Virome Variation during Sea Star Wasting Disease Progression in Pisaster ochraceus (Asteroidea, Echinodermata)

Abstract
1. Introduction
2. Materials and Methods
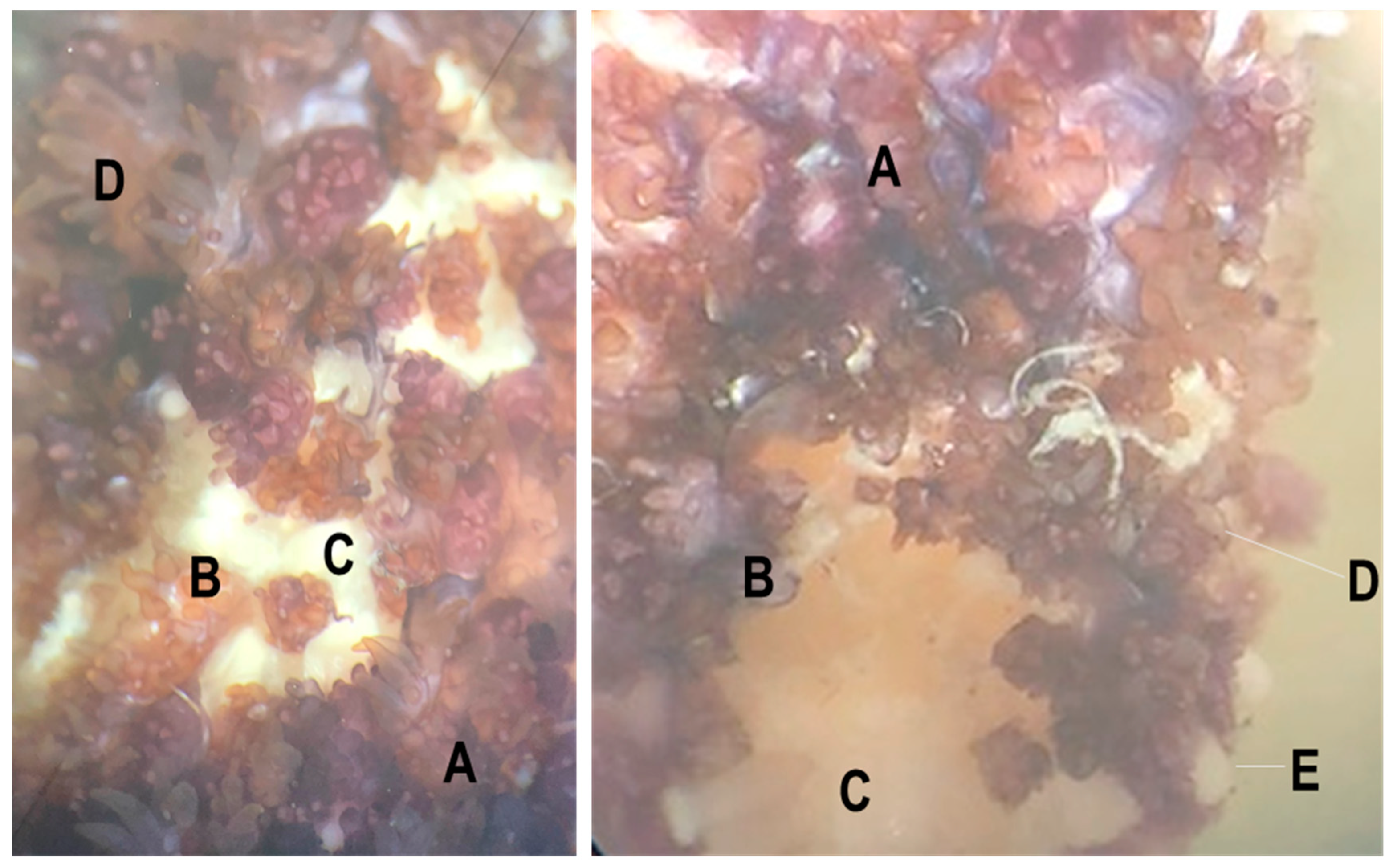
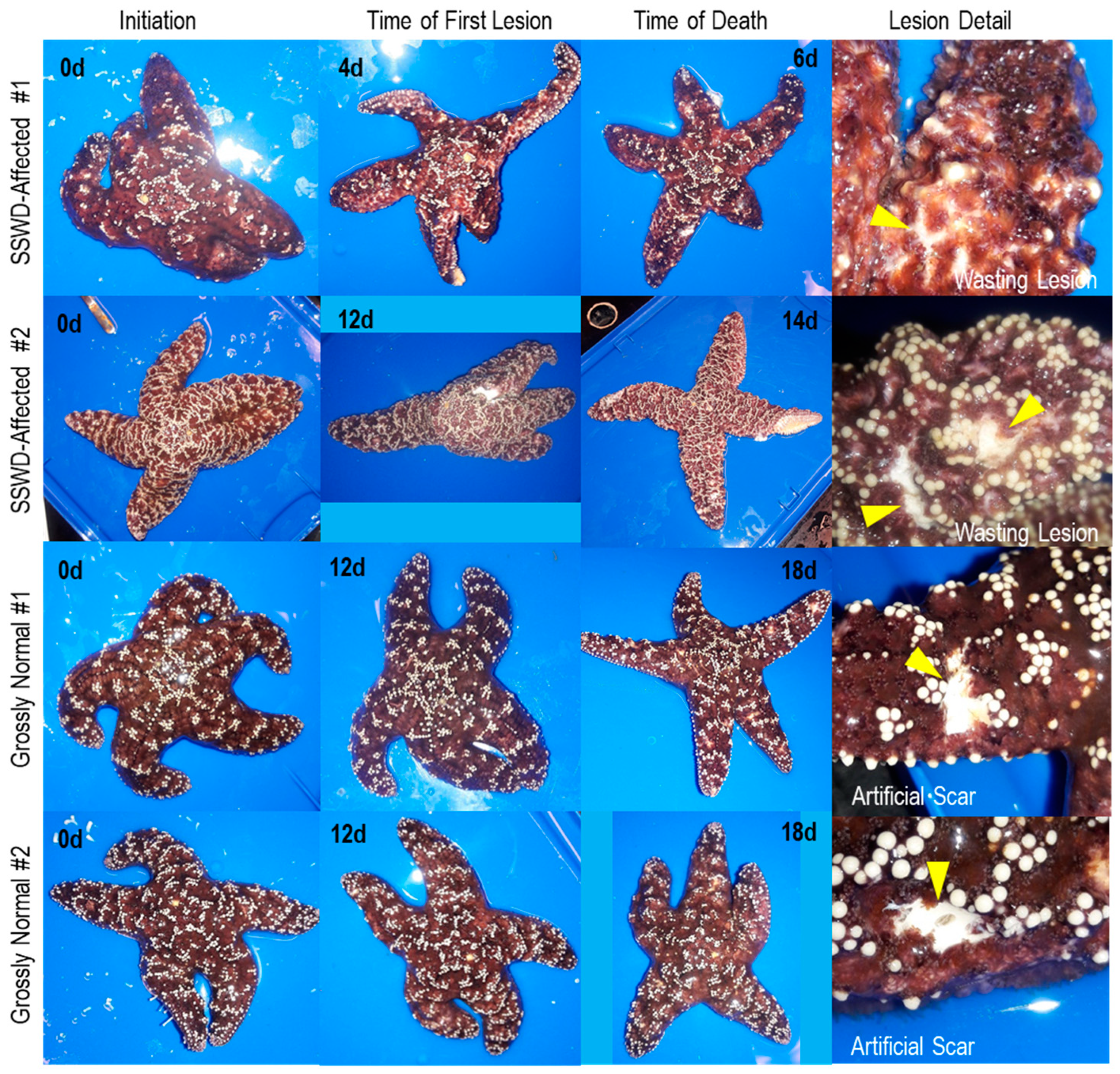
2.1. Survey Design
2.2. Viral Metagenome Preparation
2.3. Bioinformatic Processing
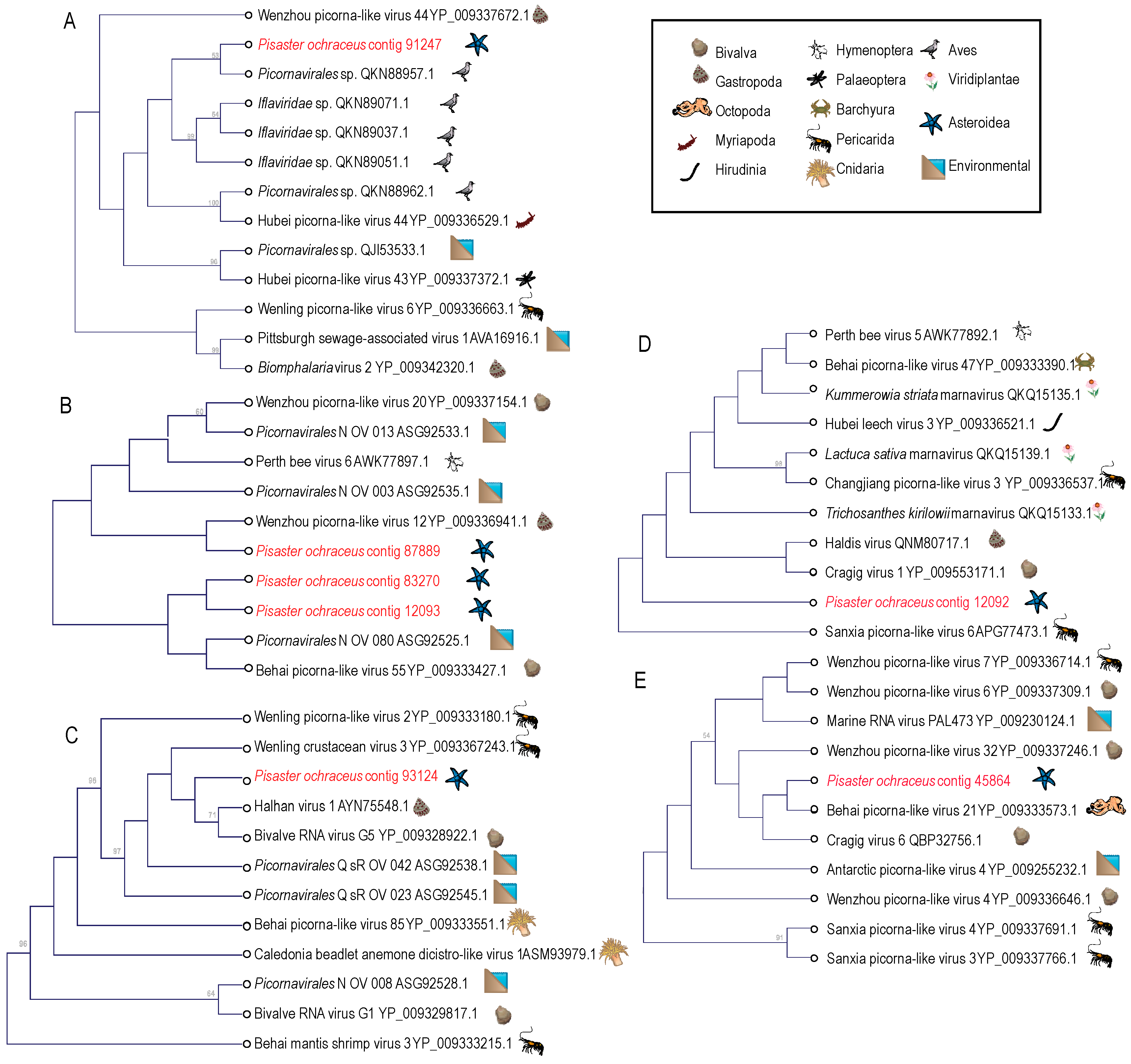
2.4. Phylogenetic Analyses
2.5. Statistical Analyses
3. Results and Discussion
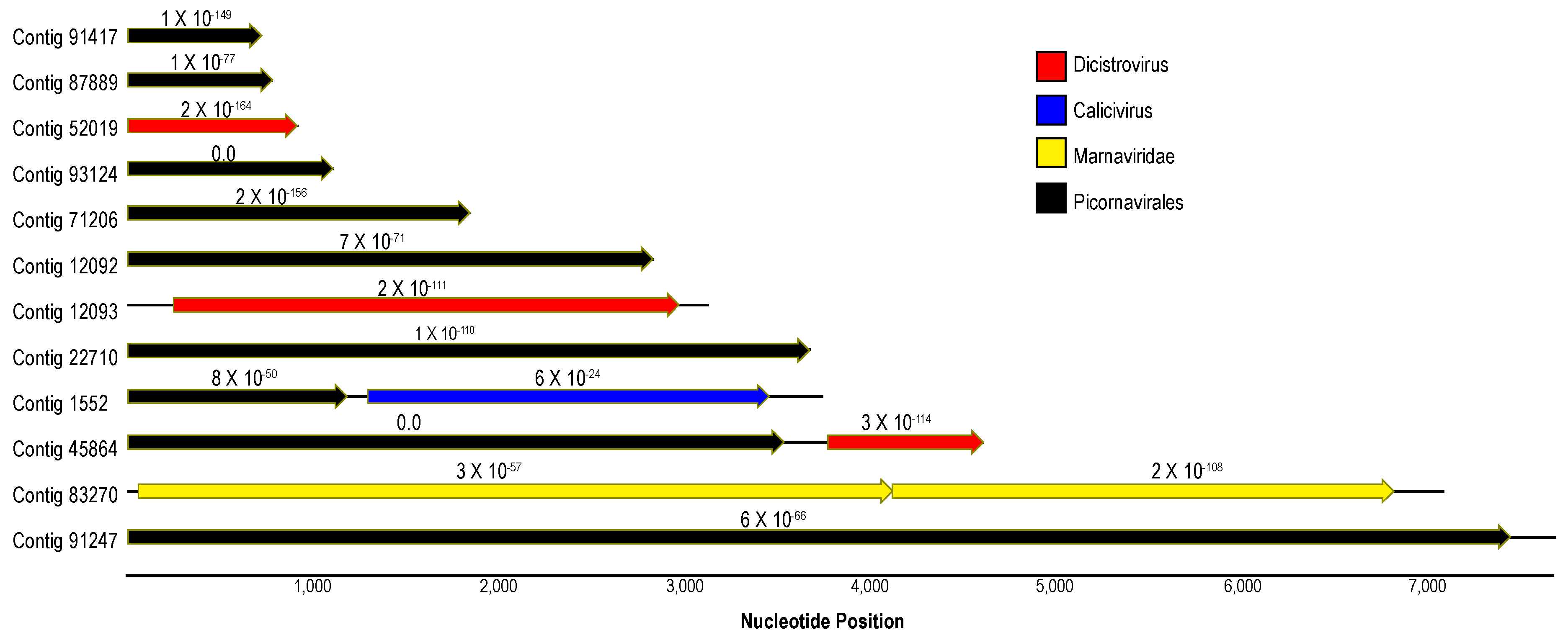
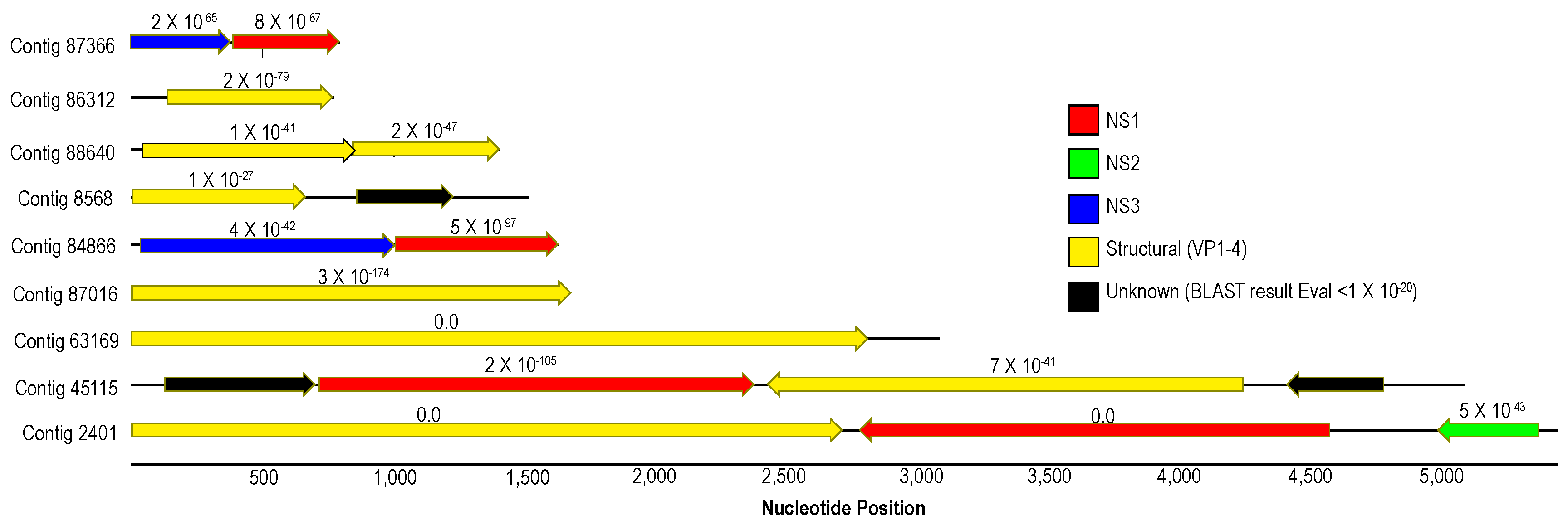
3.1. Description of Viruses Recovered in Viral Metagenomes
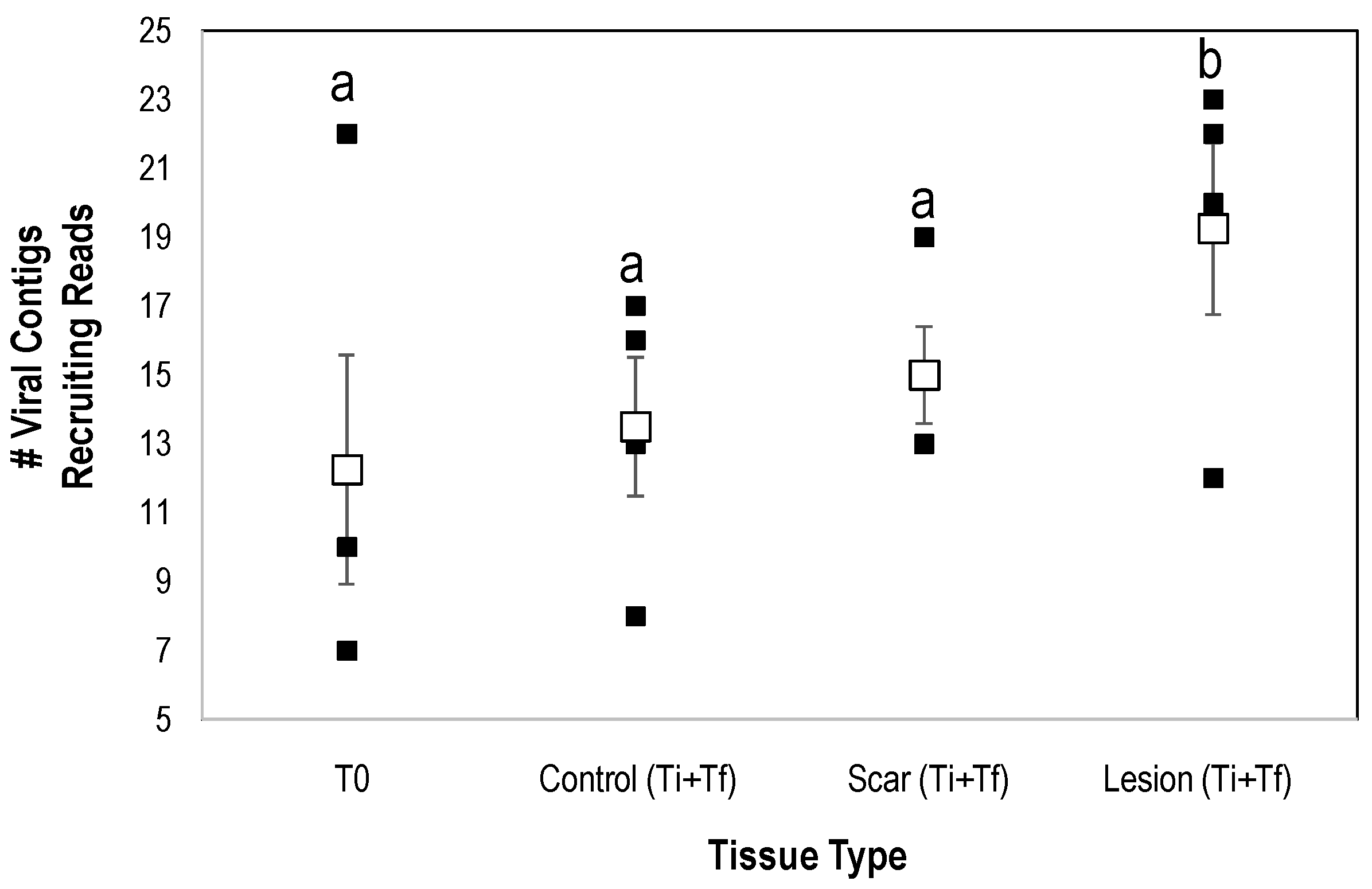
3.2. Analysis of Virome Association with Sea Star Wasting Disease
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mead, A.D. Twenty-Eighth Annual Report of the Commissioners of Inland Fisheries, Made to the General Assembly at Its January Session, 1898; Forgotten Books: Providence, RI, USA, 1898; pp. 11–29. [Google Scholar]
- Hewson, I.; Button, J.B.; Gudenkauf, B.M.; Miner, B.; Newton, A.L.; Gaydos, J.K.; Wynne, J.; Groves, C.J.; Hendler, G.; Murray, M.; et al. Densovirus associated with sea-star wasting disease and mass mortality. Proc. Natl. Acad. Sci. USA 2014, 111, 17276–17283. [Google Scholar] [CrossRef] [PubMed]
- Hewson, I.; Sullivan, B.; Jackson, E.W.; Xu, Q.; Long, H.; Lin, C.; Quijano Cardé, E.M.; Seymour, J.; Siboni, N.; Jones, M.R.L.; et al. Perspective: Something old, something new? Review of wasting and other mortality in Asteroidea (Echinodermata). Front. Mar. Sci. 2019, 6, 406. [Google Scholar] [CrossRef]
- Bucci, C.; Francoeur, M.; McGreal, J.; Smolowitz, R.; Zazueta-Novoa, V.; Wessel, G.M.; Gomez-Chiarri, M. Sea star wasting disease in Asterias forbesi along the Atlantic coast of North America. PLoS ONE 2017, 12, e0188523. [Google Scholar] [CrossRef] [PubMed]
- Hewson, I.; Bistolas, K.S.I.; Quijano Carde, E.M.; Button, J.B.; Foster, P.J.; Flanzenbaum, J.M.; Kocian, J.; Lewis, C.K. Investigating the complex association between viral ecology, environment and Northeast Pacific sea star wasting. Front. Mar. Sci. 2018, 5, 77. [Google Scholar] [CrossRef]
- Jackson, E.W.; Wilhelm, R.C.; Johnson, M.R.; Lutz, H.L.; Danforth, I.; Gaydos, J.K.; Hart, M.W.; Hewson, I. Diversity of sea star-associated densoviruses and transcribed endogenized viral elements of densovirus origin. J. Virol. 2020, 15, 17. [Google Scholar] [CrossRef] [PubMed]
