Hepatitis B Virus (HBV) Subviral Particles as Protective Vaccines and Vaccine Platforms


Abstract
1. Introduction
2. Hepatitis B Virus, Classification, and Gene Products
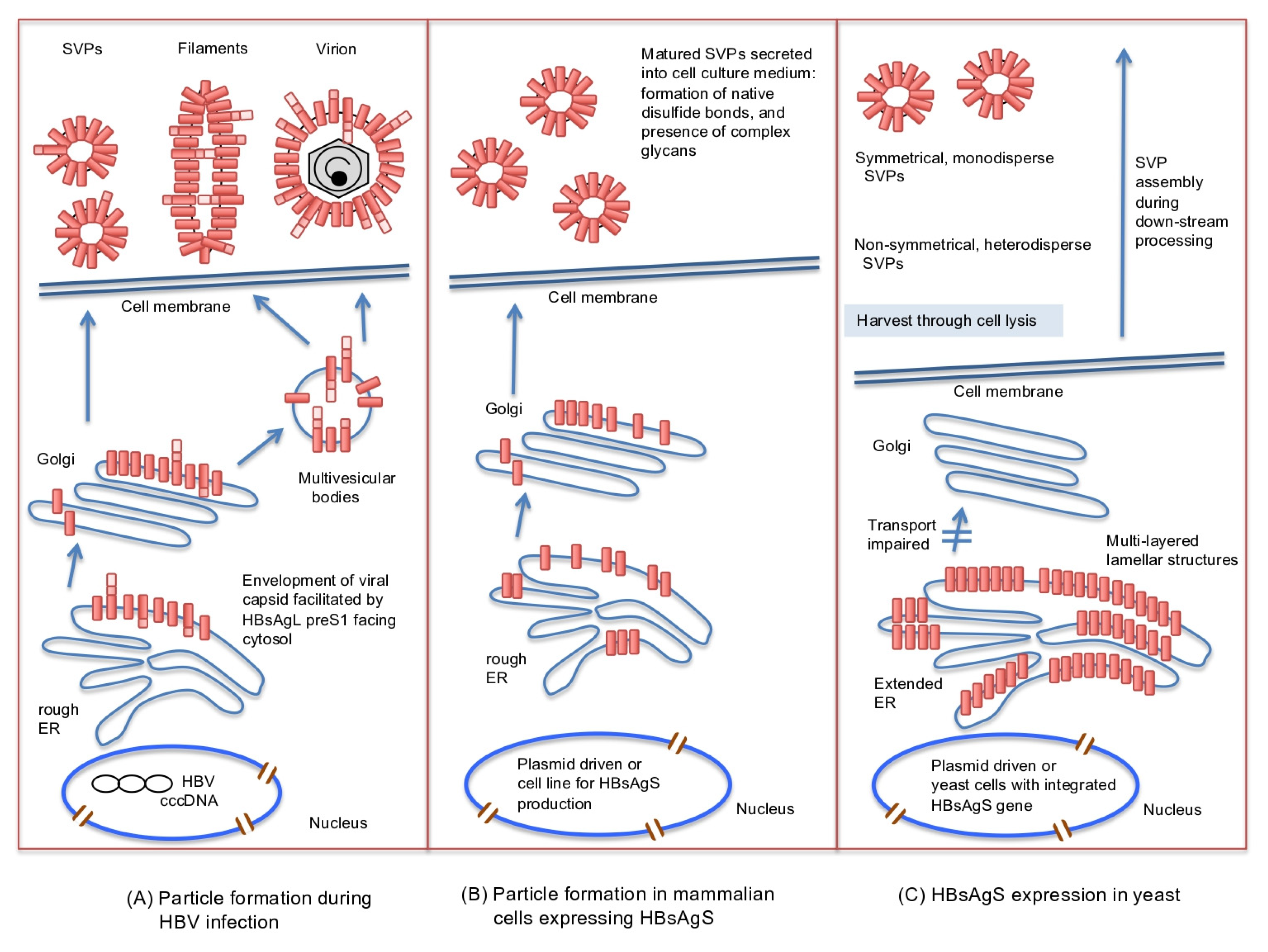
3. Role of HBsAg in HBV, Filament, and SVP Formation
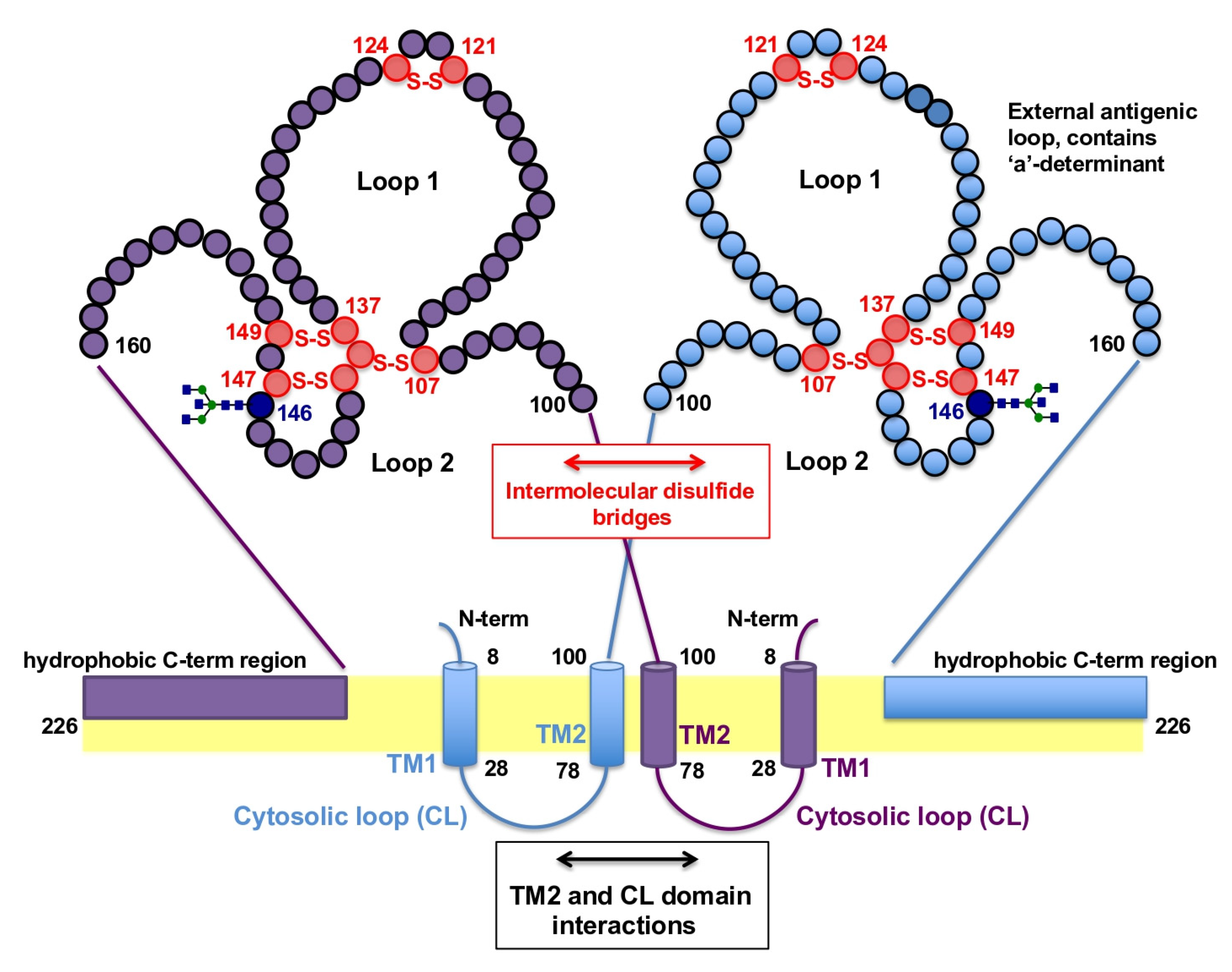
3.1. Topology of HBsAgS
3.2. Topology of HBsAgM
3.3. Topology of HBsAgL
4. Biochemical Properties of SVPs
5. The First Vaccine Generation Against HBV: Vaccine Derived from Patient Plasma
6. The Second Vaccine Generation Against HBV: Yeast-Derived Recombinant HBsAgS SVPs
7. Third Generation Vaccine Concepts against HBV
8. HBsAgS SVPs as Platforms for Medically Relevant Antigenic Sequences
9. Enhancement of Platform Immunogenicity through Biochemical Modifications
10. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Huzair, F.; Sturdy, S. Biotechnology and the transformation of vaccine innovation: The case of the hepatitis B vaccines 1968–2000. Stud. Hist. Philos. Biol. Biomed. Sci. 2017, 64, 11–21. [Google Scholar] [CrossRef]
- Millman, I. The development of the hepatitis B vaccine. In Hepatitis B. The Virus, the Disease, and the Vaccine; Millman, I., Eisenstein, T.K., Blumberg, B.S., Eds.; Plenum Publishing Corp.: New York, NY, USA, 1984; pp. 137–147. [Google Scholar]
- World Health Organization. Hepatitis B. Key Facts; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- World Health Organization. Prevention and Control of Viral Hepatitis Infection. Framework for Global Action; World Health Organization: Geneva, Switzerland, 2012. [Google Scholar]
- Adepoju, P. RTS, S malaria vaccine pilots in three African countries. Lancet 2019, 393, 1685. [Google Scholar] [CrossRef]
- Schuerman, L. RTS, S malaria vaccine could provide major public health benefits. Lancet 2019, 394, 735–736. [Google Scholar] [CrossRef]
- Cohen, J.; Nussenzweig, V.; Nussenzweig, R.; Vekemans, J.; Leach, A. From the circumsporozoite protein to the RTS, S/AS candidate vaccine. Hum. Vaccines 2010, 6, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Mohsen, M.O.; Zha, L.; Cabral-Miranda, G.; Bachmann, M.F. Major findings and recent advances in virus-like particle (VLP)-based vaccines. Semin. Immunol. 2017, 34, 123–132. [Google Scholar] [CrossRef]
- López-Sagaseta, J.; Malito, E.; Rappuoli, R.; Bottomley, M.J. Self-assembling protein nanoparticles in the design of vaccines. Comput. Struct. Biotechnol. J. 2016, 14, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.; Jiang, X. Recent advancements in combination subunit vaccine development. Hum. Vaccines Immunother. 2017, 13, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Yuen, M.-F.; Chen, D.-S.; Dusheiko, G.M.; Janssen, H.L.A.; Lau, D.T.Y.; Locarnini, S.A.; Peters, M.G.; Lai, C.-L. Hepatitis B virus infection. Nat. Rev. Dis. Primers 2018, 4, 18035. [Google Scholar] [CrossRef] [PubMed]
- Seeger, C.; Mason, W.S. Molecular biology of hepatitis B virus infection. Virology 2015, 479–480, 672–686. [Google Scholar] [CrossRef]
- Gerlich, W.H. Medical virology of hepatitis B: How it began and where we are now. Virol. J. 2013, 10, 239. [Google Scholar] [CrossRef]
- Kramvis, A. Genotypes and genetic variability of hepatitis B virus. Intervirology 2014, 57, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Norder, H.; Couroucé, A.M.; Coursaget, P.; Echevarria, J.M.; Lee, S.D.; Mushahwar, I.K.; Robertson, B.H.; Locarnini, S.; Magnius, L.O. Genetic diversity of hepatitis B virus strains derived worldwide: Genotypes, subgenotypes and HBsAg subtypes. Intervirology 2004, 47, 289–309. [Google Scholar] [CrossRef] [PubMed]
- Slagle, B.L.; Bouchard, M.J. Role of HBx in hepatitis B virus persistence and its therapeutic implications. Curr. Opin. Virol. 2018, 30, 32–38. [Google Scholar] [CrossRef]
- Bruss, V. Hepatitis B virus morphogenesis. World J. Gastroenterol. 2007, 13, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Le Bouvier, G.L. The heterogeneity of Australia antigen. J. Infect. Dis. 1971, 123, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Bancroft, W.H.; Mundon, F.K.; Russell, P.K. Detection of additional antigenic determinants of hepatitis B antigen. J. Immunol. 1972, 109, 842–848. [Google Scholar]
- Jazayeri, S.M.; Alavian, S.M.; Dindoost, P.; Thomas, H.C.; Karayiannis, P. Molecular variants of hepatitis B surface antigen (HBsAg). In Viral Hepatitis, 4th ed.; Thomas, H.C., Lok, A.S.F., Locarnini, S.A., Zuckerman, A.J., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2014; pp. 107–126. [Google Scholar]
- Désiré, N.; Ngo, Y.; Franetich, J.-F.; Dembele, L.; Mazier, D.; Vaillant, J.-C.; Poynard, T.; Thibault, V. Definition of an HBsAg to DNA international unit conversion factor by enrichment of circulating hepatitis B forms. J. Viral Hepat. 2015, 22, 718–726. [Google Scholar]
- Brunetto, M.R. A new role for an old marker, HBsAg. J. Hepatol. 2010, 52, 475–477. [Google Scholar] [CrossRef]
- Dreesman, G.R.; Hollinger, F.B.; Suriano, J.R.; Fujioka, R.S.; Brunschwig, J.P.; Melnick, J.L. Biophysical and biochemical heterogeneity of purified hepatitis B antigen. J. Virol. 1972, 10, 469–476. [Google Scholar] [CrossRef]
- Dane, D.S.; Cameron, C.H.; Briggs, M. Virus-like particles in serum of patients with Australia-antigen associated hepatitis. Lancet 1970, 1, 695–698. [Google Scholar] [CrossRef]
- Rydell, G.E.; Prakash, K.; Norder, H.; Lindh, M. Hepatitis B surface antigen on subviral particles reduces the neutralizing effect of anti-HBs antibodies on hepatitis B viral particles in vitro. Virology 2017, 509, 67–70. [Google Scholar] [CrossRef] [PubMed]
- Hilleman, M.R.; Bertland, A.U.; Buynak, E.B.; Lampson, G.P.; McAleer, W.J.; McLean, A.A.; Roehm, R.R.; Tytell, A.A. Clinical and laboratory studies of HBsAg vaccine. In Viral Hepatitis; Vyas, G.N., Cohen, S.N., Schmid, R., Eds.; The Franklin Institute Press: Philadelphia, PA, USA, 1978; pp. 525–537. [Google Scholar]
