Prophylactic Hepatitis E Vaccines: Antigenic Analysis and Serological Evaluation

 , and
, and
Abstract
1. Introduction
2. The Design of Hepatitis E Vaccines
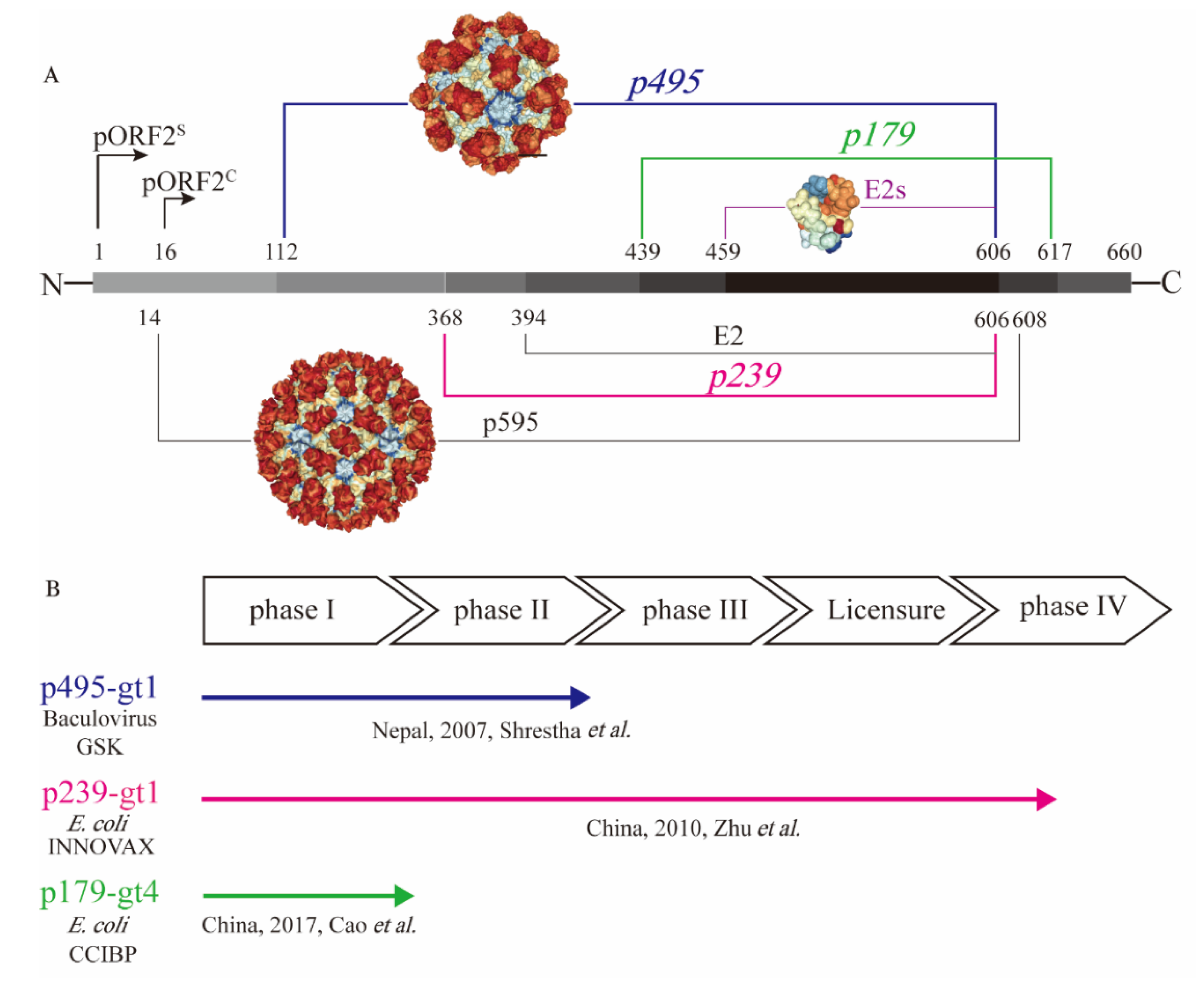
2.1. Molecular Structure of Different Truncated Versions of pORF2
2.2. Hepatitis E Vaccines
3. Analysis of the Native-Like Epitopes on Recombinant Antigens
3.1. Epitope-Specific Analysis of the HEV Vaccine Antigen by mAbs
3.2. The Application of an In Vitro Relative Potency Assay (IVRP)
4. The Serological Evaluation
4.1. The Binding Antibody Analysis
4.2. The Neutralizing Antibody Analysis
5. Can Current Vaccines Protect against Hetero-Genotypes?
6. Conclusions and Prospects
Funding
Conflicts of Interest
References
- Khuroo, M.S. Study of an epidemic of non-A, non-B hepatitis. Possibility of another human hepatitis virus distinct from post-transfusion non-A, non-B type. Am. J. Med. 1980, 68, 818–824. [Google Scholar] [CrossRef]
- Webb, G.W.; Dalton, H.R. Hepatitis E: An underestimated emerging threat. Adv. Infect. Dis. 2019, 6, 2049936119837162. [Google Scholar] [CrossRef]
- World Health Organization. Recommendations to Assure the Quality, Safety and Efficacy of Recombinant Hepatitis E Vaccines. Available online: https://www.who.int/biologicals/BS.2018.2348.Recommendations_HEP_E_vaccines.pdf (accessed on 10 December 2019).
- World Health Organization. Global Hepatitis Report. 2017. Available online: https://www.who.int/hepatitis/publications/global-hepatitis-report2017/en/ (accessed on 8 January 2020).
- Kamar, N.; Bendall, R.; Legrand-Abravanel, F.; Xia, N.S.; Ijaz, S.; Izopet, J.; Dalton, H.R. Hepatitis E. Lancet 2012, 379, 2477–2488. [Google Scholar] [CrossRef]
- Gallian, P.; Pouchol, E.; Djoudi, R.; Lhomme, S.; Mouna, L.; Gross, S.; Bierling, P.; Assal, A.; Kamar, N.; Mallet, V.; et al. Transfusion-Transmitted Hepatitis E Virus Infection in France. Transfus. Med. Rev. 2019, 33, 146–153. [Google Scholar] [CrossRef]
- Vollmer, T.; Diekmann, J.; Eberhardt, M.; Knabbe, C.; Dreier, J. Hepatitis E in blood donors: Investigation of the natural course of asymptomatic infection, Germany, 2011. Euro. Surveill. 2016, 21. [Google Scholar] [CrossRef]
