Low Host Abundance and High Temperature Determine Switching from Lytic to Lysogenic Cycles in Planktonic Microbial Communities in a Tropical Sea (Red Sea)


Abstract
:1. Introduction
2. Materials and Methods
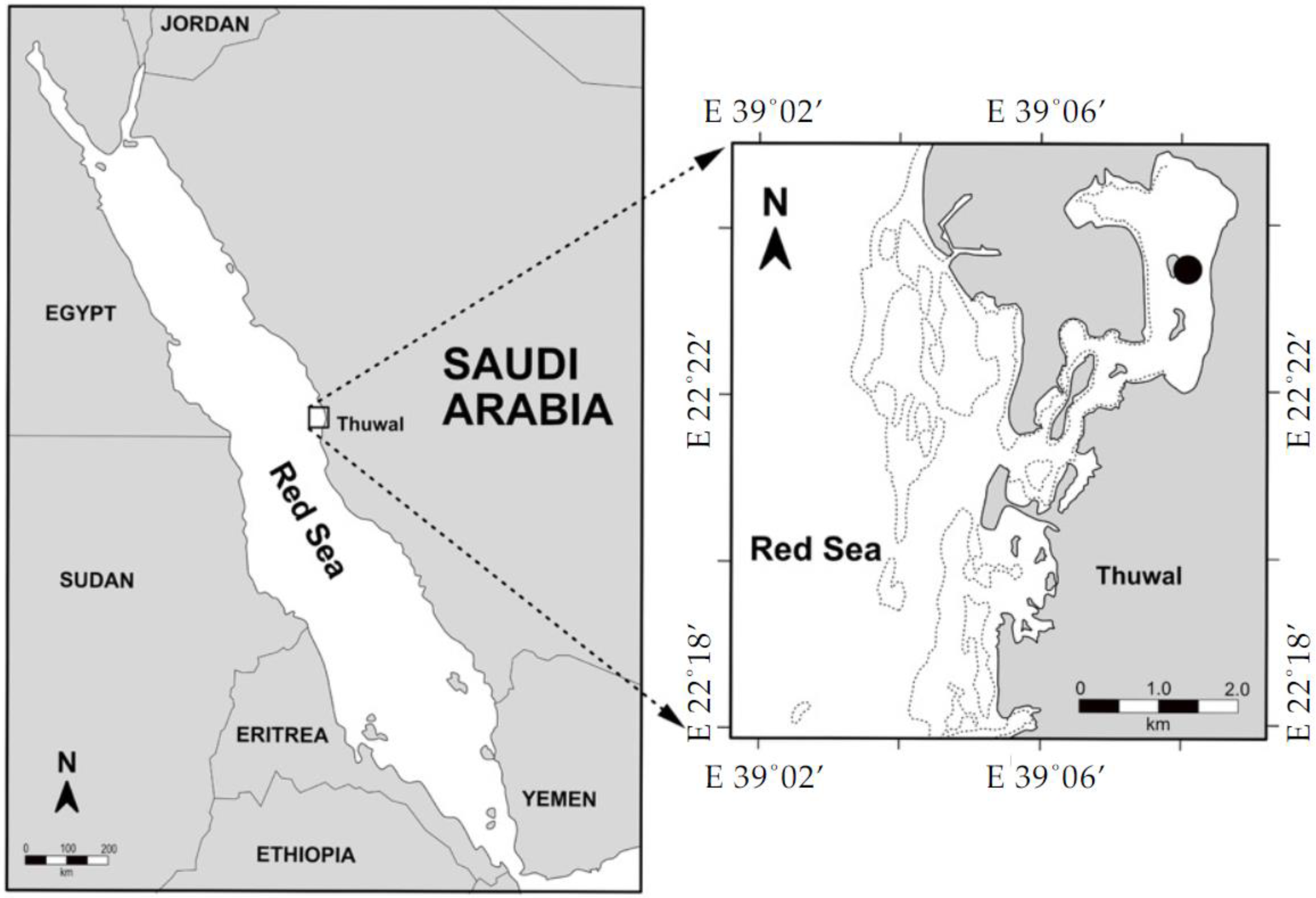
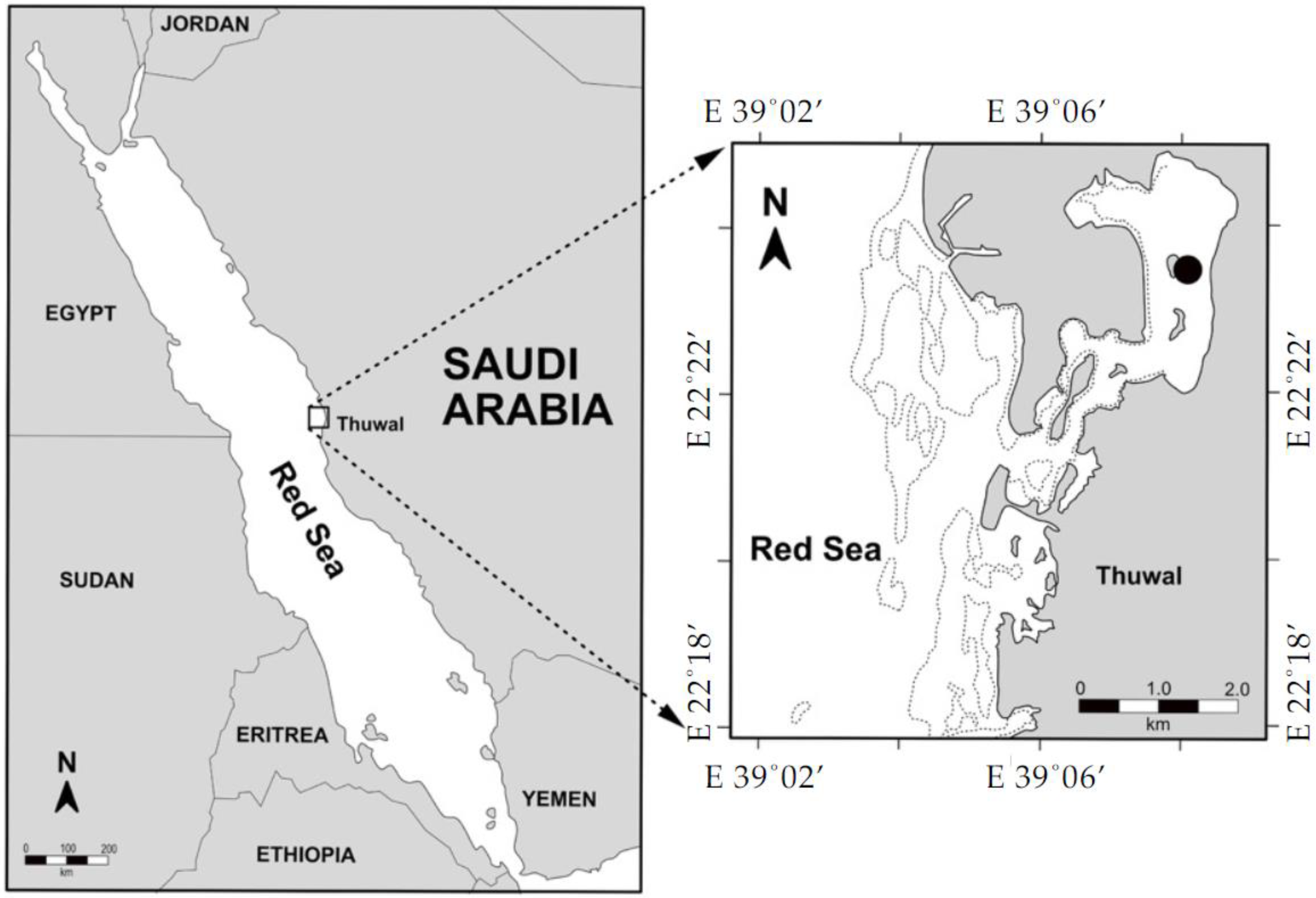
2.1. Study Sites and Sample Collection
2.2. Environmental Parameters Measurements
2.3. Virus Reduction
2.4. Incubations for Measurements of Lytic Viral Production and Lysogeny Induction
2.5. Viral Production Rates
2.6. Flow Cytometry
2.7. Burst Size and the Percentage of Lysogenic Bacteria
2.8. Statistical Analysis
3. Results
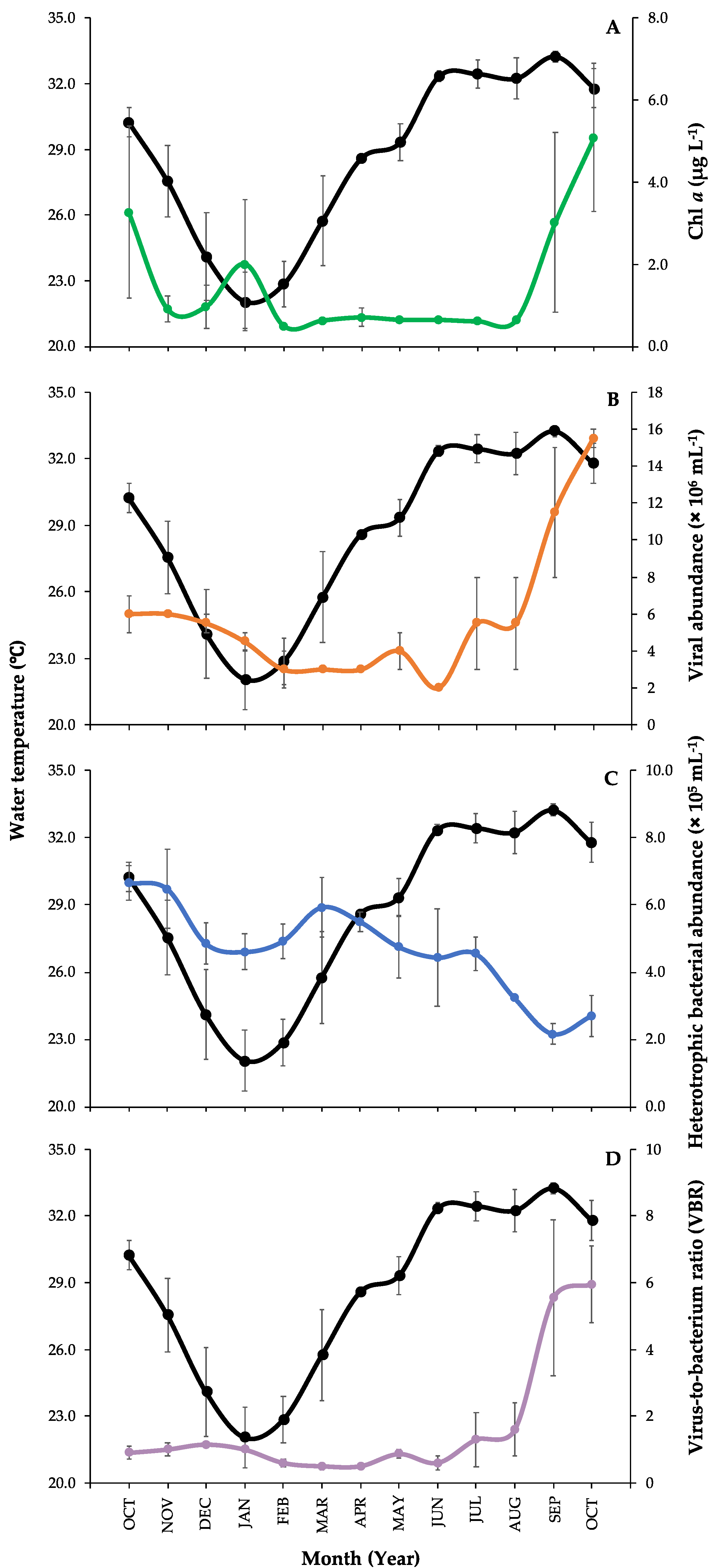
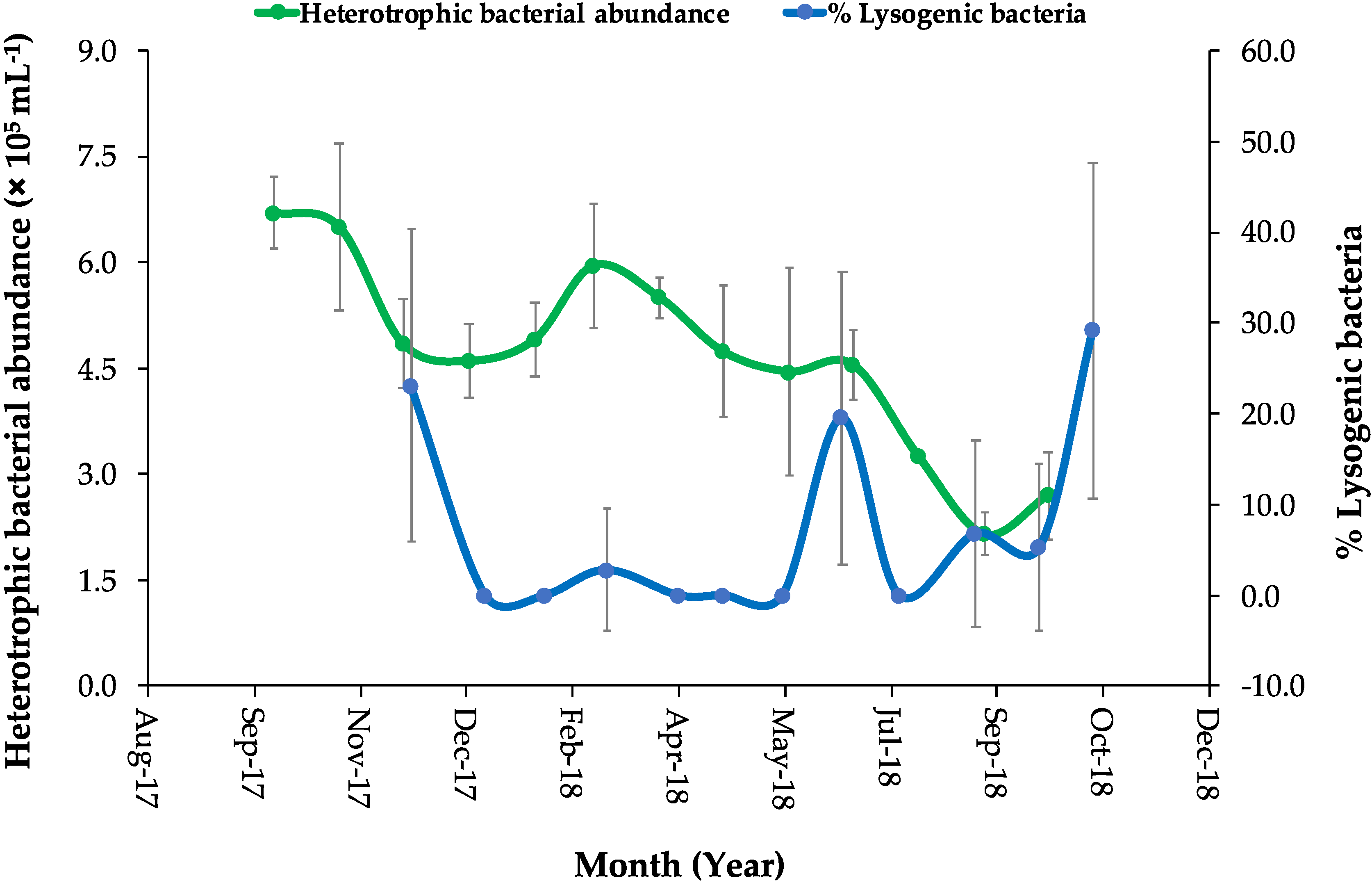
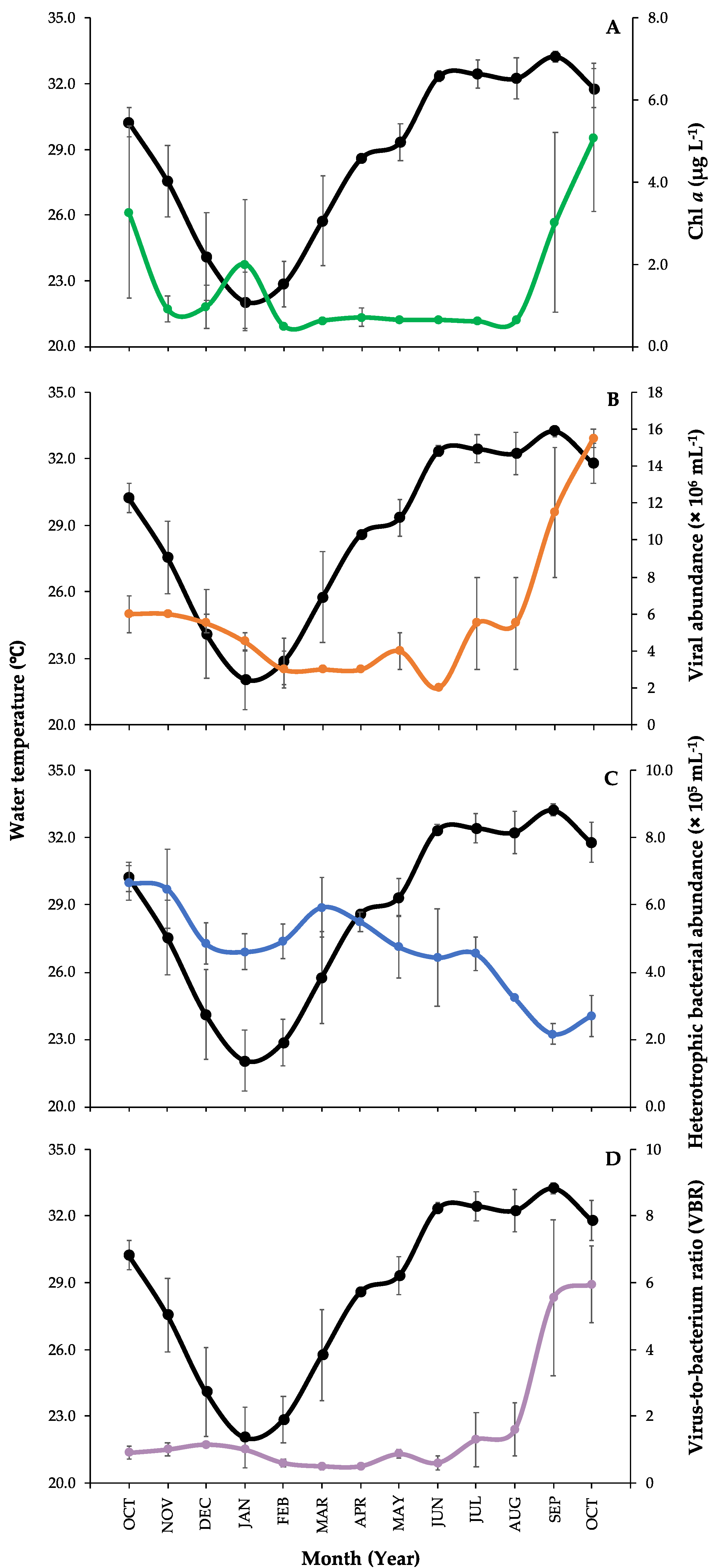
3.1. Environmental Parameters Measurements and Microbial Abundances
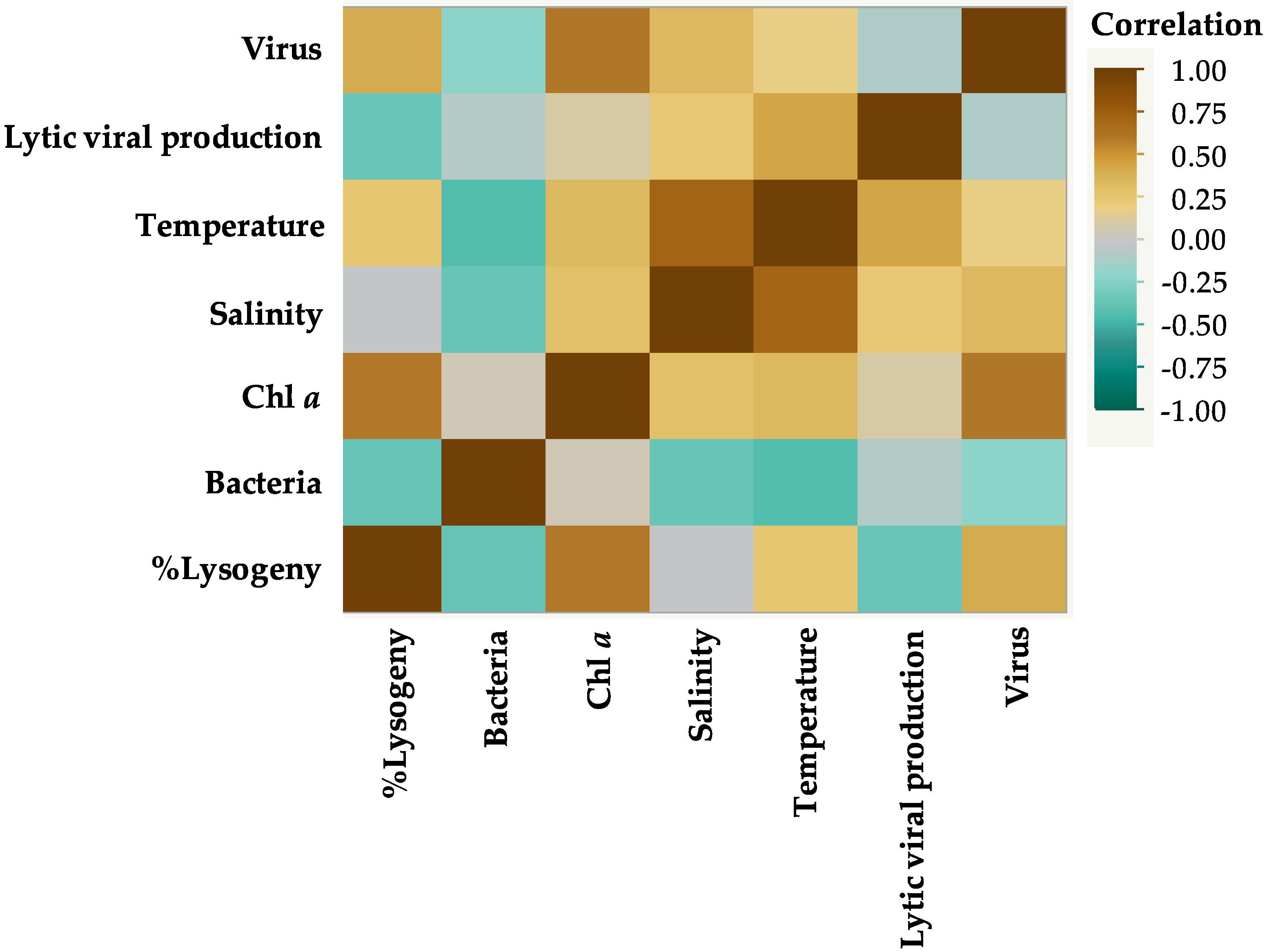
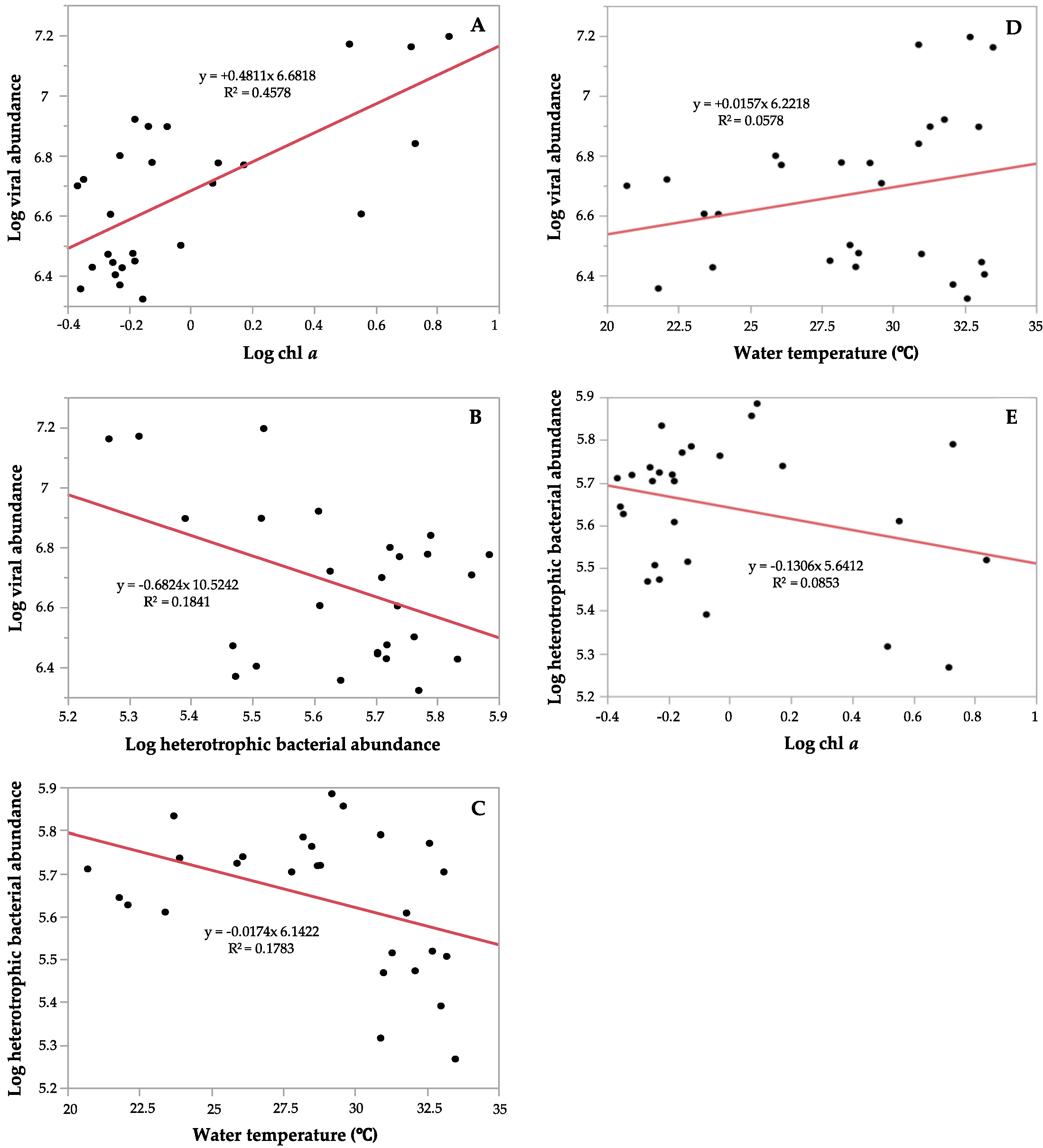
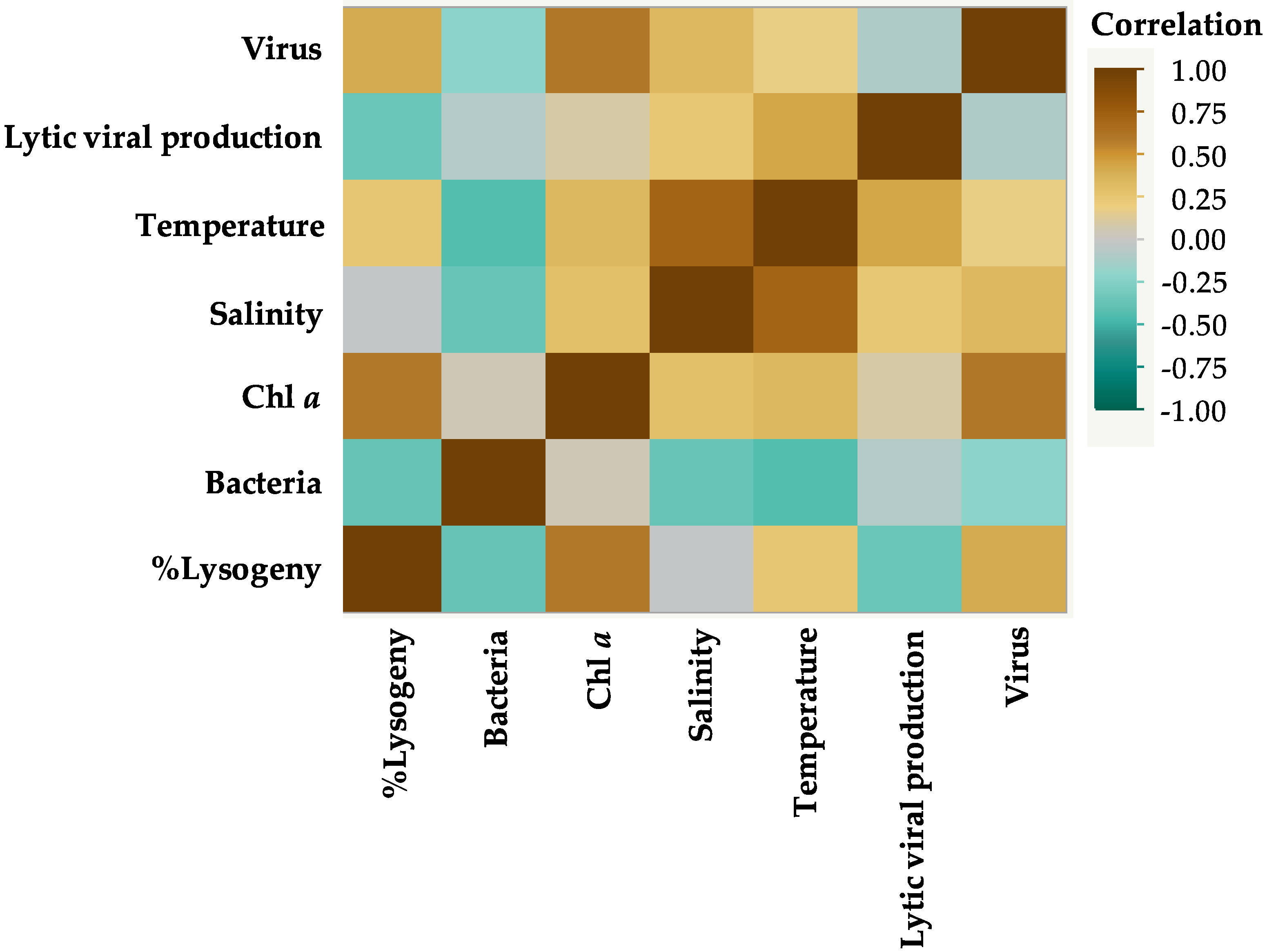
3.2. Correlations between Environmental Parameters and Microbial Communities
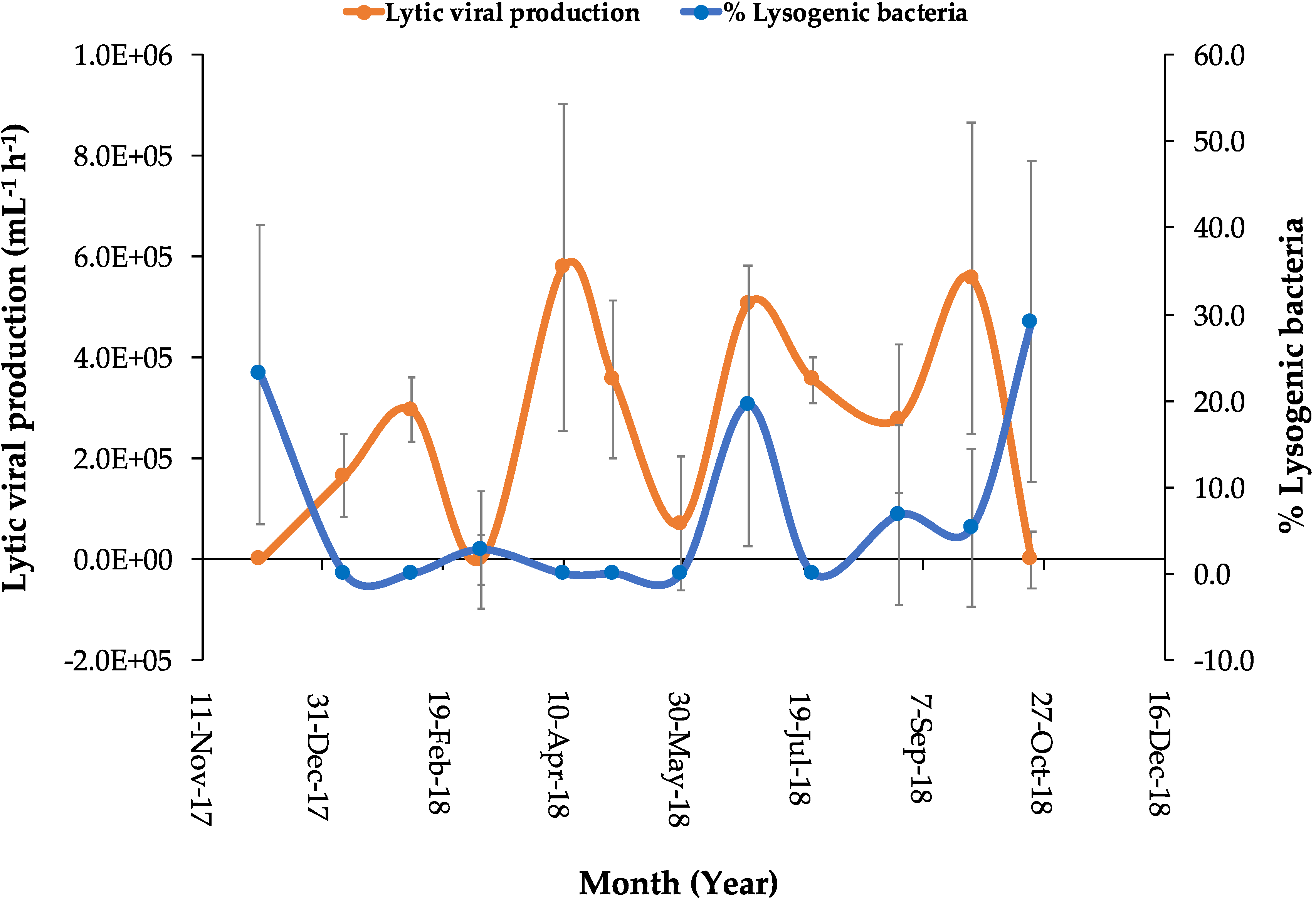
3.3. Viral Production and the Percentage of Lysogeny
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Breitbart, M. Marine viruses: Truth or dare. Annu. Rev. Mar. Sci. 2012, 4, 425–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suttle, C.A. The significance of viruses to mortality in aquatic microbial communities. Microb. Ecol. 1994, 28, 237–243. [Google Scholar] [CrossRef]
- Fuhrman, J.A. Marine viruses and their biogeochemical and ecological effects. Nature 1999, 399, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Wommack, K.E.; Colwell, R.R. Virioplankton: Viruses in aquatic ecosystems. Microbiol. Mol. Biol. Rev. 2000, 64, 69–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boras, J.A.; Montserrat Sala, M.; Baltar, F.; Arístegui, J.; Duarte, C.M.; Vaqué, D. Effect of viruses and protists on bacteria in eddies of the Canary Current region (subtropical northeast Atlantic). Limnol. Oceanogr. 2010, 55, 885–898. [Google Scholar] [CrossRef]
- Lara, E.; Vaqué, D.; Sà, E.L.; Boras, J.A.; Gomes, A.; Borrull, E.; Diez-Vives, C.; Teira, E.; Pernice, M.C.; Garcia, F.C.; et al. Unveiling the role and life strategies of viruses from the surface to the dark ocean. Sci. Adv. 2017, 3, e1602565. [Google Scholar] [CrossRef] [Green Version]
- Paul, J.H. Prophages in marine bacteria: Dangerous molecular time bombs or the key to survival in the seas? ISME J. 2008, 2, 579–589. [Google Scholar] [CrossRef]
