Characterization and Incidence of the First Member of the Genus Mitovirus Identified in the Phytopathogenic Species Fusarium oxysporum


Abstract
1. Introduction
2. Materials and Methods
2.1. Fungal Isolates and Culture Conditions
2.2. dsRNA Extractions
2.3. cDNA Synthesis, Cloning, and Sequencing
2.4. Molecular and Phylogenetic Analysis
2.5. Incidence of Mitovirus FodMV1 in the Collection Analyzed
3. Results
3.1. Identification of Viral Infections in a Collection of F. oxysporum f. sp. dianthi Isolates
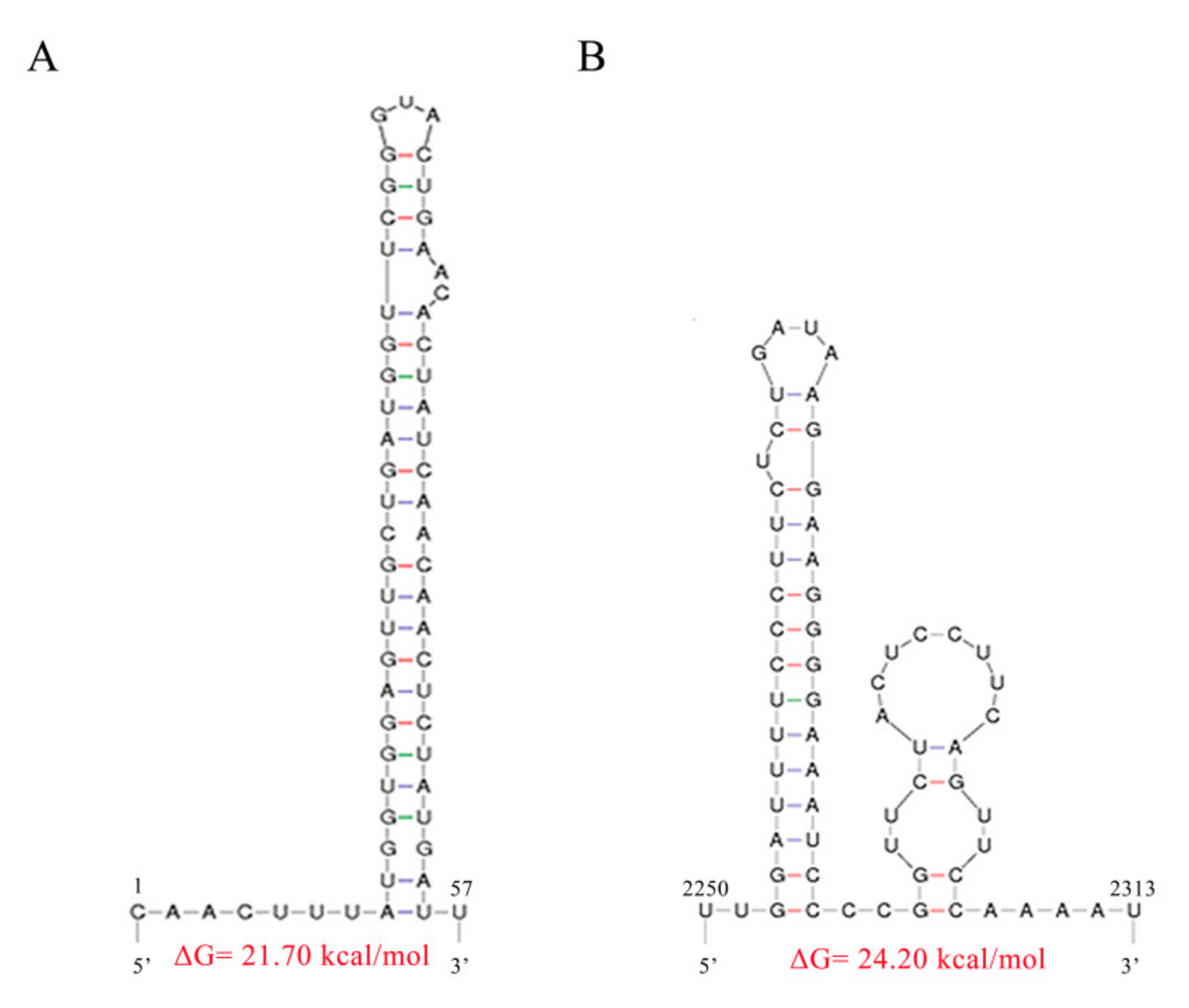
3.2. Molecular Characterization of a Novel Mitovirus Infecting F. oxysporum f.sp. dianthi
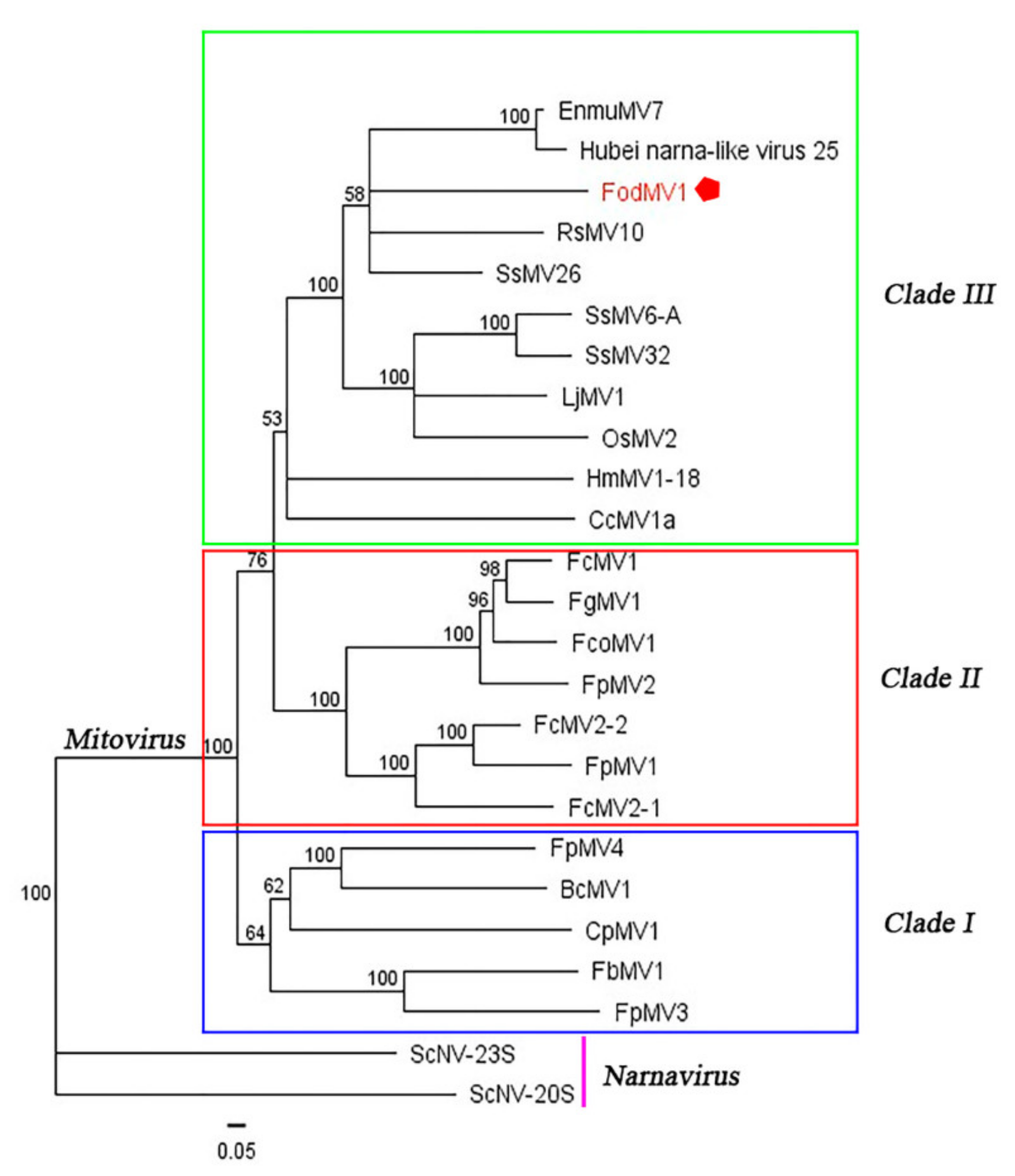
3.3. Phylogenetic Relationship between FodMV1 and Other Mitoviruses.
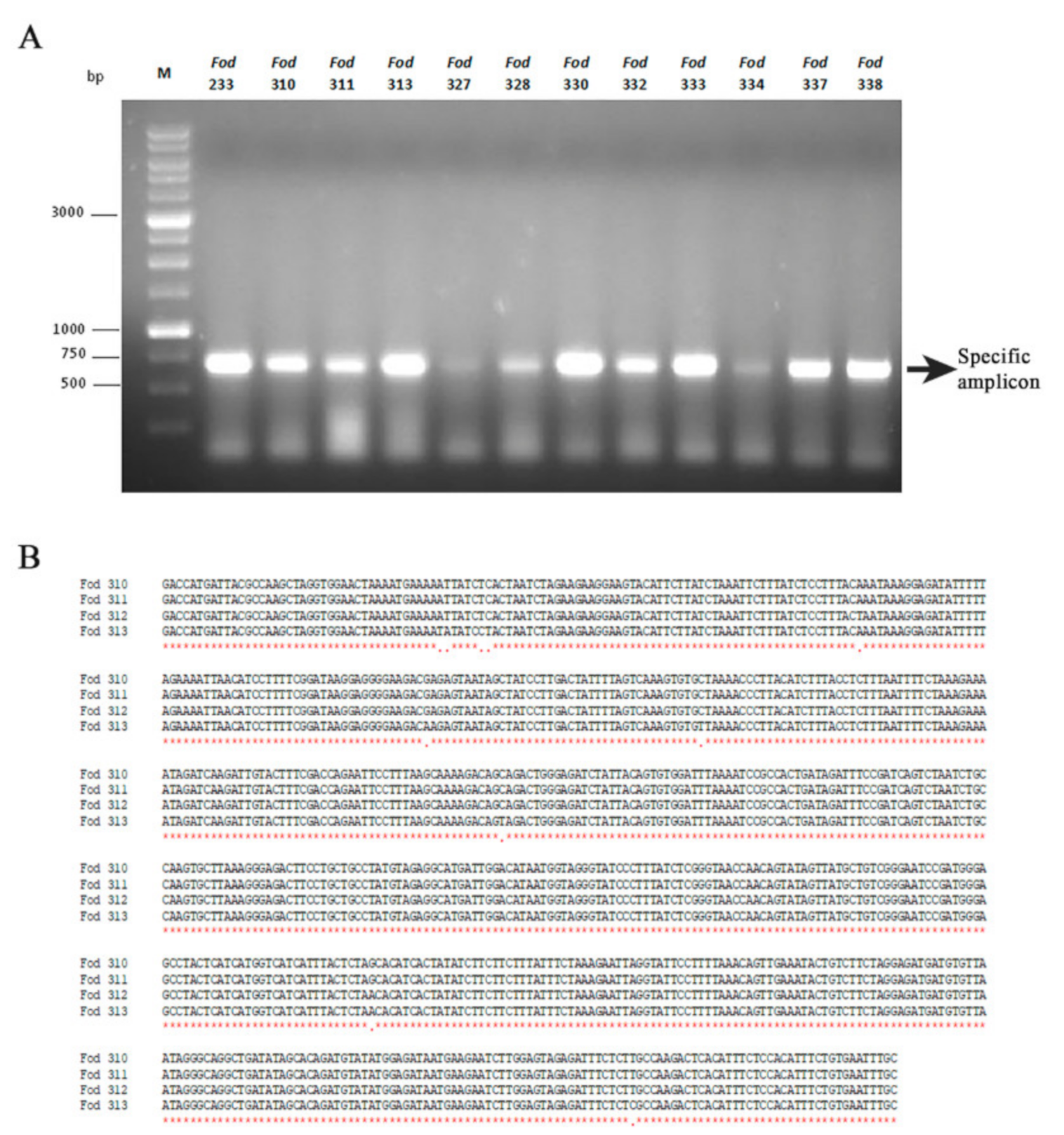
3.4. Incidence of Mitovirus FodMV1 in the Collection Analyzed
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Garibaldi, A.; Gullino, M.L. Attempts of biocontrol of fusarium-wilt of carnation in Italy. Phytopathology 1987, 77, 1721. [Google Scholar]
- Baayen, R.P. Fusarium wilt of carnation. Disease development, resistance mechanism of the host and taxonomy of the pathogen. Ph.D. Thesis, Degree-Granting University of Utretcht, Utrecht, The Netherlands, 1988. [Google Scholar]
- Ghabrial, S.A.; Suzuki, N. Viruses of plant pathogenic fungi. Annu Rev. Phytopathol 2009, 47, 353–384. [Google Scholar] [CrossRef] [PubMed]
- Pearson, M.N.; Beever, R.E.; Boine, B.; Arthur, K. Mycoviruses of filamentous fungi and their relevance to plant pathology. Mol Plant. Pathol 2009, 10, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Son, M.; Yu, J.; Kim, K.H. Five questions about mycoviruses. Plos Pathog 2015, 11, e1005172. [Google Scholar] [CrossRef] [PubMed]
- Ghabrial, S.A.; Caston, J.R.; Jiang, D.; Nibert, M.L.; Suzuki, N. 50-plus years of fungal viruses. Virology 2015, 356–368, 479–480. [Google Scholar] [CrossRef]
- Abbas, A. A Review Paper on Mycoviruses. J. Plant. Pathol. Microbiol. 2016, 7, 2. [Google Scholar] [CrossRef]
- Nibert, M.L.; Debat, H.J.; Manny, A.R.; Grigoriev, I.V.; De Fine Licht, H.H. Mitovirus and mitochondrial coding sequences from basal fungus Entomophthora muscae. Viruses 2019, 11, 351. [Google Scholar] [CrossRef]
