Preliminary Identification of Potential Vaccine Targets for the COVID-19 Coronavirus (SARS-CoV-2) Based on SARS-CoV Immunological Studies


Abstract
1. Introduction
2. Materials and Methods
2.1. Acquisition and Processing of Sequence Data
2.2. Acquisition and Filtering of Epitope Data
2.3. Population-Coverage-Based T Cell Epitope Selection
2.4. Constructing the Phylogenetic Tree
2.5. Data and Code Availability
3. Results
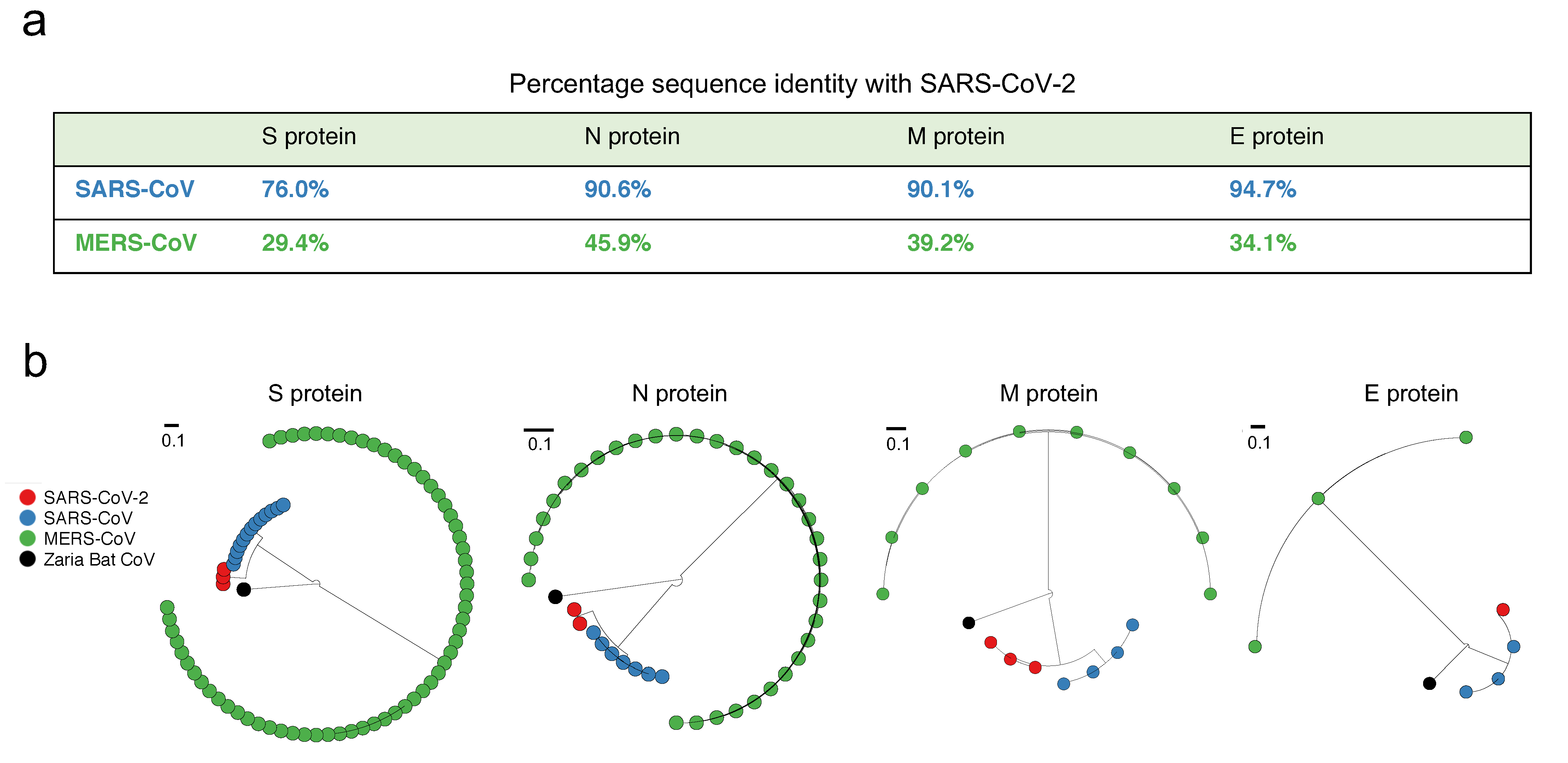
3.1. Structural Proteins of SARS-CoV-2 Are Genetically Similar to SARS-CoV, but Not to MERS-CoV
3.2. Mapping the SARS-CoV-Derived T Cell Epitopes That Are Identical in SARS-CoV-2, and Determining Those With Greatest Estimated Population Coverage
3.3. Mapping the SARS-CoV-Derived B cell Epitopes that Are Identical in SARS-CoV-2
4. Discussion
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Wang, C.; Horby, P.W.; Hayden, F.G.; Gao, G.F. A novel coronavirus outbreak of global health concern. Lancet 2020, 395, 470–473. [Google Scholar] [CrossRef]
- Centers-of-Disease-Control-and-Prevention Confirmed 2019-nCoV cases globally. Available online: https://www.cdc.gov/coronavirus/2019-ncov/locations-confirmed-cases.html (accessed on 31 January 2020).