- Howard-Varona, C.; Hargreaves, K.R.; Abedon, S.T.; Sullivan, M.B. Lysogeny in nature: Mechanisms, impact and ecology of temperate phages. ISME J. 2017, 11, 1511–1520. [Google Scholar] [CrossRef] [Green Version]
- Suttle, C.A.J.N. Viruses in the sea. Nature 2005, 437, 356–361. [Google Scholar] [CrossRef]
- Suttle, C.A.; Chen, F. Mechanisms and rates of decay of marine viruses in seawater. Appl. Environ. Microbiol. 1992, 58, 3721–3729. [Google Scholar] [CrossRef] [Green Version]
- Noble, R.T.; Fuhrman, J.A. Virus decay and its causes in coastal waters. Appl. Environ. Microbiol. 1997, 63, 77–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, H.; Sobsey, M.D. Comparative Survival of Indicator Viruses and Enteric Viruses in Seawater and Sediment. Water Sci. Technol. 1993, 27, 425–428. [Google Scholar] [CrossRef]
- Danovaro, R.; Corinaldesi, C.; Dell’anno, A.; Fuhrman, J.A.; Middelburg, J.J.; Noble, R.T.; Suttle, C.A. Marine viruses and global climate change. FEMS Microbiol. Rev. 2011, 35, 993–1034. [Google Scholar] [CrossRef]
- Williamson, S.J.; Houchin, L.A.; McDaniel, L.; Paul, J.H. Seasonal variation in lysogeny as depicted by prophage induction in Tampa Bay, Florida. Appl Environ. Microbiol. 2002, 68, 4307–4314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDaniel, L.D.; delaRosa, M.; Paul, J.H. Temperate and lytic cyanophages from the Gulf of Mexico. J. Mar. Biol. Assoc. UK 2006, 86, 517–527. [Google Scholar] [CrossRef]
- Stewart, F.M.; Levin, B.R. The Population Biology of Bacterial-Viruses—Why Be Temperate. Theor. Popul. Biol. 1984, 26, 93–117. [Google Scholar] [CrossRef]
- Jiang, S.C.; Paul, J.H. Occurrence of lysogenic bacteria in marine microbial communities as determined by prophage induction. Mar. Ecol. Prog. Ser. 1996, 142, 27–38. [Google Scholar] [CrossRef]
- Knowles, B.; Silveira, C.; Bailey, B.; Barott, K.; Cantu, V.; Cobián-Güemes, A.; Coutinho, F.; Dinsdale, E.; Felts, B.; Furby, K.; et al. Lytic to temperate switching of viral communities. Nature 2016, 531, 466–470. [Google Scholar] [CrossRef]
- Ashy, R.A. Lysogeny and Phage Dynamics in the Red Sea Ecosystem. Ph.D. Thesis, King Abdullah University of Science and Technology, Thuwal, Saudi Arabia, 2 December 2019. [Google Scholar]
- Boras, J.A.; Sala, M.M.; Vazquez-Dominguez, E.; Weinbauer, M.G.; Vaque, D. Annual changes of bacterial mortality due to viruses and protists in an oligotrophic coastal environment (NW Mediterranean). Environ. Microbiol. 2009, 11, 1181–1193. [Google Scholar] [CrossRef]
- Mojica, K.D.; Brussaard, C.P. Factors affecting virus dynamics and microbial host-virus interactions in marine environments. FEMS Microbiol. Ecol. 2014, 89, 495–515. [Google Scholar] [CrossRef] [Green Version]
