Loop-Mediated Isothermal Amplification (LAMP) for the Diagnosis of Zika Virus: A Review


Abstract
1. Introduction
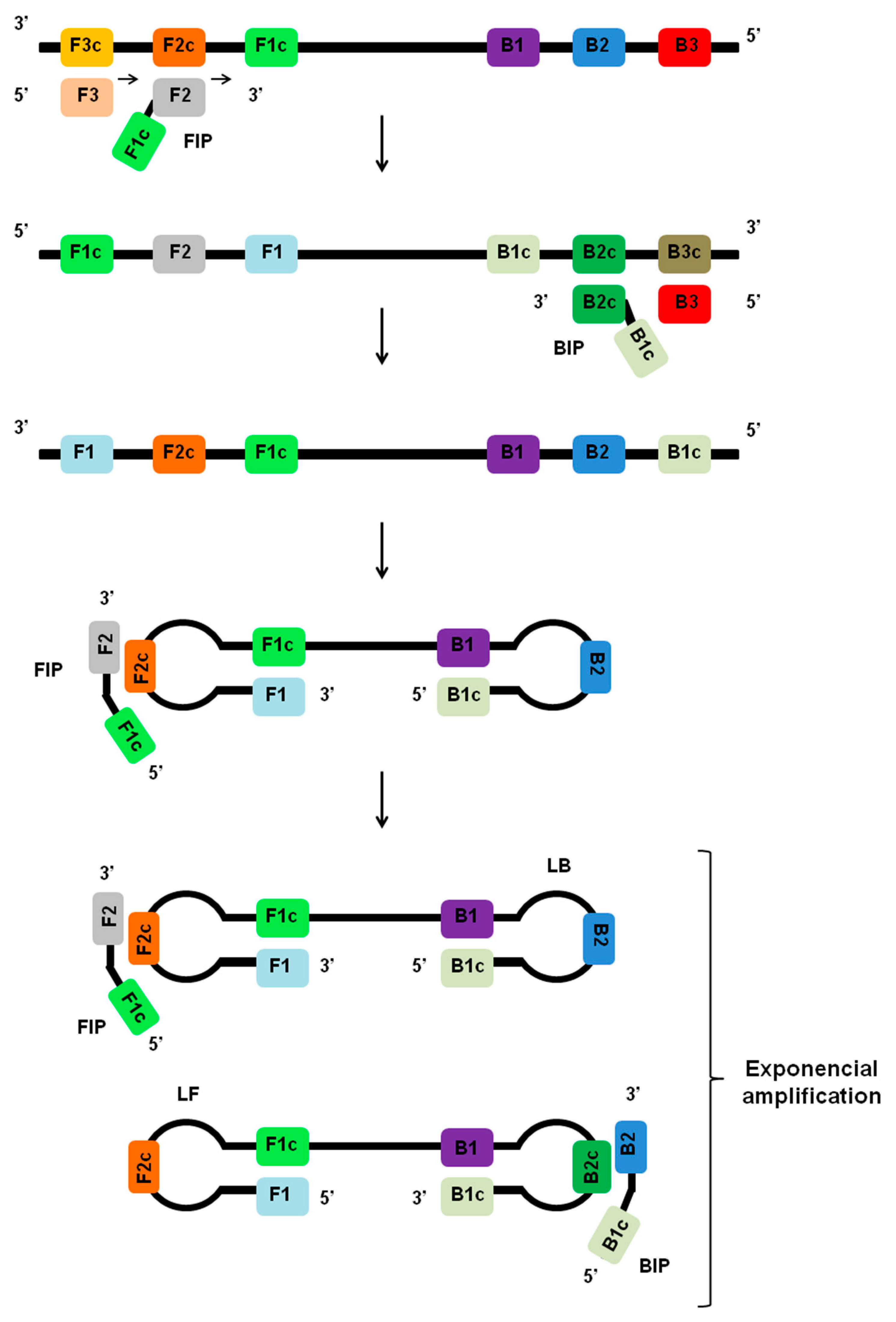
2. Principles of LAMP Assay
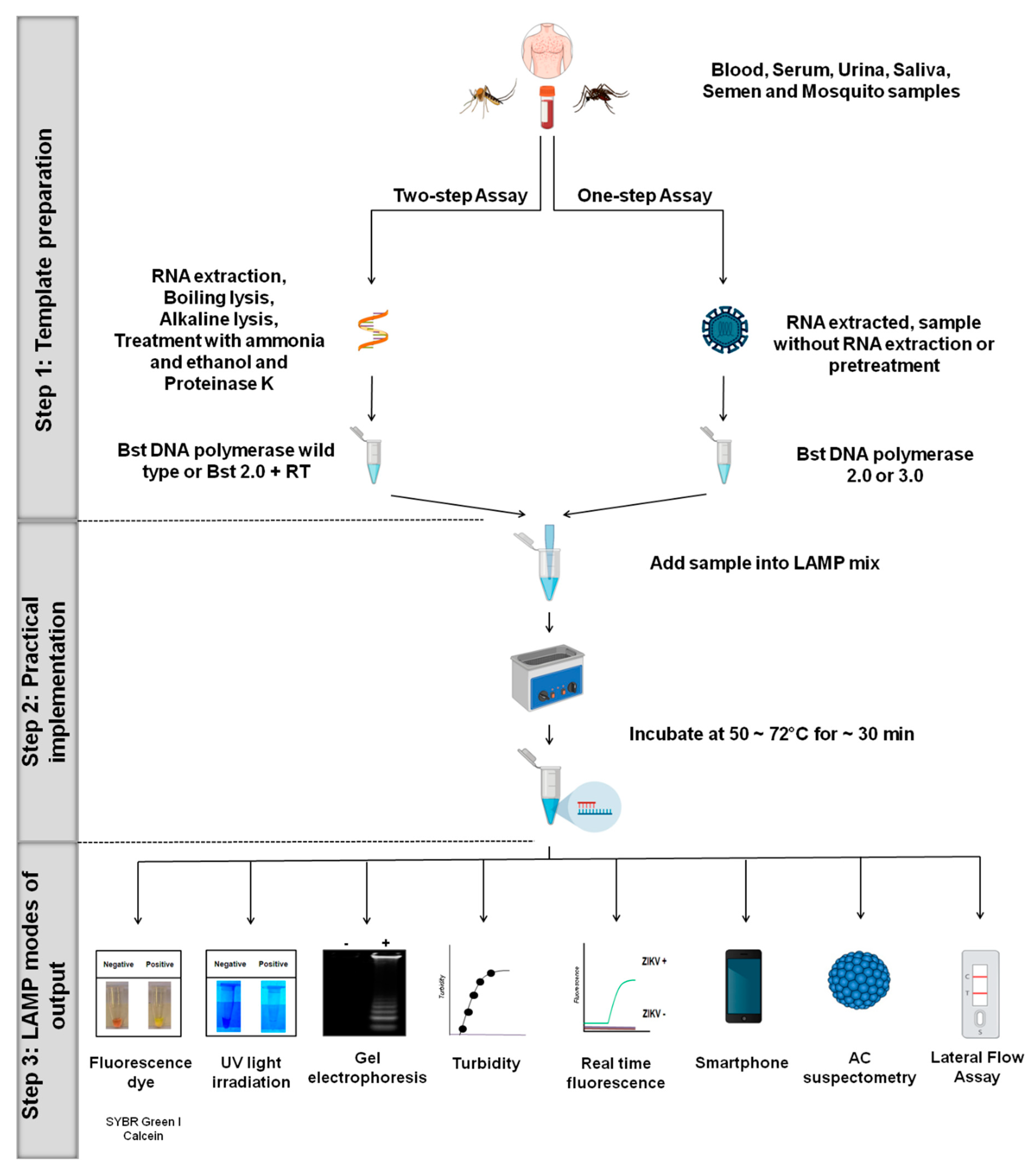
3. Principles LAMP Platforms for ZIKV
3.1. Step 1: Template Preparation
3.2. Step 2: Practical Implementation
3.3. Step 3: LAMP Modes of Output
4. LAMP Specificity
5. LAMP Sensitivity
6. LAMP Assay Evaluation Using Clinical Samples
7. LAMP for Detection of ZIKV in Mosquito Samples
8. Multiplexing
9. Clinical Application and Public Health Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Haddow, A.D.; Schuh, A.J.; Yasuda, C.Y.; Kasper, M.R.; Heang, V.; Huy, R.; Guzman, H.; Tesh, R.B.; Weaver, S.C. Genetic characterization of Zika virus strains: Geographic expansion of the Asian lineage. PLoS Negl. Trop. Dis. 2012, 6, e1477. [Google Scholar] [CrossRef]
- Faye, O.; Freire, C.C.; Iamarino, A.; de Oliveira, J.V.; Diallo, M.; Zanotto, P.M.; Sall, A.A. Molecular evolution of Zika virus during its emergence in the 20(th) century. PLoS Negl. Trop. Dis. 2014, 8, e2636. [Google Scholar] [CrossRef]
- Petersen, L.R.; Jamieson, D.J.; Powers, A.M.; Honein, M.A. Zika Virus. N. Engl. J. Med. 2016, 374, 1552–1563. [Google Scholar] [CrossRef]
- Chambers, T.J.; Hahn, C.S.; Galler, R.; Rice, C.M. Flavivirus genome organization, expression, and replication. Ann. Rev. Microbiol. 1990, 44, 649–688. [Google Scholar] [CrossRef]
- Kuno, G.; Chang, G.J. Full-length sequencing and genomic characterization of Bagaza, Kedougou, and Zika viruses. Arch. Virol. 2007, 152, 687–696. [Google Scholar] [CrossRef]
- Lanciotti, R.S.; Kosoy, O.L.; Laven, J.J.; Velez, J.O.; Lambert, A.J.; Johnson, A.J.; Stanfield, S.M.; Duffy, M.R. Genetic and serologic properties of Zika virus associated with an epidemic, Yap State, Micronesia, 2007. Emerg. Infect. Dis. 2008, 14, 1232–1239. [Google Scholar] [CrossRef]
- Waggoner, J.J.; Pinsky, B.A. Zika Virus: Diagnostics for an Emerging Pandemic Threat. J. Clin. Microbiol. 2016, 54, 860–867. [Google Scholar] [CrossRef]
- Dick, G.W.; Kitchen, S.F.; Haddow, A.J. Zika virus. I. Isolations and serological specificity. Trans. R Soc. Trop. Med. Hyg. 1952, 46, 509–520. [Google Scholar] [CrossRef]
- Ioos, S.; Mallet, H.P.; Leparc Goffart, I.; Gauthier, V.; Cardoso, T.; Herida, M. Current Zika virus epidemiology and recent epidemics. Med. Mal. Infect. 2014, 44, 302–307. [Google Scholar] [CrossRef]
- Musso, D. Zika Virus Transmission from French Polynesia to Brazil. Emerg. Infect. Dis. 2015, 21, 1887. [Google Scholar] [CrossRef]
- Gatherer, D.; Kohl, A. Zika virus: A previously slow pandemic spreads rapidly through the Americas. J. Gen. Virol. 2016, 97, 269–273. [Google Scholar] [CrossRef]
- Oehler, E.; Watrin, L.; Larre, P.; Leparc-Goffart, I.; Lastere, S.; Valour, F.; Baudouin, L.; Mallet, H.; Musso, D.; Ghawche, F. Zika virus infection complicated by Guillain-Barre syndrome—Case report, French Polynesia, December 2013. Euro surveillance: Bulletin Europeen sur les maladies transmissibles = European communicable disease bulletin. Eurosurveillance 2014, 19, 20720. [Google Scholar] [CrossRef]
- Campos, G.S.; Bandeira, A.C.; Sardi, S.I. Zika Virus Outbreak, Bahia, Brazil. Emerg. Infect. Dis. 2015, 21, 1885–1886. [Google Scholar] [CrossRef]
- Campos, T.L.; Duraes-Carvalho, R.; Rezende, A.M.; de Carvalho, O.V.; Kohl, A.; Wallau, G.L.; Pena, L.J. Revisiting Key Entry Routes of Human Epidemic Arboviruses into the Mainland Americas through Large-Scale Phylogenomics. Int. J. Genom. 2018, 2018, 6941735. [Google Scholar] [CrossRef]
- Rasmussen, S.A.; Jamieson, D.J.; Honein, M.A.; Petersen, L.R. Zika Virus and Birth Defects--Reviewing the Evidence for Causality. N. Engl. J. Med. 2016, 374, 1981–1987. [Google Scholar] [CrossRef]