- Sahli, R.; Fraga, M.; Semela, D.; Moradpour, D.; Gouttenoire, J. Rabbit HEV in immunosuppressed patients with hepatitis E acquired in Switzerland. J. Hepatol. 2019, 70, 1023–1025. [Google Scholar] [CrossRef]
- Yapa, C.M.; Furlong, C.; Rosewell, A.; Ward, K.A.; Adamson, S.; Shadbolt, C.; Kok, J.; Tracy, S.L.; Bowden, S.; Smedley, E.J.; et al. First reported outbreak of locally acquired hepatitis E virus infection in Australia. Med. J. Aust. 2016, 204, 274. [Google Scholar] [CrossRef]
- Nanmoku, K.; Owada, Y.; Oshiro, Y.; Kurosawa, A.; Kubo, T.; Shinzato, T.; Shimizu, T.; Kimura, T.; Sakuma, Y.; Ishikawa, N.; et al. Prevalence and characteristics of hepatitis E virus infection in kidney transplant recipients: A single-center experience in Japan. Transpl. Infect. Dis. 2019, 21, e13033. [Google Scholar] [CrossRef]
- Donnelly, M.C.; Scobie, L.; Crossan, C.L.; Dalton, H.; Hayes, P.C.; Simpson, K.J. Review article: Hepatitis E-a concise review of virology, epidemiology, clinical presentation and therapy. Aliment. Pharm. 2017, 46, 126–141. [Google Scholar] [CrossRef]
- Lee, G.H.; Tan, B.H.; Teo, E.C.; Lim, S.G.; Dan, Y.Y.; Wee, A.; Aw, P.P.; Zhu, Y.; Hibberd, M.L.; Tan, C.K.; et al. Chronic Infection With Camelid Hepatitis E Virus in a Liver Transplant Recipient Who Regularly Consumes Camel Meat and Milk. Gastroenterology 2016, 150, 355–357. [Google Scholar] [CrossRef]
- Woo, P.C.; Lau, S.K.; Teng, J.L.; Cao, K.Y.; Wernery, U.; Schountz, T.; Chiu, T.H.; Tsang, A.K.; Wong, P.C.; Wong, E.Y.; et al. New Hepatitis E Virus Genotype in Bactrian Camels, Xinjiang, China, 2013. Emerg. Infect. Dis. 2016, 22, 2219–2221. [Google Scholar] [CrossRef] [PubMed]
- Debing, Y.; Moradpour, D.; Neyts, J.; Gouttenoire, J. Update on hepatitis E virology: Implications for clinical practice. J. Hepatol. 2016, 65, 200–212. [Google Scholar] [CrossRef] [PubMed]
- Nan, Y.; Yu, Y.; Ma, Z.; Khattar, S.K.; Fredericksen, B.; Zhang, Y.J. Hepatitis E virus inhibits type I interferon induction by ORF1 products. J. Virol. 2014, 88, 11924–11932. [Google Scholar] [CrossRef]
- Nair, V.P.; Anang, S.; Subramani, C.; Madhvi, A.; Bakshi, K.; Srivastava, A.; Nayak, B.; Surjit, M. Endoplasmic Reticulum Stress Induced Synthesis of a Novel Viral Factor Mediates Efficient Replication of Genotype-1 Hepatitis E Virus. PLOS Pathog. 2016, 12, e1005521. [Google Scholar] [CrossRef]