- World-Health-Organization Statement on the second meeting of the International Health Regulations (2005) Emergency Committee regarding the outbreak of novel coronavirus (2019-nCoV). Available online: https://www.who.int/news-room/detail/30-01-2020-statement-on-the-second-meeting-of-the-international-health-regulations-(2005)-emergency-committee-regarding-the-outbreak-of-novel-coronavirus-(2019-ncov) (accessed on 31 January 2020).
- World-Health-Organization Coronavirus disease (COVID-19) outbreak. Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019 (accessed on 31 January 2020).
- World-Health-Organization Statement on the meeting of the International Health Regulations (2005) Emergency Committee regarding the outbreak of novel coronavirus (2019-nCoV). Available online: https://www.who.int/news-room/detail/23-01-2020-statement-on-the-meeting-of-the-international-health-regulations-(2005)-emergency-committee-regarding-the-outbreak-of-novel-coronavirus-(2019-ncov) (accessed on 31 January 2020).
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef]
- Heymann, D.L. Data sharing and outbreaks: Best practice exemplified. Lancet 2020, 395, 469–470. [Google Scholar] [CrossRef]
- Liu, X.; Wang, X.-J. Potential inhibitors for 2019-nCoV coronavirus M protease from clinically approved medicines. bioRxiv 2020, 2020.01.29.924100. [Google Scholar]
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020. [Google Scholar] [CrossRef] [PubMed]
- World-Health-Organization Update 49 - SARS case fatality ratio, incubation period. Available online: https://www.who.int/csr/sars/archive/2003_05_07a/en/ (accessed on 31 January 2020).
- World-Health-Organization Middle East respiratory syndrome coronavirus (MERS-CoV). Available online: https://www.who.int/emergencies/mers-cov/en/ (accessed on 31 January 2020).
- Lu, R.; Zhao, X.; Li, J.; Niu, P.; Yang, B.; Wu, H.; Wang, W.; Song, H.; Huang, B.; Zhu, N.; et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: Implications for virus origins and receptor binding. Lancet 2020, 6736, 1–10. [Google Scholar] [CrossRef]
- Letko, M.; Munster, V. Functional assessment of cell entry and receptor usage for lineage B β-coronaviruses, including 2019-nCoV. bioRxiv 2020, 2020.01.22.915660. [Google Scholar]
- Hoffmann, M.; Kleine-Weber, H.; Kruger, N.; Muller, M.; Drosten, C.; Pohlmann, S. The novel coronavirus 2019 (2019-nCoV) uses the SARS-coronavirus receptor ACE2 and the cellular protease TMPRSS2 for entry into target cells. bioRxiv 2020, 2020.01.31.929042. [Google Scholar]
- Yang, Z.-Y.; Kong, W.-P.; Huang, Y.; Roberts, A.; Murphy, B.R.; Subbarao, K.; Nabel, G.J. A DNA vaccine induces SARS coronavirus neutralization and protective immunity in mice. Nature 2004, 428, 561–564. [Google Scholar] [CrossRef]
- Deming, D.; Sheahan, T.; Heise, M.; Yount, B.; Davis, N.; Sims, A.; Suthar, M.; Harkema, J.; Whitmore, A.; Pickles, R.; et al. Vaccine efficacy in senescent mice challenged with recombinant SARS-CoV bearing epidemic and zoonotic spike variants. PLoS Med. 2006, 3, e525. [Google Scholar] [CrossRef]
