
Neonatal Development in Prenatally Zika Virus-Exposed Infant Macaques with Dengue Immunity

 , , ,
, , ,  , , , add
Show full author list
, , , add
Show full author list
Abstract
:1. Introduction
2. Materials and Methods
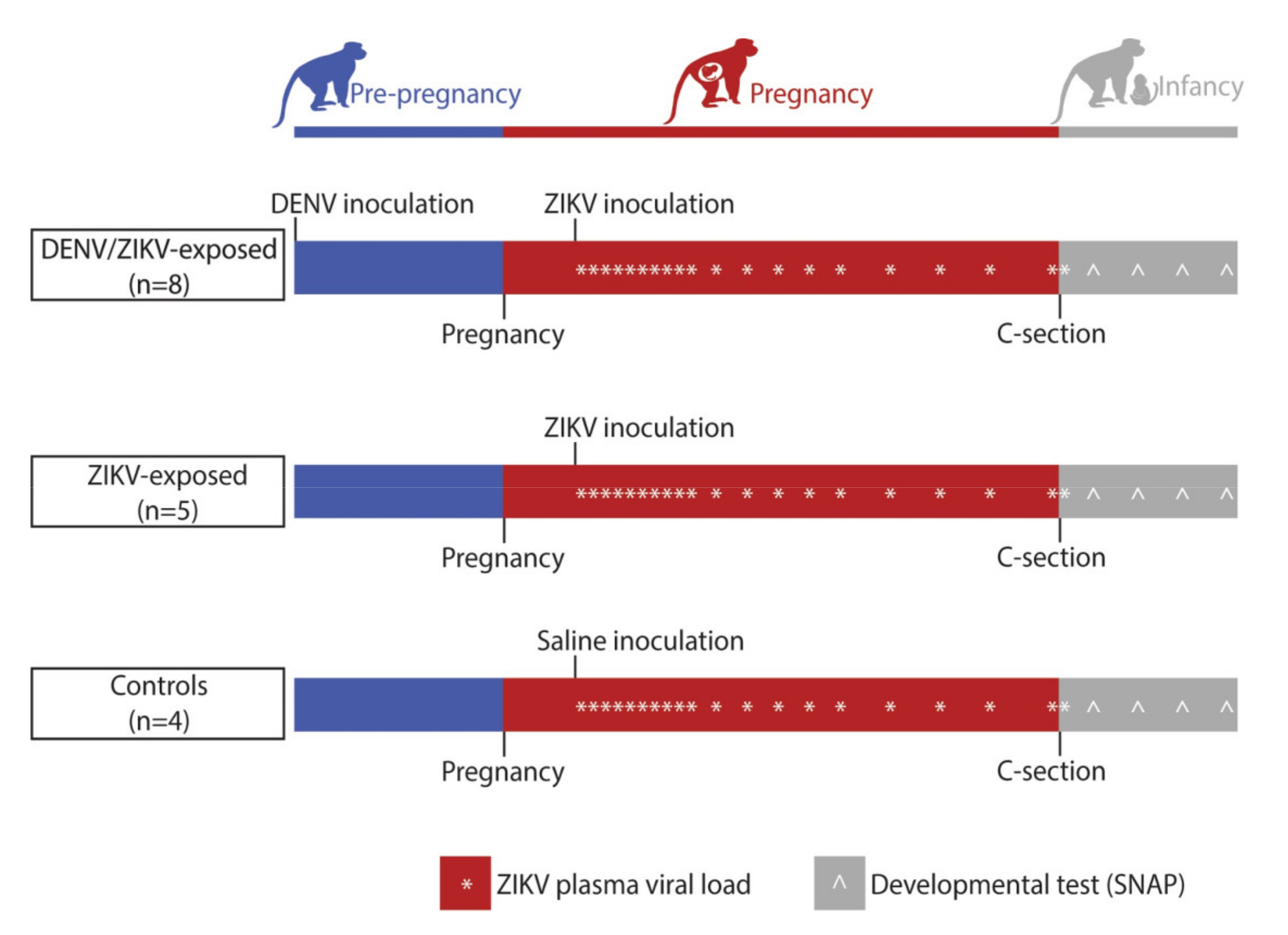
2.1. Study Design
2.2. Ethics Statement
2.3. Care and Use of Macaques
2.4. Inoculation and Monitoring
2.5. Pregnancy Monitoring and Fetal Measurements
2.6. Cesarean Delivery
2.7. Placental Pathology
2.8. vRNA Isolation from Plasma and Tissues
2.9. Plaque Reduction Neutralization Test (PRNT)
2.10. Infant Care
2.11. Neurobehavioral Assessments
2.12. Statistical Analyses
2.12.1. Neonatal Development
2.12.2. Pregnancy and Infant Individual Characteristics
2.12.3. Predictors of Neonatal Neurobehavior
3. Results
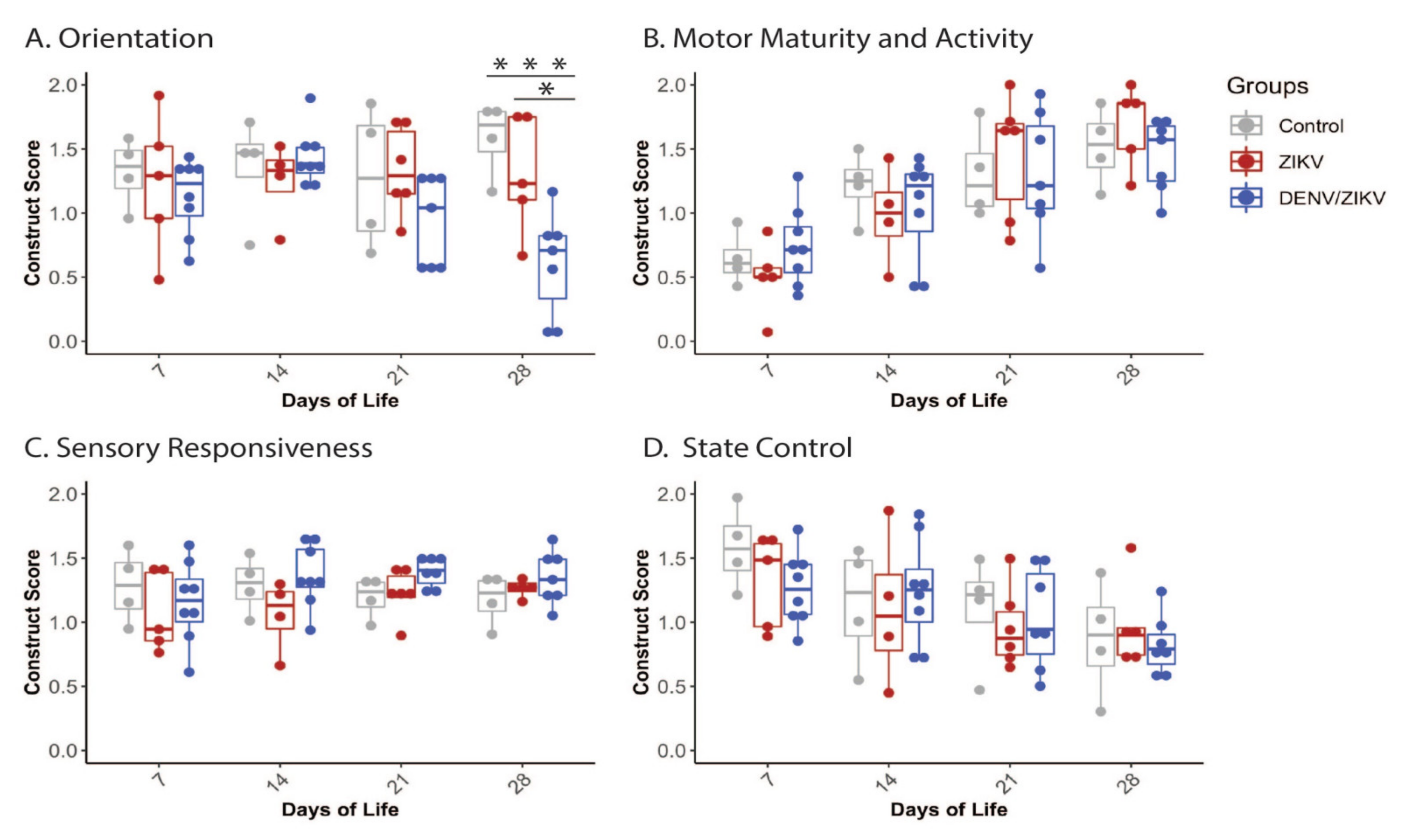
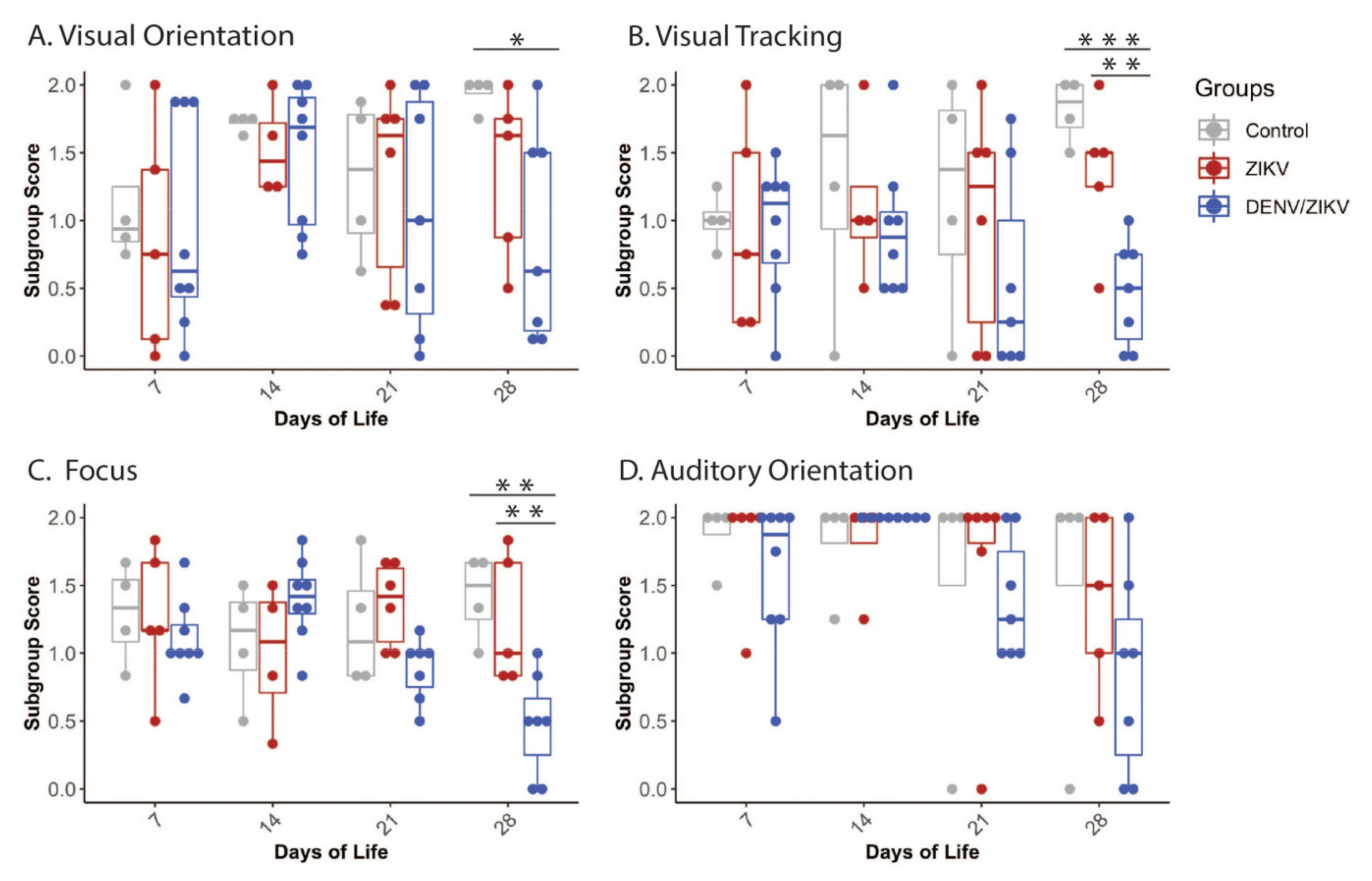
3.1. Neonatal Development
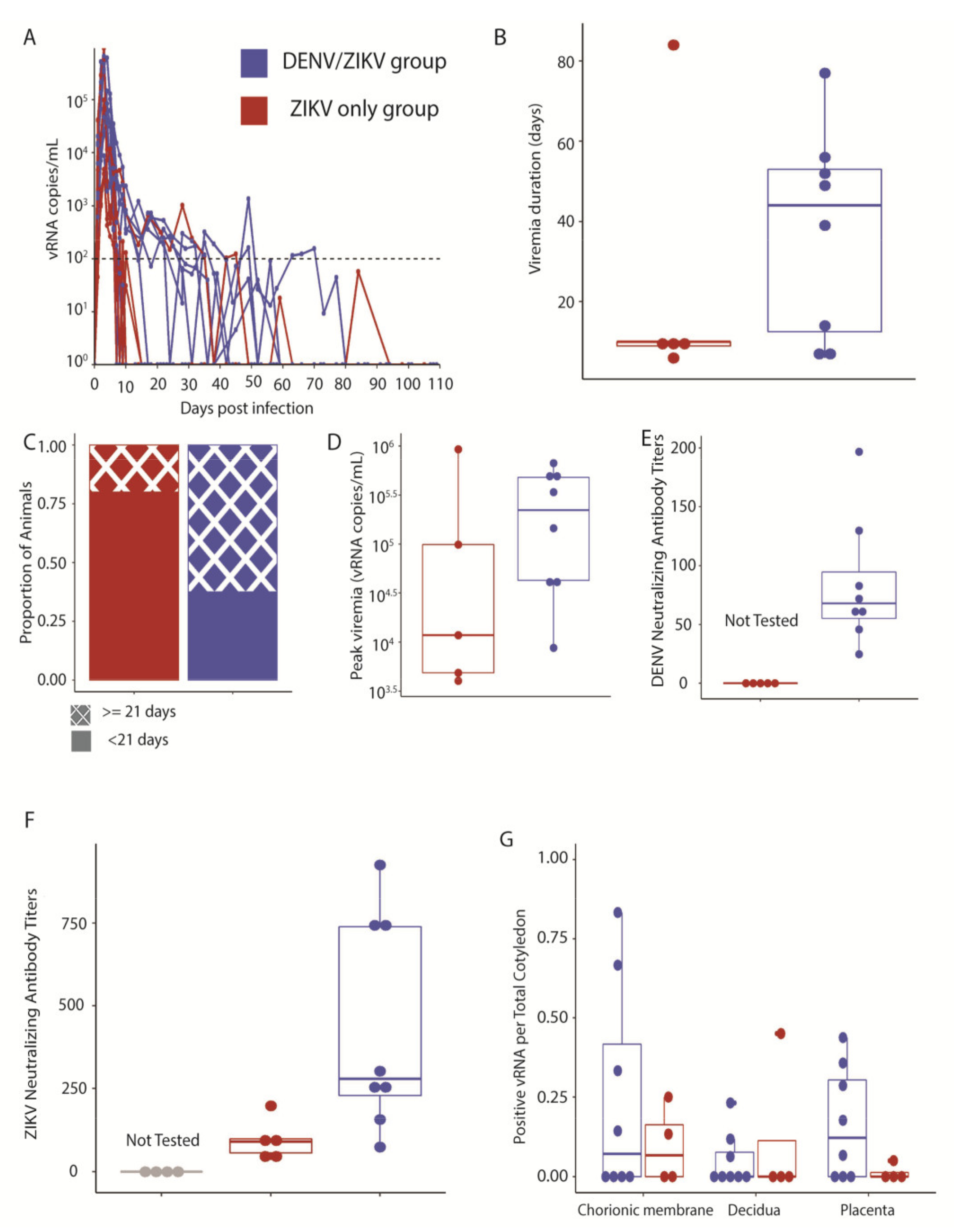
3.2. Maternal ZIKV Infection Characteristics
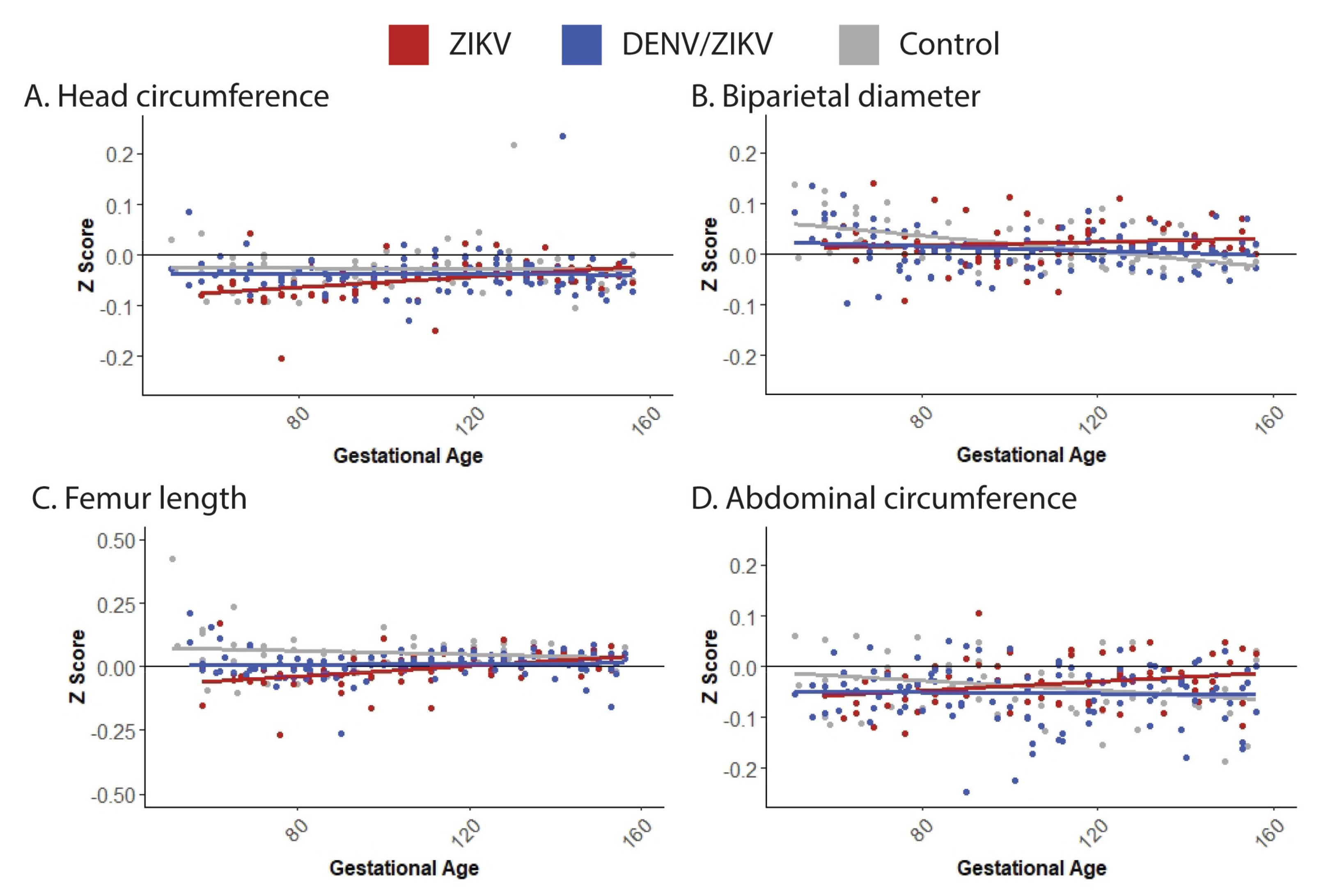
3.3. Fetal Growth, Placental Pathology and Infant Health
3.4. Predictors of Neonatal Neurodevelopment
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mulkey, S.B.; Arroyave-Wessel, M.; Peyton, C.; Bulas, D.I.; Fourzali, Y.; Jiang, J.; Russo, S.; McCarter, R.; Msall, M.E.; Du Plessis, A.J.; et al. Neurodevelopmental Abnormalities in Children within Utero Zika Virus Exposure Without Congenital Zika Syndrome. JAMA Pediatr. 2020, 174, 269–276. [Google Scholar] [CrossRef]
- Nielsen-Saines, K.; Brasil, P.; Kerin, T.; Vasconcelos, Z.; Gabaglia, C.R.; Damasceno, L.; Pone, M.V.D.S.; De Carvalho, L.M.A.; Pone, S.M.; Zin, A.A.; et al. Delayed childhood neurodevelopment and neurosensory alterations in the second year of life in a prospective cohort of ZIKV-exposed children. Nat. Med. 2019, 25, 1213–1217. [Google Scholar] [CrossRef] [PubMed]
- Peçanha, P.M.; Junior, S.C.G.; Pone, S.M.; Pone, M.V.D.S.; Vasconcelos, Z.; Zin, A.; Vilibor, R.H.H.; Costa, R.P.; Meio, M.D.B.B.; Nielsen-Saines, K.; et al. Neurodevelopment of children exposed intra-uterus by Zika virus: A case series. PLoS ONE 2020, 15, e0229434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vianna, R.A.D.O.; Lovero, K.L.; de Oliveira, S.A.; Fernandes, A.; dos Santos, T.C.S.; Lima, L.C.S.D.S.; Carvalho, F.R.; Quintans, M.D.S.; Bueno, A.C.; Torbey, A.F.M.; et al. Children Born to Mothers with Rash During Zika Virus Epidemic in Brazil: First 18 Months of Life. J. Trop. Pediatr. 2019, 65, 592–602. [Google Scholar] [CrossRef] [PubMed]
- Rice, M.E.; Galang, R.R.; Roth, N.M.; Ellington, S.R.; Moore, C.A.; Valencia-Prado, M.; Ellis, E.M.; Tufa, A.J.; Taulung, L.A.; Alfred, J.M.; et al. Vital Signs:Zika-Associated Birth Defects and Neurodevelopmental Abnormalities Possibly Associated with Congenital Zika Virus Infection—U.S. Territories and Freely Associated States, 2018. MMWR Morb. Mortal. Wkly. Rep. 2018, 67, 858–867. [Google Scholar] [CrossRef] [Green Version]