- Kamar, N.; Izopet, J.; Pavio, N.; Aggarwal, R.; Labrique, A.; Wedemeyer, H.; Dalton, H.R. Hepatitis E virus infection. Nat. Rev. Dis. Primers 2017, 3, 17086. [Google Scholar] [CrossRef]
- Feng, Z.; Hirai-Yuki, A.; McKnight, K.L.; Lemon, S.M. Naked Viruses That Aren’t Always Naked: Quasi-Enveloped Agents of Acute Hepatitis. Annu. Rev. Virol. 2014, 1, 539–560. [Google Scholar] [CrossRef]
- Marion, O.; Lhomme, S.; Nayrac, M.; Dubois, M.; Pucelle, M.; Requena, M.; Migueres, M.; Abravanel, F.; Peron, J.M.; Carrere, N.; et al. Hepatitis E virus replication in human intestinal cells. Gut 2019. [Google Scholar] [CrossRef]
- Takahashi, M.; Tanaka, T.; Takahashi, H.; Hoshino, Y.; Nagashima, S.; Mizuo, H.; Yazaki, Y.; Takagi, T.; Azuma, M.; Kusano, E.; et al. Hepatitis E Virus (HEV) strains in serum samples can replicate efficiently in cultured cells despite the coexistence of HEV antibodies: Characterization of HEV virions in blood circulation. J. Clin. Microbiol. 2010, 48, 1112–1125. [Google Scholar] [CrossRef]
- Mori, Y.; Matsuura, Y. Structure of hepatitis E viral particle. Virus Res. 2011, 161, 59–64. [Google Scholar] [CrossRef]
- Li, S.W.; Zhao, Q.; Wu, T.; Chen, S.; Zhang, J.; Xia, N.S. The development of a recombinant hepatitis E vaccine HEV 239. Hum. Vaccin. Immunother. 2015, 11, 908–914. [Google Scholar] [CrossRef]
- Zhang, X.; Xin, L.; Li, S.; Fang, M.; Zhang, J.; Xia, N.; Zhao, Q. Lessons learned from successful human vaccines: Delineating key epitopes by dissecting the capsid proteins. Hum. Vaccin. Immunother. 2015, 11, 1277–1292. [Google Scholar] [CrossRef]
- Innis, B.L.; Lynch, J.A. Immunization against Hepatitis E. Cold Spring Harb. Perspect. Med. 2018, 8, a032573. [Google Scholar] [CrossRef]
- Wang, X.; Li, M.; Lin, Z.; Pan, H.; Tang, Z.; Zheng, Z.; Li, S.; Zhang, J.; Xia, N.; Zhao, Q. Multifaceted characterization of recombinant protein-based vaccines: An immunochemical toolbox for epitope-specific analyses of the hepatitis E vaccine. Vaccine 2018, 36, 7650–7658. [Google Scholar] [CrossRef]
- Group, C.-V.W. A-VAX: Applying Quality by Design to Vaccines. 2012. Available online: https://www.dcvmn.org/IMG/pdf/a-vax-applying-qbd-to-vaccines_2012.pdf. (accessed on 10 December 2019).