- Maupas, P.; Goudeau, A.; Coursaget, P.; Drucker, J.; Barin, F.; André, M. Immunization against hepatitis B in man: A pilot study of two years’ duration. In Viral Hepatitis; Vyas, G.N., Cohen, S.N., Schmid, R., Eds.; The Franklin Institute Press: Philadelphia, PA, USA, 1978; pp. 539–556. [Google Scholar]
- McAleer, W.J.; Buynak, E.B.; Maigetter, R.Z.; Wampler, D.E.; Miller, W.J.; Hilleman, M.R. Human hepatitis B vaccine from recombinant yeast. Nature 1984, 307, 178–180. [Google Scholar] [CrossRef] [PubMed]
- Murphy, B.L.; Maynard, J.E.; Le Bouvier, G.L. Viral subtypes and cross-presentation in hepatitis B virus infections of chimpanzees. Intervirology 1974, 3, 378–381. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Liu, K. Complete and incomplete hepatitis B virus particles: Formation, function, and application. Viruses 2017, 9, 56. [Google Scholar] [CrossRef]
- Selzer, L.; Zlotnick, A. Assembly and release of hepatitis B virus. Cold Spring Harb. Perspect. Med. 2015, 5, a021394. [Google Scholar] [CrossRef]
- Sureau, C.; Salisse, J. A conformational heparan sulfate binding site essential to infectivity overlaps with the conserved hepatitis B virus a-determinant. Hepatology 2013, 57, 985–994. [Google Scholar] [CrossRef]
- Bruss, V.; Vieluf, K. Functions of the internal pre-S domain of the large surface protein in hepatitis B virus particle morphogenesis. J. Virol. 1995, 69, 6652–6657. [Google Scholar] [CrossRef]
- Schulze, A.; Gripon, P.; Urban, S. Hepatitis B virus infection initiates with a large surface protein-dependent binding to heparan sulfate proteoglycans. Hepatology 2007, 46, 1759–1768. [Google Scholar] [CrossRef]
- Leistner, C.M.; Gruen-Bernhard, S.; Glebe, D. Role of glycosaminoglycans for binding and infection of hepatitis B virus. Cell. Microbiol. 2008, 10, 122–133. [Google Scholar] [CrossRef]
- Neurath, A.R.; Kent, S.B.H.; Strick, N.; Parker, K. Identification and chemical synthesis of a host cell receptor binding site on hepatitis B virus. Cell 1986, 46, 429–436. [Google Scholar] [CrossRef]
- Yan, H.; Zhong, G.; Xu, G.; He, W.; Jing, Z.; Gao, Z.; Huang, Y.; Qi, Y.; Peng, B.; Wang, H.; et al. Sodium taurocholate cotransporting polypeptide is a functional receptor for human hepatitis B and D virus. eLife 2012, 1, e00049. [Google Scholar] [CrossRef]
- Lambert, C.; Prange, R. Chaperone action in the posttranslational topological reorientation of the hepatitis B virus large envelope protein: Implications for translocational regulation. Proc. Natl. Acad. Sci. USA 2003, 100, 5199–5204. [Google Scholar] [CrossRef] [PubMed]
- Cho, D.Y.; Yang, G.H.; Ryu, C.J.; Hong, H.J. Molecular chaperone GRP78/BiP interacts with the large surface protein of hepatitis B virus in vitro and in vivo. J. Virol. 2003, 77, 2784–2788. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Löffler-Mary, H.; Werr, M.; Prange, R. Sequence-specific repression of cotranslational translocation of the hepatitis B virus envelope proteins coincides with binding of heat shock protein Hsc70. Virology 1997, 235, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Stieler, J.T.; Prange, R. Involvement of ESCRT-II in hepatitis B virus morphogenesis. PLoS ONE 2014, 9, e91279. [Google Scholar] [CrossRef]
- Lambert, C.; Döring, T.; Prange, R. Hepatitis B virus maturation is sensitive to functional inhibition of ESCRT-III, Vps4, and γ2-Adaptin. J. Virol. 2007, 81, 9050–9060. [Google Scholar] [CrossRef]
- Watanabe, T.; Sorensen, E.M.; Naito, A.; Schott, M.; Kim, S.; Ahlquist, P. Involvement of host cellular multivesicular body functions in hepatitis B virus budding. Proc. Natl. Acad. Sci. USA 2007, 104, 10205–10210. [Google Scholar] [CrossRef]
- Jiang, B.; Himmelsbach, K.; Ren, H.; Boller, K.; Hildt, E. Subviral hepatitis B virus filaments, like infectious viral particles, are released via multivesicular bodies. J. Virol. 2016, 90, 3330–3341. [Google Scholar] [CrossRef]
- Chairez, R.; Hollinger, F.B.; Melnick, J.L.; Dreesman, G.R. Biophysical properties of purified morphologic forms of hepatitis B antigen. Intervirology 1974, 3, 129–140. [Google Scholar] [CrossRef]
- Short, J.M.; Chen, S.; Roseman, A.M.; Butler, P.J.G.; Crowther, R.A. Structure of hepatitis B surface antigen from subviral tubes determined by electron cryomicroscopy. J. Mol. Biol. 2009, 390, 135–141. [Google Scholar] [CrossRef]
- Dryden, K.A.; Wieland, S.F.; Whitten-Bauer, C.; Gerin, J.L.; Chisari, F.V.; Yeager, M. Native hepatitis B virions and capsids visualized by electron cryomicroscopy. Mol. Cell 2006, 22, 843–850. [Google Scholar] [CrossRef]
- Cao, J.; Zhang, J.; Lu, Y.; Luo, S.; Zhang, J.; Zhu, P. Cryo-EM structure of native spherical subviral particles isolated from HBV carriers. Virus Res. 2019, 259, 90–96. [Google Scholar] [CrossRef]
- Gilbert, R.J.C.; Beales, L.; Blond, D.; Simon, M.N.; Lin, B.Y.; Chisari, F.V.; Stuart, D.I.; Rowlands, D.J. Hepatitis B small surface antigen particles are octahedral. Proc. Natl. Acad. Sci. USA 2005, 102, 14783–14788. [Google Scholar] [CrossRef] [PubMed]
- Crowley, C.W.; Liu, C.-C.; Levinson, A.D. Plasmid-directed synthesis of hepatitis B surface antigen in monkey cells. Moll. Cell. Biol. 1983, 3, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-C.; Yansura, D.; Levinson, A.D. Direct expression of hepatitis B surface antigen in monkey cells from an SV40 vector. DNA 1982, 1, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Moriarty, A.M.; Hoyer, B.H.; Shih, J.W.K.; Gerin, J.L.; Hamer, D.H. Expression of the hepatitis B virus surface antigen gene in cell culture by using a simian virus 40 vector. Proc. Natl. Acad. Sci. USA 1981, 78, 2606–2610. [Google Scholar] [CrossRef] [PubMed]