- Graham, R.L.; Becker, M.M.; Eckerle, L.D.; Bolles, M.; Denison, M.R.; Baric, R.S. A live, impaired-fidelity coronavirus vaccine protects in an aged, immunocompromised mouse model of lethal disease. Nat. Med. 2012, 18, 1820–1826. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Shen, X.; Yang, R.F.; Li, Y.X.; Ji, Y.Y.; He, Y.Y.; De Shi, M.; Lu, W.; Shi, T.L.; Wang, J.; et al. Identification of an epitope of SARS-coronavirus nucleocapsid protein. Cell Res. 2003, 13, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wen, J.; Li, J.; Yin, J.; Zhu, Q.; Wang, H.; Yang, Y.; Qin, E.; You, B.; Li, W.; et al. Assessment of immunoreactive synthetic peptides from the structural proteins of severe acute respiratory syndrome coronavirus. Clin. Chem. 2003, 49, 1989–1996. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Shi, Y.; Li, P.; Li, L.; Yi, Y.; Ma, Q.; Cao, C. Profile of antibodies to the nucleocapsid protein of the severe acute respiratory syndrome (SARS)-associated coronavirus in probable SARS patients. Clin. Vaccine Immunol. 2004, 11, 227–228. [Google Scholar] [CrossRef] [PubMed]
- Tang, F.; Quan, Y.; Xin, Z.-T.; Wrammert, J.; Ma, M.-J.; Lv, H.; Wang, T.-B.; Yang, H.; Richardus, J.H.; Liu, W.; et al. Lack of peripheral memory B cell responses in recovered patients with severe acute respiratory syndrome: A six-year follow-up study. J. Immunol. 2011, 186, 7264–7268. [Google Scholar] [CrossRef]
- Peng, H.; Yang, L.-T.; Wang, L.-Y.; Li, J.; Huang, J.; Lu, Z.-Q.; Koup, R.A.; Bailer, R.T.; Wu, C.-Y. Long-lived memory T lymphocyte responses against SARS coronavirus nucleocapsid protein in SARS-recovered patients. Virology 2006, 351, 466–475. [Google Scholar] [CrossRef]
- Fan, Y.-Y.; Huang, Z.-T.; Li, L.; Wu, M.-H.; Yu, T.; Koup, R.A.; Bailer, R.T.; Wu, C.-Y. Characterization of SARS-CoV-specific memory T cells from recovered individuals 4 years after infection. Arch. Virol. 2009, 154, 1093–1099. [Google Scholar] [CrossRef]
- Ng, O.-W.; Chia, A.; Tan, A.T.; Jadi, R.S.; Leong, H.N.; Bertoletti, A.; Tan, Y.-J. Memory T cell responses targeting the SARS coronavirus persist up to 11 years post-infection. Vaccine 2016, 34, 2008–2014. [Google Scholar] [CrossRef]
- Liu, W.J.; Zhao, M.; Liu, K.; Xu, K.; Wong, G.; Tan, W.; Gao, G.F. T-cell immunity of SARS-CoV: Implications for vaccine development against MERS-CoV. Antiviral Res. 2017, 137, 82–92. [Google Scholar] [CrossRef]
- Li, C.K.-F.; Wu, H.; Yan, H.; Ma, S.; Wang, L.; Zhang, M.; Tang, X.; Temperton, N.J.; Weiss, R.A.; Brenchley, J.M.; et al. T cell responses to whole SARS coronavirus in humans. J. Immunol. 2008, 181, 5490–5500. [Google Scholar] [CrossRef]
- Channappanavar, R.; Fett, C.; Zhao, J.; Meyerholz, D.K.; Perlman, S. Virus-specific memory CD8 T cells provide substantial protection from lethal severe acute respiratory syndrome coronavirus infection. J. Virol. 2014, 88, 11034–11044. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Pickett, B.E.; Sadat, E.L.; Zhang, Y.; Noronha, J.M.; Squires, R.B.; Hunt, V.; Liu, M.; Kumar, S.; Zaremba, S.; Gu, Z.; et al. ViPR: An open bioinformatics database and analysis resource for virology research. Nucleic Acids Res. 2012, 40, D593–D598. [Google Scholar] [CrossRef] [PubMed]
- Vita, R.; Mahajan, S.; Overton, J.A.; Dhanda, S.K.; Martini, S.; Cantrell, J.R.; Wheeler, D.K.; Sette, A.; Peters, B. The immune epitope database (IEDB): 2018 update. Nucleic Acids Res. 2019, 47, D339–D343. [Google Scholar] [CrossRef]
- Mirarab, S.; Nguyen, N.; Guo, S.; Wang, L.-S.; Kim, J.; Warnow, T. PASTA: Ultra-large multiple sequence alignment for nucleotide and amino-acid sequences. J. Comput. Biol. 2015, 22, 377–386. [Google Scholar] [CrossRef]
- Huson, D.H.; Scornavacca, C. Dendroscope 3: An interactive tool for rooted phylogenetic trees and networks. Syst. Biol. 2012, 61, 1061–1067. [Google Scholar] [CrossRef]
- Ahmed, S.F. Data and software code for reproducing results of this paper. Available online: https://github.com/faraz107/2019-nCoV-T-Cell-Vaccine-Candidates (accessed on 31 January 2020).