- Huang, X.; Wang, X.; Zhang, J.; Xia, N.; Zhao, Q. Escherichia coli-derived virus-like particles in vaccine development. Npj. Vaccines 2017, 2, 3. [Google Scholar] [CrossRef]
- Yin, X.; Ying, D.; Lhomme, S.; Tang, Z.; Walker, C.M.; Xia, N.; Zheng, Z.; Feng, Z. Origin, antigenicity, and function of a secreted form of ORF2 in hepatitis E virus infection. Proc. Natl. Acad. Sci. USA 2018, 115, 4773–4778. [Google Scholar] [CrossRef]
- Yamashita, T.; Mori, Y.; Miyazaki, N.; Cheng, R.H.; Yoshimura, M.; Unno, H.; Shima, R.; Moriishi, K.; Tsukihara, T.; Li, T.C.; et al. Biological and immunological characteristics of hepatitis E virus-like particles based on the crystal structure. Proc. Natl. Acad. Sci. USA 2009, 106, 12986–12991. [Google Scholar] [CrossRef]
- Huang, C.C.; Nguyen, D.; Fernandez, J.; Yun, K.Y.; Fry, K.E.; Bradley, D.W.; Tam, A.W.; Reyes, G.R. Molecular cloning and sequencing of the Mexico isolate of hepatitis E virus (HEV). Virology 1992, 191, 550–558. [Google Scholar] [CrossRef]
- Li, S.; Tang, X.; Seetharaman, J.; Yang, C.; Gu, Y.; Zhang, J.; Du, H.; Shih, J.W.; Hew, C.L.; Sivaraman, J.; et al. Dimerization of hepatitis E virus capsid protein E2s domain is essential for virus-host interaction. PLOS Pathog. 2009, 5, e1000537. [Google Scholar] [CrossRef]
- Tang, X.; Yang, C.; Gu, Y.; Song, C.; Zhang, X.; Wang, Y.; Zhang, J.; Hew, C.L.; Li, S.; Xia, N.; et al. Structural basis for the neutralization and genotype specificity of hepatitis E virus. Proc. Natl. Acad. Sci. USA 2011, 108, 10266–10271. [Google Scholar] [CrossRef]
- Gu, Y.; Tang, X.; Zhang, X.; Song, C.; Zheng, M.; Wang, K.; Zhang, J.; Ng, M.H.; Hew, C.L.; Li, S.; et al. Structural basis for the neutralization of hepatitis E virus by a cross-genotype antibody. Cell Res. 2015, 25, 604–620. [Google Scholar] [CrossRef]
- Zhang, J.Z.; Ng, M.H.; Xia, N.S.; Lau, S.H.; Che, X.Y.; Chau, T.N.; Lai, S.T.; Im, S.W. Conformational antigenic determinants generated by interactions between a bacterially expressed recombinant peptide of the hepatitis E virus structural protein. J. Med. Virol. 2001, 64, 125–132. [Google Scholar] [CrossRef]
- Li, S.W.; Zhang, J.; Li, Y.M.; Ou, S.H.; Huang, G.Y.; He, Z.Q.; Ge, S.X.; Xian, Y.L.; Pang, S.Q.; Ng, M.H.; et al. A bacterially expressed particulate hepatitis E vaccine: Antigenicity, immunogenicity and protectivity on primates. Vaccine 2005, 23, 2893–2901. [Google Scholar] [CrossRef]
- Cao, Y.F.; Tao, H.; Hu, Y.M.; Shi, C.B.; Wu, X.; Liang, Q.; Chi, C.P.; Li, L.; Liang, Z.L.; Meng, J.H.; et al. A phase 1 randomized open-label clinical study to evaluate the safety and tolerability of a novel recombinant hepatitis E vaccine. Vaccine 2017, 35, 5073–5080. [Google Scholar] [CrossRef]
- Cao, Y.; Bing, Z.; Guan, S.; Zhang, Z.; Wang, X. Development of new hepatitis E vaccines. Hum. Vaccin. Immunother. 2018, 14, 2254–2262. [Google Scholar] [CrossRef]
- Shrestha, M.P.; Scott, R.M.; Joshi, D.M.; Mammen, M.P., Jr.; Thapa, G.B.; Thapa, N.; Myint, K.S.; Fourneau, M.; Kuschner, R.A.; Shrestha, S.K.; et al. Safety and efficacy of a recombinant hepatitis E vaccine. N. Engl. J. Med. 2007, 356, 895–903. [Google Scholar] [CrossRef]
- Zhu, F.C.; Zhang, J.; Zhang, X.F.; Zhou, C.; Wang, Z.Z.; Huang, S.J.; Wang, H.; Yang, C.L.; Jiang, H.M.; Cai, J.P.; et al. Efficacy and safety of a recombinant hepatitis E vaccine in healthy adults: A large-scale, randomised, double-blind placebo-controlled, phase 3 trial. Lancet 2010, 376, 895–902. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, X.F.; Huang, S.J.; Wu, T.; Hu, Y.M.; Wang, Z.Z.; Wang, H.; Jiang, H.M.; Wang, Y.J.; Yan, Q.; et al. Long-term efficacy of a hepatitis E vaccine. N. Engl. J. Med. 2015, 372, 914–922. [Google Scholar] [CrossRef]