- Dubois, M.F.; Pourcel, C.; Rousset, S.; Chany, C.; Tiollais, P. Excretion of hepatitis B surface antigen particles from mouse cells transformed with cloned viral DNA. Proc. Natl. Acad. Sci. USA 1980, 77, 4549–4553. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Kim, B.K.; Choi, E.-C. Physiochemical properties of recombinant hepatitis B surface antigen expressed in mammalian cell (C127). Arch. Pharm. Res. 1998, 21, 521–526. [Google Scholar] [CrossRef]
- Eble, B.E.; Lingappa, V.R.; Ganem, D. Hepatitis B surface antigen: An unusual secreted protein initially synthesized as a transmembrane polypeptide. Mol. Cell. Biol. 1986, 6, 1454–1463. [Google Scholar] [CrossRef]
- Eble, B.E.; MacRae, D.R.; Lingappa, V.R.; Ganem, D. Multiple topogenic sequences determine the transmembrane orientation of hepatitis B surface antigen. Mol. Cell. Biol. 1987, 7, 3591–3601. [Google Scholar] [CrossRef]
- Suffner, S.; Gerstenberg, N.; Patra, M.; Ruibal, P.; Orabi, A.; Schindler, M.; Bruss, V. Domains of the hepatitis B virus small surface protein S mediating oligomerization. J. Virol. 2018, 92, e02232-17. [Google Scholar] [CrossRef] [PubMed]
- Stirk, H.J.; Thornton, J.M.; Howard, C.R. A topological model for hepatitis B surface antigen. Intervirology 1992, 33, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Berting, A.; Hahnen, J.; Kröger, M.; Gerlich, W.H. Computer-aided studies on the spatial structure of the small hepatitis B surface protein. Intervirology 1995, 38, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Komla-Soukha, I.; Sureau, C. A tryptophan-rich motif in the carboxyl terminus of the small envelope protein of hepatitis B virus is central to the assembly of hepatitis delta virus particles. J. Virol. 2006, 80, 4648–4655. [Google Scholar] [CrossRef] [PubMed]
- Paulij, W.P.; de Wit, P.L.M.; Sünnen, C.M.G.; van Roosmalen, M.H.; Petersen-van Ettekoven, A.; Cooreman, M.P.; Heijtink, R.A. Localization of a unique hepatitis B virus epitope sheds new light on the structure of hepatitis B virus surface antigen. J. Gen. Virol. 1999, 80, 2121–2126. [Google Scholar] [CrossRef] [PubMed]
- Mangold, C.M.T.; Unckell, F.; Werr, M.; Streeck, R.E. Analysis of intermolecular disulfide bonds and free sulfhydryl groups in hepatitis B surface antigen particles. Arch. Virol. 1997, 142, 2257–2267. [Google Scholar] [CrossRef] [PubMed]
- Mangold, C.M.T.; Streeck, R.E. Mutational analysis of the cysteine residues in the hepatitis B virus small envelope protein. J. Virol. 1993, 67, 4588–4597. [Google Scholar] [CrossRef]
- Mangold, C.M.T.; Unckell, F.; Werr, M.; Streeck, R.E. Secretion and antigenicity of hepatitis B virus small envelope proteins lacking cysteines in the major antigenic region. Virology 1995, 211, 535–543. [Google Scholar] [CrossRef]
- Cheong, W.S.; Hyakumura, M.; Yuen, L.; Warner, N.; Locarnini, S.; Netter, H.J. Modulation of the immunogenicity of virus-like particles composed of mutant hepatitis B virus envelope subunits. Antivir. Res. 2012, 93, 209–218. [Google Scholar] [CrossRef]
- Wunderlich, G.; Bruss, V. Characterization of early hepatitis B virus surface protein oligomers. Arch. Virol. 1996, 141, 1191–1205. [Google Scholar] [CrossRef]
- Patzer, E.J.; Nakamura, G.R.; Yaffe, A. Intracellular transport and secretion of hepatitis B surface antigen in mammalian cells. J. Virol. 1984, 51, 346–353. [Google Scholar] [CrossRef]
- Lu, X.; Mehta, A.; Dwek, R.; Butters, T.; Block, T. Evidence that N-linked glycosylation is necessary for hepatitis B virus secretion. Virology 1995, 213, 660–665. [Google Scholar] [CrossRef] [PubMed]
- Ito, K.; Qin, Y.; Guarnieri, M.; Garcia, T.; Kwei, K.; Mizokami, M.; Zhang, J.; Li, J.; Wands, J.R.; Tong, S. Impairment of hepatitis B virus virion secretion by single-amino-acid substitutions in the small envelope protein and rescue by a novel glycosylation site. J. Virol. 2010, 84, 12850–12861. [Google Scholar] [CrossRef] [PubMed]
- Julithe, R.; Abou-Jaoudé, G.; Sureau, C. Modification of the hepatitis B virus envelope protein glycosylation pattern interferes with secretion of viral particles, infectivity, and susceptibility to neutralizing antibodies. J. Virol. 2014, 88, 9049–9059. [Google Scholar] [CrossRef] [PubMed]
- Eble, B.E.; Lingappa, V.R.; Ganem, D. The N-terminal (pre-S2) domain of a hepatitis B virus surface glycoprotein is translocated across membranes by downstream signal sequences. J. Virol. 1990, 64, 1414–1419. [Google Scholar] [CrossRef] [PubMed]
- Stibbe, W.; Gerlich, W.H. Characterization of pre-s gene products in hepatitis B surface antigen. Dev. Biol. Stand. 1983, 54, 33–43. [Google Scholar]
- Schmitt, S.; Glebe, D.; Tolle, T.K.; Lochnit, G.; Linder, D.; Geyer, R.; Gerlich, W.H. Structure of pre-S2 N- and O-linked glycans in surface proteins from different genotypes of hepatitis B virus. J. Gen. Virol. 2004, 85, 2045–2053. [Google Scholar] [CrossRef]
- Cheng, K.-C.; Moss, B. Selective synthesis and secretion of particles composed of the hepatitis B virus middle surface protein directed by a recombinant vaccinia virus: Induction of antibodies to pre-S and S epitopes. J. Virol. 1987, 61, 1286–1290. [Google Scholar] [CrossRef]
- McLachlan, A.; Milich, D.R.; Raney, A.K.; Riggs, M.G.; Hughes, J.L.; Sorge, J.; Chisari, F.V. Expression of hepatitis B virus surface and core antigens: Influences of pre-S and precore sequences. J. Virol. 1987, 61, 683–692. [Google Scholar] [CrossRef]
- Molnar-Kimber, K.L.; Jarocki-Witek, V.; Dheer, S.K.; Vernon, S.K.; Conley, A.J.; Davis, A.R.; Hung, P.P. Distinctive properties of the hepatitis B virus envelope proteins. J. Virol. 1988, 62, 407–416. [Google Scholar] [CrossRef]