- Li, F. Structure of SARS coronavirus spike receptor-binding domain complexed with receptor. Science. 2005, 309, 1864–1868. [Google Scholar] [CrossRef]
- Dahirel, V.; Shekhar, K.; Pereyra, F.; Miura, T.; Artyomov, M.; Talsania, S.; Allen, T.M.; Altfeld, M.; Carrington, M.; Irvine, D.J.; et al. Coordinate linkage of HIV evolution reveals regions of immunological vulnerability. Proc. Natl. Acad. Sci. 2011, 108, 11530–11535. [Google Scholar] [CrossRef]
- Quadeer, A.A.; Louie, R.H.Y.; Shekhar, K.; Chakraborty, A.K.; Hsing, I.-M.; McKay, M.R. Statistical linkage analysis of substitutions in patient-derived sequences of genotype 1a hepatitis C virus nonstructural protein 3 exposes targets for immunogen design. J. Virol. 2014, 88, 7628–7644. [Google Scholar] [CrossRef][Green Version]
- Ahmed, S.F.; Quadeer, A.A.; Morales-Jimenez, D.; McKay, M.R. Sub-dominant principal components inform new vaccine targets for HIV Gag. Bioinformatics 2019, 35, 3884–3889. [Google Scholar] [CrossRef]
- Quadeer, A.A.; Morales-Jimenez, D.; McKay, M.R. Co-evolution networks of HIV/HCV are modular with direct association to structure and function. PLOS Comput. Biol. 2018, 14, e1006409. [Google Scholar] [CrossRef]
- Prabakaran, P.; Gan, J.; Feng, Y.; Zhu, Z.; Choudhry, V.; Xiao, X.; Ji, X.; Dimitrov, D.S. Structure of severe acute respiratory syndrome coronavirus receptor-binding domain complexed with neutralizing antibody. J. Biol. Chem. 2006, 281, 15829–15836. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Chakraborti, S.; He, Y.; Roberts, A.; Sheahan, T.; Xiao, X.; Hensley, L.E.; Prabakaran, P.; Rockx, B.; Sidorov, I.A.; et al. Potent cross-reactive neutralization of SARS coronavirus isolates by human monoclonal antibodies. Proc. Natl. Acad. Sci. 2007, 104, 12123–12128. [Google Scholar] [CrossRef]
- Hwang, W.C.; Lin, Y.; Santelli, E.; Sui, J.; Jaroszewski, L.; Stec, B.; Farzan, M.; Marasco, W.A.; Liddington, R.C. Structural basis of neutralization by a human anti-severe acute respiratory syndrome spike protein antibody, 80R. J. Biol. Chem. 2006, 281, 34610–34616. [Google Scholar] [CrossRef] [PubMed]
- UniProt UniProtKB - P59594 (SPIKE_CVHSA). Available online: https://www.uniprot.org/uniprot/P59594 (accessed on 31 January 2020).
