1. Introduction
Global food security is constantly threatened by emerging pathogens, such as plant viruses. Plant viruses cause outbreaks that reduce the yield and the quality of the crops that sustain the food chain [
1]. The globalization of the vegetable trade, together with the limited effectiveness of monitoring and eradication measures, favor the worldwide spread of plant viruses. In addition, the frequency and scale of plant virus outbreaks are predicted to increase due to climate change, which may facilitate the expansion of new or existing viruses to geographic areas in which they were not previously present or epidemic [
2]. This is the case of tomato brown rugose fruit virus (ToBRFV) (genus
Tobamovirus, family
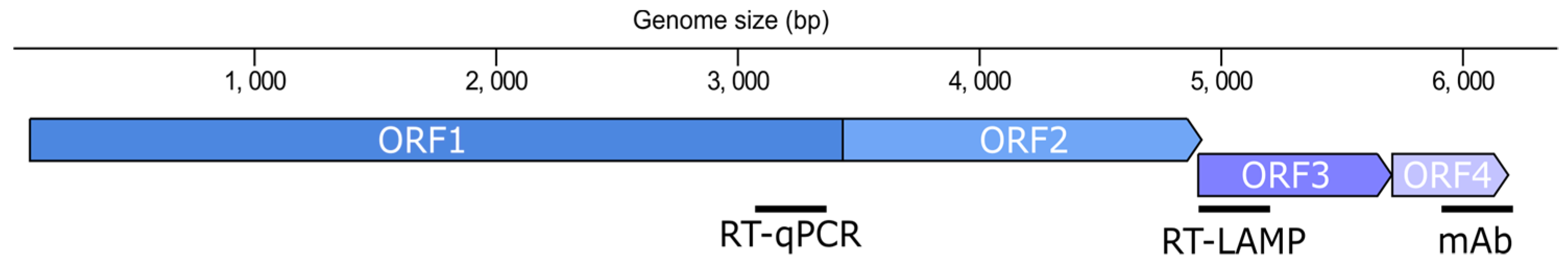
Virgaviridae). ToBRFV, as other tobamoviruses, has a particle with a rod-shaped morphology formed by multimers of the capsid protein (CP) that encapsulate a positive single-stranded RNA (ssRNA(+)) genome of 6.2 to 6.4 kb encoding four open reading frames (ORFs) (
Figure 1). ORF1 and ORF2 are separated by a leaky stop codon and encode non-structural proteins that assemble the RNA-dependent RNA polymerase (RdRP). ORF1 encodes the small subunit (124–132 kDa) and ORF2 the large subunit of the RdRP (181–189 kDa) which contains the polymerase domain. The downstream ORFs encode the 28–31 kDa movement protein (MP) and the 17–18 kDa CP, which are translated from their respective subgenomic RNAs.
The tobamovirus group includes well-known viruses that affect tomato and other solanaceous species, such as tobacco mosaic virus (TMV) and tomato mosaic virus (ToMV) among others [
3,
4]. Thus far, the
Tm-1, Tm-2/
Tm-22 resistance genes have routinely been used in tomato hybrid breeding to protect new varieties against TMV and ToMV [
5,
6]. However, the
Tm-22 genetic resistance which lasted unbroken for over sixty years has now been overcome by ToBRFV. Apparently, the MP of ToBRFV is the genetic determinant for
Tm-22 resistance-breaking [
7]. Similarly, the
L gene alleles have been used for the protection of most pepper commercial cultivars against tobamoviruses. ToBRFV induces a hypersensitive response in plants harboring these resistance genes [
8], and infections of sweet pepper lacking these resistances have been reported in Italy [
9]. On the other hand, the viral particle of ToBRFV is very stable and resilient to commonly used disinfectants [
10], which may facilitate its mechanical transmission even by pollinators [
11]. Taken together, the particular features of ToBRFV have contributed to its rapid spread worldwide [
12,
13,
14,
15,
16,
17,
18,
19,
20,
21] since it was first reported in Israel and Jordan in 2014 [
8,
22]. In an attempt to delay or arrest ToBRFV expansion in Europe, it has been included in the European Plant Protection Organization (EPPO) alert list (Commission Implementing Decision EU 2019/1615) and in the list of quarantine bodies (Commission regulation EU 2019/2072).
Detection of infected plants or seeds is critical for the success of the intervention and eradication strategies to prevent further expansion of ToBRFV. In this matter, the EPPO published a Standard that describes a diagnostic protocol for the detection and identification of this virus (PM7/146(1)). This protocol distinguishes between plant and seed materials. Thus, symptomatic plant material can be processed by enzyme-linked immunosorbent assay (ELISA), while the recommendation for asymptomatic material is the analysis by coupled reverse transcription-polymerase chain reaction (RT-PCR) and real-time quantitative PCR (RT-qPCR). In contrast, seed material must be exclusively analyzed by RT-qPCR. The RT-PCR should be performed using primers from Alkowni et al., 2019 [
19] or Rodriguez-Mendoza et al., 2019 [
23], while the RT-qPCR should be carried out by using a duplex real-time test with CaTa28 and CSP1325 primers and probe proposed by the International Seed Federation (ISF) in 2020. Apart from the EPPO’s Standards, other RT-qPCR tests have been developed for ToBRFV detection and compared with RT-PCR tests [
24]. As an alternative to these tests, isothermal amplification techniques such as loop-mediated isothermal amplification (LAMP) [
25] are gaining popularity due to their simplicity and similar performance compared to RT-qPCR. Many tests have been developed using LAMP to diagnose plant viruses, including ToBRFV [
26,
27].
In spite of the existence of a handful of resources for the detection of ToBRFV, a comprehensive comparison that facilitates their selection is still missing. In this work, we have designed, assessed, and compared new tests for the specific and sensitive detection of the ToBRFV viral particle and its genome. To this end, we have targeted the CP and two different ORFs (
RdRP and
MP) (
Figure 1) by employing three different methods: DAS-ELISA, RT-qPCR, and RT-LAMP. Altogether, we provide an array of new resources for ToBRFV testing at the protein and nucleic acid levels that are specific to this virus and show an equivalent sensitivity to those of the EPPO-recommended tests.