- Sheu, S.Y.; Lo, S.J. Biogenesis of the hepatitis B viral middle (M) surface protein in a human hepatoma cell line: Demonstration of an alternative secretion pathway. J. Gen. Virol. 1994, 75, 3031–3039. [Google Scholar] [CrossRef] [PubMed]
- Fernholz, D.; Galle, P.R.; Stemler, M.; Brunetto, M.; Bonino, F.; Will, H. Infectious hepatitis B virus variant defective in pre-S2 protein expression in a chronic carrier. Virology 1993, 194, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Ostapchuk, P.; Hearing, P.; Ganem, D. A dramatic shift in the transmembrane topology of a viral envelope glycoprotein accompanies hepatitis B viral morphogenesis. EMBO J. 1994, 13, 1048–1057. [Google Scholar] [CrossRef] [PubMed]
- Prange, R.; Streeck, R.E. Novel transmembrane topology of the hepatitis B virus envelope proteins. EMBO J. 1995, 14, 247–256. [Google Scholar] [CrossRef]
- Seitz, S.; Iancu, C.; Volz, T.; Mier, W.; Dandri, M.; Urban, S.; Bartenschlager, R. A slow maturation process renders hepatitis B virus infectious. Cell Host Microbe 2016, 20, 25–35. [Google Scholar] [CrossRef]
- Ni, Y.; Sonnabend, J.; Seitz, S.; Urban, S. The pre-S2 domain of the hepatitis B virus is dispensable for infectivity but serves a spacer function for L-protein-connected virus assembly. J. Virol. 2010, 84, 3879–3888. [Google Scholar] [CrossRef]
- Bruss, V.; Gerhardt, E.; Vieluf, K.; Wunderlich, G. Functions of the large hepatitis B virus surface protein in viral particle morphogenesis. Intervirology 1996, 39, 23–31. [Google Scholar] [CrossRef]
- Gripon, P.; Le Seyec, J.; Rumin, S.; Guguen-Guillouzo, C. Myristylation of the hepatitis B virus large surface protein is essential for viral infectivity. Virology 1995, 213, 292–299. [Google Scholar] [CrossRef]
- Xu, Z.; Bruss, V.; Yen, T.S.B. Formation of intracellular particles by hepatitis B virus large surface protein. J. Virol. 1997, 71, 5487–5494. [Google Scholar] [CrossRef]
- Persing, D.H.; Varmus, H.E.; Ganem, D. Inhibition of secretion of hepatitis B surface antigen by a related presurface polypeptide. Science 1986, 234, 1388–1391. [Google Scholar] [CrossRef]
- Ou, J.H.; Rutter, W.J. Regulation of secretion of the hepatitis B virus major surface antigen by the preS-1 protein. J. Virol. 1987, 61, 782–786. [Google Scholar] [CrossRef] [PubMed]
- Heermann, K.H.; Goldmann, U.; Schwartz, W.; Seyffarth, T.; Baumgarten, H.; Gerlich, W.H. Large surface proteins of hepatitis B virus containing the pre-s sequence. J. Virol. 1984, 52, 396–402. [Google Scholar] [CrossRef] [PubMed]
- Gavilanes, F.; Gonzalez-Ros, J.M.; Peterson, D.L. Structure of hepatitis B surface antigen. J. Biol. Chem. 1982, 257, 7770–7777. [Google Scholar] [PubMed]
- Peterson, D.L. The structure of hepatitis B surface antigen and its antigenic sites. BioEssays 1987, 6, 258–262. [Google Scholar] [CrossRef]
- Shih, C.; Li, L.S.; Roychoudhury, S.; Ho, M.H. In vitro propagation of human hepatitis B virus in a rat hepatoma cell line. Proc. Natl. Acad. Sci. USA 1989, 86, 6323–6327. [Google Scholar] [CrossRef]
- Hruska, J.F.; Robinson, W.S. The proteins of hepatitis B Dane particle cores. J. Med. Virol. 1977, 1, 119–131. [Google Scholar] [CrossRef]
- Sukeno, N.; Shirachi, R.; Yamaguchi, J.; Ishida, N. Reduction and reoxidation of Australia antigen: Loss and reconstitution of particle structure and antigenicity. J. Virol. 1972, 9, 182–183. [Google Scholar] [CrossRef]
- Vyas, G.N.; Rao, K.R.; Ibrahim, A.B. Australia antigen (hepatitis B antigen): A conformational antigen dependent of disulfide bonds. Science 1972, 178, 1300–1301. [Google Scholar] [CrossRef]
- Imai, M.; Gotoh, A.; Nishioka, K.; Kurashina, S.; Miyakawa, Y.; Mayumi, M. Antigenicity of reduced and alkylated Australia antigen. J. Immunol. 1974, 112, 416–419. [Google Scholar]
- Huovila, A.-P.J.; Eder, A.M.; Fuller, S.D. Hepatitis B surface antigen assembles in a post-ER, pre-Golgi compartment. J. Cell Biol. 1992, 118, 1305–1320. [Google Scholar] [CrossRef]
- Patzer, E.J.; Nakamura, G.R.; Simonsen, C.C.; Levinson, A.D.; Brands, R. Intracellular assembly and packaging of hepatitis B surface antigen particles occur in the endoplasmic reticulum. J. Virol. 1986, 58, 884–892. [Google Scholar] [CrossRef] [PubMed]
- Satoh, O.; Umeda, M.; Imai, H.; Tunoo, H.; Inoue, K. Lipid composition of hepatitis B surface antigen particles and the particle-producing human hepatoma cell lines. J. Lipid Res. 1990, 31, 1293–1300. [Google Scholar] [PubMed]
- Satoh, O.; Imai, H.; Yoneyama, T.; Miyamura, T.; Utsumi, H.; Inoue, K.; Umeda, M. Membrane structure of the hepatitis B virus surface antigen particle. J. Biochem. 2000, 127, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Gavilanes, F.; Gomez-Gutierrez, J.; Aracil, M.; Gonzales-Ros, J.M.; Ferragut, J.A.; Guerrero, E.; Peterson, D.L. Hepatitis B surface antigen. Role of lipids in maintaining the structural and antigenic properties of protein components. Biochem. J. 1990, 265, 857–864. [Google Scholar] [CrossRef]
- Guerrero, E.; Gavilanes, F.; Peterson, D.L. Model for the protein arrangement in HBsAg particles based on physical and chemical studies. In Viral Hepatitis and Liver Disease; Zuckerman, A.J., Ed.; Alan, R. Liss Inc.: New York, NY, USA, 1988; pp. 606–613. [Google Scholar]
- Greiner, V.J.; Egelé, C.; Oncul, S.; Ronzon, F.; Manin, C.; Klymchenko, A.; Mély, Y. Characterization of the lipid and protein organization in HBsAg viral particles by steady-state and time-resolved fluorescence spectroscopy. Biochimie 2010, 92, 994–1002. [Google Scholar] [CrossRef]
- Milhiet, P.-E.; Dosset, P.; Godefroy, C.; Le Grimellec, C.; Guigner, J.M.; Larquet, E.; Ronzon, F.; Manin, C. Nanoscale topography of hepatitis B antigen particles by atomic force microscopy. Biochimie 2011, 93, 254–259. [Google Scholar] [CrossRef]