- Meneses, J.D.A.; Ishigami, A.C.; de Mello, L.M.; de Albuquerque, L.L.; de Brito, C.A.A.; Cordeiro, M.T.; Pena, L.J. Lessons Learned at the Epicenter of Brazil’s Congenital Zika Epidemic: Evidence From 87 Confirmed Cases. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2017, 64, 1302–1308. [Google Scholar] [CrossRef]
- Diallo, D.; Sall, A.A.; Diagne, C.T.; Faye, O.; Ba, Y.; Hanley, K.A.; Buenemann, M.; Weaver, S.C.; Diallo, M. Zika virus emergence in mosquitoes in southeastern Senegal, 2011. PLoS ONE 2014, 9, e109442. [Google Scholar] [CrossRef]
- Guo, X.X.; Li, C.X.; Deng, Y.Q.; Xing, D.; Liu, Q.M.; Wu, Q.; Sun, A.J.; Dong, Y.D.; Cao, W.C.; Qin, C.F.; et al. Culex pipiens quinquefasciatus: A potential vector to transmit Zika virus. Emerging Microbes Infect. 2016, 5, e102. [Google Scholar] [CrossRef]
- Guedes, D.R.; Paiva, M.H.; Donato, M.M.; Barbosa, P.P.; Krokovsky, L.; Rocha, S.W.D.S.; Saraiva, K.; Crespo, M.M.; Rezende, T.M.; Wallau, G.L.; et al. Zika virus replication in the mosquito Culex quinquefasciatus in Brazil. Emerg. Microbes Infect. 2017, 6, e69. [Google Scholar] [CrossRef]
- Elizondo-Quiroga, D.; Medina-Sánchez, A.; Sánchez-González, J.M.; Eckert, K.A.; Villalobos-Sánchez, E.; Navarro-Zúñiga, A.R.; Sánchez-Tejeda, G.; Correa-Morales, F.; González-Acosta, C.; Arias, C.F.; et al. Author Correction: Zika Virus in Salivary Glands of Five Different Species of Wild-Caught Mosquitoes from Mexico. Sci. Rep. 2018, 8, 7887. [Google Scholar] [CrossRef]
- Smartt, C.T.; Shin, D.; Kang, S.; Tabachnick, W.J. (Diptera: Culicidae) From Florida Transmitted Zika Virus. Front. Microbiol. 2018, 9, 768. [Google Scholar] [CrossRef]
- Musso, D.; Nhan, T.; Robin, E.; Roche, C.; Bierlaire, D.; Zisou, K.; Shan Yan, A.; Cao-Lormeau, V.M.; Broult, J. Potential for Zika virus transmission through blood transfusion demonstrated during an outbreak in French Polynesia, November 2013 to February 2014. Euro surveillance: Bulletin Europeen sur les maladies transmissibles = European communicable disease bulletin. Eurosurveillance 2014, 19, 20761. [Google Scholar]
- Cunha, M.S.; Esposito, D.L.; Rocco, I.M.; Maeda, A.Y.; Vasami, F.G.; Nogueira, J.S.; de Souza, R.P.; Suzuki, A.; Addas-Carvalho, M.; Barjas-Castro, M.E.L.; et al. First Complete Genome Sequence of Zika Virus (Flaviviridae, Flavivirus) from an Autochthonous Transmission in Brazil. Genome Announc. 2016, 4, e00032-16. [Google Scholar] [CrossRef]
- Hills, S.L.; Russell, K.; Hennessey, M.; Williams, C.; Oster, A.M.; Fischer, M.; Mead, P. Transmission of Zika Virus Through Sexual Contact with Travelers to Areas of Ongoing Transmission—Continental United States, 2016. MMWR Morb. Mortal. Wkly. Rep. 2016, 65, 215–216. [Google Scholar] [CrossRef]
- Faria, N.R.; Azevedo, R.D.S.D.; Kraemer, M.U.G.; Souza, R.; Cunha, M.S.; Hill, S.C.; Thézé, J.; Bonsall, M.B.; Bowden, T.A.; Rissanen, I.; et al. Zika virus in the Americas: Early epidemiological and genetic findings. Science 2016, 352, 345–349. [Google Scholar] [CrossRef]
- Wikan, N.; Smith, D.R. Zika virus: History of a newly emerging arbovirus. Lancet Infect. Dis. 2016, 16, e119–e126. [Google Scholar] [CrossRef]
- Yaren, O.; Alto, B.W.; Gangodkar, P.V.; Ranade, S.R.; Patil, K.N.; Bradley, K.M.; Yang, Z.; Phadke, N.; Benner, S.A. Point of sampling detection of Zika virus within a multiplexed kit capable of detecting dengue and chikungunya. BMC Infect. Dis. 2017, 17, 293. [Google Scholar] [CrossRef]
- Kurosaki, Y.; Martins, D.B.G.; Kimura, M.; Catena, A.D.S.; Borba, M.A.C.S.; Mattos, S.D.S.; Abe, H.; Yoshikawa, R.; de Lima Filho, J.L.; Yasuda, J. Development and evaluation of a rapid molecular diagnostic test for Zika virus infection by reverse transcription loop-mediated isothermal amplification. Sci. Rep. 2017, 7, 13503. [Google Scholar] [CrossRef]
- Faye, O.; Diallo, D.; Diallo, M.; Weidmann, M.; Sall, A.A. Quantitative real-time PCR detection of Zika virus and evaluation with field-caught mosquitoes. Virol. J. 2013, 10, 311. [Google Scholar] [CrossRef]