2. Materials and Methods
2.1. Virus Isolates and Plant Inoculation
The Spanish ToBRFV isolate from Vicar (Almeria, Spain) [
18] was provided by the “Laboratorio de Producción y Sanidad Vegetal” (La Mojonera, Almería, Spain). Isolates of other tobamoviruses used in this study were acquired from the DSMZ (Leibniz, Germany) collection: TMV (PV-1252), ToMV (PV-0141), pepper mild mottle virus (PMMoV, PV-0093), tobacco mild green mottle virus (TMGMV, PV-0124). Approximately 50 mg of dried plant tissue was homogenized for each isolate in 2 mL of 30 mM phosphate buffer pH = 8 using a mortar and pestle. Homogenates were used to mechanically inoculate leaves of 25–26-day-old
N. benthamiana plants (4–5-true leaves) and the first pair of true leaves of 7–10-day-old tomato plants (cultivar M82). For this, the leaves to be inoculated were first dusted with carborundum powder (600 mesh) and then manually rubbed with the homogenate. The plants thus inoculated were kept separately in a confined greenhouse under controlled conditions set with a 16/8 h photoperiod and a 26/22 °C day/night cycle. After 10 to 15 days, systemic leaves that showed obvious symptoms of infection were collected, cut, mixed, and divided into samples of approximately 100 mg. The samples were frozen in liquid nitrogen, ground with a Retsch Mixer Mill MM400 (ThermoFisher, Hampton, NH, USA) for 1 min at 30 Hz, and stored at −80 °C for later analyses.
2.2. Design and Production of Monoclonal Antibodies against ToBRFV
The monoclonal antibodies (MoAbs) were produced by GenScript (Piscataway, NJ, USA) using standard methods [
28,
29]. Briefly, the ToBRFV CP epitope (
Figure 2) was synthesized and used as an immunogen to be injected into five 6-week-old BALB/c mice. Production of hybridoma-secreting MoAbs against the ToBRFV was performed according to [
30] with minor modifications. Hybridoma supernatants were screened for the presence of anti-CP peptide antibodies by indirect ELISA in 96-well plates and Western-blotting. Positive hybridoma clones were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM) high glucose (WISENT Bioproducts, Quebec, Canada), supplemented with 10% fetal bovine serum (WISENT Bioproducts) and 1% Penicillin-Streptomycin (Sigma-Aldrich, Saint Louis, MO, USA). Cells were allowed to grow at 37 °C and supplemented with 5.5% of CO
2. The hybridoma was then injected intraperitoneally into pristine primed syngeneic BALB/c mice to produce ascites. After 7–10 days, the ascites samples were collected and their titers were determined by indirect ELISA. The isotypes of the MoAbs were determined using an isotyping kit following the manufacturer’s instructions (Sigma-Aldrich). The anti-CP peptide IgG was purified from ascites with an immobilized protein-G affinity column (GE Healthcare, Wauwatosa, WI, USA) according to the manufacturer’s manual. Purified antibodies were stored at −80 °C.
2.3. Western Blot
Healthy and ToBRFV-, TMV- or ToMV-infected tomato leaves were ground in liquid nitrogen using a mortar and a pestle. The plant material was solubilized in 4 mL per g of RIPA buffer (10 mM Tris-HCl pH = 7.5; 150 mM NaCl; 0.5 mM EDTA; 0.1% SDS; 1% Triton X-100; 1% Deoxycholate) and mixed thoroughly. The homogenate was centrifuged at 3000× g for 15 min at 4 °C, then the supernatant was transferred to a new tube and centrifuged at 12,000× g for 15 min at 4 °C. The extracts were kept at −20 °C. The proteins were resolved in 15% SDS-PAGE gels and blotted onto nitrocellulose membranes (GE Healthcare) using a Trans-Blot (Bio-Rad, Hercules, CA, USA). After blocking, the membranes were incubated with a 1/250 dilution of antisera from immunized mice against the ToBRFV CP peptide or with 1/2 hybridoma supernatant dilutions. Next, an HRP-conjugated anti-mouse secondary antibody (Promega, Madison, WI, USA) was used at a 1/2500 dilution. As a control, a commercial ToBRFV antibody (DSMZ, Leibniz, Germany) was used at 1/1000 dilution and detected with an HRP-conjugated anti-rabbit secondary antibody (Promega) at 1/2500 dilution. The membranes were developed using SuperSignal West Pico PLUS (ThermoFisher, Hampton, NH, USA) and an Amersham Imager 600 (GE Healthcare Life Sciences, Wauwatosa, WI, USA).
2.4. DAS-ELISA
Plant extracts were obtained by homogenizing ground tissue with extraction buffer (137 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, 1.8 mM KH2PO4, 0.05% Tween-20, 20 g/L polyvinyl pyrrolidone (K10-K40), 2g/L bovine serum albumin, pH = 7.4) in a 1:5 ratio (μg: mL). The homogenate was centrifuged for 10 min at 13,000 rpm. The supernatant was recovered and used in the ELISA as follows. A 96-well plate (ThermoFisher) was coated with 50 μL/well of the capture antibody diluted in coating buffer (15 mM Na2CO3, 35 mM NaCO3, pH = 9.6) and incubated for 3 h at 37 °C. This was followed by four washes with 200 μL/well of wash buffer (137 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, 1.8 mM KH2PO4, 0.05% Tween-20, pH = 7.4). Afterwards, 50 μL/well of the plant extract were added and incubated overnight at 4 °C. Once washed four times with 200 uL/well of wash buffer, 50 μL/well of the secondary antibody (tagged with the alkaline phosphatase reporter enzyme) was added, and the plate incubated for 3 h at 37 °C, after which the washing step was repeated as above. Lastly, the enzymatic activity was measured after adding 50 μL of the substrate solution (1 mg/mL 4-nitrophenylphosphate-di-Na-salt in substrate buffer (1 M diethanolamine, pH = 9.8) and incubating the plate for 60 min, then the absorbance was read at 405 nm using a TECAN Sunrise (Männedorf, Switzerland). The AbCam Alkaline phosphatase Conjugation Kit—Lightning-Link® (AbCam, Cambridge, UK) was used to conjugate the secondary antibody following the manufacturer´s instructions.