- Jackson, E.W.; Pepe-Ranney, C.; Johnson, M.R.; Distel, D.L.; Hewson, I. A highly prevalent and pervasive densovirus discovered among sea stars from the North American Atlantic coast. Appl. Environ. Microbiol. 2020, 86. [Google Scholar] [CrossRef] [PubMed]
- Aquino, C.A.; Besemer, R.M.; DeRito, C.M.; Kocian, J.; Porter, I.R.; Raimondi, P.T.; Rede, J.E.; Schiebelhut, L.M.; Sparks, J.P.; Wares, J.P.; et al. Evidence that non-pathogenic microorganisms drive sea star wasting disease through boundary layer oxygen diffusion limitation. bioRxiv 2020. [Google Scholar] [CrossRef]
- Aalto, E.A.; Lafferty, K.D.; Sokolow, S.H.; Grewelle, R.E.; Ben-Horin, T.; Boch, C.A.; Raimondi, P.T.; Bograd, S.J.; Hazen, E.L.; Jacox, M.G.; et al. Models with environmental drivers offer a plausible mechanism for the rapid spread of infectious disease outbreaks in marine organisms. Sci. Rep. 2020, 10, 5975. [Google Scholar] [CrossRef]
- Eisenlord, M.E.; Groner, M.L.; Yoshioka, R.M.; Elliott, J.; Maynard, J.; Fradkin, S.; Turner, M.; Pyne, K.; Rivlin, N.; van Hooidonk, R.; et al. Ochre star mortality during the 2014 wasting disease epizootic: Role of population size structure and temperature. Phil. Trans. R. Soc. B 2016, 371, 20150212. [Google Scholar] [CrossRef]
- Menge, B.A.; Cerny-Chipman, E.B.; Johnson, A.; Sullivan, J.; Gravem, S.; Chan, F. Sea star wasting disease in the keystone predator Pisaster ochraceus in Oregon: Insights into differential population impacts, recovery, predation rate, and temperature effects from long-term research. PLoS ONE 2016, 11, e0153994. [Google Scholar]
- Lloyd, M.M.; Pespeni, M.H. Microbiome shifts with onset and progression of Sea Star Wasting Disease revealed through time course sampling. Sci. Rep. 2018, 8, 12. [Google Scholar] [CrossRef] [PubMed]
- Newton, A.L.; Smolowitz, R. Chapter 41—Invertebrates. In Pathology of Wildlife and Zoo Animals; Terio, K.A., McAloose, D., Leger, J.S., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 1019–1052. [Google Scholar]
- Nuñez-Pons, L.; Work, T.M.; Angulo-Preckler, C.; Moles, J.; Avila, C. Exploring the pathology of an epidermal disease affecting a circum-Antarctic sea star. Sci. Rep. 2018, 8, 12. [Google Scholar]
- Thurber, R.V.; Haynes, M.; Breitbart, M.; Wegley, L.; Rohwer, F. Laboratory procedures to generate viral metagenomes. Nat. Protoc. 2009, 4, 470–483. [Google Scholar] [CrossRef] [PubMed]
