; Bah, M.A.; Kanu, M.C.; Kanu, O.T.; Lavalie, E.G.; et al. Reservoir displacement by an invasive rodent reduces Lassa virus zoonotic spillover risk. Nat. Commun. 2024, 15, 3589. [Google Scholar]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Davis, N.A.; Kenyon, M.A.; Ghersi, B.M.; Sparks, J.L.D.; Gass, J.D., Jr. Assessing the Environmental Drivers of Lassa Fever in West Africa: A Systematic Review. Viruses 2025, 17, 504. https://doi.org/10.3390/v17040504
Davis NA, Kenyon MA, Ghersi BM, Sparks JLD, Gass JD Jr. Assessing the Environmental Drivers of Lassa Fever in West Africa: A Systematic Review. Viruses. 2025; 17(4):504. https://doi.org/10.3390/v17040504
Chicago/Turabian StyleDavis, Natalie A., Madeline A. Kenyon, Bruno M. Ghersi, Jessica L. Decker Sparks, and Jonathon D. Gass, Jr. 2025. "Assessing the Environmental Drivers of Lassa Fever in West Africa: A Systematic Review" Viruses 17, no. 4: 504. https://doi.org/10.3390/v17040504
APA StyleDavis, N. A., Kenyon, M. A., Ghersi, B. M., Sparks, J. L. D., & Gass, J. D., Jr. (2025). Assessing the Environmental Drivers of Lassa Fever in West Africa: A Systematic Review. Viruses, 17(4), 504. https://doi.org/10.3390/v17040504