- King, A.M.; Adams, M.J.; Lefkowitz, E.J. Virus taxonomy: Classification and nomenclature of viruses: Ninth Report of the International Committee on Taxonomy of Viruses; Elsevier: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Herrero, N.; Dueñas, E.; Quesada-Moraga, E.; Zabalgogeazcoa, I. Prevalence and diversity of viruses in the entomopathogenic fungus Beauveria bassiana. Appl Environ. Microbiol. 2012, 78, 8523–8530. [Google Scholar] [CrossRef]
- Kotta-Loizou, I.; Coutts, R.H.A. Studies on the virome of the entomopathogenic fungus Beauveria bassiana reveal novel dsRNA elements and mild hypervirulence. PLoS Pathog. 2017, 13, e1006183. [Google Scholar] [CrossRef]
- Ikeda, Y.; Shimura, H.; Kitahara, R.; Masuta, C.; Ezawa, T. A novel virus-like double-stranded RNA in an obligate biotroph arbuscular mycorrhizal fungus: A hidden player in mycorrhizal symbiosis. Mol. Plant. Microbe Interact. 2012, 25, 1005–1012. [Google Scholar] [CrossRef]
- Cook, S.; Chung, B.Y.; Bass, D.; Moureau, G.; Tang, S.; McAlister, E.; Culverwell, C.L.; Glücksman, E.; Wang, H.; Brown, T.D.; et al. Novel virus discovery and genome reconstruction from field RNA samples reveals highly divergent viruses in dipteran hosts. PLoS ONE 2013, 8, e80720. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Lin, X.D.; Tian, J.H.; Chen, L.J.; Chen, X.; Li, C.X.; Qin, X.C.; Li, J.; Cao, J.P.; Eden, J.S.; et al. Redefining the invertebrate RNA virosphere. Nature 2016, 540, 539–543. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Jiang, D. New insights into mycoviruses and exploration for the biological control of crop fungal diseases. Annu Rev. Phytopathol. 2014, 52, 45–68. [Google Scholar] [CrossRef] [PubMed]
- Lemus-Minor, C.G.; Cañizares, M.C.; García-Pedrajas, M.D.; Pérez-Artés, E. Fusarium oxysporum f. sp. dianthi virus 1 accumulation is correlated with changes in virulence and other phenotypic traits of its fungal host. Phytopathology 2018, 108, 957–963. [Google Scholar] [CrossRef]
- Yu, X.; Li, B.; Fu, Y.; Xie, J.; Cheng, J.; Ghabrial, S.A.; Li, G.; Yi, X.; Jiang, D. Extracellular transmission of a DNA mycovirus and its use as a natural fungicide. Proc. Natl. Acad. Sci.USA 2013, 110, 1452–1457. [Google Scholar] [CrossRef]