- Walls, A.C.; Park, Y.-J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, function and antigenicity of the SARS-CoV-2 spike glycoprotein. bioRxiv 2020, 2020.02.19.956581. [Google Scholar]
- Walls, A.C.; Xiong, X.; Park, Y.-J.; Tortorici, M.A.; Snijder, J.; Quispe, J.; Cameroni, E.; Gopal, R.; Dai, M.; Lanzavecchia, A.; et al. Unexpected receptor functional mimicry elucidates activation of coronavirus fusion. Cell 2019, 176, 1026–1039.e15. [Google Scholar] [CrossRef] [PubMed]
- Walls, A.C.; Tortorici, M.A.; Snijder, J.; Xiong, X.; Bosch, B.-J.; Rey, F.A.; Veesler, D. Tectonic conformational changes of a coronavirus spike glycoprotein promote membrane fusion. Proc. Natl. Acad. Sci. 2017, 114, 11157–11162. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Gui, M.; Wang, X.; Xiang, Y. Cryo-EM structure of the SARS coronavirus spike glycoprotein in complex with its host cell receptor ACE2. PLOS Pathog. 2018, 14, e1007236. [Google Scholar] [CrossRef]
- Wrapp, D.; Wang, N.; Corbett, K.S.; Goldsmith, J.A.; Hsieh, C.-L.; Abiona, O.; Graham, B.S.; McLellan, J.S. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science. 2020, 2011, eabb2507. [Google Scholar] [CrossRef]
- Tian, X.; Li, C.; Huang, A.; Xia, S.; Lu, S.; Shi, Z.; Lu, L.; Jiang, S.; Yang, Z.; Wu, Y.; et al. Potent binding of 2019 novel coronavirus spike protein by a SARS coronavirus-specific human monoclonal antibody. Emerg. Microbes Infect. 2020, 9, 382–385. [Google Scholar] [CrossRef]
- Ferguson, A.L.; Mann, J.K.; Omarjee, S.; Ndung’u, T.; Walker, B.D.; Chakraborty, A.K. Translating HIV sequences into quantitative fitness landscapes predicts viral vulnerabilities for rational immunogen design. Immunity 2013, 38, 606–617. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, A.K.; Barton, J.P. Rational design of vaccine targets and strategies for HIV: A crossroad of statistical physics, biology, and medicine. Reports Prog. Phys. 2017, 80, 032601. [Google Scholar] [CrossRef] [PubMed]
- Quadeer, A.A.; Louie, R.H.Y.; McKay, M.R. Identifying immunologically-vulnerable regions of the HCV E2 glycoprotein and broadly neutralizing antibodies that target them. Nat. Commun. 2019, 10, 2073. [Google Scholar] [CrossRef] [PubMed]
- Louie, R.H.Y.; Kaczorowski, K.J.; Barton, J.P.; Chakraborty, A.K.; McKay, M.R. Fitness landscape of the human immunodeficiency virus envelope protein that is targeted by antibodies. Proc. Natl. Acad. Sci. 2018, 115, E564–E573. [Google Scholar] [CrossRef]
- Quadeer, A.A.; Barton, J.P.; Chakraborty, A.K.; McKay, M.R. Deconvolving mutational patterns of poliovirus outbreaks reveals its intrinsic fitness landscape. Nat. Commun. 2020, 11, 377. [Google Scholar] [CrossRef]