- Yu, X.Y.; Chen, Z.P.; Wang, S.Y.; Pan, H.R.; Wang, Z.F.; Zhang, Q.F.; Shen, L.Z.; Zheng, X.P.; Yan, C.F.; Lu, M.; et al. Safety and immunogenicity of hepatitis E vaccine in elderly people older than 65years. Vaccine 2019, 37, 4581–4586. [Google Scholar] [CrossRef]
- Zhang, X.; Wei, M.; Pan, H.; Lin, Z.; Wang, K.; Weng, Z.; Zhu, Y.; Xin, L.; Zhang, J.; Li, S.; et al. Robust manufacturing and comprehensive characterization of recombinant hepatitis E virus-like particles in Hecolin((R)). Vaccine 2014, 32, 4039–4050. [Google Scholar] [CrossRef]
- Zhao, M.; Li, X.J.; Tang, Z.M.; Yang, F.; Wang, S.L.; Cai, W.; Zhang, K.; Xia, N.S.; Zheng, Z.Z. A Comprehensive Study of Neutralizing Antigenic Sites on the Hepatitis E Virus (HEV) Capsid by Constructing, Clustering, and Characterizing a Tool Box. J. Biol. Chem. 2015, 290, 19910–19922. [Google Scholar] [CrossRef]
- Weng, Z.; Zhao, Q. Utilizing ELISA to monitor protein-protein interaction. Methods Mol. Biol. 2015, 1278, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Wang, X.; Fang, M.; Xia, N.; Zhao, Q. Detection of subtle differences in analogous viral capsid proteins by allowing unrestricted specific interaction in solution competition ELISA. J. Virol. Methods 2016, 236, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Wang, X.; Zhang, Z.; Li, Y.; Lin, Z.; Pan, H.; Gu, Y.; Li, S.; Zhang, J.; Xia, N.; et al. Demonstration of real-time and accelerated stability of hepatitis E vaccine with a combination of different physicochemical and immunochemical methods. J. Pharm. Biomed. 2020, 177. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, T.; Cao, L.; Wang, X.; Cao, J.; Huang, X.; Cai, Y.; Lin, Z.; Pan, H.; Yuan, Q.; et al. Simultaneous in situ visualization and quantitation of dual antigens adsorbed on adjuvants using high content analysis. Nanomed. (Lond) 2019, 14, 2535–2548. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, M.; Yang, F.; Li, Y.; Zheng, Z.; Zhang, X.; Lin, Q.; Wang, Y.; Li, S.; Xia, N.; et al. Comparable quality attributes of hepatitis E vaccine antigen with and without adjuvant adsorption-dissolution treatment. Hum. Vaccin. Immunother. 2015, 11, 1129–1139. [Google Scholar] [CrossRef][Green Version]
- Zhao, Q.; Li, S.; Yu, H.; Xia, N.; Modis, Y. Virus-like particle-based human vaccines: Quality assessment based on structural and functional properties. Trends Biotechnol. 2013, 31, 654–663. [Google Scholar] [CrossRef]
- Sitrin, R.D.; Zhao, Q.; Potter, C.S.; Carragher, B.; Washabaugh, M.W. Recombinant Virus-like Particle Protein Vaccines. In Vaccine Analysis: Strategies; Nunnally, B., Turula, V., Sitrin, R., Eds.; Springer: Berlin, Germany, 2015. [Google Scholar]
- Wang, X.; An, Z.; Luo, W.; Xia, N.; Zhao, Q. Molecular and functional analysis of monoclonal antibodies in support of biologics development. Protein Cell 2018, 9, 74–85. [Google Scholar] [CrossRef]
- Morgeaux, S.; Poirier, B.; Ragan, C.I.; Wilkinson, D.; Arabin, U.; Guinet-Morlot, F.; Levis, R.; Meyer, H.; Riou, P.; Shaid, S.; et al. Replacement of in vivo human rabies vaccine potency testing by in vitro glycoprotein quantification using ELISA - Results of an international collaborative study. Vaccine 2017, 35, 966–971. [Google Scholar] [CrossRef]
- Zhao, Q.; Towne, V.; Brown, M.; Wang, Y.; Abraham, D.; Oswald, C.B.; Gimenez, J.A.; Washabaugh, M.W.; Kennedy, R.; Sitrin, R.D. In-depth process understanding of RECOMBIVAX HB(R) maturation and potential epitope improvements with redox treatment: Multifaceted biochemical and immunochemical characterization. Vaccine 2011, 29, 7936–7941. [Google Scholar] [CrossRef]