- Blumberg, B.S. Australia antigen and the biology of hepatitis B. Science 1977, 197, 17–25. [Google Scholar] [CrossRef]
- Couroucé-Pauty, A.M.; Delons, S.; Soulier, J.P. Attempt to prevent hepatitis by using specific anti-HBs immunoglobulin. Am. J. Med. Sci. 1975, 270, 375–383. [Google Scholar]
- Iwarson, S.; Kjellman, H.; Ahlmén, J.; Ljunggren, C.; Eriksson, E.; Selander, D.; Hermodsson, S. Hepatitis B immune serum globulin and standard gamma globulin in prevention of hepatitis B infection among hospital staff: A preliminary report. Am. J. Med. Sci. 1975, 270, 385–389. [Google Scholar] [CrossRef]
- Hilleman, M.R.; Buynak, E.B.; Roehm, R.R.; Tytell, A.A.; Bertland, A.U.; Lampson, G.P. Purified and inactivated human hepatitis B vaccine: Progress report. Am. J. Med. Sci. 1975, 270, 401–404. [Google Scholar] [CrossRef]
- Purcell, R.H.; Gerin, J.L. Hepatitis B subunit vaccine: A preliminary report of safety and efficacy tests in chimpanzees. Am. J. Med. Sci. 1975, 270, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Okochi, K.; Murakami, S.; Ninomiya, K.; Kaneko, M. Australia antigen, transfusion and hepatitis. Vox Sang. 1970, 18, 289–300. [Google Scholar] [CrossRef]
- London, W.T.; Drew, J.S.; Lustbader, E.D.; Werner, B.G.; Blumberg, B.S. Host responses to hepatitis B infection in patients in a chronic hemodialysis unit. Kidney Int. 1977, 12, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Krugman, S.; Giles, J.P.; Hammond, J. Viral Hepatitis, type B (MS-2 strain). Studies on active immunization. JAMA 1971, 217, 41–45. [Google Scholar] [CrossRef]
- Buynak, E.B.; Roehm, R.R.; Tytell, A.A.; Bertland, A.U., II; Lampson, G.P.; Hilleman, M.R. Vaccine against human hepatitis B. J. Am. Med. Assoc. 1976, 235, 2832–2834. [Google Scholar] [CrossRef]
- Gerin, J.L.; Holland, P.V.; Purcell, R.H. Australia antigen: Large-scale purification from human serum and biochemical studies of its proteins. J. Virol. 1971, 7, 569–576. [Google Scholar] [CrossRef]
- Maupas, P.; Goudeau, A.; Coursaget, P.; Drucker, J.; Bagros, P. Immunisation against hepatitis B in man. Lancet 1976, 307, 1367–1370. [Google Scholar] [CrossRef]
- McMahon, B.J.; Helminiak, C.; Wainwright, R.B.; Bulkow, L.; Trimble, B.A.; Wainwright, K. Frequency of adverse reactions to hepatitis B vaccine in 43,618 persons. Am. J. Med. 1992, 92, 254–256. [Google Scholar] [CrossRef]
- Szmuness, W.; Stevens, C.E.; Zang, E.A.; Harley, E.J.; Kellner, A. A controlled clinical trial of the efficacy of the hepatitis B vaccine (Heptavax B): A final report. Hepatol 1981, 1, 377–385. [Google Scholar] [CrossRef]
- Shaw, F.E.; Graham, D.J.; Guess, H.A.; Milstien, J.B.; Johnson, J.M.; Schatz, G.C.; Hadler, S.C.; Kuritsky, J.N.; Hiner, E.E.; Bregman, D.J.; et al. Postmarketing surveillance for neurologic adverse events reported after hepatitis B vaccination. Experience of the first three years. Am. J. Epidemiol. 1988, 127, 337–352. [Google Scholar] [CrossRef]
- Neurath, A.R.; Strick, N.; Kent, S.B.H.; Offensperger, W.; Wahl, S.; Christman, J.K.; Acs, G. Enzyme-linked immunoassay of pre-S gene-coded sequences in hepatitis B vaccines. J. Virol. Method 1985, 12, 185–192. [Google Scholar] [CrossRef]
- Neurath, A.R.; Kent, S.B.H.; Strick, N.; Taylor, P.; Stevens, C.E. Hepatitis B virus contains pre-S gene-encoded domains. Nature 1985, 315, 154–156. [Google Scholar] [CrossRef] [PubMed]
- Coursaget, P.; Bringer, L.; Sarr, G.; Bourdil, C.; Fritzell, B.; Blondeau, C.; Yvonnet, B.; Chiron, J.P.; Jeannée, E.; Guindo, S.; et al. Comparative immunogenicity in children of mammalian cell-derived recombinant hepatitis B vaccine and plasma-derived hepatitis B vaccine. Vaccine 1992, 10, 379–382. [Google Scholar] [CrossRef]
- Zuckerman, A.J. Hepatitis-B vaccine. Safety criteria and non-B infection. Lancet 1976, 307, 1396–1397. [Google Scholar] [CrossRef]
- Hilleman, M.R.; Ellis, R. Vaccines made from recombinant yeast cells. Vaccine 1986, 4, 75–76. [Google Scholar] [CrossRef]
- Stephenne, J. Recombinant versus plasma-derived hepatitis B vaccines: Issues of safety, immunogenicity and cost-effectiveness. Vaccine 1988, 6, 299–303. [Google Scholar] [CrossRef]
- Coursaget, P.; Adamowicz, P.; Bourdil, C.; Yvonnet, B.; Buisson, Y.; Barrès, J.L.; Saliou, P.; Chiron, J.P.; Diop Mar, I. Anti-preS2 antibodies in natural hepatitis B virus infection and after immunization. Vaccine 1988, 6, 357–361. [Google Scholar] [CrossRef]
- Lau, J.Y.N.; Lai, C.L.; Wu, P.C.; Lin, H.J. Comparison of two plasma-derived hepatitis B vaccines: Long-term report of a prospective, randomized trial. J. Gastroenterol. Hepatol. 1989, 4, 331–337. [Google Scholar] [CrossRef]
- Lee, H.S.; Kim, C.Y. Seroepidemiology of HBV and HCV in Korea: The decreasing prevalence rate of HBV infection after launching HB vaccination program. Int. Hepatol. Commun. 1996, 5, 53–61. [Google Scholar] [CrossRef]
- Kalayanarooj, S.; Vaughn, D.W.; Ariyasriwatana, C.; Snitbhan, R. Protective antibody after a ‘one dollar’ hepatitis B vaccine. Southeast Asian J. Trop. Med. Public Health 1996, 27, 659–663. [Google Scholar]
- Schoub, B.D.; Matai, U.; Singh, B.; Blackburn, N.K.; Levin, J.B. Universal immunization of infants with low doses of a low-cost, plasma-derived hepatitis B vaccine in South Africa. Bull. World Health Organ. 2002, 80, 277–281. [Google Scholar] [PubMed]
- Coates, T.; Wilson, R.; Patrick, G.; André, F.; Watson, V. Hepatitis B vaccines: Assessment of the seroprotective efficacy of two recombinant DNA vaccines. Clin. Ther. 2001, 23, 392–403. [Google Scholar] [CrossRef]