- Bardina, S.V.; Bunduc, P.; Tripathi, S.; Duehr, J.; Frere, J.J.; Brown, J.; Nachbagauer, R.; Foster, G.A.; Krysztof, D.; Tortorella, D.; et al. Enhancement of Zika virus pathogenesis by preexisting antiflavivirus immunity. Science 2017, 356, 175–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCracken, M.; Gromowski, G.D.; Friberg, H.L.; Lin, X.; Abbink, P.; De La Barrera, R.; Eckles, K.H.; Garver, L.S.; Boyd, M.; Jetton, D.; et al. Impact of prior flavivirus immunity on Zika virus infection in rhesus macaques. PLoS Pathog. 2017, 13, e1006487. [Google Scholar] [CrossRef] [PubMed]
- Pantoja, P.; Pérez-Guzmán, E.X.; Rodríguez, I.V.; White, L.J.; González, O.; Serrano, C.; Giavedoni, L.; Hodara, V.; Cruz, L.; Arana, T.; et al. Zika virus pathogenesis in rhesus macaques is unaffected by pre-existing immunity to dengue virus. Nat. Commun. 2017, 8, 15674. [Google Scholar] [CrossRef]
- Petzold, S.; Agbaria, N.; Deckert, A.; Dambach, P.; Winkler, V.; Drexler, J.F.; Horstick, O.; Jaenisch, T. Congenital abnormalities associated with Zika virus infection–Dengue as potential co-factor? A systematic review. PLoS Negl. Trop. Dis. 2021, 15, e0008984. [Google Scholar] [CrossRef]
- Halai, U.-A.; Nielsen-Saines, K.; Moreira, M.L.; De Sequeira, P.C.; Junior, J.P.P.; Zin, A.D.A.; Cherry, J.; Gabaglia, C.R.; Gaw, S.; Adachi, K.; et al. Maternal Zika Virus Disease Severity, Virus Load, Prior Dengue Antibodies, and Their Relationship to Birth Outcomes. Clin. Infect. Dis. 2017, 65, 877–883. [Google Scholar] [CrossRef] [Green Version]
- Brasil, P.; Pereira, J.P.; Moreira, M.E.; Nogueira, R.M.R.; Damasceno, L.; Wakimoto, M.; Rabello, R.S.; Valderramos, S.G.; Halai, U.-A.; Salles, T.S.; et al. Zika Virus Infection in Pregnant Women in Rio de Janeiro. N. Engl. J. Med. 2016, 375, 2321–2334. [Google Scholar] [CrossRef]
- Moreira-Soto, A.; Cabral, R.; Pedroso, C.; Eschbach-Bludau, M.; Rockstroh, A.; Vargas, L.A.; Postigo-Hidalgo, I.; Luz, E.; Sampaio, G.S.; Drosten, C.; et al. Exhaustive TORCH Pathogen Diagnostics Corroborate Zika Virus Etiology of Congenital Malformations in Northeastern Brazil. mSphere 2018, 3, e00278-18. [Google Scholar] [CrossRef] [Green Version]
- Pedroso, C.; Fischer, C.; Feldmann, M.; Sarno, M.; Luz, E.; Moreira-Soto, A.; Cabral, R.; Netto, E.M.; Brites, C.; Kümmerer, B.M.; et al. Cross-Protection of Dengue Virus Infection against Congenital Zika Syndrome, Northeastern Brazil. Emerg. Infect. Dis. 2019, 25, 1485–1493. [Google Scholar] [CrossRef] [Green Version]