- Gilbert, K.B.; Holcomb, E.E.; Allscheid, R.L.; Carrington, J.C. Hiding in plain sight: New virus genomes discovered via a systematic analysis of fungal public transcriptomes. PLoS ONE 2019, 14, e0219207. [Google Scholar] [CrossRef]
- Hillman., B.I.; Esteban., R. Narnaviridae. In Virus Taxonomy: Classification and nomenclature of viruses. Ninth report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier: San Diego, CA, USA, 2011; pp. 1055–1060. [Google Scholar]
- Hillman, B.I.; Cai, G. The family Narnaviridae: Simplest of RNA viruses. Adv. Virus Res. 2013, 86, 149–176. [Google Scholar] [CrossRef]
- Nibert, M.L.; Vong, M.; Fugate, K.K.; Debat, H.J. Evidence for contemporary plant mitoviruses. Virology 2018, 518, 14–24. [Google Scholar] [CrossRef]
- Nerva, L.; Vigani, G.; Di Silvestre, D.; Ciuffo, M.; Forgia, M.; Chitarra, W.; Turina, M. Biological and molecular characterization of Chenopodium quinoa mitovirus 1 reveals a distinct sRNA response compared to cytoplasmic RNA viruses. J. Virol. 2019, 93, e01998-e18. [Google Scholar] [CrossRef]
- Wu, M.; Zhang, L.; Li, G.; Jiang, D.; Ghabrial, S.A. Genome characterization of a debilitation-associated mitovirus infecting the phytopathogenic fungus Botrytis cinerea. Virology 2010, 406, 117–126. [Google Scholar] [CrossRef]
- Xu, Z.; Wu, S.; Liu, L.; Cheng, J.; Fu, Y.; Jiang, D.; Xie, J. A mitovirus related to plant mitochondrial gene confers hypovirulence on the phytopathogenic fungus Sclerotinia sclerotiorum. Virus Res. 2015, 197, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Dover, S.L.; Cole, T.E.; Brasier, C.M.; Buck, K.W. Multiple mitochondrial viruses in an isolate of the Dutch elm disease fungus Ophiostoma novo-ulmi. Virology 1999, 258, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Nibert, M.L. Mitovirus UGA(Trp) codon usage parallels that of host mitochondria. Virology 2017, 507, 96–100. [Google Scholar] [CrossRef] [PubMed]