- Abd El Wahed, A.; Sanabani, S.S.; Faye, O.; Pessôa, R.; Patriota, J.V.; Giorgi, R.R.; Patel, P.; Böhlken-Fascher, S.; Landt, O.; Niedrig, M.; et al. Rapid Molecular Detection of Zika Virus in Acute-Phase Urine Samples Using the Recombinase Polymerase Amplification Assay. PLoS Curr. 2017, 9. [Google Scholar] [CrossRef]
- Nicolini, A.M.; McCracken, K.E.; Yoon, J.Y. Future developments in biosensors for field-ready Zika virus diagnostics. J. Biol. Eng. 2017, 11, 7. [Google Scholar] [CrossRef]
- Meagher, R.J.; Negrete, O.A.; Van Rompay, K.K. Engineering Paper-Based Sensors for Zika Virus. Trends Mol. Med. 2016, 22, 529–530. [Google Scholar] [CrossRef]
- Felix, A.C.; Souza, N.C.S.; Figueiredo, W.M.; Costa, A.A.; Inenami, M.; da Silva, R.M.G.; Levi, J.E.; Pannuti, C.S.; Romano, C.M. Cross reactivity of commercial anti-dengue immunoassays in patients with acute Zika virus infection. J. Med. Virol. 2017, 89, 1477–1479. [Google Scholar] [CrossRef]
- Notomi, T.; Okayama, H.; Masubuchi, H.; Yonekawa, T.; Watanabe, K.; Amino, N.; Hase, T. Loop-mediated isothermal amplification of DNA. Nucleic Acids Res. 2000, 28, E63. [Google Scholar] [CrossRef]
- Nemoto, M.; Imagawa, H.; Tsujimura, K.; Yamanaka, T.; Kondo, T.; Matsumura, T. Detection of equine rotavirus by reverse transcription loop-mediated isothermal amplification (RT-LAMP). J. Vet. Med. Sci. 2010, 72, 823–826. [Google Scholar] [CrossRef]
- Notomi, T.; Mori, Y.; Tomita, N.; Kanda, H. Loop-mediated isothermal amplification (LAMP): Principle, features, and future prospects. J. Microbiol. 2015, 53, 1–5. [Google Scholar] [CrossRef]
- Song, J.; Mauk, M.G.; Hackett, B.A.; Cherry, S.; Bau, H.H.; Liu, C. Instrument-Free Point-of-Care Molecular Detection of Zika Virus. Anal. Chem. 2016, 88, 7289–7294. [Google Scholar] [CrossRef]
- Tian, B.; Qiu, Z.; Ma, J.; Zardán Gómez de la Torre, T.; Johansson, C.; Svedlindh, P.; Strömberg, M. Attomolar Zika virus oligonucleotide detection based on loop-mediated isothermal amplification and AC susceptometry. Biosens. Bioelectron. 2016, 86, 420–425. [Google Scholar] [CrossRef]
- Wang, X.; Yin, F.; Bi, Y.; Cheng, G.; Li, J.; Hou, L.; Li, Y.; Yang, B.; Liu, W.; Yang, L. Rapid and sensitive detection of Zika virus by reverse transcription loop-mediated isothermal amplification. J. Virol Methods 2016, 238, 86–93. [Google Scholar] [CrossRef]
- Lee, D.; Shin, Y.; Chung, S.; Hwang, K.S.; Yoon, D.S.; Lee, J.H. Simple and Highly Sensitive Molecular Diagnosis of Zika Virus by Lateral Flow Assays. Anal. Chem. 2016, 88, 12272–12278. [Google Scholar] [CrossRef]
- Chotiwan, N.; Brewster, C.D.; Magalhaes, T.; Weger-Lucarelli, J.; Duggal, N.K.; Rückert, C.; Nguyen, C.; Garcia Luna, S.M.; Fauver, J.R.; Andre, B.; et al. Rapid and specific detection of Asian- and African-lineage Zika viruses. Sci. Transl. Med. 2017, 9, eaag0538. [Google Scholar] [CrossRef]
- Lamb, L.E.; Bartolone, S.N.; Tree, M.O.; Conway, M.J.; Rossignol, J.; Smith, C.P.; Chancellor, M.B. Rapid Detection of Zika Virus in Urine Samples and Infected Mosquitos by Reverse Transcription-Loop-Mediated Isothermal Amplification. Sci. Rep. 2018, 8, 3803. [Google Scholar] [CrossRef]
- Zhao, J.; Feng, R. Sensitive and rapid detection of Zika virus by loop-mediated isothermal amplification. Virus Genes 2018, 55, 43–50. [Google Scholar] [CrossRef]
- Kaarj, K.; Akarapipad, P.; Yoon, J.Y. Simpler, Faster, and Sensitive Zika Virus Assay Using Smartphone Detection of Loop-mediated Isothermal Amplification on Paper Microfluidic Chips. Sci. Rep. 2018, 8, 12438. [Google Scholar] [CrossRef]
- Bui, T.T.; Moi, M.L.; Morita, K.; Hasebe, F. Development of a universal and lineage-specific primer sets for Zika virus (ZIKV) rapid detection in blood and urine samples by using one-step reverse transcription loop-mediated isothermal amplification (RT-LAMP). JPN J. Infect. Dis. 2019. [Google Scholar] [CrossRef]