- Ng, F.F.T.; Wheeler, E.; Greig, D.; Waltzek, T.B.; Gulland, F.; Breitbart, M. Metagenomic identification of a novel anellovirus in Pacific harbor seal (Phoca vitulina richardsii) lung samples and its detection in samples from multiple years. J. Gen. Virol. 2011, 92, 1318–1323. [Google Scholar] [CrossRef]
- Hewson, I.; Johnson, M.R.; Tibbetts, I.R. An unconventional flavivirus and other RNA viruses in the sea cucumber (Holothuroidea; Echinodermata) virome. Viruses 2020, 12, 1057. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Hewson, I. Technical pitfalls that bias comparative microbial community analyses of aquatic disease. Dis. Aquat. Org. 2019, 137, 109–124. [Google Scholar] [CrossRef]
- Kumar, G.; Li, M.; Knyaz, C.; Tamura, K. MEGAX: Molecular evolutionary genetics analysis across computing platforms. Molec. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar]
- Shi, M.; Lin, X.-D.; Tian, J.-H.; Chen, L.-J.; Chen, X.; Li, C.-X.; Qin, X.-C.; Li, J.; Cao, J.-P.; Eden, J.-S.; et al. Redefining the invertebrate RNA virosphere. Nature 2016, 540, 539–543. [Google Scholar] [CrossRef]
- Shi, M.; Lin, X.-D.; Vasilakis, N.; Tian, J.-H.; Li, C.-X.; Chen, L.-J.; Eastwood, G.; Diao, X.-N.; Chen, M.-H.; Chen, X.; et al. Divergent viruses discovered in arthropods and vertebrates revise the evolutionary history of the Flaviviridae and related viruses. J. Virol. 2016, 90, 659. [Google Scholar] [CrossRef] [PubMed]
- Wille, M.; Eden, J.-S.; Shi, M.; Klaassen, M.; Hurt, A.C.; Holmes, E.C. Virus–virus interactions and host ecology are associated with RNA virome structure in wild birds. Mol. Ecol. 2018, 27, 5263–5278. [Google Scholar] [CrossRef] [PubMed]
- Debat, H.J. An RNA virome associated to the golden orb-weaver spider Nephila clavipes. Front. Microbiol. 2017, 8, 2097. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; White, V.L.; Schlub, T.; Eden, J.-S.; Hoffmann, A.A.; Holmes, E.C. No detectable effect of Wolbachia on the prevalence and abundance of the RNA virome of Drosophila melanogaster. Proc. R. Soc. B 2018, 285, 20181165. [Google Scholar] [CrossRef] [PubMed]
- Niu, J.; Li, X.-L.; Wu, Y.-L.; Sun, Q.-Z.; Zhang, W.; Cao, M.; Wang, J.-J. RNA virome screening in diverse but ecologically related citrus pests reveals potential virus-host interactions. J. Invertebr. Pathol. 2020, 170, 107329. [Google Scholar] [CrossRef]
- Ott Rutar, S.; Kordis, D. Analysis of the RNA virome of basal hexapods. PeerJ 2020, 8, e8336. [Google Scholar] [CrossRef]
- Wille, M.; Harvey, E.; Shi, M.; Gonzalez-Acuña, D.; Holmes, E.C.; Hurt, A.C. Sustained RNA virome diversity in Antarctic penguins and their ticks. ISME J. 2020, 14, 1768–1782. [Google Scholar] [CrossRef]