- Paquin, B.; Laforest, M.J.; Forget, L.; Roewer, I.; Wang, Z.; Longcore, J.; Lang, B.F. The fungal mitochondrial genome project: Evolution of fungal mitochondrial genomes and their gene expression. Curr. Genet. 1997, 31, 380–395. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Xie, J.; Cheng, J.; Li, B.; Chen, T.; Fu, Y.; Li, G.; Wang, M.; Jin, H.; Wan, H.; et al. Fungal DNA virus infects a mycophagous insect and utilizes it as a transmission vector. Proc. Natl. Acad. Sci. USA 2016, 113, 12803–12808. [Google Scholar] [CrossRef] [PubMed]
- Marzano, S.-Y.L.; Nelson, B.D.; Ajayi-Oyetunde, O.; Bradley, C.A.; Hughes, T.J.; Hartman, G.L.; Eastburn, D.M.; Domier, L.L. Identification of diverse mycoviruses through metatranscriptomics characterization of the viromes of five major fungal plant pathogens. J. Virol. 2016, 90, 6846–6863. [Google Scholar] [CrossRef] [PubMed]
- Polashock, J.J.; Bedker, P.J.; Hillman, B.I. Movement of a small mitochondrial double-stranded RNA element of Cryphonectria parasitica: Ascospore inheritance and implications for mitochondrial recombination. Mol. Gen. Genet. 1997, 256, 566–571. [Google Scholar] [CrossRef]
- Xie, J.; Ghabrial, S.A. Molecular characterization of two mitoviruses co-infecting a hypovirulent isolate of the plant pathogenic fungus Sclerotinia sclerotiorum. Virology 2012, 428, 77–85. [Google Scholar] [CrossRef]
- Deng, F.; Xu, R.; Boland, G.J. Hypovirulence-associated double-stranded RNA from Sclerotinia homoeocarpa is conspecific with Ophiostoma novo-ulmi mitovirus 3a-Ld. Phytopathology 2003, 93, 1407–1414. [Google Scholar] [CrossRef]
- Lakshman, D.K.; Jian, J.; Tavantzis, S.M. A double-stranded RNA element from a hypovirulent strain of Rhizoctonia solani occurs in DNA form and is genetically related to the pentafunctional AROM protein of the shikimate pathway. Proc. Natl. Acad. Sci. USA 1998, 95, 6425–6429. [Google Scholar] [CrossRef]
- Vainio, E.J.; Martínez-Álvarez, P.; Bezos, D.; Hantula, J.; Diez, J.J. Fusarium circinatum isolates from northern Spain are commonly infected by three distinct mitoviruses. Arch. Virol. 2015, 160, 2093–2098. [Google Scholar] [CrossRef] [PubMed]
- Osaki, H.; Sasaki, A.; Nomiyama, K.; Sekiguchi, H.; Tomioka, K.; Takehara, T. Isolation and characterization of two mitoviruses and a putative alphapartitivirus from Fusarium spp. Virus Genes 2015, 50, 466–473. [Google Scholar] [CrossRef] [PubMed]