- Costa, K.C.M.; Brancaglion, G.A.; Almeida, C.A.D.F.; de Amorim, G.E.S.; Veloso, L.L.; Lião, L.D.S.; de Souza, G.A.P.; Pinheiro, B.P.; Ângelo, M.L.; Ruginsk, S.G.; et al. No effect of prior Dengue virus 1 infection in mouse dams on long-term behavioral profiles in offspring infected with Zika virus during gestation. Neurosci. Lett. 2020, 739, 135448. [Google Scholar] [CrossRef]
- Mohr, E.L. Modeling Zika Virus-Associated Birth Defects in Nonhuman Primates. J. Pediatr. Infect. Dis. Soc. 2018, 7, S60–S66. [Google Scholar] [CrossRef] [PubMed]
- Dudley, D.M.; Aliota, M.T.; Mohr, E.L.; Newman, C.M.; Golos, T.G.; Friedrich, T.C.; O’Connor, D.H. Using Macaques to Address Critical Questions in Zika Virus Research. Annu. Rev. Virol. 2019, 6, 481–500. [Google Scholar] [CrossRef] [Green Version]
- Koenig, M.R.; Razo, E.; Mitzey, A.; Newman, C.; Dudley, D.M.; Breitbach, M.E.; Semler, M.R.; Stewart, L.M.; Weiler, A.M.; Rybarczyk, S.; et al. Quantitative definition of neurobehavior, vision, hearing and brain volumes in macaques congenitally exposed to Zika virus. PLoS ONE 2020, 15, e0235877. [Google Scholar] [CrossRef] [PubMed]
- Sackett, G.P. CHAPTER 1—Developmental Disabilities and Primate Models Defined. Primate Models of Children’s Health and Developmental Disabilities; Academic Press: New York, NY, USA, 2008; pp. 1–10. [Google Scholar]
- Crooks, C.M.; Weiler, A.M.; Rybarczyk, S.L.; Bliss, M.I.; Jaeger, A.S.; Murphy, M.E.; Simmons, H.A.; Mejia, A.; Fritsch, M.K.; Hayes, J.M.; et al. Prior dengue immunity enhances Zika virus infection of the maternal-fetal interface in rhesus macaques. bioRxiv 2021. [Google Scholar] [CrossRef]
- Weatherall, D. The Use of Non-Human Primates in Research, The Royal Society. 2006. Available online: https://royalsociety.org/topics-policy/publications/2006/weatherall-report/ (accessed on 20 June 2021).
- Tarantal, A.F. Ultrasound Imaging in Rhesus (Macaca mulatta) and Long-Tailed (Macaca fascicularis) Macaques: Reproductive and Research Applications. Ultrasound Imaging; Elsevier Ltd.: Amsterdam, The Netherlands, 2005. [Google Scholar]
- Nguyen, S.M.; Antony, K.M.; Dudley, D.M.; Kohn, S.; Simmons, H.A.; Wolfe, B.; Salamat, M.S.; Teixeira, L.B.C.; Wiepz, G.J.; Thoong, T.H.; et al. Highly efficient maternal-fetal Zika virus transmission in pregnant rhesus macaques. PLoS Pathog. 2017, 13, e1006378. [Google Scholar] [CrossRef] [Green Version]
- Dudley, D.M.; Aliota, M.; Mohr, E.L.; Weiler, A.M.; Lehrer-Brey, G.; Weisgrau, K.L.; Mohns, M.S.; Breitbach, M.E.; Rasheed, M.N.; Newman, C.M.; et al. A rhesus macaque model of Asian-lineage Zika virus infection. Nat. Commun. 2016, 7, 12204. [Google Scholar] [CrossRef] [PubMed]
- Hansen, S.G.; Piatak, M.; Ventura, A.B.; Hughes, C.M.; Gilbride, R.M.; Ford, J.C.; Oswald, K.; Shoemaker, R.; Li, Y.; Lewis, M.S.; et al. Immune clearance of highly pathogenic SIV infection. Nature 2013, 502, 100–104. [Google Scholar] [CrossRef] [Green Version]