- Pettersson, J.H.O.; Shi, M.; Eden, J.-S.; Holmes, E.C.; Hesson, J.C. Meta-Transcriptomic comparison of the RNA viromes of the mosquito vectors Culex pipiens and Culex torrentium in northern Europe. Viruses 2019, 11, 1033. [Google Scholar] [CrossRef]
- Mahar, J.E.; Shi, M.; Hall, R.N.; Strive, T.; Holmes, E.C. Comparative analysis of RNA virome composition in rabbits and associated ectoparasites. J. Virol. 2020, 94, e02119-19. [Google Scholar] [CrossRef]
- Öhlund, P.; Hayer, J.; Lundén, H.; Hesson, J.C.; Blomström, A.-L. Viromics reveal a number of novel RNA viruses in swedish mosquitoes. Viruses 2019, 11, 1027. [Google Scholar] [CrossRef]
- Bennett, A.J.; Bushmaker, T.; Cameron, K.; Ondzie, A.; Niama, F.R.; Parra, H.-J.; Mombouli, J.-V.; Olson, S.H.; Munster, V.J.; Goldberg, T.L. Diverse RNA viruses of arthropod origin in the blood of fruit bats suggest a link between bat and arthropod viromes. Virology 2019, 528, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Hewson, I.; Bistolas, K.S.I.; Button, J.B.; Jackson, E.W. Occurrence and seasonal dynamics of RNA viral genotypes in three contrasting temperate lakes. PLoS ONE 2018, 13, e0194419. [Google Scholar] [CrossRef] [PubMed]
- Miranda, J.A.; Culley, A.I.; Schvarcz, C.R.; Steward, G.F. RNA viruses as major contributors to Antarctic virioplankton. Environ. Microbiol. 2016, 18, 3714–3727. [Google Scholar] [CrossRef] [PubMed]
- Lang, A.S.; Rise, M.L.; Culley, A.I.; Steward, G.F. RNA viruses in the sea. FEMS Microbiol. Rev. 2009, 33, 295–323. [Google Scholar] [CrossRef]
- Culley, A.I.; Lang, A.S.; Suttle, C.A. The complete genomes of three viruses assembled from shotgun libraries of marine RNA virus communities. Virol. J. 2007, 4, 69. [Google Scholar] [CrossRef]
- Culley, A.I.; Steward, G.F. New genera of RNA viruses in subtropical seawater, inferred from polymerase gene sequences. Appl. Environ. Microbiol. 2007, 73, 5937–5944. [Google Scholar] [CrossRef]
- Culley, A.I.; Lang, A.S.; Suttle, C.A. High diversity of unknown picorna-like viruses in the sea. Nature 2003, 424, 1054–1057. [Google Scholar] [CrossRef]
- Culley, A.I.; Lang, A.S.; Suttle, C.A. Metagenomic analysis of coastal RNA virus communities. Science 2006, 312, 1795–1798. [Google Scholar] [CrossRef]
- Rosani, U.; Gerdol, M. A bioinformatics approach reveals seven nearly-complete RNA-virus genomes in bivalve RNA-seq data. Virus Res. 2017, 239, 33–42. [Google Scholar] [CrossRef]