- Osaki, H.; Sasaki, A.; Nomiyama, K.; Tomioka, K. Multiple virus infection in a single strain of Fusarium poae shown by deep sequencing. Virus Genes 2016, 52, 835–847. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, Y.; Abraham, A.; Uesaka, K.; Kondo, H.; Suga, H.; Suzuki, N.; Chiba, S. Novel mitoviruses and a unique tymo-like virus in hypovirulent and virulent strains of the fusarium head blight bungus, Fusarium boothii. Viruses 2018, 10, 584. [Google Scholar] [CrossRef]
- Gómez-Lama Cabanás, C.; Valverde-Corredor, A.; Pérez-Artés, E. Molecular analysis of Spanish populations of Fusarium oxysporum f. sp dianthi demonstrates a high genetic diversity and identifies virulence groups in races 1 and 2 of the pathogen. Eur. J. Plant. Pathol 2012, 132, 561–576. [Google Scholar] [CrossRef]
- Valverde, R.A.; Nameth, S.T.; Jordan, R.L. Analysis of double-stranded-RNA for plant-virus diagnosis. Plant. Dis. 1990, 1990 74, 255–258. [Google Scholar]
- Xie, J.; Wei, D.; Jiang, D.; Fu, Y.; Li, G.; Ghabrial, S.; Peng, Y. Characterization of debilitation-associated mycovirus infecting the plant-pathogenic fungus Sclerotinia sclerotiorum. J. Gen. Virol. 2006, 87, 241–249. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Zuker, M.; Mathews, D.H.; Turner, D.H. Algorithms and Thermodynamics for RNA Secondary Structure Prediction: A Practical Guide. In RNA Biochemistry and Biotechnology; Barciszewski, J., Clark, B.F.C., Eds.; NATO ASI Series; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1999; pp. 11–43. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Hong, Y.; Cole, T.E.; Brasier, C.M.; Buck, K.W. Evolutionary relationships among putative RNA-dependent RNA polymerases encoded by a mitochondrial viruslike RNA in the dutch elm disease fungus, Ophiostoma novo-ulmi, by other viruses and virus-like RNAs and by the arabidopsis mitochondrial genome. Virology 1998, 246, 158–169. [Google Scholar] [CrossRef]
- Buck, K.W. Comparison of the replication of positive-stranded RNA viruses of plants and animals. Adv. Virus Res. 1996, 47, 159–251. [Google Scholar] [PubMed]