- Mostafa, M.M.; Al-Ghobashy, M.A.; Fathalla, F.A.; Salem, M.Y. Optimization and validation of ELISA immunoassay for the evaluation of in-vitro relative potency of a four-component human papillomavirus vaccine products. Biologicals 2016, 44, 596–599. [Google Scholar] [CrossRef]
- Khateri, M.; Abdoli, A.; Motevalli, F.; Fotouhi, F.; Bolhassani, A.; Arashkia, A.; Jazaeri, E.O.; Shahbazi, S.; Mehrbod, P.; Naziri, H.; et al. Evaluation of autophagy induction on HEV 239 vaccine immune response in a mouse model. Iubmb. Life 2018, 70, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Pinto, L.A.; Dillner, J.; Beddows, S.; Unger, E.R. Immunogenicity of HPV prophylactic vaccines: Serology assays and their use in HPV vaccine evaluation and development. Vaccine 2018, 36, 4792–4799. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Ge, S.X.; Huang, G.Y.; Li, S.W.; He, Z.Q.; Wang, Y.B.; Zheng, Y.J.; Gu, Y.; Ng, M.H.; Xia, N.S. Evaluation of antibody-based and nucleic acid-based assays for diagnosis of hepatitis E virus infection in a rhesus monkey model. J. Med. Virol. 2003, 71, 518–526. [Google Scholar] [CrossRef] [PubMed]
- Tsarev, S.A.; Tsareva, T.S.; Emerson, S.U.; Govindarajan, S.; Shapiro, M.; Gerin, J.L.; Purcell, R.H. Recombinant vaccine against hepatitis E: Dose response and protection against heterologous challenge. Vaccine 1997, 15, 1834–1838. [Google Scholar] [CrossRef]
- Wen, J.; Behloul, N.; Dai, X.; Dong, C.; Liang, J.; Zhang, M.; Shi, C.; Meng, J. Immunogenicity difference between two hepatitis E vaccines derived from genotype 1 and 4. Antivir. Res. 2016, 128, 36–42. [Google Scholar] [CrossRef]
- Tam, A.W.; White, R.; Reed, E.; Short, M.; Zhang, Y.; Fuerst, T.R.; Lanford, R.E. In vitro propagation and production of hepatitis E virus from in vivo-infected primary macaque hepatocytes. Virology 1996, 215, 1–9. [Google Scholar] [CrossRef]
- Tanaka, T.; Takahashi, M.; Kusano, E.; Okamoto, H. Development and evaluation of an efficient cell-culture system for Hepatitis E virus. J. Gen. Virol. 2007, 88, 903–911. [Google Scholar] [CrossRef]
- Shukla, P.; Nguyen, H.T.; Torian, U.; Engle, R.E.; Faulk, K.; Dalton, H.R.; Bendall, R.P.; Keane, F.E.; Purcell, R.H.; Emerson, S.U. Cross-species infections of cultured cells by hepatitis E virus and discovery of an infectious virus-host recombinant. Proc. Natl. Acad. Sci. USA 2011, 108, 2438–2443. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, C.; Qi, Y.; Geng, Y. Hepatitis E Virus. Adv. Exp. Med. Biol. 2016, 948, 1–16. [Google Scholar] [CrossRef]
- Liu, C.; Cai, W.; Yin, X.; Tang, Z.; Wen, G.; Ambardekar, C.; Li, X.; Ying, D.; Feng, Z.; Zheng, Z.; et al. An Optimized High-Throughput Neutralization Assay for Hepatitis E Virus (HEV) Involving Detection of Secreted Porf2. Viruses 2019, 11. [Google Scholar] [CrossRef]
- Takahashi, M.; Hoshino, Y.; Tanaka, T.; Takahashi, H.; Nishizawa, T.; Okamoto, H. Production of monoclonal antibodies against hepatitis E virus capsid protein and evaluation of their neutralizing activity in a cell culture system. Arch. Virol. 2008, 153, 657–666. [Google Scholar] [CrossRef] [PubMed]
- Tam, A.W.; White, R.; Yarbough, P.O.; Murphy, B.J.; McAtee, C.P.; Lanford, R.E.; Fuerst, T.R. In vitro infection and replication of hepatitis E virus in primary cynomolgus macaque hepatocytes. Virology 1997, 238, 94–102. [Google Scholar] [CrossRef]