- Moniruzzaman, M.; Wurch, L.L.; Alexander, H.; Dyhrman, S.T.; Gobler, C.J.; Wilhelm, S.W. Virus-host infection dynamics of marine single-celled eukaryotes resolved from metatranscriptomics. bioRxiv 2016, 093716. [Google Scholar] [CrossRef]
- Sakuna, K.; Elliman, J.; Owens, L. Discovery of a novel Picornavirales, Chequa iflavirus, from stressed redclaw crayfish (Cherax quadricarinatus) from farms in northern Queensland, Australia. Virus Res. 2017, 238, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Lachnit, T.; Thomas, T.; Steinberg, P. Expanding our understanding of the seaweed holobiont: RNA viruses of the red alga Delisea pulchra. Front. Microbiol. 2016, 6, 1489. [Google Scholar] [CrossRef] [PubMed]
- Tai, V.; Lawrence, J.E.; Lang, A.S.; Chan, A.M.; Culley, A.I.; Suttle, C.A. Characterization of HaRNAV, a single-stranded RNA virus causing lysis of Heterosigma akashiwo (Raphidophyceae). J. Phycol. 2003, 39, 343–352. [Google Scholar] [CrossRef]
- Toyoda, K.; Kimura, K.; Osada, K.; Williams, D.M.; Adachi, T.; Yamada, K.; Tomaru, Y. Novel marine diatom ssRNA virus NitRevRNAV infecting Nitzschia reversa. Plant Ecol. Evol. 2019, 152, 178–187. [Google Scholar] [CrossRef]
- Bonami, J.R.; Hasson, K.W.; Mari, J.; Poulos, B.T.; Lightner, D.V. Taura syndrome of marine penaeid shrimp: Characterization of the viral agent. J. Gen. Virol. 1997, 78, 313–319. [Google Scholar] [CrossRef]
- Munday, B.L.; Kwang, J.; Moody, N. Betanodavirus infections of teleost fish: A review. J. Fish Dis. 2002, 25, 127–142. [Google Scholar] [CrossRef]
- Yong, C.Y.; Yeap, S.K.; Omar, A.R.; Tan, W.S. Advances in the study of nodavirus. PeerJ 2017, 5, e3841. [Google Scholar] [CrossRef]
- Jean-Michel, A.; François, H.; Donald, V.L.; Rita, M.R.; Jocelyne, M.; Jean-Robert, B. A viral disease associated with mortalities in hatchery-reared postlarvae of the giant freshwater prawn Macrobrachium rosenbergii. Dis. Aquat. Org. 1999, 38, 177–181. [Google Scholar]
- Gomez, D.K.; Baeck, G.W.; Kim, J.H.; Choresca, C.H.; Park, S.C. Molecular detection of betanodaviruses from apparently healthy wild marine invertebrates. J. Invertebr. Pathol. 2008, 97, 197–202. [Google Scholar] [CrossRef]
- Gomez, D.K.; Lim, D.J.; Baeck, G.W.; Youn, H.J.; Shin, N.S.; Youn, H.Y.; Hwang, C.Y.; Park, J.H.; Park, S.C. Detection of betanodaviruses in apparently healthy aquarium fishes and invertebrates. J. Vet. Sci. 2006, 7, 369–374. [Google Scholar] [CrossRef]