- Buck, K.W.; Brasier, C.M. Viruses of the dutch elm disease fungus. In dsRNA Genetic Elements: Concepts and Applications in Agriculture, Forestry and Medicine; Tavantzis, S.M., Ed.; CRC Press LCC: Boca Raton, FL, USA, 2002; pp. 165–190. [Google Scholar]
- Gómez-Lama Cabanás, C.; Pérez-Artés, E. New evidence of intra-race diversity in Fusarium oxysporum f. sp. dianthi populations based on Vegetative Compatibility Groups. Eur. J. Plant. Pathol. 2014, 139, 445–451. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate (s)a | Geographic Origin b | Source | Year | Race-Group Assignation by PCR Pattern c |
|---|---|---|---|---|
| Fod 108, 109, 110, 111, 120, 121 | Chipiona(Ca) | Plant | 2008 | R1t |
| Fod 112, 113, 114, 117 | Chipiona(Ca) | Plant | 2008 | R2I |
| Fod 118, 119 | Chipiona(Ca) | Plant | 2008 | R2II |
| Fod 115 | Chipiona(Ca) | Plant | 2008 | - |
| Fod 124, 127, 132, 134, 136, 138 | Chipiona(Ca) | Plant | 2009 | R2I |
| Fod 122, 128, 130, 140, 142 | Chipiona(Ca) | Plant | 2009 | R1t |
| Fod 183, 185, 187 | Chipiona(Ca) | Plant | 2010 | R2I |
| Fod 181, 189, 191 | Chipiona(Ca) | Plant | 2010 | R1t |
| Fod 179, 195, | Chipiona(Ca) | Soil | 2010 | R2II |
| Fod 197 | Chipiona(Ca) | Soil | 2010 | R1t |
| Fod 144, 146, 148, 150, 158, 160, 162, 164 | La Colonia(Se) | Plant | 2010 | R2I |
| Fod 152, 154 | Lebrija(Se) | Plant | 2010 | R1t |
| Fod 156 | Lebrija(Se) | Plant | 2010 | R2II |
| Fod 223, 225, 227, 229, 231, 233, 235, 237, 239, 241, 245, 247, 249, 251, 253 | Chipiona(Ca) | Plant | 2011 | R2I |
| Fod 200, 201, 203, 205 | Chipiona(Ca) | Plant | 2011 | R1t |
| Fod 271 | Chipiona(Ca) | Soil | 2011 | R2I |
| Fod 256, 258, 259, 260, 261, 264, 265, 267, 269, 272, 275, 281 | Chipiona(Ca) | Soil | 2011 | R2II |
| Fod 270, 280 | Chipiona(Ca) | Soil | 2011 | R1t |
| Fod 207, 210, 211, 213, 215, 217, 219, 221 | Lebrija(Se) | Plant | 2011 | R2I |
| Fod 327, 328, 329, 330, 331, 332, 333, 334, 335, 336, 341.1, 357, 358, 359, 360, 361, 364, 368, 373.2, 433, 434, 435, 436, 453 | Chipiona(Ca) | Plant | 2012 | R2I |
| Fod 365, 452 | Chipiona(Ca) | Plant | 2012 | R2II |
| Fod 341.2, 342, 343, 344, 345, 346, 347, 348, 349, 350, 351, 363, 366, 367, 371, 372, 373.1, 471 | Chipiona(Ca) | Plant | 2012 | R1t |