- Lindsey, H.S.; Calisher, C.H.; Mathews, J.H. Serum dilution neutralization test for California group virus identification and serology. J. Clin. Microbiol. 1976, 4, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Goodlin, B.L.; Sackett, G.P. Parturition in Macaca nemestrina. Am. J. Primatol. 1983, 4, 283–307. [Google Scholar] [CrossRef]
- Laughlin, N.K.; Lasky, R.E.; Giles, N.L.; Luck, M.L. Lead Effects on Neurobehavioral Development in the Neonatal Rhesus Monkey (Macaca mulatta). Neurotoxicol. Teratol. 1999, 21, 627–638. [Google Scholar] [CrossRef]
- Schneider, M.L.; Moore, C.F.; Kraemer, G.W.; Roberts, A.D.; DeJesus, O.T. The impact of prenatal stress, fetal alcohol exposure, or both on development: Perspectives from a primate model. Psychoneuroendocrinology 2001, 27, 285–298. [Google Scholar] [CrossRef]
- Schneider, M.L.; Roughton, E.C.; Koehler, A.J.; Lubach, G.R. Growth and Development Following Prenatal Stress Exposure in Primates: An Examination of Ontogenetic Vulnerability. Child. Dev. 1999, 70, 263–274. [Google Scholar] [CrossRef]
- Levin, E.D.; Schneider, M.L.; Ferguson, S.; Schantz, S.L.; Bowman, R.E. Behavioral effects of developmental lead exposure in rhesus monkeys. Dev. Psychobiol. 1988, 21, 371–382. [Google Scholar] [CrossRef]
- Coe, C.L.; Lubach, G.R.; Crispen, H.R.; Shirtcliff, E.A.; Schneider, M.L. Challenges to maternal wellbeing during pregnancy impact temperament, attention, and neuromotor responses in the infant rhesus monkey. Dev. Psychobiol. 2010, 52, 625–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brazelton, T.B. Assessment of the infant at risk. Clin. Obstetr. Gynecol. 1973, 16, 361–375. [Google Scholar] [CrossRef]
- Short, S.J.; Lubach, G.R.; Karasin, A.I.; Olsen, C.W.; Styner, M.; Knickmeyer, R.; Gilmore, J.H.; Coe, C.L. Maternal Influenza Infection During Pregnancy Impacts Postnatal Brain Development in the Rhesus Monkey. Biol. Psychiatry 2010, 67, 965–973. [Google Scholar] [CrossRef] [Green Version]
- Schneider, M.L.; Suomi, S.J. Neurobehavioral assessment in rhesus monkey neonates (Macaca mulatta): Developmental changes, behavioral stability, and early experience. Infant Behav. Dev. 1992, 15, 155–177. [Google Scholar] [CrossRef]
- Schneider, M.L.; Moore, C.F. Effect of prenatal stress on development: A nonhuman primate model. In Minnesota Symposia on Child Psychology; Lawrence Erlbaum: Hillsdale, NJ, USA, 2000. [Google Scholar]
- Schneider, M.L.; Moore, C.F.; Adkins, M.M. The Effects of Prenatal Alcohol Exposure on Behavior: Rodent and Primate Studies. Neuropsychol. Rev. 2011, 21, 186–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, M.L.; Moore, C.F.; Larson, J.A.; Barr, C.S.; DeJesus, O.T.; Roberts, A.D. Timing of moderate level prenatal alcohol exposure influences gene expression of sensory processing behavior in rhesus monkeys. Front. Integr. Neurosci. 2009, 3, 30. [Google Scholar] [CrossRef] [Green Version]