- Mann, J.K.; Barton, J.P.; Ferguson, A.L.; Omarjee, S.; Walker, B.D.; Chakraborty, A.; Ndung’u, T. The fitness landscape of HIV-1 Gag: Advanced modeling approaches and validation of model predictions by in vitro testing. PLoS Comput. Biol. 2014, 10, e1003776. [Google Scholar] [CrossRef]
- Ramaiah, A.; Arumugaswami, V. Insights into cross-species evolution of novel human coronavirus 2019-nCoV and defining immune determinants for vaccine development. bioRxiv 2020, 2020.01.29.925867. [Google Scholar]



{kind=link}
{kind=link}
| Filtering Criteria | Number of Epitopes | |
|---|---|---|
| Positive T cell assays | T cell epitopes | 115 |
| Positive major histocompatibility complex (MHC) binding assays | T cell epitopes | 959 |
| Positive B cell assays | Linear B cell epitopes | 298 |
| Discontinuous B cell epitopes | 6 | |
| Protein | IEDB ID | Epitope | MHC Allele1 | MHC Allele Class 1 |
|---|---|---|---|---|
| N | 125100 | ILLNKHID | HLA-A*02:01 | I |
| N | 1295 | AFFGMSRIGMEVTPSGTW | NA | NA |
| N | 190494 | MEVTPSGTWL | HLA-B*40:01 | I |
| N | 21347 | GMSRIGMEV | HLA-A*02:01 | I |
| N | 27182 | ILLNKHIDA | HLA-A*02:01 | I |
| N | 2802 | ALNTPKDHI | HLA-A*02:01 | I |
| N | 28371 | IRQGTDYKHWPQIAQFA | NA | NA |
| N | 31166 | KHWPQIAQFAPSASAFF | NA | NA |
| N | 34851 | LALLLLDRL | HLA-A*02:01 | I |
| N | 37473 | LLLDRLNQL | HLA-A*02:01 | I |
| N | 37611 | LLNKHIDAYKTFPPTEPK | NA | NA |
| N | 38881 | LQLPQGTTL | HLA-A*02:01 | I |
| N | 3957 | AQFAPSASAFFGMSR | NA | II |
| N | 3958 | AQFAPSASAFFGMSRIGM | NA | NA |
| N | 55683 | RRPQGLPNNTASWFT | NA | I |
| N | 74517 | YKTFPPTEPKKDKKKK | NA | NA |
| S | 100048 | GAALQIPFAMQMAYRF | HLA-DRA*01:01, HLA-DRB1*07:01 | II |
| S | 100300 | MAYRFNGIGVTQNVLY | HLA-DRB1*04:01 | II |
| S | 100428 | QLIRAAEIRASANLAATK | HLA-DRB1*04:01 | II |
| S | 16156 | FIAGLIAIV | HLA-A*02:01 | I |
| S | 2801 | ALNTLVKQL | HLA-A*02:01 | I |
| S | 36724 | LITGRLQSL | HLA-A2 | I |
| S | 44814 | NLNESLIDL | HLA-A*02:01 | I |
| S | 50311 | QALNTLVKQLSSNFGAI | HLA-DRB1*04:01 | II |
| S | 54680 | RLNEVAKNL | HLA-A*02:01 | I |
| S | 69657 | VLNDILSRL | HLA-A*02:01 | I |
| S | 71663 | VVFLHVTYV | HLA-A*02:01 | I |
| Epitopes1 | MHC Allele Class | MHC Allele | Global Accumulated Population Coverage2 (%) | Accumulated Population Coverage in China (%) |
|---|---|---|---|---|
| FIAGLIAIV, GLIAIVMVTI, IITTDNTFV, ALNTLVKQL, LITGRLQSL, LLLQYGSFC, LQYGSFCT, NLNESLIDL, RLDKVEAEV, RLNEVAKNL, RLQSLQTYV, VLNDILSRL, VVFLHVTYV, ILLNKHID, FPRGQGVPI, LLLLDRLNQ, GMSRIGMEV, ILLNKHIDA, ALNTPKDHI, LALLLLDRL, LLLDRLNQL, LLLLDRLNQL, LQLPQGTTL, AQFAPSASA, TTLPKGFYA, VLQLPQGTTL | I | HLA-A*02:01 | 39.08 | 14.62 |
| GYQPYRVVVL, PYRVVVLSF, LSPRWYFYY | I | HLA-A*24:02 | 55.48 | 36.11 |
| DSFKEELDKY, LIDLQELGKY, PYRVVVLSF, GTTLPKGFY, VTPSGTWLTY | I | HLA-A*01:01 | 66.78 | 39.09 |