| Fod 337, 338, 339, 340, 369, 370, 375, 437, 438, 439, 442, 443, 444, 445, 446, 451, 469 | Chipiona(Ca) | Soil | 2012 | R2I |
| Fod 362, 376, 440, 441, 447, 448, 450, 454, 459, 463, 476 | Chipiona(Ca) | Soil | 2012 | R2II |
| Fod 352, 353, 354, 355, 356, 374, 449, 455, 456, 457, 458, 460, 461, 462, 464, 465, 466, 467, 468, 470, 472, 473, 474, 475, 477, 478, 479, 480, 481, 482, 483, 484, 485, 486, 487 | Chipiona(Ca) | Soil | 2012 | R1t |
| Fod 383, 384, 385, 386, 387, 388 | Lebrija(Se) | Plant | 2012 | R2I |
| Fod 377, 378, 379, 380, 381, 382, 391, 392, 393, 394 | Lebrija(Se) | Plant | 2012 | R1t |
| Fod 400, 402 | Lebrija(Se) | Soil | 2012 | R2I |
| Fod 389, 390, 395, 403, 404, 405 | Lebrija(Se) | Soil | 2012 | R2II |
| Fod 396, 397, 398, 399, 401 | Lebrija(Se) | Soil | 2012 | R1t |
| Fod 282, 283, 291, 292, 293, 294, 295, 296, 299, 300, 301, 316, 317, 318, 320, 321, 322, 323 | Colombia | Plant | 2012 | R1t |
| Fod 303, 304, 305, 306, 307, 308, 309, 310, 311 | Colombia | Plant | 2012 | R2I |
| Fod 297, 298, 316, 319, 322 | Colombia | Plant | 2012 | R2II |
| Fod 284, 287, 288, 289 | Colombia | Soil | 2012 | R2II |
| Fod 302, 312, 313, 314, 315 | Colombia | Soil | 2012 | R2I |
| Fod 285, 286, 290, 324, 325, 326 | Colombia | Soil | 2012 | R1t |
| Virusa | Acronym | Length (nt/aa Size) | Overlap (aa Identities %) | Bit score /e-value | Query Cover | GenBank Accesión No. |
|---|---|---|---|---|---|---|
| Rhizoctonia solani mitovirus 10 | RsMV10 | 2701/752 | 345/563(61) | 435/6e-138 | 70% | ALD89102.1 |
| Entomophthora muscae mitovirus 7 | EnmuMV7 | 2300/701 | 315/551(57) | 385/1e-119 | 69% | QDH86825.1 |
| Hubei narna-like virus 25 | HNV25 | 2375/701 | 328/619(52) | 357/1e-108 | 77% | YP_009336494.1 |
| Loramyces juncicola mitovirus 1 | LjMV1 | 2416/688 | 317/560(56) | 352/7e-11 | 70% | AZT88622.1 |
| Sclerotinia sclerotiorum mitovirus 6 | SsMV6 | 2566/703 | 311/562(5) | 325/2e-96 | 70% | AHF48622.1 |
| Sclerotinia sclerotiorum mitovirus 26 | SsMV26 | 1262/420 | 242/391(61) | 311/1e-94 | 48% | AWY10984.1 |
| Host Isolatea | RdRp (% nt Identity)b | Geographic Originc | Source | Year | Race group Assignation by PCR Patternd |
|---|---|---|---|---|---|