- Wong, Y.P.; Othman, S.; Lau, Y.L.; Radu, S.; Chee, H.Y. Loop-mediated isothermal amplification (LAMP): A versatile technique for detection of micro-organisms. J. Appl. Microbiol. 2018, 124, 626–643. [Google Scholar] [CrossRef]
- Salinas, N.R.; Little, D.P. Electric LAMP: Virtual Loop-Mediated Isothermal AMPlification. ISRN Bioinform. 2012, 2012, 696758. [Google Scholar] [CrossRef]
- Santiago-Felipe, S.; Tortajada-Genaro, L.A.; Carrascosa, J.; Puchades, R.; Maquieira, Á. Real-time loop-mediated isothermal DNA amplification in compact disc micro-reactors. Biosens. Bioelectron. 2016, 79, 300–306. [Google Scholar] [CrossRef]
- Balasuriya, U.B.; Lee, P.Y.; Tiwari, A.; Skillman, A.; Nam, B.; Chambers, T.M.; Tsai, Y.L.; Ma, L.J.; Yang, P.C.; Chang, H.F.; et al. Rapid detection of equine influenza virus H3N8 subtype by insulated isothermal RT-PCR (iiRT-PCR) assay using the POCKIT™ Nucleic Acid Analyzer. J. Virol. Methods 2014, 207, 66–72. [Google Scholar] [CrossRef]
- Lau, Y.L.; Lai, M.Y.; Teoh, B.T.; Abd-Jamil, J.; Johari, J.; Sam, S.S.; Tan, K.K.; AbuBakar, S. Colorimetric Detection of Dengue by Single Tube Reverse-Transcription-Loop-Mediated Isothermal Amplification. PLoS ONE 2015, 10, e0138694. [Google Scholar] [CrossRef]
- Lopez-Jimena, B.; Bekaert, M.; Bakheit, M.; Frischmann, S.; Patel, P.; Simon-Loriere, E.; Lambrechts, L.; Duong, V.; Dussart, P.; Harold, G.; et al. Development and validation of four one-step real-time RT-LAMP assays for specific detection of each dengue virus serotype. PLoS Negl. Trop. Dis. 2018, 12, e0006381. [Google Scholar] [CrossRef]
- Lopez-Jimena, B.; Wehner, S.; Harold, G.; Bakheit, M.; Frischmann, S.; Bekaert, M.; Faye, O.; Sall, A.A.; Weidmann, M. Development of a single-tube one-step RT-LAMP assay to detect the Chikungunya virus genome. PLoS Negl. Trop. Dis. 2018, 12, e0006448. [Google Scholar] [CrossRef]
- Silva, S.J.R.D.; Paiva, M.H.S.; Guedes, D.R.D.; Krokovsky, L.; Melo, F.L.; Silva, M.A.L.D.; Silva, A.D.; Ayres, C.F.J.; Pena, L.J. Development and Validation of Reverse Transcription Loop-Mediated Isothermal Amplification (RT-LAMP) for Rapid Detection of ZIKV in Mosquito Samples from Brazil. Sci. Rep. 2019, 9, 4494. [Google Scholar] [CrossRef]
- Chen, H.W.; Weissenberger, G.; Atkins, E.; Chao, C.C.; Suputtamongkol, Y.; Ching, W.M. Highly Sensitive Loop-Mediated Isothermal Amplification for the Detection of Leptospira. Int. J. Bacteriol. 2015, 2015, 147173. [Google Scholar] [CrossRef]
- Chen, H.W.; Weissenberger, G.; Ching, W.M. Development of Lyophilized Loop-Mediated Isothermal Amplification Reagents for the Detection of Leptospira. Mil. Med. 2016, 181 (Suppl. 5), 227–231. [Google Scholar] [CrossRef]
- Nagamine, K.; Hase, T.; Notomi, T. Accelerated reaction by loop-mediated isothermal amplification using loop primers. Mol. Cell Probes 2002, 16, 223–229. [Google Scholar] [CrossRef]
- Guo, X.G.; Zhou, Y.Z.; Li, Q.; Wang, W.; Wen, J.Z.; Zheng, L.; Wang, Q. Rapid and reliable diagnostic method to detect Zika virus by real-time fluorescence reverse transcription loop-mediated isothermal amplification. AMB Express 2018, 8, 60. [Google Scholar] [CrossRef]
- Priye, A.; Bird, S.W.; Light, Y.K.; Ball, C.S.; Negrete, O.A.; Meagher, R.J. A smartphone-based diagnostic platform for rapid detection of Zika, chikungunya, and dengue viruses. Sci. Rep. 2017, 7, 44778. [Google Scholar] [CrossRef]
- Calvert, A.E.; Biggerstaff, B.J.; Tanner, N.A.; Lauterbach, M.; Lanciotti, R.S. Rapid colorimetric detection of Zika virus from serum and urine specimens by reverse transcription loop-mediated isothermal amplification (RT-LAMP). PLoS ONE 2017, 12, e0185340. [Google Scholar] [CrossRef]
- Ganguli, A.; Ornob, A.; Yu, H.; Damhorst, G.L.; Chen, W.; Sun, F.; Bhuiya, A.; Cunningham, B.T.; Bashir, R. Hands-free smartphone-based diagnostics for simultaneous detection of Zika, Chikungunya, and Dengue at point-of-care. Biomed. Microdevices 2017, 19, 73. [Google Scholar] [CrossRef]