- Hernández-Bernal, F.; Aguilar-Betancourt, A.; Aljovin, V.; Arias, G.; Valenzuela, C.; de Alejo, K.P.; Hernández, K.; Oquendo, O.; Figueredo, N.; Figueroa, N.; et al. Comparison of four recombinant hepatitis B vaccines applied on an accelerated schedule in healthy adults. Hum. Vaccine 2011, 7, 1026–1036. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Iino, S.; Shiraki, K.; Akahane, Y.; Okamoto, H.; Domoto, K.; Mishiro, S. Safety and efficacy of a recombinant yeast-derived pre-S2 + S-containing hepatitis B vaccine (TGP-943): Phase 1, 2 and 3 clinical testing. Vaccine 1994, 12, 1090–1096. [Google Scholar] [CrossRef]
- Teles, S.A.; Martins, R.M.B.; Lopes, C.L.R.; dos Santos Carneiro, M.A.; Souza, K.P.; Yoshida, C.F.T. Immunogenicity of a recombinant hepatitis B vaccine (Euvax-B) in haemodialysis patients and staff. Eur.J. Epidemiol. 2001, 17, 145–149. [Google Scholar] [CrossRef]
- Abraham, P.; Mistry, F.P.; Bapat, M.R.; Sharma, G.; Reddy, G.R.; Prasad, K.S.N.; Ramanna, V. Evaluation of a new recombinant DNA hepatitis B vaccine (Shanvac-B). Vaccine 1999, 17, 1125–1129. [Google Scholar] [CrossRef]
- Anonymous. A two-dose hepatitis B vaccine for adults (Heplisav-B). Med. Lett. Drugs Ther. 2018, 60, 17–18. [Google Scholar]
- Zhu, F.; Deckx, H.; Roten, R.; Michiels, B.; Sarnecki, M. Comparative efficacy, safety and immunogenicity of Hepavax-Gene TF and Engerix-B recombinant hepatitis B vaccines in neonates in China. Pediatr. Infect. Dis. J. 2017, 36, 94–101. [Google Scholar] [CrossRef]
- Shouval, D.; Roggendorf, H.; Roggendorf, M. Enhanced immune response to hepatitis B vaccination through immunization with a Pre-S1/Pre-S2/S vaccine. Med. Microbiol. Immunol. 2015, 204, 57–68. [Google Scholar] [CrossRef]
- Young, M.D.; Schneider, D.L.; Zuckerman, A.J.; Du, W.; Dickson, B.; Maddrey, W.C. Adult hepatitis B vaccination using a novel triple antigen recombinant vaccine. Hepatology 2001, 34, 372–376. [Google Scholar] [CrossRef]
- Valenzuela, P.; Medina, A.; Rutter, W.J.; Ammerer, G.; Hall, B.D. Synthesis and assembly of hepatitis B virus surface antigen particles in yeast. Nature 1982, 298, 347–350. [Google Scholar] [CrossRef] [PubMed]
- Harford, N.; Cabezon, T.; Crabeel, M.; Simoen, E.; Rutgers, A.; De Wilde, M. Expression of hepatitis B surface antigen in yeast. Dev. Biol. Stand. 1983, 54, 125–130. [Google Scholar] [PubMed]
- Miyanohara, A.; Toh-e, A.; Nozaki, C.; Hamada, F.; Ohtomo, N.; Matsubara, K. Expression of hepatitis B surface antigen gene in yeast. Proc. Natl. Acad. Sci. USA 1983, 80, 1–5. [Google Scholar] [CrossRef] [PubMed]
- André, F.E. Overview of a 5-year clinical experience with a yeast-derived hepatitis B vaccine. Vaccine 1990, 8 (Suppl. 1), S74–S78. [Google Scholar]
- Wolters, B.; Junge, U.; Dziuba, S.; Roggendorf, M. Immunogenicity of combined hepatitis A and B vaccine in elderly persons. Vaccine 2003, 21, 3623–3628. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, C.; Liu, Z.; Zou, G.; Li, J.; Lu, M. Host genetic determinants of hepatitis B virus infection. Front. Genet. 2019, 10, 696. [Google Scholar] [CrossRef]
- Li, Z.K.; Nie, J.J.; Li, J.; Zhuang, H. The effec t of HLA on immunological response to hepatitis B vaccine in healthy people: A meta-analysis. Vaccine 2013, 31, 4355–4361. [Google Scholar] [CrossRef]
- Milich, D.R.; Leroux-Roels, G.G. Immunogenetics of the response to HBsAg vaccination. Autoimmun. Rev. 2003, 2, 248–257. [Google Scholar] [CrossRef]
- Cregg, J.M.; Tschopp, J.F.; Stillman, C.; Siegel, R.; Akong, M.; Craig, W.S.; Buckholz, R.G.; Madden, K.R.; Kellaris, P.A.; Davis, G.R.; et al. High-level expression and efficient assembly of hepatitis B surface antigen in the methylotrophic yeast, Pichia pastoris. Biotechnology 1987, 5, 479–485. [Google Scholar] [CrossRef]
- Lünsdorf, H.; Gurramkonda, C.; Adnan, A.; Khanna, N.; Rinas, U. Virus-like particle production with yeast: Ultrastructural and immunocytochemical insights into Pichia pastoris producing high levels of the hepatitis B surface antigen. Microb. Cell Factories 2011, 10, 48. [Google Scholar] [CrossRef]
- Zahid, M.; Lünsdorf, H.; Rinas, U. Assessing stability and assembly of the hepatitis B surface antigen into virus-like particles during down-stream processing. Vaccine 2015, 33, 3739–3745. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Wang, Y.; Freed, D.; Fu, T.-M.; Gimenez, J.A.; Sitrin, R.D.; Washabaugh, M.W. Maturation of recombinant hepatitis B virus surface antigen particles. Hum. Vaccines 2006, 2, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Gurramkonda, C.; Zahid, M.; Nemani, S.K.; Adnan, A.; Gudi, S.K.; Khanna, N.; Ebensen, T.; Lünsdorf, H.; Guzmán, C.A.; Rinas, U. Purification of hepatitis B surface antigen virus-like particles from recombinant Pichia pastoris and in vivo analysis of their immunogenic properties. J. Chromatogr. B 2013, 940, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Towne, V.; Brown, M.; Wang, Y.; Abraham, D.; Oswald, C.B.; Gimenez, J.A.; Washabaugh, M.W.; Kennedy, R.; Sitrin, R.D. In-depth process understanding of RECOMBIVAX HB® maturation and potential epitope improvements with redox treatment: Multifaceted biochemical and immunochemical characterization. Vaccine 2011, 29, 7936–7941. [Google Scholar] [CrossRef] [PubMed]
- Wampler, D.E.; Lehman, E.D.; Boger, J.; McAleer, W.J.; Scolnick, E.M. Multiple chemical forms of hepatitis B surface antigen produced in yeast. Proc. Natl. Acad. Sci. USA 1985, 82, 6830–6834. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Sugahara, K.; Shiosaki, K.; Mizokami, H.; Takeo, K. Fine structure of hepatitis B virus surface antigen produced by recombinant yeast: Comparison with HBsAg of human origin. FEMS Microbiol. Lett. 1998, 165, 363–367. [Google Scholar] [CrossRef]
- Sonveaux, N.; Conrath, K.; Capiau, C.; Brasseur, R.; Goormaghtigh, E.; Ruysschaert, J.M. The topology of the S protein in the yeast-derived hepatitis B surface antigen particles. J. Biol. Chem. 1994, 269, 25637–25645. [Google Scholar]