| Fod 124 | 97% | Chipiona(Ca) | Plant | 2009 | R2I |
| Fod 233 | 96% | Chipiona(Ca) | Plant | 2011 | R2I |
| Fod 235 | 95% | Chipiona(Ca) | Plant | 2011 | R2I |
| Fod 237 | 95% | Chipiona(Ca) | Plant | 2011 | R2I |
| Fod 253 | 95% | Chipiona(Ca) | Plant | 2011 | R2I |
| Fod 219 | 95% | Lebrija(Se) | Plant | 2011 | R2I |
| Fod 215 | 97% | Lebrija(Se) | Plant | 2011 | R2I |
| Fod 221 | 95% | Lebrija(Se) | Plant | 2011 | R2I |
| Fod 327 | 96% | Chipiona(Ca) | Plant | 2012 | R2I |
| Fod 328 | 95% | Chipiona(Ca) | Plant | 2012 | R2I |
| Fod 330 | 96% | Chipiona(Ca) | Plant | 2012 | R2I |
| Fod 331 | 97% | Chipiona(Ca) | Plant | 2012 | R2I |
| Fod 332 | 95% | Chipiona(Ca) | Plant | 2012 | R2I |
| Fod 333 | 96% | Chipiona(Ca) | Plant | 2012 | R2I |
| Fod 334 | 97% | Chipiona(Ca) | Plant | 2012 | R2I |
| Fod 341.1 | 95% | Chipiona(Ca) | Plant | 2012 | R2I |
| Fod 337 | 97% | Chipiona(Ca) | Soil | 2012 | R2I |
| Fod 338 | 95% | Chipiona(Ca) | Soil | 2012 | R2I |
| Fod 340 | 96% | Chipiona(Ca) | Soil | 2012 | R2I |
| Fod 310 | 100% | Colombia | Plant | 2012 | R2I |
| Fod 311 | 100% | Colombia | Plant | 2012 | R2I |
| Fod 313 | 100% | Colombia | Soil | 2012 | R2I |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torres-Trenas, A.; Pérez-Artés, E. Characterization and Incidence of the First Member of the Genus Mitovirus Identified in the Phytopathogenic Species Fusarium oxysporum. Viruses 2020, 12, 279. https://doi.org/10.3390/v12030279
Torres-Trenas A, Pérez-Artés E. Characterization and Incidence of the First Member of the Genus Mitovirus Identified in the Phytopathogenic Species Fusarium oxysporum. Viruses. 2020; 12(3):279. https://doi.org/10.3390/v12030279
Chicago/Turabian StyleTorres-Trenas, Almudena, and Encarnación Pérez-Artés. 2020. "Characterization and Incidence of the First Member of the Genus Mitovirus Identified in the Phytopathogenic Species Fusarium oxysporum" Viruses 12, no. 3: 279. https://doi.org/10.3390/v12030279
APA StyleTorres-Trenas, A., & Pérez-Artés, E. (2020). Characterization and Incidence of the First Member of the Genus Mitovirus Identified in the Phytopathogenic Species Fusarium oxysporum. Viruses, 12(3), 279. https://doi.org/10.3390/v12030279