- Castro, T.; Sabalza, M.; Barber, C.; Abrams, W.; Da Costa, A.C.; De Pádua Milagres, F.A.; Braz-Silva, P.H.; Malamud, D.; Gallottini, M. Rapid diagnosis of Zika virus through saliva and urine by Loop-mediated isothermal amplification (LAMP). J. Oral Microbiol. 2018, 10, 1510712. [Google Scholar] [CrossRef]
- Bartolone, S.N.; Tree, M.O.; Conway, M.J.; Chancellor, M.B.; Lamb, L.E. Reverse Transcription-Loop-mediated Isothermal Amplification (RT-LAMP) Assay for Zika Virus and Housekeeping Genes in Urine, Serum, and Mosquito Samples. J. Vis. Exp. 2018, 139, e58436. [Google Scholar] [CrossRef]
- Sabalza, M.; Yasmin, R.; Barber, C.A.; Castro, T.; Malamud, D.; Kim, B.J.; Zhu, H.; Montagna, R.A.; Abrams, W.R. Detection of Zika virus using reverse-transcription LAMP coupled with reverse dot blot analysis in saliva. PLoS ONE 2018, 13, e0192398. [Google Scholar] [CrossRef]
- Neves Estrela, P.F.; de Melo Mendes, G.; de Oliveira, K.G.; Bailã, A.M.; de Almeida Soares, C.M.; Assunção, N.A.; Mendes Duarte, G.R. Ten-minute direct detection of Zika virus in serum samples by RT-LAMP. J. Virol. Methods 2019, 113675. [Google Scholar] [CrossRef]
- Chander, Y.; Koelbl, J.; Puckett, J.; Moser, M.J.; Klingele, A.J.; Liles, M.R.; Carrias, A.; Mead, D.A.; Schoenfeld, T.W. A novel thermostable polymerase for RNA and DNA loop-mediated isothermal amplification (LAMP). Front. Microbiol. 2014, 5, 395. [Google Scholar] [CrossRef]
- Poole, C.B.; Tanner, N.A.; Zhang, Y.; Evans, T.C.; Carlow, C.K. Diagnosis of brugian filariasis by loop-mediated isothermal amplification. PLoS Negl. Trop. Dis. 2012, 6, e1948. [Google Scholar] [CrossRef]
- Tanner, N.A.; Zhang, Y.; Evans, T.C. Simultaneous multiple target detection in real-time loop-mediated isothermal amplification. Biotechniques 2012, 53, 81–89. [Google Scholar] [CrossRef]
- Karthik, K.; Rathore, R.; Thomas, P.; Arun, T.R.; Viswas, K.N.; Dhama, K.; Agarwal, R.K. New closed tube loop mediated isothermal amplification assay for prevention of product cross-contamination. MethodsX 2014, 1, 137–143. [Google Scholar] [CrossRef]
- Tao, Z.Y.; Zhou, H.Y.; Xia, H.; Xu, S.; Zhu, H.W.; Culleton, R.L.; Han, E.T.; Lu, F.; Fang, Q.; Gu, Y.P.; et al. Adaptation of a visualized loop-mediated isothermal amplification technique for field detection of Plasmodium vivax infection. Parasit Vectors 2011, 4, 115. [Google Scholar] [CrossRef]
- Hong, M.; Zha, L.; Fu, W.; Zou, M.; Li, W.; Xu, D. A modified visual loop-mediated isothermal amplification method for diagnosis and differentiation of main pathogens from Mycobacterium tuberculosis complex. World J. Microbiol. Biotechnol. 2012, 28, 523–531. [Google Scholar] [CrossRef]
- Liang, C.; Cheng, S.; Chu, Y.; Wu, H.; Zou, B.; Huang, H.; Xi, T.; Zhou, G. A closed-tube detection of loop-mediated isothermal amplification (LAMP) products using a wax-sealed fluorescent intercalator. J. NanoSci. Nanotechnol. 2013, 13, 3999–4005. [Google Scholar] [CrossRef]
- Lucchi, N.W.; Ljolje, D.; Silva-Flannery, L.; Udhayakumar, V. Use of Malachite Green-Loop Mediated Isothermal Amplification for Detection of Plasmodium spp. Parasites. PLoS ONE 2016, 11, e0151437. [Google Scholar] [CrossRef]
- Nzelu, C.O.; Gomez, E.A.; Cáceres, A.G.; Sakurai, T.; Martini-Robles, L.; Uezato, H.; Mimori, T.; Katakura, K.; Hashiguchi, Y.; Kato, H. Development of a loop-mediated isothermal amplification method for rapid mass-screening of sand flies for Leishmania infection. Acta Trop. 2014, 132, 1–6. [Google Scholar] [CrossRef]
- Stranieri, A.; Lauzi, S.; Giordano, A.; Paltrinieri, S. Reverse transcriptase loop-mediated isothermal amplification for the detection of feline coronavirus. J. Virol. Methods 2017, 243, 105–108. [Google Scholar] [CrossRef]
- Inoshima, Y.; Takasu, M.; Ishiguro, N. Establishment of an on-site diagnostic procedure for detection of orf virus from oral lesions of Japanese serows (Capricornis crispus) by loop-mediated isothermal amplification. J. Vet. Med. Sci. 2017, 78, 1841–1845. [Google Scholar] [CrossRef]