- Nagasaki, K.; Yamaguchi, M. Effect of temperature on the algicidal activity and the stability of HaV (Heterosigma akashiwo virus). Aquat. Microb. Ecol. 1998, 15, 211–216. [Google Scholar] [CrossRef] [Green Version]
- Baudoux, A.-C.; Brussaard, C.P. Characterization of different viruses infecting the marine harmful algal bloom species Phaeocystis globosa. Virology 2005, 341, 80–90. [Google Scholar] [CrossRef] [Green Version]
- Tomaru, Y.; Tanabe, H.; Yamanaka, S.; Nagasaki, K. Effects of temperature and light on stability of microalgal viruses, HaV, HcV and HcRNAV. Plankton Biol. Ecol. 2005, 52, 1–6. [Google Scholar]
- Martínez, J.M.; Boere, A.; Gilg, I.; van Lent, J.W.; Witte, H.J.; van Bleijswijk, J.D.; Brussaard, C.P. New lipid envelope-containing dsDNA virus isolates infecting Micromonas pusilla reveal a separate phylogenetic group. Aquat. Microb. Ecol. 2015, 74, 17–28. [Google Scholar] [CrossRef]
- Mojica, K.D.; Huisman, J.; Wilhelm, S.W.; Brussaard, C.P. Latitudinal variation in virus-induced mortality of phytoplankton across the North Atlantic Ocean. ISME J. 2016, 10, 500–513. [Google Scholar] [CrossRef] [Green Version]
- McDaniel, L.; Houchin, L.A.; Williamson, S.J.; Paul, J.H. Lysogeny in marine Synechococcus. Nature 2002, 415, 496. [Google Scholar] [CrossRef] [PubMed]
- Weinbauer, M.G.; Brettar, I.; Höfle, M.G. Lysogeny and virus-induced mortality of bacterioplankton in surface, deep, and anoxic marine waters. Limnol. Oceanogr. 2003, 48, 1457–1465. [Google Scholar] [CrossRef] [Green Version]
- Demory, D.; Arsenieff, L.; Simon, N.; Six, C.; Rigaut-Jalabert, F.; Marie, D.; Ge, P.; Bigeard, E.; Jacquet, S.; Sciandra, A.; et al. Temperature is a key factor in Micromonas-virus interactions. ISME J. 2017, 11, 601–612. [Google Scholar] [CrossRef] [Green Version]
- Kendrick, B.J.; DiTullio, G.R.; Cyronak, T.J.; Fulton, J.M.; Van Mooy, B.A.; Bidle, K.D. Temperature-induced viral resistance in Emiliania huxleyi (Prymnesiophyceae). PLoS ONE 2014, 9, e112134. [Google Scholar] [CrossRef]
- Tomaru, Y.; Kimura, K.; Yamaguchi, H. Temperature alters algicidal activity of DNA and RNA viruses infecting Chaetoceros tenuissimus. Aquat. Microb. Ecol. 2014, 73, 171–183. [Google Scholar] [CrossRef] [Green Version]
- Wilson, W.H.; Francis, I.; Ryan, K.; Davy, S.K. Temperature induction of viruses in symbiotic dinoflagellates. Aquat. Microb. Ecol. 2001, 25, 99–102. [Google Scholar] [CrossRef] [Green Version]