- Van der Meeren, P.; Van Criekinge, W.; Vanderdeelen, J.; Baert, L. Phospholipid composition of r-DNA hepatitis B surface antigens. Int. J. Pharm. 1994, 106, 89–92. [Google Scholar] [CrossRef]
- Diminsky, D.; Schirmbeck, R.; Reimann, J.; Barenholz, Y. Comparison between hepatitis B surface antigen (HBsAg) particles derived from mammalian cells (CHO) and yeast cells (Hansenula polymorpha): Composition, structure and immunogenicity. Vaccine 1997, 15, 637–647. [Google Scholar] [CrossRef]
- Brown, S.E.; Stanley, C.; Howard, C.R.; Zuckerman, A.J.; Steward, M.W. Antibody responses to recombinant and plasma derived hepatitis B vaccines. Br. Med. J. 1986, 292, 159–161. [Google Scholar] [CrossRef]
- Franchi, L.; Núñez, G. The Nlrp3 inflammasome is critical for aluminium hydroxide-mediated IL-1β secretion but dispensible for adjuvant activity. Eur. J. Immunol. 2008, 38, 2085–2089. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Willingham, S.B.; Ting, J.P.-Y.; Re, F. Cutting Edge: Inflammasome activation by alum and alum’s adjuvant effect are mediated by NLRP3. J. Immunol. 2008, 181, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Kool, M.; Pétrilli, V.; De Smedt, T.; Rolaz, A.; Hammad, H.; van Nimwegen, M.; Bergen, I.M.; Castillo, R.; Lambrecht, B.N.; Tschopp, J. Cutting edge: Alum adjuvant stimulates inflammatory dendritic cells through activation of the NALP3 inflammasome. J. Immunol. 2008, 181, 3755–3759. [Google Scholar] [CrossRef] [PubMed]
- HogenEsch, H. Mechanism of immunopotentiation and safety of aluminum adjuvants. Front. Immunol. 2013, 3, 406. [Google Scholar] [CrossRef] [PubMed]
- Iyer, S.; Robinett, R.S.R.; HogenEsch, H.; Hem, S.L. Mechanism of adsorption of hepatitis B surface antigen by aluminum hydroxide adjuvant. Vaccine 2004, 22, 1475–1479. [Google Scholar] [CrossRef] [PubMed]
- Hansen, B.; Belfast, M.; Soung, G.; Song, L.; Egan, P.M.; Capen, R.; HogenEsch, H.; Mancinelli, R.; Hem, S.L. Effect of the strength of adsorption of hepatitis B surface antigen to aluminum hydroxide adjuvant on the immune response. Vaccine 2009, 27, 888–892. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; HogenEsch, H.; Hem, S.L. Change in the degree of adsorption of proteins by aluminum-containing adjuvants following exposure to interstitial fluid: Freshly prepared and aged model vaccines. Vaccine 2002, 20, 80–85. [Google Scholar] [CrossRef]
- Greiner, V.J.; Ronzon, F.; Larquet, E.; Desbat, B.; Estèves, C.; Bonvin, J.; Gréco, F.; Manin, C.; Klymchenko, A.S.; Mély, Y. The structure of HBsAg particles is not modified upon their adsorption on aluminium hydroxide gel. Vaccine 2012, 30, 5240–5245. [Google Scholar] [CrossRef]
- Jackson, S.; Lentino, J.; Kopp, J.; Murray, L.; Ellison, W.; Rhee, M.; Shockey, G.; Akella, L.; Erby, K.; Heyward, W.L.; et al. Immunogenicity of a two-dose investigational hepatitits B vaccine, HBsAg-1018, using a toll-like receptor 9 agonist adjuvant compared with a licensed hepatitis B vaccine in adults. Vaccine 2018, 36, 668–674. [Google Scholar] [CrossRef]
- Neurath, A.R.; Seto, B.; Strick, N. Antibodies to synthetic peptides from the preS1 region of the hepatitis B virus (HBV) envelope (env) protein are virus-neutralizing and protective. Vaccine 1989, 7, 234–236. [Google Scholar] [CrossRef]
- Neurath, A.R.; Kent, S.B.H.; Parker, K.; Prince, A.M.; Strick, N.; Brotman, B.; Sproul, P. Anrtibodies to a synthetic peptide from the preS 120-145 region of the hepatitis B virus envelope are virus-neutralizing. Vaccine 1986, 4, 35–37. [Google Scholar] [CrossRef]
- Itoh, Y.; Takai, E.; Ohnuma, H.; Kitajima, K.; Tsuda, F.; Machida, A.; Mishiro, S.; Nakamura, T.; Miyakawa, Y.; Mayumi, M. A synthetic peptide vaccine involving the product of the pre-S(2) region of hepatitis B virus DNA: Protective efficacy in chimpanzees. Proc. Natl. Acad. Sci. USA 1986, 83, 9174–9178. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, C.; Penna, A.; Bertoletti, A.; Cavalli, A.; Valli, A.; Schianchi, C.; Fiaccadori, F. The preS1 antigen of hepatitis B virus is highly immunogenic at the T cell level in man. J. Clin. Investig. 1989, 84, 1314–1319. [Google Scholar] [CrossRef] [PubMed]
- Milich, D.R. T- and B-cell recognition of hepatitis B viral antigens. Immunol. Today 1988, 9, 380–386. [Google Scholar] [CrossRef]
- Milich, D.R.; McLachlan, A.; Chisari, F.V.; Kent, S.B.H.; Thornton, G.B. Immune response to the pre-S(1) region of the hepatitis B surface antigen (HBsAg): A pre-S(1)-specific T cell response can bypass nonresponsiveness to the pre-S(2) and S regions of HBsAg. J. Immunol. 1986, 137, 315–322. [Google Scholar]
- Shouval, D.; Ilan, Y.; Adler, R.; Deepen, R.; Panet, A.; Even-Chen, Z.; Gorecki, M.; Gerlich, W.H. Improved immunogenicity in mice of a mammalian cell-derived recombinant hepatitis B vaccine containing pre-S1 and pre-S2 antigens as compared with conventional yeast-derived vaccines. Vaccine 1994, 12, 1453–1459. [Google Scholar] [CrossRef]
- Kobayashi, M.; Asano, T.; Utsunomiya, M.; Itoh, Y.; Fijisawa, Y.; Nishimura, O.; Kato, K.; Kakinuma, A. Recombinant hepatitis B virus surface antigen carrying the pre-S2 region derived from yeast: Purification and characterization. J. Biotechnol. 1988, 8, 1–22. [Google Scholar] [CrossRef]
- Itoh, Y.; Fujisawa, Y. Synthesis in yeast of hepatitis B virus surface antigen modified P31 particles by gene modification. Biochem. Biophys. Res. Commun. 1986, 141, 942–948. [Google Scholar] [CrossRef]
- De Wilde, M.; Rutgers, T.; Cabezon, T.; Hauser, P.; van Opstal, O.; Harford, N.; van Wijnendaele, F.; Desmons, P.; Comberbach, M.; Roelants, P.; et al. PreS-containing HBsAg particles from Saccharomyces cerevisiae: Production, antigenicity, and immunogenicity. In Viral Hepatitis and Liver Disease; Hollinger, F.B., Lemon, S.M., Margolis, H., Eds.; Williams & Wilkins: Baltimore, MD, USA, 1991; pp. 732–736. [Google Scholar]