- Walker, C.L.; Merriam, A.A.; Ohuma, E.O.; Dighe, M.K.; Gale, M.; Rajagopal, L.; Papageorghiou, A.T.; Gyamfi-Bannerman, C.; Waldorf, K.M.A. Femur-sparing pattern of abnormal fetal growth in pregnant women from New York City after maternal Zika virus infection. Am. J. Obstet. Gynecol. 2018, 219, 187.e1–187.e20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, M.L.; Moore, C.F.; Kraemer, G.W. Moderate alcohol during pregnancy: Learning and behavior in adolescent rhesus monkeys. Alcohol. Clin. Experim. Res. 2001, 25, 1383–1392. [Google Scholar] [CrossRef]
- Stringer, E.M.; Martinez, E.; Blette, B.; Ruiz, C.E.T.; Boivin, M.; Zepeda, O.; Stringer, J.S.A.; Morales, M.; Ortiz-Pujols, S.; Familiar, I.; et al. Neurodevelopmental Outcomes of Children Following in Utero Exposure to Zika in Nicaragua. Clin. Infect. Dis. 2021, 72, e146–e153. [Google Scholar] [CrossRef] [PubMed]
- Pomar, L.; Lambert, V.; Matheus, S.; Pomar, C.; Hcini, N.; Carles, G.; Rousset, D.; Vouga, M.; Panchaud, A.; Baud, D. Prolonged Maternal Zika Viremia as a Marker of Adverse Perinatal Outcomes. Emerg. Infect. Dis. 2021, 27, 490–498. [Google Scholar] [CrossRef]
- Thomas, S.J.; Jarman, R.; Endy, T.P.; Kalayanarooj, S.; Vaughn, D.W.; Nisalak, A.; Putnak, R.; Anderson, K.B.; Gibbons, R.V.; Libraty, D.H. Dengue Plaque Reduction Neutralization Test (PRNT) in Primary and Secondary Dengue Virus Infections: How Alterations in Assay Conditions Impact Performance. Am. J. Trop. Med. Hyg. 2009, 81, 825–833. [Google Scholar] [CrossRef] [Green Version]
- Katzelnick, L.C.; Gresh, L.; Halloran, M.E.; Mercado, J.C.; Kuan, G.; Gordon, A.; Balmaseda, A.; Harris, E. Antibody-dependent enhancement of severe dengue disease in humans. Science 2017, 358, 929–932. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Dam ID | Inoculation Prior to Pregnancy | Inoculation during Pregnancy | Gestational Day at Inoculation | Dam Age at Inoculation during Pregnancy (Years) | Gestational Day at Infant Delivery | Number of Previous Pregnancies | Pregnancy Data Presented in Previous Publication |
|---|---|---|---|---|---|---|---|---|
| Control | 044-105 | - | PBS | 42 | 11 | 161 | 3 | - |
| 044-106 | - | PBS | 48 | 15 | 160 | 6 | - | |
| 044-107 | - | PBS | 48 | 8 | 163 | 5 | - | |
| 044-108 | - | PBS | 48 | 12 | 159 | 5 | - | |
| ZIKV | 044-101 | DMEM + FBS | ZIKV | 48 | 16 | 160 | 6 | (18) |
| 044-102 | DMEM + FBS | ZIKV | 45 | 17 | 160 | 7 | (18) | |
| 044-103 | DMEM + FBS | ZIKV | 45 | 13 | 159 | 3 | (18) | |
| 044-104 | DMEM + FBS # | ZIKV | 45 | 7 | 159 | 2 | (18) | |