- Meng, J.; Dubreuil, P.; Pillot, J. A new PCR-based seroneutralization assay in cell culture for diagnosis of hepatitis E. J. Clin. Microbiol. 1997, 35, 1373–1377. [Google Scholar] [CrossRef]
- Emerson, S.U.; Clemente-Casares, P.; Moiduddin, N.; Arankalle, V.A.; Torian, U.; Purcell, R.H. Putative neutralization epitopes and broad cross-genotype neutralization of Hepatitis E virus confirmed by a quantitative cell-culture assay. J. Gen. Virol. 2006, 87, 697–704. [Google Scholar] [CrossRef]
- Cai, W.; Tang, Z.M.; Wen, G.P.; Wang, S.L.; Ji, W.F.; Yang, M.; Ying, D.; Zheng, Z.Z.; Xia, N.S. A high-throughput neutralizing assay for antibodies and sera against hepatitis E virus. Sci. Rep. 2016, 6, 25141. [Google Scholar] [CrossRef]
- He, S.; Miao, J.; Zheng, Z.; Wu, T.; Xie, M.; Tang, M.; Zhang, J.; Ng, M.H.; Xia, N. Putative receptor-binding sites of hepatitis E virus. J. Gen. Virol. 2008, 89, 245–249. [Google Scholar] [CrossRef]
- Zheng, Z.Z.; Miao, J.; Zhao, M.; Tang, M.; Yeo, A.E.; Yu, H.; Zhang, J.; Xia, N.S. Role of heat-shock protein 90 in hepatitis E virus capsid trafficking. J. Gen. Virol. 2010, 91, 1728–1736. [Google Scholar] [CrossRef]
- Wu, X.; Chen, P.; Lin, H.; Su, Y.; Hao, X.; Cao, Y.; Li, L.; Zhu, F.; Liang, Z. Dynamics of 8G12 competitive antibody in “prime-boost” vaccination of Hepatitis E vaccine. Hum. Vaccin. Immunother. 2017, 13, 1–6. [Google Scholar] [CrossRef][Green Version]
- Chen, S.; Huang, X.; Li, Y.; Wang, X.; Pan, H.; Lin, Z.; Zheng, Q.; Li, S.; Zhang, J.; Xia, N.; et al. Altered antigenicity and immunogenicity of human papillomavirus virus-like particles in the presence of thimerosal. Eur. J. Pharm. Biopharm. 2019, 141, 221–231. [Google Scholar] [CrossRef]
- Smith, G.; Raghunandan, R.; Wu, Y.; Liu, Y.; Massare, M.; Nathan, M.; Zhou, B.; Lu, H.; Boddapati, S.; Li, J.; et al. Respiratory syncytial virus fusion glycoprotein expressed in insect cells form protein nanoparticles that induce protective immunity in cotton rats. PLoS ONE 2012, 7, e50852. [Google Scholar] [CrossRef]
- Sudenga, S.L.; Torres, B.N.; Botha, M.H.; Zeier, M.; Abrahamsen, M.E.; Glashoff, R.H.; Engelbrecht, S.; Schim Van der Loeff, M.F.; Van der Laan, L.E.; Kipping, S.; et al. HPV serostatus pre- and post-vaccination in a randomized phase II preparedness trial among young Western Cape, South African women: The evri trial. Papillomavirus Res. 2017, 3, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Szkolnicka, D.; Pollan, A.; Da Silva, N.; Oechslin, N.; Gouttenoire, J.; Moradpour, D. Recombinant Hepatitis E Viruses Harboring Tags in the ORF1 Protein. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [PubMed]
- Purcell, R.H.; Nguyen, H.; Shapiro, M.; Engle, R.E.; Govindarajan, S.; Blackwelder, W.C.; Wong, D.C.; Prieels, J.P.; Emerson, S.U. Pre-clinical immunogenicity and efficacy trial of a recombinant hepatitis E vaccine. Vaccine 2003, 21, 2607–2615. [Google Scholar] [CrossRef]
- World Health Organization; Regional Office for Africa. Weekly Bulletin on Outbreak and other Emergencies: Week 14: 01–07 April 2019. Available online: https://apps.who.int/iris/handle/10665/1642?locale=en (accessed on 10 December 2019).
- Holm, D.K.; Moessner, B.K.; Engle, R.E.; Zaaijer, H.L.; Georgsen, J.; Purcell, R.H.; Christensen, P.B. Declining prevalence of hepatitis E antibodies among Danish blood donors. Transfusion 2015, 55, 1662–1667. [Google Scholar] [CrossRef]
- Effectiveness Trial to Evaluate Protection of Pregnant Women by Hepatitis E Vaccine in Bangladesh. Available online: https://clinicaltrials.gov/ct2/show/NCT02759991?term=NCT02759991&draw=2&rank=1 (accessed on 9 January 2020).