2.5. RNA Extraction
RNA extraction was performed using the NucleoSpin RNA plant kit (MACHEREY-NAGEL, Düren, Germany) following the manufacturer’s instructions. The RNA extracts were checked by electrophoresis in a 1% agarose gel, and the RNA concentration was measured with a Nanodrop ™ One (ThermoFisher), and adjusted to a working concentration of 10 ng/µL, to be used as a template for the RT-LAMP and RT-qPCR. A healthy plant RNA extract was used as the diluent to adjust the concentrations.
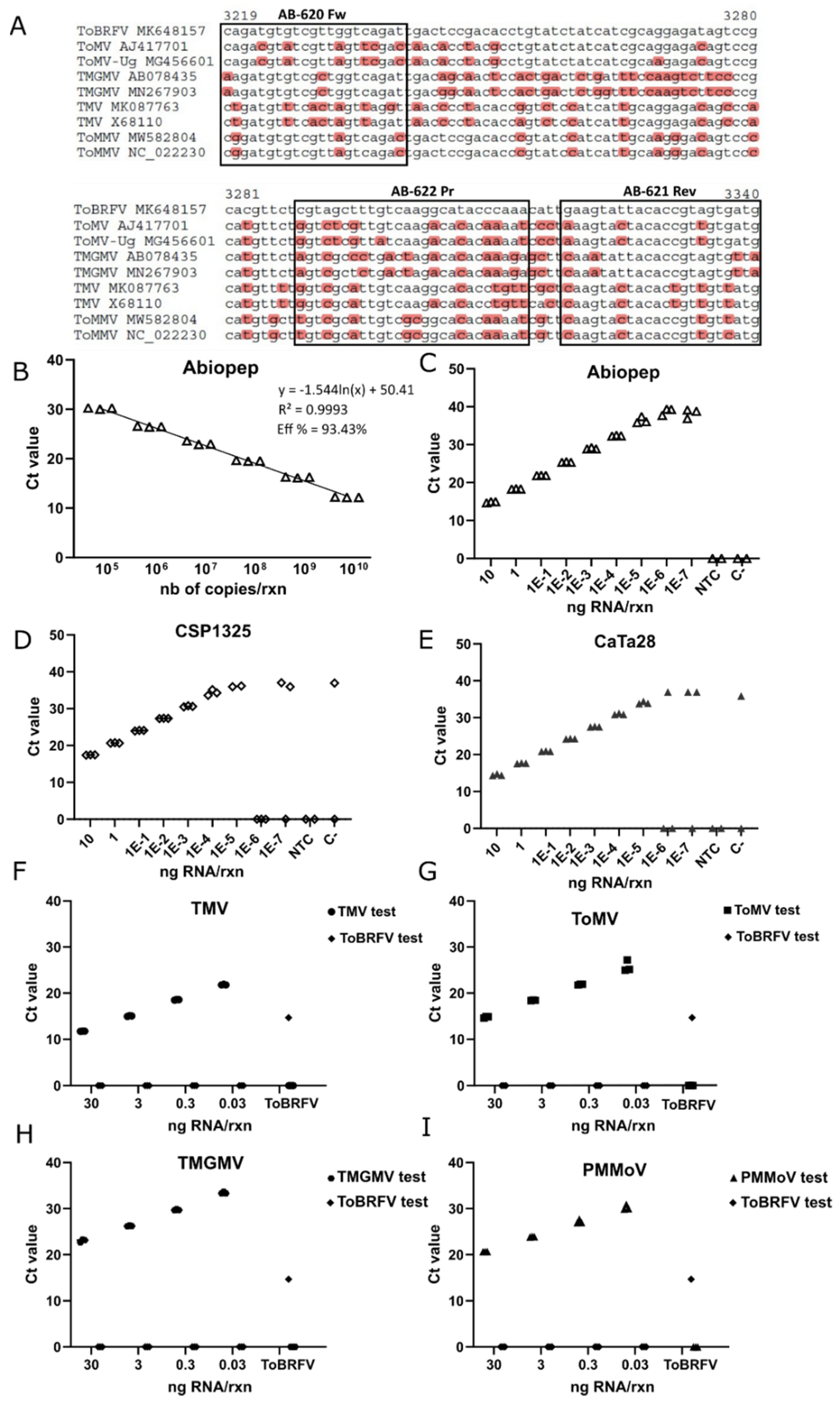
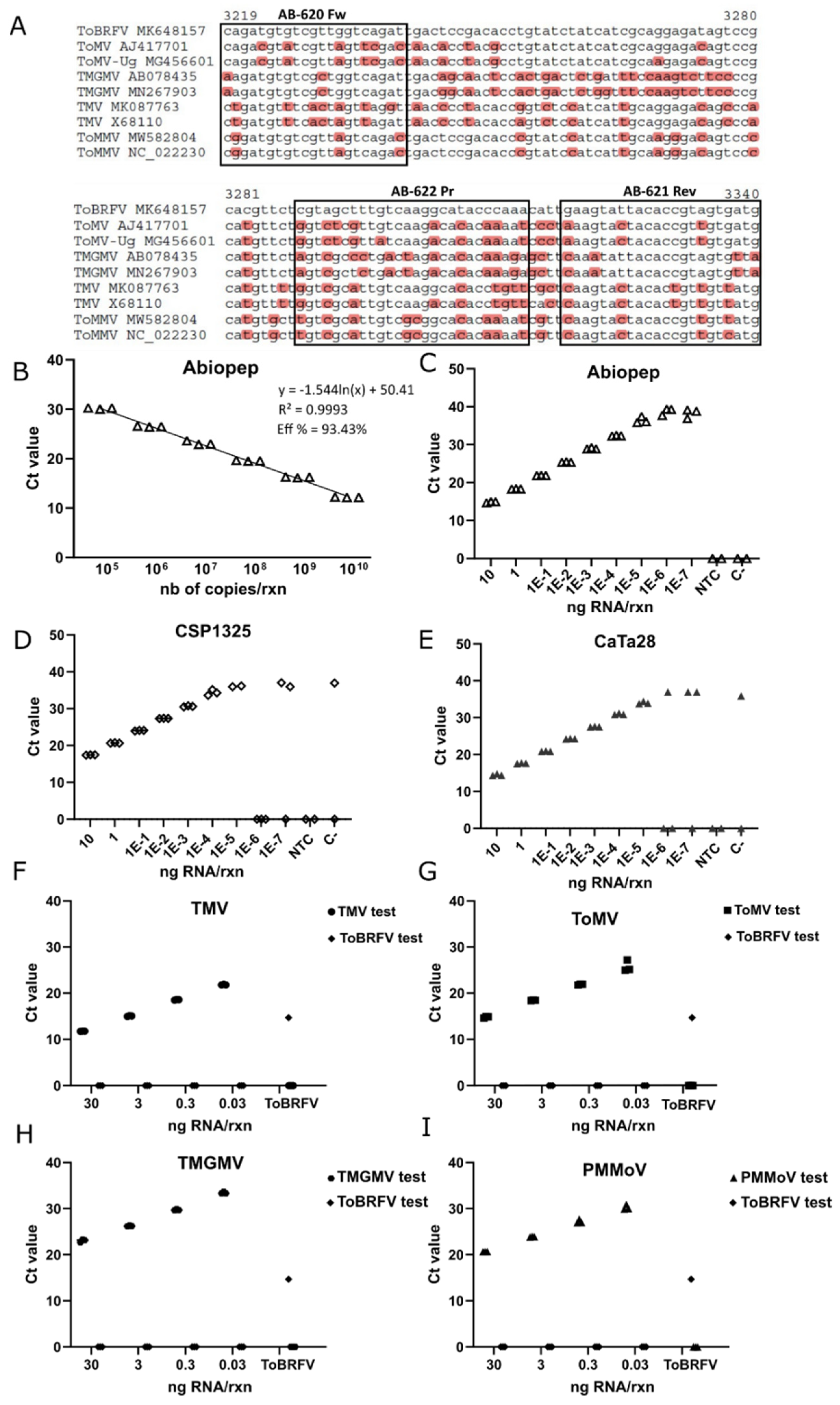
2.6. Primers and Probe Design for One-Step RT-qPCR
A total of 71 complete sequences for ToBRFV, 31 complete sequences for the ToMV, 40 complete sequences for TMV, 16 complete sequences for PMMoV and 26 partial sequences for TMGMV were retrieved from the NCBI database to assess inclusivity for each RT-qPCR test. Sequence alignments were performed with the ClustalW tool [
31] included in the MEGA7 software [
32]. For RT-qPCR tests, a 122 bp region within the