- Lee, S.H.; Baek, Y.H.; Kim, Y.H.; Choi, Y.K.; Song, M.S.; Ahn, J.Y. One-Pot Reverse Transcriptional Loop-Mediated Isothermal Amplification (RT-LAMP) for Detecting MERS-CoV. Front. Microbiol. 2016, 7, 2166. [Google Scholar] [CrossRef]
- Oscorbin, I.P.; Belousova, E.A.; Zakabunin, A.I.; Boyarskikh, U.A.; Filipenko, M.L. Comparison of fluorescent intercalating dyes for quantitative loop-mediated isothermal amplification (qLAMP). Biotechniques 2016, 61, 20–25. [Google Scholar] [CrossRef]
- Wu, R.; Liu, X.; Guo, B.; Chen, F.; Wang, X. Development of double loop-mediated isothermal amplification to detect Listeria monocytogenes in food. Curr. Microbiol. 2014, 69, 839–845. [Google Scholar] [CrossRef]
- Woźniakowski, G.; Samorek-Salamonowicz, E.; Kozdruń, W. Comparison of loop-mediated isothermal amplification and PCR for the detection and differentiation of Marek’s disease virus serotypes 1, 2, and 3. Avian Dis. 2013, 57 (Suppl. 2), 539–543. [Google Scholar]
- Wassermann, M.; Mackenstedt, U.; Romig, T. A loop-mediated isothermal amplification (LAMP) method for the identification of species within the Echinococcus granulosus complex. Vet. Parasitol. 2014, 200, 97–103. [Google Scholar] [CrossRef]
- Mwendwa, F.; Mbae, C.K.; Kinyua, J.; Mulinge, E.; Mburugu, G.N.; Njiru, Z.K. Stem loop-mediated isothermal amplification test: Comparative analysis with classical LAMP and PCR in detection of Entamoeba histolytica in Kenya. BMC Res. Notes 2017, 10, 142. [Google Scholar] [CrossRef]
- Watts, M.R.; James, G.; Sultana, Y.; Ginn, A.N.; Outhred, A.C.; Kong, F.; Verweij, J.J.; Iredell, J.R.; Chen, S.C.; Lee, R. A loop-mediated isothermal amplification (LAMP) assay for Strongyloides stercoralis in stool that uses a visual detection method with SYTO-82 fluorescent dye. Am. J. Trop. Med. Hyg. 2014, 90, 306–311. [Google Scholar] [CrossRef]
- Fischbach, J.; Xander, N.C.; Frohme, M.; Glökler, J.F. Shining a light on LAMP assays—A comparison of LAMP visualization methods including the novel use of berberine. Biotechniques 2015, 58, 189–194. [Google Scholar] [CrossRef]
- Niessen, L. Current state and future perspectives of loop-mediated isothermal amplification (LAMP)-based diagnosis of filamentous fungi and yeasts. Appl. Microbiol. Biotechnol. 2015, 99, 553–574. [Google Scholar] [CrossRef]
- Zhou, D.; Guo, J.; Xu, L.; Gao, S.; Lin, Q.; Wu, Q.; Wu, L.; Que, Y. Establishment and application of a loop-mediated isothermal amplification (LAMP) system for detection of cry1Ac transgenic sugarcane. Sci. Rep. 2014, 4, 4912. [Google Scholar] [CrossRef]
- Mori, Y.; Notomi, T. Loop-mediated isothermal amplification (LAMP): A rapid, accurate, and cost-effective diagnostic method for infectious diseases. J. Infect. Chemother. Off. J. Jpn. Soc. Chemother. 2009, 15, 62–69. [Google Scholar] [CrossRef]
- Pessôa, R.; Patriota, J.V.; Lourdes de Souza, M.; Felix, A.C.; Mamede, N.; Sanabani, S.S. Investigation Into an Outbreak of Dengue-like Illness in Pernambuco, Brazil, Revealed a Cocirculation of Zika, Chikungunya, and Dengue Virus Type 1. Medicine 2016, 95, e3201. [Google Scholar] [CrossRef]
- Magalhaes, T.; Braga, C.; Cordeiro, M.T.; Oliveira, A.L.S.; Castanha, P.M.S.; Maciel, A.P.R.; Amancio, N.M.L.; Gouveia, P.N.; Peixoto-da-Silva, V.J.; Peixoto, T.F.L.; et al. Zika virus displacement by a chikungunya outbreak in Recife, Brazil. PLoS Negl. Trop. Dis. 2017, 11, e0006055. [Google Scholar] [CrossRef]
- Cevallos, V.; Ponce, P.; Waggoner, J.J.; Pinsky, B.A.; Coloma, J.; Quiroga, C.; Morales, D.; Cárdenas, M.J. Zika and Chikungunya virus detection in naturally infected Aedes aegypti in Ecuador. Acta Trop. 2018, 177, 74–80. [Google Scholar] [CrossRef]