| 044-109 | - | ZIKV | 48 | 16 | 160 | 7 | - | |
| DENV/ZIKV | 042-102 | DENV | ZIKV | 46 | 7 | 160 | 2 | (18) |
| 042-104 | DENV | ZIKV | 46 | 12 | 160 | 5 | (18) | |
| 042-101 | DENV | ZIKV | 48 | 13 | 160 | 5 | (18) | |
| 042-103 | DENV | ZIKV | 45 | 6 | 159 | 1 | (18) | |
| 042-107 | DENV | ZIKV | 48 | 11 | 160 | 3 | (18) | |
| 042-105 | DENV | ZIKV | 49 | 15 | 161 | 6 | (18) | |
| 042-108 | DENV | ZIKV | 45 | 8 | 161 | 3 | (18) |
| Group | Dam | Infarctions/ Total Cotyledons | Chronic Histiocytic Intervillositis | Villous Stromal Calcifications | Vasculopathy | Total Placental Weight (g) |
|---|---|---|---|---|---|---|
| Control | 044-105 | 1/17 | Absent | Present | Absent | 111.08 |
| 044-106 | 1/8 | Absent | Present | Absent | 106.5 | |
| 044-107 | 0/9 | Absent | Present | Present | 144.48 | |
| 044-108 | 5/11 | Absent | Present | Absent | 122.92 | |
| ZIKV | 044-101 | 5/20 | Absent | Present | Absent | 172.59 |
| 044-102 | 6/15 | Absent | Present | Absent | 123.87 | |
| 044-103 | 0/16 | Absent | Absent | Absent | 134.49 | |
| 044-104 | 2/11 | Absent | Absent | Absent | 120.48 | |
| DENV/ZIKV | 042-101 | 3/14 | Absent | Present | Absent | 104.4 |
| 042-102 | 1/13 | Present | Present | Absent | 111.9 | |
| 042-103 | 0/17 | Absent | Present | Absent | 120.06 | |
| 042-104 | 4/15 | Absent | Present | Absent | 95.39 | |
| 042-105 | 4/16 | Absent | Absent | Absent | 119.97 | |
| 042-106 | 8/15 | Absent | Present | Present | 120.14 | |
| 042-107 | 4/14 | Absent | Present | Absent | 139.74 | |
| 042-108 | 5/14 | Absent | Present | Absent | 129.54 |
| Outcomes | Predictors | Univariate Regression Analysis | Multivariate Regression Analysis | ||||
|---|---|---|---|---|---|---|---|
| Correlation Coefficient | p-Value | Standardized Regression Coefficient | p-Value | Variance Inflation Factor | R2 Final Multivariate Model | ||
| Orientation construct | Duration of maternal plasma viremia | 0.10 | 0.7651 | −0.92 | 0.0112 | 3.80 | 0.85 |
| Peak maternal plasma viral load | −0.21 | 0.5193 | |||||
| Fraction of vRNA -positive cotyledons | −0.21 | 0.4479 | 0.72 | 0.0197 | 3.10 | ||
| DENV immunity (PRNT90) | 0.08 | 0.7654 | |||||
| Infarctions/total cotyledons | −0.09 | 0.7603 | 0.49 | 0.0153 | 1.30 | ||
| Fetal head circumference trajectory | 0.01 | 0.9827 | 0.47 | 0.0169 | 1.20 | ||
| Fetal biparietal diameter growth trajectory | −0.39 | 0.1331 | |||||
| Total placental weight | 0.29 | 0.2974 | 1.08 | 0.0008 | 1.50 | ||
| Birth weight | 0.14 | 0.6117 | |||||
| Focus subgroup of Orientation construct | Duration of maternal plasma viremia | 0.01 | 0.9736 | −1.07 | 0.0171 | 3.8 | 0.76 |
| Peak maternal plasma viral load | −0.21 | 0.5198 | |||||
| Fraction of vRNA -positive cotyledons | −0.26 | 0.3471 | 0.84 | 0.029 | 3.1 | ||
| DENV immunity (PRNT90) | 0.07 | 0.7967 | |||||