- Tsai, A.Y.; Gong, G.C.; Liu, H.B. Seasonal variations in virioplankton and picoplankton in semi-enclosed and open coastal waters. Terr. Atoms. Ocean. Sci. 2018, 29, 465–472. [Google Scholar] [CrossRef] [Green Version]
- Wells, L.E.; Deming, J.W. Effects of temperature, salinity and clay particles on inactivation and decay of cold-active marine Bacteriophage 9A. Aquat. Microb. Ecol. 2006, 45, 31–39. [Google Scholar] [CrossRef]
- Rasul, N.M.; Stewart, I.C.; Nawab, Z.A. Introduction to the Red Sea: Its origin, structure, and environment. In The Red Sea; Springer: Berlin, Germany, 2015; pp. 1–28. [Google Scholar]
- Chaidez, V.; Dreano, D.; Agusti, S.; Duarte, C.M.; Hoteit, I. Decadal trends in Red Sea maximum surface temperature. Sci. Rep. 2017, 7, 8144. [Google Scholar] [CrossRef] [Green Version]
- Acker, J.; Leptoukh, G.; Shen, S.; Zhu, T.; Kempler, S. Remotely-sensed chlorophyll a observations of the northern Red Sea indicate seasonal variability and influence of coastal reefs. J. Mar. Syst. 2008, 69, 191–204. [Google Scholar] [CrossRef]
- Banguera-Hinestroza, E.; Eikrem, W.; Mansour, H.; Solberg, I.; Curdia, J.; Holtermann, K.; Edvardsen, B.; Kaartvedt, S. Seasonality and toxin production of Pyrodinium bahamense in a Red Sea lagoon. Harmful Algae 2016, 55, 163–171. [Google Scholar] [CrossRef]
- Sofianos, S.S.; Johns, W.E.; Murray, S.P. Heat and freshwater budgets in the Red Sea from direct observations at Bab el Mandeb. Deep Sea Res. Part II Top. Stud. Oceanogr. 2002, 49, 1323–1340. [Google Scholar] [CrossRef]
- Weisse, T. The Microbial Loop in the Red-Sea. Dynamics of Pelagic Bacteria and Heterotrophic Nanoflagellates. Mar. Ecol. Prog. Ser. 1989, 55, 241–250. [Google Scholar] [CrossRef]
- Pearman, J.K.; Ellis, J.; Irigoien, X.; Sarma, Y.V.B.; Jones, B.H.; Carvalho, S. Microbial planktonic communities in the Red Sea: High levels of spatial and temporal variability shaped by nutrient availability and turbulence. Sci. Rep. 2017, 7, 6611. [Google Scholar] [CrossRef] [Green Version]
- Silva, L.; Calleja, M.L.; Huete-Stauffer, T.M.; Ivetic, S.; Ansari, M.I.; Viegas, M.; Moran, X.A.G. Low Abundances but High Growth Rates of Coastal Heterotrophic Bacteria in the Red Sea. Front. Microbiol. 2018, 9, 3244. [Google Scholar] [CrossRef] [Green Version]
- Hadas, E.; Marie, D.; Shpigel, M.; Ilan, M. Virus predation by sponges is a new nutrient-flow pathway in coral reef food webs. Limnol. Oceanogr. 2006, 51, 1548–1550. [Google Scholar] [CrossRef] [Green Version]
- Danovaro, R.; Bongiorni, L.; Corinaldesi, C.; Giovannelli, D.; Damiani, E.; Astolfi, P.; Greci, L.; Pusceddu, A. Sunscreens cause coral bleaching by promoting viral infections. Environ. Health Perspect. 2008, 116, 441–447. [Google Scholar] [CrossRef] [Green Version]