- Ng, T.F.F.; Alavandi, S.; Varsani, A.; Burghart, S.; Breitbart, M. Metagenomic identification of a nodavirus and a circular ssDNA virus in semi-purified viral nucleic acids from the hepatopancreas of healthy Farfantepenaeus duorarum shrimp. Dis. Aquat. Org. 2013, 105, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Gomez, D.K.; Baeck, G.W.; Kim, J.H.; Choresca, C.H.; Park, S.C. Genetic analysis of betanodaviruses in subclinically infected aquarium fish and invertebrates. Curr. Microbiol. 2008, 56, 499–504. [Google Scholar] [CrossRef] [PubMed]
- Bergoin, M.; Tijssen, P. Molecular biology of Densovirinae. Contrib. Microbiol. 2000, 4, 12–32. [Google Scholar] [PubMed]
- Tijssen, P.; Bergoin, M. Densonucleosis viruses constitute an increasingly diversified subfamily among the parvoviruses. Semin. Virol. 1995, 6, 347–355. [Google Scholar] [CrossRef]
- Kang, Y.J.; Huang, W.; Zhao, A.L.; Lai, D.D.; Shao, L.; Shen, Y.Q.; Deng, X.; Delwart, E.; Zhang, W. Densoviruses in oyster Crassostrea ariakensis. Arch. Virol. 2017, 162, 2153–2157. [Google Scholar] [CrossRef]
- Richard, J.C.; Leis, E.; Dunn, C.D.; Agbalog, R.; Waller, D.; Knowles, S.; Putnam, J.; Goldberg, T.L. Mass mortality in freshwater mussels (Actinonaias pectorosa) in the Clinch River, USA, linked to a novel densovirus. Sci. Rep. 2020, 10, 14498. [Google Scholar] [CrossRef]
- Gudenkauf, B.M.; Hewson, I. Comparative metagenomics of viral assemblages inhabiting four phyla of marine invertebrates. Front. Mar. Sci. 2016, 3, 23. [Google Scholar] [CrossRef]
- Bochow, S.; Condon, K.; Elliman, J.; Owens, L. First complete genome of an Ambidensovirus; Cherax quadricarinatus densovirus, from freshwater crayfish Cherax quadricarinatus. Mar. Genom. 2015, 24, 305–312. [Google Scholar] [CrossRef]
- Fargette, D.; Pinel, A.; Abubakar, Z.; Traoré, O.; Brugidou, C.; Fatogoma, S.; Hébrard, E.; Choisy, M.; Séré, Y.; Fauquet, C.; et al. Inferring the evolutionary history of Rice Yellow Mottle Virus from genomic, phylogenetic, and phylogeographic studies. J. Virol. 2004, 78, 3252–3261. [Google Scholar] [CrossRef]
- Tamm, T.; Truve, E. Sobemoviruses. J. Virol. 2000, 74, 6231–6241. [Google Scholar] [CrossRef]
- Laffy, P.W.; Botté, E.S.; Wood-Charlson, E.M.; Weynberg, K.D.; Rattei, T.; Webster, N.S. Thermal stress modifies the marine sponge virome. Environ. Microbiol. Rep. 2019, 11, 690–698. [Google Scholar] [CrossRef] [PubMed]