| Infarctions/total cotyledons | −0.07 | 0.8174 | 0.49 | 0.0395 | 1.3 | ||
| Fetal head circumference trajectory | 0.06 | 0.8223 | 0.5 | 0.0342 | 1.2 | ||
| Fetal biparietal diameter growth trajectory | −0.38 | 0.1417 | |||||
| Total placental weight | 0.31 | 0.2591 | 1.04 | 0.003 | 1.5 | ||
| Birth weight | 0.04 | 0.8821 | |||||
| Visual Orientation subgroup of Orientation construct | Duration of maternal plasma viremia | −0.1 | 0.7684 | −0.63 | 0.0003 | 7.90 | 0.98 |
| Peak maternal plasma viral load | 0.05 | 0.8793 | |||||
| Fraction of vRNA -positive cotyledons | −0.40 | 0.1428 | 0.30 | 0.0018 | 4.60 | ||
| DENV immunity (PRNT90) | −0.03 | 0.9124 | 0.44 | 0.0001 | 2.10 | ||
| Infarctions/total cotyledons | 0.18 | 0.5189 | 0.17 | 0.0078 | 3.40 | ||
| Fetal head circumference trajectory | −0.53 | 0.0331 * | |||||
| Fetal biparietal diameter growth trajectory | −0.78 | 0.0004 * | −0.77 | <0.0001 | 3.40 | ||
| Total placental weight | 0.48 | 0.0729 | |||||
| Birth weight | 0.13 | 0.6220 | 0.57 | 0.0002 | 4.80 | ||
| Visual tracking subgroup of Orientation construct | Duration of maternal plasma viremia | 0.12 | 0.7141 | No significant predictors | |||
| Peak maternal plasma viral load | −0.06 | 0.8449 | |||||
| Fraction of vRNA -positive cotyledons | −0.11 | 0.6921 | |||||
| DENV immunity (PRNT90) | −0.05 | 0.8572 | |||||
| Infarctions/total cotyledons | −0.29 | 0.2990 | |||||
| Fetal head circumference trajectory | −0.15 | 0.5875 | |||||
| Fetal biparietal diameter growth trajectory | −0.4 | 0.1244 | |||||
| Total placental weight | 0.44 | 0.0971 | |||||
| Birth weight | 0.19 | 0.4732 | |||||
| Auditory orientation subgroup of Orientation construct | Duration of maternal plasma viremia | 0.21 | 0.5101 | −1.8 | 0.0011 | 8.1 | 0.94 |
| Peak maternal plasma viral load | −0.32 | 0.3148 | −0.77 | 0.0045 | 3.2 | ||
| Fraction of vRNA -positive cotyledons | −0.05 | 0.8646 | 1.88 | 0.0008 | 7.4 | ||
| DENV immunity (PRNT90) | 0.17 | 0.5205 | |||||
| Infarctions/total cotyledons | 0.07 | 0.8151 | |||||
| Fetal head circumference trajectory | 0.32 | 0.2323 | 0.57 | 0.0066 | 2.2 | ||
| Fetal biparietal diameter growth trajectory | −0.07 | 0.8054 | −0.94 | 0.0012 | 2.3 | ||
| Total placental weight | 0.05 | 0.8643 | |||||
| Birth weight | 0.04 | 0.8884 | 1.27 | 0.0011 | 4 | ||
| Motor maturity & activity construct | Duration of maternal plasma viremia | 0.57 | 0.0541 | 0.52 | |||
| Peak maternal plasma viral load | 0.40 | 0.1925 | 0.56 | 0.0577 | 1.2 | ||
| Fraction of vRNA -positive cotyledons | 0.43 | 0.1084 | |||||
| DENV immunity (PRNT90) | 0.26 | 0.3248 | |||||
| Infarctions/total cotyledons | −0.14 | 0.6248 | −0.75 | 0.0776 | 2.6 | ||
| Fetal head circumference trajectory | −0.01 | 0.9567 | 0.83 | 0.0332 | 1.9 | ||