- Antunes, A.; Alam, I.; Simoes, M.F.; Daniels, C.; Ferreira, A.J.S.; Siam, R.; El-Dorry, H.; Bajic, V.B. First Insights into the Viral Communities of the Deep-sea Anoxic Brines of the Red Sea. Genom. Proteom. Bioinform. 2015, 13, 304–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prabowo, D.A.; Agustí, S. Free-living dinoflagellates of the central Red Sea, Saudi Arabia: Variability, new records and potentially harmful species. Mar. Pollut. Bull. 2019, 141, 629–648. [Google Scholar] [CrossRef]
- Wilhelm, S.W.; Brigden, S.M.; Suttle, C.A. A dilution technique for the direct measurement of viral production: A comparison in stratified and tidally mixed coastal waters. Microb. Ecol. 2002, 43, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Weinbauer, M.G.; Rowe, J.M.; Wilhelm, S. Determining rates of virus production in aquatic systems by the virus reduction approach. In Manual of Aquatic Viral Ecology; ASLO: Waco, TX, USA, 2010; pp. 1–8. [Google Scholar] [CrossRef]
- Rowe, J.M.; DeBruyn, J.M.; Poorvin, L.; LeCleir, G.R.; Johnson, Z.I.; Zinser, E.R.; Wilhelm, S.W. Viral and bacterial abundance and production in the Western Pacific Ocean and the relation to other oceanic realms. FEMS Microbiol. Ecol. 2012, 79, 359–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinbauer, M.G.; Suttle, C.A. Potential significance of lysogeny to bacteriophage production and bacterial mortality in coastal waters of the gulf of Mexico. Appl Environ. Microbiol. 1996, 62, 4374–4380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinbauer, M.G.; Suttle, C.A. Lysogeny and prophage induction in coastal and offshore bacterial communities. Aquat. Microb. Ecol. 1999, 18, 217–225. [Google Scholar] [CrossRef] [Green Version]
- Weinbauer, M.G. Ecology of prokaryotic viruses. FEMS Microbiol. Rev. 2004, 28, 127–181. [Google Scholar] [CrossRef] [Green Version]
- Agustí, S.; Sánchez, M.C. Cell viability in natural phytoplankton communities quantified by a membrane permeability probe. Limnol. Oceanogr. 2002, 47, 818–828. [Google Scholar] [CrossRef] [Green Version]
- Lasternas, S.; Agustí, S. The percentage of living bacterial cells related to organic carbon release from senescent oceanic phytoplankton. Biogeosciences 2014, 11, 6377–6387. [Google Scholar] [CrossRef] [Green Version]
- Brum, J.R.; Hurwitz, B.L.; Schofield, O.; Ducklow, H.W.; Sullivan, M.B. Seasonal time bombs: Dominant temperate viruses affect Southern Ocean microbial dynamics. ISME J. 2016, 10, 437–449. [Google Scholar] [CrossRef] [PubMed]