- Sánchez-Carbonel, J.; Tantaléan-Yépez, D.; Aguilar-Luis, M.A.; Silva-Caso, W.; Weilg, P.; Vásquez-Achaya, F.; Costa, L.; Martins-Luna, J.; Sandoval, I.; Del Valle-Mendoza, J. Identification of infection by Chikungunya, Zika, and Dengue in an area of the Peruvian coast. Molecular diagnosis and clinical characteristics. BMC Res. Notes 2018, 11, 175. [Google Scholar] [CrossRef]
- Couto-Lima, D.; Madec, Y.; Bersot, M.I.; Campos, S.S.; Motta, M.A.; Santos, F.B.D.; Vazeille, M.; Vasconcelos, P.F.D.C.; Lourenço-de-Oliveira, R.; Failloux, A.B. Potential risk of re-emergence of urban transmission of Yellow Fever virus in Brazil facilitated by competent Aedes populations. Sci. Rep. 2017, 7, 4848. [Google Scholar] [CrossRef]
- Hsieh, K.; Mage, P.L.; Csordas, A.T.; Eisenstein, M.; Soh, H.T. Simultaneous elimination of carryover contamination and detection of DNA with uracil-DNA-glycosylase-supplemented loop-mediated isothermal amplification (UDG-LAMP). Chem. Commun. 2014, 50, 3747–3749. [Google Scholar] [CrossRef]
- Ayllón, T.; Campos, R.M.; Brasil, P.; Morone, F.C.; Câmara, D.C.P.; Meira, G.L.S.; Tannich, E.; Yamamoto, K.A.; Carvalho, M.S.; Pedro, R.S.; et al. Early Evidence for Zika Virus Circulation among Aedes aegypti Mosquitoes, Rio de Janeiro, Brazil. Emerg Infect. Dis. 2017, 23, 1411–1412. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Procedure | Samples | ZIKV Strains Used | Target Region of the Primers | Analytical Sensitivity (Limit of Detection) | Pretreatment or Need for RNA Extraction from the Sample | Validation with Clinical Samples | Reference |
|---|---|---|---|---|---|---|---|
| One-step RT-LAMP Detection: development of a tape based on lateral flow assay (LFA) | Human blood spiked with RNA ZIKV | MR 766 Uganda (AY632535) | Envelope Protein | 10° ZIKV RNA Copy | No | No | [40] |
| Development of a microfluidic cassette based on RT-LAMP Detection: through the naked eye with the aid of leuco crystal violet (LCV) | Saliva spiked with ZIKV | MEX 2-81 (Mosquito/2016/México) | Envelope Protein | 5 PFU | Yes | No | [37] |
| One-step LAMP Detection: development of LAMP protocol coupled with AC susceptometry | Human serum spiked with synthetic oligonucleotides of ZIKV | Synthetic oligonucleotides of ZIKV | NS5 | 1 aM | No | No | [38] |
| One-step RT-LAMP Detection: turbidity monitoring coupled to RT-qPCR. Through the naked eye with calcein and UV light | Saliva, urine, and serum experimentally infected with ZIKV | ZIKV_SMGC-1 China (KX266255) | NS1 | 0.02 PFU/test | Yes | No | [39] |
| One-step RT-LAMP Detection: through the colorimetric technology | Clinical samples including urine and human serum | Puerto Rico (PRVABC 59) | Envelope Protein | 1.2 RNA copies/μL | Yes | Yes | [59] |
| One-step RT-LAMP Detection: monitoring of fluorescence coupled to RT-qPCR platform and through the naked eye | Clinical samples including serum, plasma, and human semen. In addition to blood, plasma, saliva, urine, semen and Aedes mosquito spiked with ZIKV | Puerto Rico (PRVABC59), P6-740 (HQ234449), 41525 (KU955591) and MR 766 Uganda (AY632535) | NS2A | 0.43 PFU | No | Yes | [41] |
| Development of a Trioplex assay to detect ZIKV, DENV, and CHIKV Detection: development of RT-LAMP protocol coupled with a smartphone for detecting fluorescence | Human blood spiked with ZIKV | Puerto Rico (PRVABC59), | NS1 | 1.56 PFU/mL | Yes | No | [60] |
| Development of a platform based on RT-LAMP assay Detection: fluorescence monitoring coupled to RT-qPCR platform | Clinical samples, including serum and human urine. In addition to human serum and urine spiked with ZIKV | PRABC59, 976 Uganda, ArD157995, P6-740, CPC-0740*, 41525-DAK* | Envelope Protein | 14.5 TCID50/mL | Yes | Yes | [28] |
| Development of a Trioplex assay to detect ZIKV, DENV, and CHIKV Detection: monitoring of fluorescence coupled to quenching of unincorporated amplification signal reporters (QUASR) and smartphone | Blood, urine, and saliva spiked with ZIKV | Puerto Rico (PRVABC59), Honduras (R103451), and Brazilian strain; | NS5 | 2 PFU/mL | Yes | No | [58] |