| GSFCTQLNR, GVVFLHVTY, AQALNTLVK, MTSCCSCLK, ASANLAATK, SLIDLQELGK, SVLNDILSR, TQNVLYENQK, CMTSCCSCLK, VQIDRLITGR, KTFPPTEPK, KTFPPTEPKK, LSPRWYFYY, ASAFFGMSR, ATEGALNTPK, QLPQGTTLPK, QQQGQTVTK, QQQQGQTVTK, SASAFFGMSR, SQASSRSSSR, TPSGTWLTY | I | HLA-A*03:01 | 76.14 | 41.68 |
| GSFCTQLNR, GVVFLHVTY, AQALNTLVK, MTSCCSCLK, ASANLAATK, SLIDLQELGK, SVLNDILSR, TQNVLYENQK, CMTSCCSCLK, VQIDRLITGR, KTFPPTEPK, KTFPPTEPKK, LSPRWYFYY, ASAFFGMSR, ATEGALNTPK, QLPQGTTLPK, QQQGQTVTK, QQQQGQTVTK, SASAFFGMSR, SQASSRSSSR, TPSGTWLTY | I | HLA-A*11:01 | 83.39 | 73.43 |
| GSFCTQLNR, GVVFLHVTY, AQALNTLVK, MTSCCSCLK, ASANLAATK, SLIDLQELGK, SVLNDILSR, TQNVLYENQK, CMTSCCSCLK, VQIDRLITGR, KTFPPTEPK, KTFPPTEPKK, LSPRWYFYY, ASAFFGMSR, ATEGALNTPK, QLPQGTTLPK, QQQGQTVTK, QQQQGQTVTK, SASAFFGMSR, SQASSRSSSR, TPSGTWLTY | I | HLA-A*68:01 | 85.71 | 74.25 |
| GYQPYRVVVL, PYRVVVLSF, LSPRWYFYY | I | HLA-A*23:01 | 87.72 | 74.87 |
| GSFCTQLNR, GVVFLHVTY, AQALNTLVK, MTSCCSCLK, ASANLAATK, SLIDLQELGK, SVLNDILSR, TQNVLYENQK, CMTSCCSCLK, VQIDRLITGR, KTFPPTEPK, KTFPPTEPKK, LSPRWYFYY, ASAFFGMSR, ATEGALNTPK, QLPQGTTLPK, QQQGQTVTK, QQQQGQTVTK, SASAFFGMSR, SQASSRSSSR, TPSGTWLTY | I | HLA-A*31:01 | 89.55 | 76.93 |
| FPNITNLCPF, APHGVVFLHV, FPRGQGVPI, APSASAFFGM | I | HLA-B*07:02 | 90.89 | 77.61 |
| GAALQIPFAMQMAYR, GWTFGAGAALQIPFA, IDRLITGRLQSLQTY, ISGINASVVNIQKEI, LDKYFKNHTSPDVDL, LGDISGINASVVNIQ, LGFIAGLIAIVMVTI, LNTLVKQLSSNFGAI, LQDVVNQNAQALNTL, LQSLQTYVTQQLIRA, LQTYVTQQLIRAAEI, AQKFNGLTVLPPLLT, PCSFGGVSVITPGTN, QIPFAMQMAYRFNGI, QQLIRAAEIRASANL, QTYVTQQLIRAAEIR, AYRFNGIGVTQNVLY, SSNFGAISSVLNDIL, TGRLQSLQTYVTQQL, WLGFIAGLIAIVMVT, CVNFNFNGLTGTGVL, DKYFKNHTSPDVDLG, IDAYKTFPPTEPKKD, MSRIGMEVTPSGTWL, NKHIDAYKTFPPTEP, VLQLPQGTTLPKGFY | II | HLA-DRB1*01:01 | 91.94 | 78.23 |
| FPRGQGVPI | I | HLA-B*08:01 | 92.85 | 78.41 |
| FPNITNLCPF, APHGVVFLHV, FPRGQGVPI, APSASAFFGM | I | HLA-B*35:01 | 93.53 | 79.23 |
| LQIPFAMQM, RVDFCGKGY | I | HLA-B*15:01 | 94.18 | 82.26 |
| FPNITNLCPF, APHGVVFLHV, FPRGQGVPI, APSASAFFGM | I | HLA-B*51:01 | 94.72 | 83.73 |
| YEQYIKWPWY | I | HLA-B*18:01 | 95.23 | 83.88 |
| GRLQSLQTY, RVDFCGKGY, VRFPNITNL | I | HLA-B*27:05 | 95.55 | 84 |
| MTSCCSCLK, SLIDLQELGK, CMTSCCSCLK, VQIDRLITGR, SASAFFGMSR, SQASSRSSSR | I | HLA-A*33:01 | 95.79 | 85.28 |
| LQIPFAMQM, RVDFCGKGY | I | HLA-B*58:01 | 95.99 | 86.45 |
| LQIPFAMQM, RVDFCGKGY | I | HLA-C*15:02 | 96.17 | 87.22 |
| VRFPNITNL | I | HLA-C*14:02 | 96.29 | 88.11 |
| Protein | Subunit | IEDB ID | Epitope | Protein | IEDB ID | Epitope |
|---|---|---|---|---|---|---|
| S | S2 | 10778 | DVVNQNAQALNTLVKQL | N | 15814 | FFGMSRIGMEVTPSGTW |
| S | S2 | 11038 | EAEVQIDRLITGRLQSL | N | 21065 | GLPNNTASWFTALTQHGK |
| S | S2 | 12426 | EIDRLNEVAKNLNESLIDLQELGKYEQY | N | 22855 | GTTLPK |
| S | S2 | 14626 | EVAKNLNESLIDLQELG | N | 28371 | IRQGTDYKHWPQIAQFA |