- Cochran, P.K.; Kellogg, C.A.; Paul, J.H. Prophage induction of indigenous marine lysogenic bacteria by environmental pollutants. Mar. Ecol. Prog. Ser. 1998, 164, 125–133. [Google Scholar] [CrossRef]
- Cochran, P.K.; Paul, J.H. Seasonal abundance of lysogenic bacteria in a subtropical estuary. Appl. Environ. Microbiol. 1998, 64, 2308–2312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magiopoulos, I.; Pitta, P. Viruses in a deep oligotrophic sea: Seasonal distribution of marine viruses in the epi-, meso- and bathypelagic waters of the Eastern Mediterranean. Deep Sea Res. Part II Top. Stud. Oceanogr. 2012, 66, 1–10. [Google Scholar] [CrossRef]
- Weinbauer, M.G.; Winter, C.; Höfle, M.G. Reconsidering transmission electron microscopy based estimates of viral infection of bacterio-plankton using conversion factors derived from natural communities. Aquat. Microb. Ecol. 2002, 27, 103–110. [Google Scholar] [CrossRef] [Green Version]
- Gasol, J.M.; Morán, X.A.G. Flow cytometric determination of microbial abundances and its use to obtain indices of community structure and relative activity. In Hydrocarbon and Lipid Microbiology Protocols; Springer: Berlin, Germany, 2015; pp. 159–187. [Google Scholar]
- Brussaard, C.P.; Payet, J.P.; Winter, C.; Weinbauer, M.G. Quantification of aquatic viruses by flow cytometry. In Manual of Aquatic Viral Ecology; ASLO: Waco, TX, USA, 2010; Volume 11, pp. 102–107. [Google Scholar]
- Smith, R.J.; Paterson, J.S.; Sibley, C.A.; Hutson, J.L.; Mitchell, J.G. Putative Effect of Aquifer Recharge on the Abundance and Taxonomic Composition of Endemic Microbial Communities. PLoS ONE 2015, 10, e0129004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wells, L.E.; Deming, J.W. Significance of bacterivory and viral lysis in bottom waters of Franklin Bay, Canadian Arctic, during winter. Aquat. Microb. Ecol. 2006, 43, 209–221. [Google Scholar] [CrossRef]
- Jiang, S.C.; Paul, J.H. Seasonal and Diel Abundance of Viruses and Occurrence of Lysogeny/Bacteriocinogeny in the Marine-Environment. Mar. Ecol. Prog. Ser. 1994, 104, 163–172. [Google Scholar] [CrossRef]
- Ordulj, M.; Krstulovic, N.; Santic, D.; Jozic, S.; Solic, M. Distribution of marine viruses in the Central and South Adriatic Sea. Mediterr. Mar. Sci. 2015, 16, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Yucel-Gier, G.; Kacar, A.; Gonul, L.T.; Pazi, I.; Kucuksezgin, F.; Erarslanoglu, N.; Toker, S.K. Evaluation of the relationship of picoplankton and viruses to environmental variables in a lagoon system (Cakalburnu Lagoon, Turkey). Chem. Ecol. 2018, 34, 211–228. [Google Scholar] [CrossRef]