| Development of a Trioplex kit trial to detect ZIKV, DENV, and CHIKV Detection: monitoring of fluorescence coupled to RT-qPCR platform and through the naked eye | Urine, saliva, plasma and Aedes mosquito spiked with ZIKV | Puerto Rico, (PRVABC59, KU501215.1) | NS5 | ~ 0.71 PFU equivalent viral RNAs | Yes | No | [27] |
| One-step RT-LAMP Detection: through the naked eye with SYBR Green I | Urine, serum, and infected mosquito samples | MR 766 Uganda (AY632535); MEX20 (ZK-HU 0165 P), Puerto Rico (PRVABC 59) and PB81 (H815744) | NS5 | 1 copy of the genome/rxn | No | No | [62] |
| Development of a platform based on RT-LAMP Detection: monitoring of fluorescence coupled to RT-qPCR platform | Clinical samples, including saliva and urine. In addition to saliva and urine spiked with ZIKV | (PRVABC59 – NR-50240) | Capsid | 2.2 × 103 RNA copies/mL | Yes | No | [61] |
| Development of a platform based on RT-LAMP Detection: monitoring of fluorescence coupled to RT-qPCR | Viral RNA | MR 766 Uganda | NS5 | 3.3 ng/μL | Yes | No | [57] |
| Development of RT-LAMP platform coupled with on paper microfluidic chips Detection: the color change is monitored in situ with a smartphone | Plasma and urine spiked with ZIKV | Purified ZIKV particles | NS5 | 1 copy/μL | Yes | No | [44] |
| One-step RT-LAMP Detection: through the naked eye with SYBR Green I | Clinical samples, including human urine. In addition to urine and Aedes mosquito spiked with ZIKV | MR 766 Uganda (AY632535), MEX20 (ZK-HU 0165 P), Puerto Rico (PRVABC 59), and PB81 (H815744) | NS5 | 1 copy of the genome/rxn | No | No | [42] |
| Development of an RT-LAMP platform coupled with reverse dot blot analysis (RDB) Detection: monitoring of fluorescence and reverse dot-blot for detection | Saliva spiked with ZIKV | Thailand (PLCal_ZV), Puerto Rican (NR-50244), Thailand (NR-50242), Florida (NR5024), and Honduras (NR-50358) | Capsid | 2.103 RNA copies /mL | Yes | No | [63] |
| Development of a LAMP Detection: a calcein/Mn2+ complex chromogenic method and real-time turbidity monitoring | Viral RNA | Recombinant plasmids containing NS5 gene or E (ZIKV strain Natal RGN) | NS5 and envelope protein | 0.5 × 10−9 pmol/µL DNA for NS5 and 1.12 × 10−11 pmol/µL DNA for envelope protein | Yes | No | [43] |
| One-step RT-LAMP Detection: through the naked eye with SYBR Green I | Mosquito samples including experimentally and naturally infected mosquitoes (Aedes aegypti and Culex quinquefasciatus) | PE-243 (KX197192) | Envelope protein | 10−5 PFU | No | Yes | [53] |
| One-step RT-LAMP Detection: through the naked eye with SYBR Green I | Serum spiked with ZIKV and patient samples including human serum | - | 10−3 copies of RNA (20 zepto-molar) | No | Yes | [64] | |
| One-step RT-LAMP Detection: turbidity monitoring | Serum and urine spiked with ZIKV | PRVABC59 (KX601168), MRS_OPY_Martinique_Pari_2015 (KU647676), H/PF/2013 (KJ776791), and MR766 (LC002520) | Envelope protein/NS4A | 0.17 FFU/mL–2.3 × 102 FFU/mL | Yes | No | [45] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, S.J.R.d.; Pardee, K.; Pena, L. Loop-Mediated Isothermal Amplification (LAMP) for the Diagnosis of Zika Virus: A Review. Viruses 2020, 12, 19. https://doi.org/10.3390/v12010019
Silva SJRd, Pardee K, Pena L. Loop-Mediated Isothermal Amplification (LAMP) for the Diagnosis of Zika Virus: A Review. Viruses. 2020; 12(1):19. https://doi.org/10.3390/v12010019
Chicago/Turabian StyleSilva, Severino Jefferson Ribeiro da, Keith Pardee, and Lindomar Pena. 2020. "Loop-Mediated Isothermal Amplification (LAMP) for the Diagnosis of Zika Virus: A Review" Viruses 12, no. 1: 19. https://doi.org/10.3390/v12010019
APA StyleSilva, S. J. R. d., Pardee, K., & Pena, L. (2020). Loop-Mediated Isothermal Amplification (LAMP) for the Diagnosis of Zika Virus: A Review. Viruses, 12(1), 19. https://doi.org/10.3390/v12010019