RdRP was chosen for ToBRFV, a 152 bp region of the
RdRP for ToMV, a 148 bp of the
RdRP for TMV, a 144 bp region of the
CP for PMMoV, and a 106 bp segment of the
CP for TMGMV. The Primer3 plus program was used to design the primers and probe set, and the theoretical properties of the designed assay were analyzed with the Oligo Analyzer tool from IDT (Integrated DNA Technologies, v3.1.1). The primers and probe exclusivity were also evaluated in silico against the other tobamoviruses known to affect tomato plants. Both the primers and probe were synthesized by IDT (Newark, NJ, USA) (
Table S2).
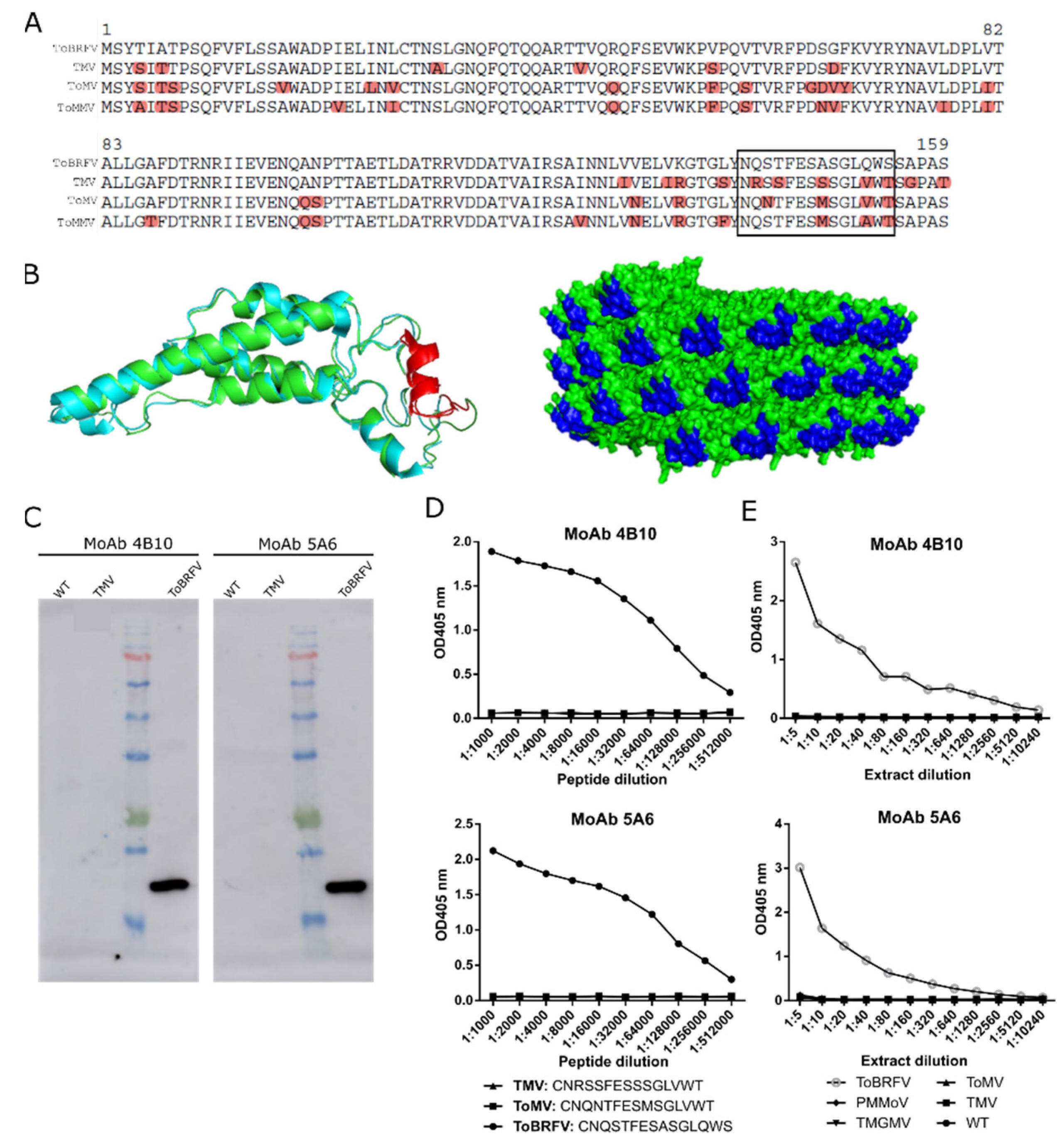
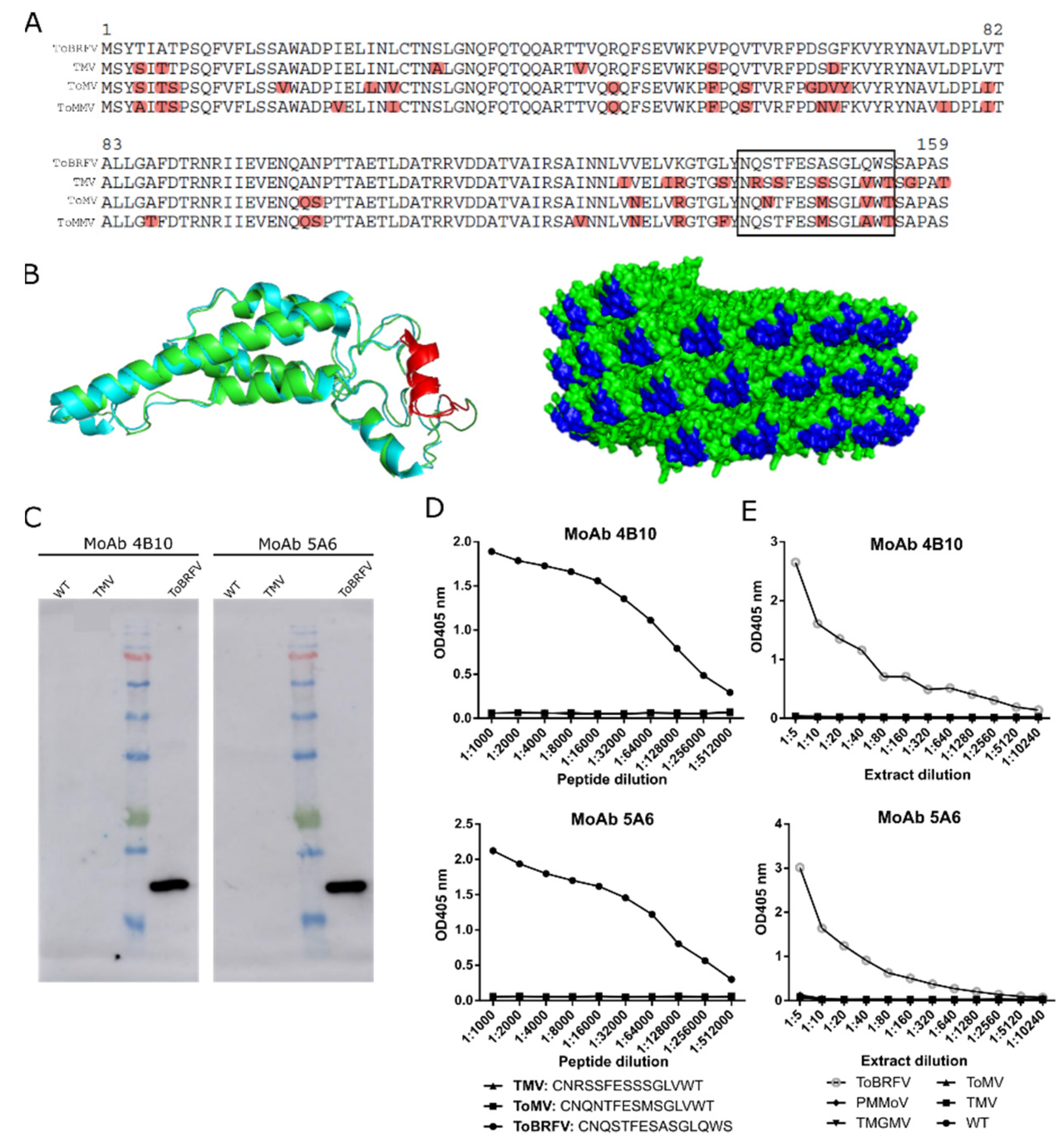
Figure 2.
Design and evaluation of two monoclonal antibodies (MoAb 4B10 and MoAb 5A6) against ToBRFV CP. (
A) Clustal alignment of the CP polypeptide sequences from ToBRFV (NC_028478), TMV (NC_001367), ToMV (NC_002692), and ToMMV (NC_022230) for the identification of a ToBRFV peptide candidate to give rise to specific ToBRFV antibodies (squared is the best candidate). (
B) On the left, ribbon representation of the structural superposition of the TMV (blue) and the predicted ToBRFV CPs (green). The selected peptide is highlighted in red. On the right, molecular surface representation of the assembled virion particle of TMV with the selected peptide marked in blue. The TMV CP structure was obtained from the protein data bank (2om3, [
33]) and the ToBRFV CP sequence from Uniprot (A0A0S2SZX3, [
34]). Protein structure visualization was performed with Pymol. (
C) Western blot analyzing the exclusivity of MoAb 4B10 and MoAb 5A6. Extracts from healthy (WT), TMV- and ToBRFV-infected plants were loaded. (
D) Indirect ELISA results showing the exclusivity of MoAb4B10 or 5A6 when using serial dilutions of synthetic peptides (squared in (A)) for ToBRFV, ToMV, and TMV. (
E) Analytical sensitivity and exclusivity of the DAS-ELISA tests devised with the monoclonal antibodies. Each data point represents a single measurement.