- Safety Study of Hepatitis E Vaccine (HEV239). Available online: https://clinicaltrials.gov/ct2/show/NCT03827395?term=NCT03827395&draw=2&rank=1 (accessed on 9 January 2020).


{kind=link}
| Vaccine Antigen. | Antibody Type | Species | Coating Antigen | Reference |
|---|---|---|---|---|
| p495 | total immunoglobulin | rhesus monkey | p495 (112−606) | Tsarev et al., 1997 [58] |
| human | Shrestha et al., 2007 [38] | |||
| p239 | IgG | rhesus monkey | E2 (394−606) * | Li et al., 2005 [35] |
| human | Zhu et al., 2010 [39] |
| Host Cell | Virus/Genotype | Type of Sample | Reference |
|---|---|---|---|
| Native virus | |||
| HepG2/C3A cells | Kernow (gt3) | serum (rhesus macaque & human) | Liu et al., 2019 [64] * |
| Huh7 cells | stool and bile-derived HEV (gt1/4) | mAb 8G12 | Gu et al., 2015 [33] |
| PLC/PRF/5 cells | JE03-1760F (gt3) | mAb H6225 | Takahashi et al., 2008 [65] |
| Primary hepatocytes | Burma (gt1) | purified IgGs (cynomolgus monkey) | Tam et al., 1997 [66] |
| PLC/PRF/5 cells | F23, SAR-55 (gt1) | serum (cynomolgus monkey & human) | Meng et al., 1997 [67] |
| HepG2/C3A cells | Sar55, Mex14, Meng (gt1) | serum (rhesus macaque) | Emerson et al., 2006 [68] |
| PLC/PRF/5 and A549 cells | JE03-1760F (gt3) | serum (human) | Tanaka et al., 2007 [61] |
| PLC/PRF/5 and A549 cells | serum-derived HEV (gt3) | serum (human) | Takahashi et al., 2010 [20] |
| Virus surrogate | |||
| HepG2 cells | recombinant protein 239 | serum (rhesus macaque) | Cai et al., 2016 [69] |
| Vaccination | Subject | Genotype 1 | Genotype 2 | Genotype 3 | Genotype 4 | Reference |
|---|---|---|---|---|---|---|
| Animal- or human-based analysis | ||||||
| p495-gt1 | human | √ | - | - | - | Shrestha et al., 2007 [38] |
| p239-gt1 | human | - | - | - | √ | Zhu et al., 2010 [39] |
| p495-gt1 | rhesus monkey | √ a | √ b | √ c | - | Purcell et al., 2003 [77] |
| p495-gt1 | rhesus monkey | √ a | √ b | - | - | Tsarev et al., 1997 [58] |
| p239-gt1 | rhesus monkey | √ | - | - | √ | Li et al., 2005 [35] |
| p179-gt4 | rhesus monkey | √ | - | - | √ | Cao et al., 2017 [36] |
| Cell-based analysis | ||||||
| p239 | HepG2/C3A | - | - | √ d | - | Liu et al., 2019 [64] |
| 8G12 | Huh7 cells | √ | - | - | √ | Gu et al., 2015 [33] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Huang, X.; Zhang, Z.; Li, S.; Zhang, J.; Xia, N.; Zhao, Q. Prophylactic Hepatitis E Vaccines: Antigenic Analysis and Serological Evaluation. Viruses 2020, 12, 109. https://doi.org/10.3390/v12010109
Li Y, Huang X, Zhang Z, Li S, Zhang J, Xia N, Zhao Q. Prophylactic Hepatitis E Vaccines: Antigenic Analysis and Serological Evaluation. Viruses. 2020; 12(1):109. https://doi.org/10.3390/v12010109
Chicago/Turabian StyleLi, Yike, Xiaofen Huang, Zhigang Zhang, Shaowei Li, Jun Zhang, Ningshao Xia, and Qinjian Zhao. 2020. "Prophylactic Hepatitis E Vaccines: Antigenic Analysis and Serological Evaluation" Viruses 12, no. 1: 109. https://doi.org/10.3390/v12010109
APA StyleLi, Y., Huang, X., Zhang, Z., Li, S., Zhang, J., Xia, N., & Zhao, Q. (2020). Prophylactic Hepatitis E Vaccines: Antigenic Analysis and Serological Evaluation. Viruses, 12(1), 109. https://doi.org/10.3390/v12010109