- Tapper, M.A.; Hicks, R.E. Temperate viruses and lysogeny in Lake Superior bacterioplankton. Limnol. Oceanogr. 1998, 43, 95–103. [Google Scholar] [CrossRef]
- Ortmann, A.C.; Lawrence, J.E.; Suttle, C.A. Lysogeny and lytic viral production during a bloom of the cyanobacterium Synechococcus spp. Microb. Ecol. 2002, 43, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Long, A.; McDaniel, L.D.; Mobberley, J.; Paul, J.H. Comparison of lysogeny (prophage induction) in heterotrophic bacterial and Synechococcus populations in the Gulf of Mexico and Mississippi River plume. ISME J. 2008, 2, 132–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowe, J.M.; Saxton, M.A.; Cottrell, M.T.; DeBruyn, J.M.; Berg, G.M.; Kirchman, D.L.; Hutchins, D.A.; Wilhelm, S.W. Constraints on viral production in the Sargasso Sea and North Atlantic. Aquat. Microb. Ecol. 2008, 52, 233–244. [Google Scholar] [CrossRef]
- Parada, V.; Herndl, G.J.; Weinbauer, M.G. Viral burst size of heterotrophic prokaryotes in aquatic systems. J. Mar. Biol. Assoc. UK 2006, 86, 613–621. [Google Scholar] [CrossRef]
- Wilcox, R.M.; Fuhrman, J.A. Bacterial-Viruses in Coastal Seawater—Lytic Rather Than Lysogenic Production. Mar. Ecol. Prog. Ser. 1994, 114, 35–45. [Google Scholar] [CrossRef]
- Bongiorni, L.; Magagnini, M.; Armeni, M.; Noble, R.; Danovaro, R. Viral production, decay rates, and life strategies along a trophic gradient in the North Adriatic Sea. Appl. Environ. Microbiol. 2005, 71, 6644–6650. [Google Scholar] [CrossRef] [Green Version]
- Winget, D.M.; Helton, R.R.; Williamson, K.E.; Bench, S.R.; Williamson, S.J.; Wommack, K.E. Repeating patterns of virioplankton production within an estuarine ecosystem. Proc. Natl. Acad. Sci. USA 2011, 108, 11506–11511. [Google Scholar] [CrossRef] [Green Version]
- White, P.A.; Kalff, J.; Rasmussen, J.B.; Gasol, J.M. The effect of temperature and algal biomass on bacterial production and specific growth rate in freshwater and marine habitats. Microb. Ecol. 1991, 21, 99–118. [Google Scholar] [CrossRef]
- Wiebe, W.J.; Sheldon, W.M.; Pomeroy, L.R. Bacterial growth in the cold: Evidence for an enhanced substrate requirement. Appl. Environ. Microbiol. 1992, 58, 359–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinbauer, M.G.; Christaki, U.; Nedoma, J.; Simek, K. Comparing the effects of resource enrichment and grazing on viral production in a meso-eutrophic reservoir. Aquat. Microb. Ecol. 2003, 31, 137–144. [Google Scholar] [CrossRef]