Figure 2.
Design and evaluation of two monoclonal antibodies (MoAb 4B10 and MoAb 5A6) against ToBRFV CP. (
A) Clustal alignment of the CP polypeptide sequences from ToBRFV (NC_028478), TMV (NC_001367), ToMV (NC_002692), and ToMMV (NC_022230) for the identification of a ToBRFV peptide candidate to give rise to specific ToBRFV antibodies (squared is the best candidate). (
B) On the left, ribbon representation of the structural superposition of the TMV (blue) and the predicted ToBRFV CPs (green). The selected peptide is highlighted in red. On the right, molecular surface representation of the assembled virion particle of TMV with the selected peptide marked in blue. The TMV CP structure was obtained from the protein data bank (2om3, [
33]) and the ToBRFV CP sequence from Uniprot (A0A0S2SZX3, [
34]). Protein structure visualization was performed with Pymol. (
C) Western blot analyzing the exclusivity of MoAb 4B10 and MoAb 5A6. Extracts from healthy (WT), TMV- and ToBRFV-infected plants were loaded. (
D) Indirect ELISA results showing the exclusivity of MoAb4B10 or 5A6 when using serial dilutions of synthetic peptides (squared in (A)) for ToBRFV, ToMV, and TMV. (
E) Analytical sensitivity and exclusivity of the DAS-ELISA tests devised with the monoclonal antibodies. Each data point represents a single measurement.
![Viruses 13 01680 g002]()
2.7. In Vitro Transcription and Standard Curve
A synthetic fragment containing part of the ToBRFV ORF1 was synthetized by GenScript (Piscataway, NJ, USA) and cloned in a pBluescript II KS (+) vector (Stratagene, La Jolla, CA, USA). In vitro transcription was performed with T3 polymerase (Promega, Madison, WI, USA) in a 20 μL reaction according to the manufacturer’s instructions. After ethanol precipitation, the transcripts were resuspended in 20 μL of RNase-free water and the RNA concentration was measured spectrophotometrically with a Nanodrop ™ One (ThermoFisher). The standard curve was prepared from serial dilutions of the in vitro transcript in a 1:10 range, diluted in healthy tomato RNA extracts to a final concentration of 16 ng/μL.
2.8. One-Step RT-qPCR
The one-step RT-qPCR reactions were performed using the KAPA PROBE FAST Universal One-Step qRT-PCR kit (KAPA Biosystems, Wilmington, MA, USA) following the manufacturer’s instructions. Two microliters of nucleic acid preparation were added to a final volume of 10 μL. The reactions were carried out in a StepOne Plus ™ Real-Time PCR System (Applied Biosystems, Waltham, MA, USA) thermocycler, and the reaction conditions were a reverse transcription step of 5 min at 42 °C, followed by PCR with a 3 min denaturation cycle at 95 °C and 40 amplification cycles of 3 s at 95 °C, and 30 s at 60 °C. Three replicates were analyzed per plot/sample and two negative controls were included in each experiment, which consisted of RNA extracts of healthy tomato leaf diluted to a final concentration of 30 ng/μL, and water in the non-template control (NTC).
2.9. RT-LAMP
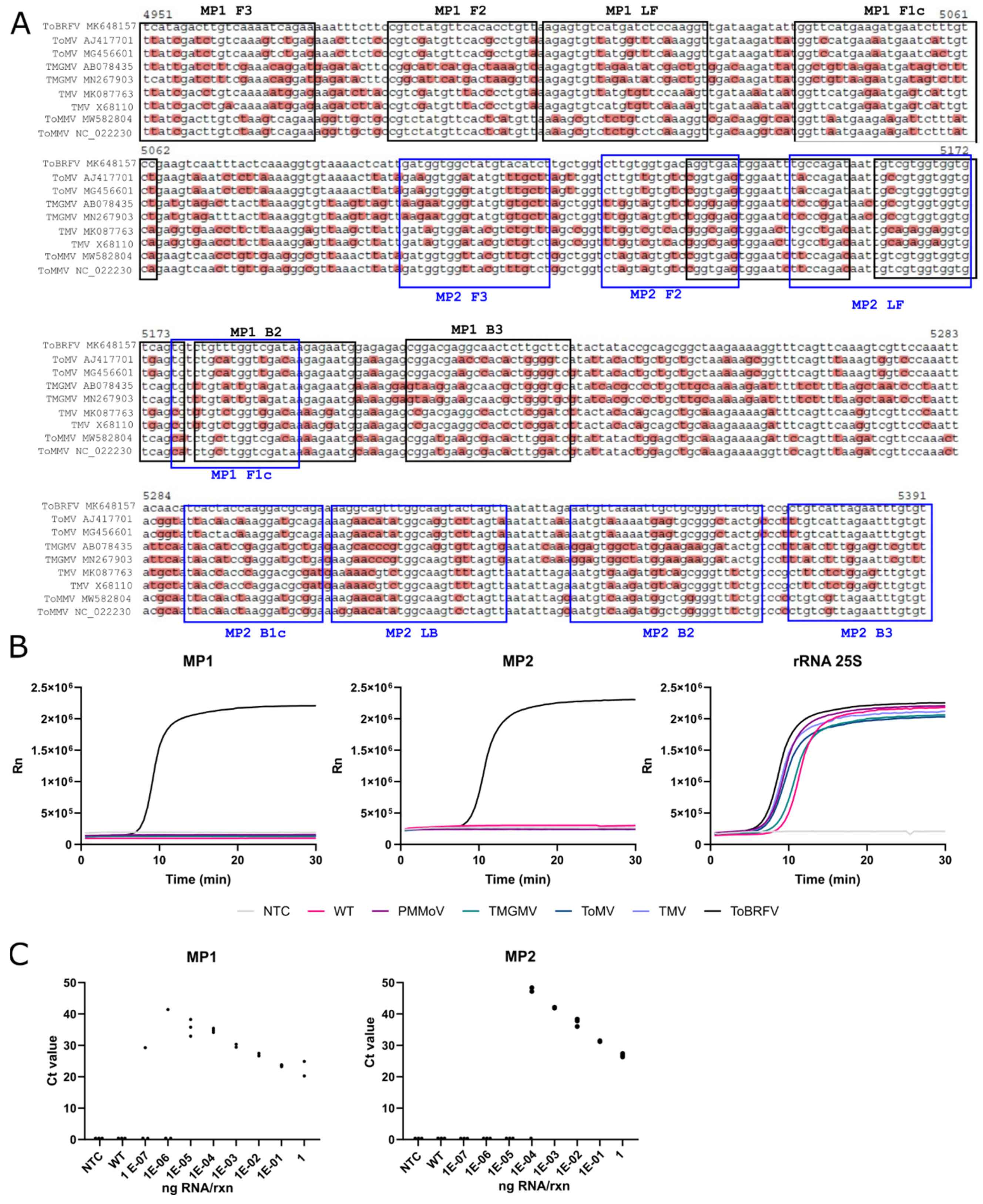
LAMP primers were designed using PrimerExplorer v.5 (
https://primerexplorer.jp/e/, accessed on 22 May 2020). Two sets of primers against the
MP of ToBRFV were designed (MP1 and MP2,
Table S2). An additional set of primers targeting the tomato ribosomal RNA 25S subunit was also designed as an internal control (rRNA 25S,
Table S2). RT-LAMP reactions were performed using the WarmStart LAMP Kit (NEB, Ipswich, MA, USA) at a final volume of 10 μL. Primers were added at a final concentration of 0.2 μM for F3 and B3, 1.6 μM for FIP and BIP primers, and 0.8 μM for LF and LB primers. Reactions were performed independently for each set of primers (MP1, MP2, and rRNA 25S) using 2 μL of input RNA. The amplification was performed at a constant temperature of 62 °C for 25–30 min (50–60 cycles of 30 s each) in an Applied Biosystems StepOnePlus Real-Time PCR System (Waltham, MA, USA), and tracked with a DNA-intercalating green fluorophore provided with the WarmStart LAMP Kit (NEB).