| Fetal biparietal diameter growth trajectory | −0.12 | 0.6707 | −1.08 | 0.0458 | 3.9 | ||
| Total placental weight | 0.62 | 0.0128 * | |||||
| Birth weight | 0.30 | 0.2648 | |||||
| Sensory responsiveness construct | Duration of maternal plasma viremia | −0.24 | 0.4620 | No significant predictors | |||
| Peak maternal plasma viral load | 0.19 | 0.5567 | |||||
| Fraction of vRNA -positive cotyledons | −0.1 | 0.7102 | |||||
| DENV immunity (PRNT90) | −0.02 | 0.9483 | |||||
| Infarctions/total cotyledons | 0.21 | 0.4580 | |||||
| Fetal head circumference trajectory | −0.44 | 0.0872 | |||||
| Fetal biparietal diameter growth trajectory | −0.05 | 0.8541 | |||||
| Total placental weight | −0.08 | 0.7905 | |||||
| Birth weight | 0.02 | 0.9309 | |||||
| State control construct | Duration of maternal plasma viremia | −0.66 | 0.0187 * | −0.51 | 0.0877 | 1.00 | 0.19 |
| Peak maternal plasma viral load | −0.63 | 0.0283 * | |||||
| Fraction of vRNA -positive cotyledons | −0.41 | 0.1240 | |||||
| DENV immunity (PRNT90) | −0.24 | 0.3743 | |||||
| Infarctions/total cotyledons | −0.05 | 0.8693 | |||||
| Fetal head circumference trajectory | −0.01 | 0.9828 | |||||
| Fetal biparietal diameter growth trajectory | −0.07 | 0.8034 | |||||
| Total placental weight | −0.28 | 0.3147 | |||||
| Birth weight | −0.07 | 0.8029 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ausderau, K.; Kabakov, S.; Razo, E.; Mitzey, A.M.; Bach, K.M.; Crooks, C.M.; Dulaney, N.; Keding, L.; Salas-Quinchucua, C.; Medina-Magües, L.G.; et al. Neonatal Development in Prenatally Zika Virus-Exposed Infant Macaques with Dengue Immunity. Viruses 2021, 13, 1878. https://doi.org/10.3390/v13091878
Ausderau K, Kabakov S, Razo E, Mitzey AM, Bach KM, Crooks CM, Dulaney N, Keding L, Salas-Quinchucua C, Medina-Magües LG, et al. Neonatal Development in Prenatally Zika Virus-Exposed Infant Macaques with Dengue Immunity. Viruses. 2021; 13(9):1878. https://doi.org/10.3390/v13091878
Chicago/Turabian StyleAusderau, Karla, Sabrina Kabakov, Elaina Razo, Ann M. Mitzey, Kathryn M. Bach, Chelsea M. Crooks, Natalie Dulaney, Logan Keding, Cristhian Salas-Quinchucua, Lex G. Medina-Magües, and et al. 2021. "Neonatal Development in Prenatally Zika Virus-Exposed Infant Macaques with Dengue Immunity" Viruses 13, no. 9: 1878. https://doi.org/10.3390/v13091878
APA StyleAusderau, K., Kabakov, S., Razo, E., Mitzey, A. M., Bach, K. M., Crooks, C. M., Dulaney, N., Keding, L., Salas-Quinchucua, C., Medina-Magües, L. G., Weiler, A. M., Bliss, M., Eickhoff, J., Simmons, H. A., Mejia, A., Antony, K. M., Morgan, T., Capuano, S., III, Schneider, M. L., ... Mohr, E. L. (2021). Neonatal Development in Prenatally Zika Virus-Exposed Infant Macaques with Dengue Immunity. Viruses, 13(9), 1878. https://doi.org/10.3390/v13091878