- Jeffrey, W.H.; Kase, J.P.; Wilhelm, S.W. UV radiation effects on heterotrophic bacterioplankton and viruses in marine ecosystems. In The Effects of UV Radiation in the Marine Environment; Stephen De Mora, S.D., Maria Vernet, Eds.; Cambridge University Press: Cambridge, UK, 2000; Volume 10, pp. 206–236. [Google Scholar]
- Danovaro, R.; Dell’Anno, A.; Corinaldesi, C.; Rastelli, E.; Cavicchioli, R.; Krupovic, M.; Noble, R.T.; Nunoura, T.; Prangishvili, D. Virus-mediated archaeal hecatomb in the deep seafloor. Sci. Adv. 2016, 2, e1600492. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lytic Virus Production (VP) (mL−1 h−1) | Lysogeny (%) | Burst Size (BS) | VBR (Ratio) | ||||
|---|---|---|---|---|---|---|---|
| Mean ± SE | (MIN/ MAX) | Mean ± SE | (MIN/ MAX) | Mean ± SE | (MIN/MAX) | Mean ± SE | (MIN/ MAX) |
| 2.6 × 105 ± 6 × 104 | N/D–5.8 × 105 | 7.2 ± 2.9 | N/D–29.1 | 15 ± 5.3 | 2.4–44.9 | 16 ± 4 | 4–79 |
| Independent Variables and Viral Abundance | Heterotrophic Bacterial Abundance | ||
|---|---|---|---|
| Interactions/Units | Parameters Estimates ± SE | t-ratio | p-value |
| Intercept | 8.3505 ± 1.0653 | 7.84 | <0.0001 |
| Virus/log10 | −0.3900 ± 0.1742 | −2.24 | 0.0367 |
| Water temperature/ (°C) | −0.0015 ± 0.0099 | −0.15 | 0.8829 |
| Chl a/Log10 (µg L−1) | 0.3144 ± 0.1371 | 2.29 | 0.0328 |
| Virus × water temperature | −0.0298 ± 0.0397 | −0.75 | 0.4612 |
| Water temperature × Chl a | 0.0693 ± 0.0343 | 2.02 | 0.0573 |
| Chl a × Virus | −0.013019 ± 0.5258 | −2.48 | 0.0224 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdulrahman Ashy, R.; Agustí, S. Low Host Abundance and High Temperature Determine Switching from Lytic to Lysogenic Cycles in Planktonic Microbial Communities in a Tropical Sea (Red Sea). Viruses 2020, 12, 761. https://doi.org/10.3390/v12070761
Abdulrahman Ashy R, Agustí S. Low Host Abundance and High Temperature Determine Switching from Lytic to Lysogenic Cycles in Planktonic Microbial Communities in a Tropical Sea (Red Sea). Viruses. 2020; 12(7):761. https://doi.org/10.3390/v12070761
Chicago/Turabian StyleAbdulrahman Ashy, Ruba, and Susana Agustí. 2020. "Low Host Abundance and High Temperature Determine Switching from Lytic to Lysogenic Cycles in Planktonic Microbial Communities in a Tropical Sea (Red Sea)" Viruses 12, no. 7: 761. https://doi.org/10.3390/v12070761
APA StyleAbdulrahman Ashy, R., & Agustí, S. (2020). Low Host Abundance and High Temperature Determine Switching from Lytic to Lysogenic Cycles in Planktonic Microbial Communities in a Tropical Sea (Red Sea). Viruses, 12(7), 761. https://doi.org/10.3390/v12070761