2.10. ToBRFV Timecourse Experiment
In total, twenty-one 5-week-old tomato plants cv. M82 (3–4 pairs of leaves approximately) were mechanically inoculated (see above). Three plants per data point (1, 2, 3, 4, 6, 8, 12 days post-inoculation) were sampled, collecting the first pair of newly emerged leaves in which the virus has replicated systemically. Then, samples were finely sliced and two subsamples prepared, one containing 100 mg for RNA extraction, and another with 200 mg for protein extraction. These subsamples were frozen in liquid nitrogen, ground with a Retsch Mixer Mill MM400 (ThermoFisher) for 1 min at 30 Hz, and stored at −80 °C for later analyses.
4. Discussion
Emergent plant viruses are a constant threat that need to be addressed quickly to avoid or limit the expansion of the new pathogens. Thus, specific testing methods that allow the identification of emergent viruses among their relatives are crucial for monitoring the progression of epidemics and implementing effective eradication measures. ToBRFV is an example of an emergent plant virus which has expanded from the Mediterranean basin to practically all the continents. Therefore, in this work, we developed an array of techniques aimed at detecting the CP and several regions of the ToBRFV genome (
Figure 1): two DAS-ELISA tests against the CP, a RT-qPCR test for the
RdRP, and a RT-LAMP test for the
MP.
The tobamovirus virion particle is composed of thousands of CP subunits that are helicoidally arranged, wrapping the ssRNA(+), and forming a rod-shaped structure [
35]. The repetition of the CP in the virion particle makes it an attractive candidate for obtaining antisera against this virus, particularly if targeting externally localized peptides. This is the most straightforward approach and the one that most of the commercial antisera follow. However, the immunization with the whole virion or CP protein yields polyclonal antisera that are likely to have cross-reactivity with other tobamoviruses, especially in the case of ToBRFV, which shares a high degree of similarity with ToMV and TMV. In contrast, MoAbs are directed against singular epitopes and are less likely to cross-react, a feature of great interest when an emergent virus is very similar to other widespread viruses. The drawback though, is that MoAbs are more difficult to obtain, and the process takes more time at a higher cost. Nevertheless, we reasoned that the extra effort and time for the development of a MoAb was justified in this case. Thus, we identified a peptide at the C-terminus of the ToBRFV CP exposed to the outer surface of the virion particle, which potentially bore enough dissimilarities with ToMV and TMV to produce specific antibodies. We used this peptide to immunize mice, and after three rounds of cloning and selection, we obtained two MoAbs which showed no cross-reactivity with the TMV CP nor with the equivalent peptides from TMV or ToMV. Next, we formulated two DAS-ELISA tests which were exclusive to ToBRFV, and which did not show cross-reactivity with TMGMV, PMMoV, ToMV, or TMV. The DAS-ELISA test formulated with the MoAb 4B10 was more sensitive than the MoAB5A6, detecting ToBRFV at least at one dilution higher. However, this difference in the analytical sensitivity was not noticed during a timecourse infection experiment, as both tests performed similarly, detecting ToBRFV after 6 dpi. From 6 to 12 dpi, the signal from both tests remained steady, perhaps indicating detection saturation. In comparison, nucleic acid amplification tests showed lower Ct values during the course of the infection, suggesting that the virus was still accumulating in the plant tissue despite the DAS-ELISA being unable to sense this increase in the viral load.