| S | S2 | 18515 | GAALQIPFAMQMAYRFN | N | 31116 | KHIDAYKTFPPTEPKKDKKK |
| S | S1 | 18594 | GAGICASY | N | 31166 | KHWPQIAQFAPSASAFF |
| S | S2 | 2092 | AISSVLNDILSRLDKVE | N | 75235 | YNVTQAFGRRGPEQTQGNF |
| S | S2 | 22321 | GSFCTQLN | N | 33669 | KTFPPTEPKKDKKKK |
| S | S2 | 27357 | ILSRLDKVEAEVQIDRL | N | 37640 | LLPAAD |
| S | S1 | 30987 | KGIYQTSN | N | 38249 | LNKHIDAYKTFPPTEPK |
| S | S2 | 3176 | AMQMAYRF | N | 38648 | LPQGTTLPKG |
| S | S2 | 32508 | KNHTSPDVDLGDISGIN | N | 38657 | LPQRQKKQ |
| S | S2 | 41177 | MAYRFNGIGVTQNVLYE | N | 48067 | PKGFYAEGSRGGSQASSR |
| S | S2 | 462 | AATKMSECVLGQSKRVD | N | 50741 | QFAPSASAFFGMSRIGM |
| S | S2 | 47479 | PFAMQMAYRFNGIGVTQ | N | 50965 | QGTDYKHW |
| S | S2 | 50311 | QALNTLVKQLSSNFGAI | N | 51483 | QLPQGTTLPKGFYAE |
| S | S2 | 51379 | QLIRAAEIRASANLAAT | N | 51484 | QLPQGTTLPKGFYAEGSR |
| S | S1 | 52020 | QQFGRD | N | 51485 | QLPQGTTLPKGFYAEGSRGGSQ |
| S | S2 | 53202 | RASANLAATKMSECVLG | N | 63729 | TFPPTEPK |
| S | S2 | 54599 | RLITGRLQSLQTYVTQQ | N | 55683 | RRPQGLPNNTASWFT |
| S | S2 | 558417 | EIDRLNEVAKNLNESLIDLQELGKYEQY | N | 60379 | SQASSRSS |
| S | S2 | 59425 | SLQTYVTQQLIRAAEIR | N | 60669 | SRGGSQASSRSSSRSR |
| S | S2 | 9094 | DLGDISGINASVVNIQK |
| IEDB ID | Associated Known Antibody | SARS-CoV S Protein Residues 1,2 |
|---|---|---|
| 910052 | S230 | G446, P462, D463, Y475 |
| 77444 | m396 | T359, T363, K365, K390, G391, D392, R395, R426, Y436, G482, Y484, T485, T486, T487, G488, I489, G490, Y491, Q492, Y494 |
| 77442 | 80R | R426, S432, T433, Y436, N437, K439, Y440, Y442, P469, P470, A471, L472, N473, C474, Y475, W476, L478, N479, D480, Y481, G482, Y484, T485, T486, T487, G488, I489, Y491, Q492 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, S.F.; Quadeer, A.A.; McKay, M.R. Preliminary Identification of Potential Vaccine Targets for the COVID-19 Coronavirus (SARS-CoV-2) Based on SARS-CoV Immunological Studies. Viruses 2020, 12, 254. https://doi.org/10.3390/v12030254
Ahmed SF, Quadeer AA, McKay MR. Preliminary Identification of Potential Vaccine Targets for the COVID-19 Coronavirus (SARS-CoV-2) Based on SARS-CoV Immunological Studies. Viruses. 2020; 12(3):254. https://doi.org/10.3390/v12030254
Chicago/Turabian StyleAhmed, Syed Faraz, Ahmed A. Quadeer, and Matthew R. McKay. 2020. "Preliminary Identification of Potential Vaccine Targets for the COVID-19 Coronavirus (SARS-CoV-2) Based on SARS-CoV Immunological Studies" Viruses 12, no. 3: 254. https://doi.org/10.3390/v12030254
APA StyleAhmed, S. F., Quadeer, A. A., & McKay, M. R. (2020). Preliminary Identification of Potential Vaccine Targets for the COVID-19 Coronavirus (SARS-CoV-2) Based on SARS-CoV Immunological Studies. Viruses, 12(3), 254. https://doi.org/10.3390/v12030254