- Grasis, J.A.; Lachnit, T.; Anton-Erxleben, F.; Lim, Y.W.; Schmieder, R.; Fraune, S.; Franzenburg, S.; Insua, S.; Machado, G.; Haynes, M.; et al. Species-specific viromes in the ancestral holobiont Hydra. PLoS ONE 2014, 9, e109952. [Google Scholar] [CrossRef] [PubMed]
- Thurber, R.L.V.; Barott, K.L.; Hall, D.; Liu, H.; Rodriguez-Mueller, B.; Desnues, C.; Edwards, R.A.; Haynes, M.; Angly, F.E.; Wegley, L.; et al. Metagenomic analysis indicates that stressors induce production of herpes-like viruses in the coral Porites compressa. Proc. Natl. Acad. Sci. USA 2008, 105, 18413–18418. [Google Scholar] [CrossRef] [PubMed]
- Frakolaki, E.; Kaimou, P.; Moraiti, M.; Kalliampakou, K.I.; Karampetsou, K.; Dotsika, E.; Liakos, P.; Vassilacopoulou, D.; Mavromara, P.; Bartenschlager, R.; et al. The role of tissue oxygen tension in Dengue virus replication. Cells 2018, 7, 241. [Google Scholar] [CrossRef]
- Moon, E.J.; Jeong, C.H.; Jeong, J.W.; Kim, K.R.; Yu, D.Y.; Murakami, S.; Kim, C.W.; Kim, K.W. Hepatitis B virus X protein induces angiogenesis by stabilizing hypoxia-inducible factor-1alpha. FASEB J. 2004, 18, 382–384. [Google Scholar] [CrossRef]
- Wakisaka, N.; Kondo, S.; Yoshizaki, T.; Murono, S.; Furukawa, M.; Pagano, J.S. Epstein-Barr virus latent membrane protein 1 induces synthesis of hypoxia-inducible factor 1α. Mol. Cell. Biol. 2004, 24, 5223–5234. [Google Scholar] [CrossRef]
- Ruan, H.; Su, H.; Hu, L.; Lamborn, K.R.; Kan, Y.W.; Deen, D.F. A hypoxia-regulated adeno-associated virus vector for cancer-specific gene therapy. Neoplasia 2001, 3, 255–263. [Google Scholar] [CrossRef][Green Version]
- Aghi, M.K.; Liu, T.-C.; Rabkin, S.; Martuza, R.L. Hypoxia enhances the replication of oncolytic Herpes Simplex virus. Mol. Ther. 2009, 17, 51–56. [Google Scholar] [CrossRef]
- Jiang, J.-H.; Wang, N.; Li, A.; Liao, W.-T.; Pan, Z.-G.; Mai, S.-J.; Li, D.-J.; Zeng, M.-S.; Wen, J.-M.; Zeng, Y.-X. Hypoxia can contribute to the induction of the Epstein-Barr virus (EBV) lytic cycle. J. Clin. Virol. 2006, 37, 98–103. [Google Scholar] [CrossRef]
- Gan, E.S.; Cheong, W.F.; Chan, K.R.; Ong, E.Z.; Chai, X.; Tan, H.C.; Ghosh, S.; Wenk, M.R.; Ooi, E.E. Hypoxia enhances antibody-dependent dengue virus infection. EMBO J. 2017, 36, 1348–1363. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Library Name | Specimen Name | Date | Tissue Type | Library Size (Reads) | Reads Matching Viruses | BioSample Accession |
|---|---|---|---|---|---|---|
| SC1 | SSWD-affected 1 | 22 July 2018 | Control | 3,658,045 | 554,809 | SRR11931187 |