RT-qPCR is the gold standard when discussing sensitivity in virus detection. This is especially important when the virus titer is low, as may occur in infected seed stocks [
36]. The EPPO recommendation is to use the CaTa28 and CSP1325 ISF-ISHI-Veg RT-qPCRs tests. The first test targets the
MP, whereas the second is directed to the
CP. In addition to this, Panno et al., 2019 [
24] reported a RT-qPCR test targeting the
MP of ToBRFV. Therefore, we decided to target the
RdRP ORF which was still untargeted. The results showed that our test could be useful for detecting ToBRFV per se, or it could be complementarily used with the aforementioned tests which target different regions of the ToBRFV genome. Under our particular conditions of reagents, equipment, and operators, the CaTa28 test was the most sensitive of either ISHI-veg tests, detecting ToBRFV in a 1E-5 ng/rxn dilution, one order of magnitude lower than CSP1325. Beyond these dilutions, the stability of the Ct measurements decreased, as reflected by some replicates which failed to amplify or produced high Ct values. In contrast, the Abiopep test continued to produce a clear signal in the rest of the dilutions, stabilizing the Ct values to around 39, which, in fact, may complicate the selection of a Ct threshold. To solve this, we collected all the Ct values coming from negative samples, including NTC, WT, and samples infected with other tobamoviruses, and observed that the lower value was close to 35. Following this cut-off value of <35, the last dilution detected by the Abiopep test was 10E-4 ng/rxn, the same as the CSP1325 test and one dilution less than the CaTa28 test. Nevertheless, it is important to remark that this threshold needs to be adjusted for each laboratory. For instance, the ISF-ISHI-Veg protocol sets a threshold at Ct <32 for the positive amplification control, whereas Botermans’ group, which followed this protocol for tracking ToBRFV outbreaks in the Netherlands, established a lower Ct threshold of <30 [
21]. Importantly, aside from the in silico analysis of the primers and probe specificity, we evaluated the exclusivity of the Abiopep RT-qPCR test by using RNA extracts from plants infected with different tobamoviruses and a specific pair of primers for each one. This assay experimentally confirmed the exclusivity of our Abiopep test, which only showed amplification for ToBRFV-positive samples, while the samples infected with the other viruses were positive with their particular set of primers and probe.
Genetic testing methods are shifting from the laboratory to the field, accelerating the decision-making process while reducing costs. To date, point-of-care testing approaches are mainly reserved for antibody tests such as immunochromatographic strips. However, these testing methods are less sensitive and specific compared with the nucleic acid amplification methods. As a result, isothermal amplification methods that do not require sophisticated equipment are gradually gaining popularity. To our knowledge, two works have been published describing LAMP tests to detect ToBRFV [
26,
27]. Both protocols consisted of one set of primers targeting the
RdRP ORF, and were compared with published RT-PCR tests [
14,
19], confirming that the LAMP method had a higher sensitivity as had been previously demonstrated for other LAMP tests targeting different plant viruses [
37,
38]. Conversely, in this work, we targeted two different segments of the
MP ORF (MP1 and MP2), adding an IC (25S) and thus facilitating the interpretation of the results. In our LAMP test, a (i) positive sample should provide a clear amplification for all three targets (MP1, MP2 and 25S), a (ii) negative sample should fail to amplify MP1 and MP2 targets while amplifying 25S, a (iii) doubtful situation would be when only one MP target (MP1 or MP2) amplified in conjunction with 25S. Examples of these outputs can be found in
Table 1. Unlike Sarkes et al., 2020 [
26] which tested the exclusivity using synthetic gene fragments of TMV and ToMV, Rizzo et al., 2021 [
27] used plant RNA extracts obtained from plants infected with different tobamoviruses. We confirmed the exclusivity of our test following the same strategy as the second study, showing that the MP1 and MP2 sets only amplified samples infected with ToBRFV, whereas the 25S set amplified all the samples except NTC. We used the same RNA extracts from ToBRFV-infected tomato plants to evaluate the sensitivity of the Abiopep RT-qPCR test and the MP1 and MP2 LAMP tests, thus allowing the comparison between both methods. Both primer sets displayed sensitivities equal to RT-qPCR tests, with a limit of detection of 10E-5 ng/rxn for the MP1 set and 10E-4 ng/rxn for the MP2 set.
Altogether, with the experiments discussed above, we evaluated fundamental parameters such as analytical sensitivity, specificity (including inclusivity and exclusivity), repeatability and reproducibility of our tests, although, like for most analyses described in the literature, with some limitations. The analytical sensitivity was evaluated for all the tests by assaying serial dilutions of an extract from an infected plant in a healthy plant tissue extract, and also in an infection timecourse experiment. Regarding the analytical specificity, only the Spanish ToBRFV isolate was evaluated to test the inclusivity since this virus is included in the EPPO’s A2 list of quarantine pests, and access to overseas isolates is restricted. Nevertheless, the relatively short evolutionary history of ToBRFV should guarantee the inclusivity of our tests as in silico analysis of the sequences showed. To ensure the exclusivity of our tests, relevant non-target tobamoviruses which are known to infect tomato and pepper (TMV, ToMV, PMMoV, TMGMV) were tested. As happened with the inclusivity analyses, the evaluation of the alignments that we performed when designing our tests should also ensure the exclusivity of our tests. Selectivity was evaluated using tomato and N. benthamiana matrices with no noticeable impact on the performance of our tests. Finally, the repeatability of the tests was shown with the twenty-one plants with different viral titers that we evaluated in the timecourse experiment. Each time point was represented by three biological replicates, each one assessed by three technical replicates.
In conclusion, we successfully achieved the goal of this work, which consisted of providing new tools for the specific and sensitive detection of ToBRFV. To our knowledge, this is the first report that gathers and describes the development, evaluation, and comparison of three testing methods with different foundations to detect ToBRFV in infected N. benthamiana and tomato leaves. Remarkably, these methods were compared using the same set of samples to provide clues about their selection. In the future, more tobamovirus isolates as well as matrices (e.g., pepper leaves, pepper and tomato seeds) need to be tested to expand our set of data referring the inclusivity, exclusivity, and selectivity of the tests. Additionally, more RT-qPCR amplification reagents need to be assessed to delimitate which one gives the best diagnostic performance. Meeting all standards of supra-national plant health authorities (e.g., the EPPO standards in Commission Implementing Regulation 1191/2020) will help in the generalization of the use of our methods.


 ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}