| SC2 | SSWD-affected 1 | 26 July 2018 | Scar | 4,099,676 | 582,091 | SRR11931186 |
| SC3 | SSWD-affected 1 | 26 July 2018 | Control | 3,592,838 | 597,395 | SRR11931179 |
| SC4 | SSWD-affected 1 | 26 July 2018 | Lesion | 3,255,385 | 169,582 | SRR11931178 |
| SC5 | SSWD-affected 1 | 27 July 2018 | Lesion | 3,443,200 | 871,889 | SRR11931177 |
| SC6 | SSWD-affected 2 | 22 July 2018 | Control | 4,948,712 | 1,007,543 | SRR11931176 |
| SC7 | SSWD-affected 2 | 4 August 2018 | Control | 2,703,665 | 624,989 | SRR11931175 |
| SC8 | SSWD-affected 2 | 4 August 2018 | Scar | 2,797,069 | 507,994 | SRR11931174 |
| SC9 | SSWD-affected 2 | 4 August 20188 | Lesion | 3,678,615 | 1,306,746 | SRR11931173 |
| SC10 | SSWD-affected 2 | 6 August 2018 | Lesion | 3,259,328 | 924,368 | SRR11931172 |
| SC11 | Grossly Normal 1 | 22 July 2018 | Control | 2,954,408 | 2349 | SRR11931185 |
| SC12 | Grossly Normal 1 | 9 August 2018 | Control | 1,919,780 | 2694 | SRR11931184 |
| SC13 | Grossly Normal 1 | 9 August 2018 | Scar | 1,939,732 | 3134 | SRR11931183 |
| SC14 | Grossly Normal 2 | 22 July 2018 | Control | 3,273,980 | 3851 | SRR11931182 |
| SC15 | Grossly Normal 2 | 9 August 2018 | Control | 1,595,206 | 1112 | SRR11931181 |
| SC16 | Grossly Normal 2 | 9 August 2018 | Scar | 1,196,532 | 882 | SRR11931180 |
| Contig # | p-Value | ||
|---|---|---|---|
| Condition | Time | Sample Type | |
| Contig 2401 | ns | 0.023 | Ns |
| Contig 12092 | 0.001 | ns | Ns |
| Contig 12093 | 0.003 | 0.022 | Ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hewson, I.; Aquino, C.A.; DeRito, C.M. Virome Variation during Sea Star Wasting Disease Progression in Pisaster ochraceus (Asteroidea, Echinodermata). Viruses 2020, 12, 1332. https://doi.org/10.3390/v12111332
Hewson I, Aquino CA, DeRito CM. Virome Variation during Sea Star Wasting Disease Progression in Pisaster ochraceus (Asteroidea, Echinodermata). Viruses. 2020; 12(11):1332. https://doi.org/10.3390/v12111332
Chicago/Turabian StyleHewson, Ian, Citlalli A. Aquino, and Christopher M. DeRito. 2020. "Virome Variation during Sea Star Wasting Disease Progression in Pisaster ochraceus (Asteroidea, Echinodermata)" Viruses 12, no. 11: 1332. https://doi.org/10.3390/v12111332
APA StyleHewson, I., Aquino, C. A., & DeRito, C. M. (2020). Virome Variation during Sea Star Wasting Disease Progression in Pisaster ochraceus (Asteroidea, Echinodermata). Viruses, 12(11), 1332. https://doi.org/10.3390/v12111332