- Kuroda, S.; Fujisawa, Y.; Iino, S.; Akahane, Y.; Suzuki, H. Induction of protection level of anti-pre-S2 antibodies in humans immunized with a novel hepatitis B vaccine consisting of M (pre-S2+S) protein particles (a third generation vaccine). Vaccine 1991, 9, 163–169. [Google Scholar] [CrossRef]
- Fujisawa, Y.; Kuroda, S.; Van Eerd, P.M.C.A.; Schellekens, H.; Kakinuma, A. Protective efficacy of a novel hepatitis B vaccine consisting of M (pre-S2+S) protein particles (a third generation vaccine). Vaccine 1990, 8, 192–198. [Google Scholar] [CrossRef]
- Kniskern, P.J.; Hagopian, A.; Burke, P.; Dunn, N.; Emini, E.A.; Miller, W.J.; Yamazaki, S.; Ellis, R.W. A candidate vaccine for hepatitis B containing the complete viral surface protein. Hepatology 1988, 8, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Biemans, R.; Thines, D.; Rutgers, T.; de Wilde, M.; Cabezon, T. The large surface protein of hepatitis B virus is retained in the yeast endoplasmic reticulum and provokes its unique enlargement. DNA Cell Biol. 1991, 10, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, S.; Otaka, S.; Miyazaki, T.; Nakao, M.; Fujisawa, Y. Hepatitis B virus envelope L protein particles. J. Biol. Chem. 1992, 267, 1953–1961. [Google Scholar] [PubMed]
- Yamada, T.; Iwabuki, H.; Kanno, T.; Tanaka, H.; Kawai, T.; Fukuda, H.; Kondo, A.; Seno, M.; Tanizawa, K.; Kuroda, S. Physicochemical and immunological characterization of hepatitis B virus envelope particels exclusively consisting of the entire L (pre-S1+pre-S2+S) protein. Vaccine 2001, 19, 3154–3163. [Google Scholar] [CrossRef]
- Leroux-Roels, G.; Desombere, I.; De Tollenaere, G.; Petit, M.A.; Desmons, P.; Hauser, P.; Delem, A.; De Grave, D.; Safary, A. Hepatitis B vaccine containing surface antigen and selected preS1 and preS2 sequences. 1. Safety and immunogenicity in young, healthy adults. Vaccine 1997, 15, 1724–1731. [Google Scholar] [CrossRef]
- Leroux-Roels, G.; Desombere, I.; Cobbaut, L.; Petit, M.A.; Desmons, P.; Hauser, P.; Delem, A.; De Grave, D.; Safary, A. Hepatitis B vaccine containing surface antigen and selected preS1 and preS2 sequences. 2. Immunogenicity in poor responders to hepatitis B vaccines. Vaccine 1997, 15, 1732–1736. [Google Scholar] [CrossRef]
- Tron, F.; Degos, F.; Bréchot, C.; Couroucé, A.M.; Goudeau, A.; Marie, F.N.; Adamowicz, P.; Saliou, P.; Laplanche, A.; Benhamou, J.P.; et al. Randomized dose range study of a recombinant hepatitis B vaccine produced in mammalian cells and containing the S and preS2 sequences. J. Infect. Dis. 1989, 160, 199–204. [Google Scholar] [CrossRef]
- Yan, D.; Wei, Y.Q.; Guo, H.C.; Sun, S.Q. The application of virus-like particles as vaccines and biological vehicles. Appl. Microbiol. Biotechnol. 2015, 99, 10415–10432. [Google Scholar] [CrossRef]
- Pushko, P.; Pumpens, P.; Grens, E. Development of virus-like particle technology from small highly symmetric to large complex virus-like particle structures. Intervirology 2013, 56, 141–165. [Google Scholar] [CrossRef]
- Ramasamy, V.; Arora, U.; Shukla, R.; Poddar, A.; Shanmugam, R.K.; White, L.J.; Mattocks, M.M.; Raut, R.; Perween, A.; Tyagi, P.; et al. A tetravalent virus-like particle vaccine designed to display domain III of dengue envelope proteins induces multi-serotype neutralizing antibodies in mice and macaques which confer protection against antibody dependent enhancement in AG129 mice. PLoS Negl. Trop. Dis. 2018, 12, 0006191. [Google Scholar] [CrossRef]
- Berkower, I.; Raymond, M.; Muller, J.; Spadaccini, A.; Aberdeen, A. Assembly, structure, and antigenic properties of virus-like particles rich in HIV-1 envelope gp120. Virology 2004, 321, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Gordon, D.M.; McGovern, T.W.; Krzych, U.; Cohen, J.C.; Schneider, I.; LaChance, R.; Heppner, D.G.; Yuan, G.; Hollingdale, M.; Slaoui, M.; et al. Safety, immunogenicity, and efficacy of a recombinantly produced Plasmodium falciparum circumsporozoite protein-hepatitis B surface antigen subunit vaccine. J. Infect. Dis. 1995, 171, 1576–1585. [Google Scholar] [CrossRef] [PubMed]
- Rutgers, T.; Gordon, D.; Gathoye, A.M.; Hollingdale, M.; Hockmeyer, W.; Rosenberg, M.; De Wilde, M. Hepatitis B surface antigen as carrier matrix for the repetitive epitope of the circumsporozoite protein of Plasmodium falciparum. Bio/Technology 1988, 6, 1065–1070. [Google Scholar] [CrossRef]
- Beaumont, E.; Patient, R.; Hourioux, C.; Dimier-Poisson, I.; Roingeard, P. Chimeric hepatitis B virus/hepatitis C virus envelope proteins elicit broadly neutralizing antibodies and constitute a potential bivalent prophylactic vaccine. Hepatology 2013, 57, 1303–1313. [Google Scholar] [CrossRef] [PubMed]
- Cervantes Gonzalez, M.; Kostrzak, A.; Guetard, D.; Pniewski, T.; Sala, M. HIV-1 derived peptides fused to HBsAg affects its immunogenicity. Virus Res. 2009, 146, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Schlienger, K.; Mancini, M.; Rivière, Y.; Dormont, D.; Tiollais, P.; Michel, M.L. Human immunodeficiency virus type 1 major nautralizing determinant exposed on hepatitis B surface antigen particles is highly immunogenic in primates. J. Virol. 1992, 66, 2570–2576. [Google Scholar] [CrossRef]
- Kingston, N.J.; Kurtovic, L.; Walsh, R.; Joe, C.; Lovrecz, G.; Locarnini, S.; Beeson, J.G.; Netter, H.J. Hepatitis B virus-like particles expressing Plasmodium falciparum epitopes induce complement-fixing antibodies against the circumsporozoite protein. Vaccine 2019, 37, 1674–1684. [Google Scholar] [CrossRef]
- Wei, S.; Lei, Y.; Yang, J.; Wang, X.; Shu, F.; Wei, X.; Lin, F.; Li, B.; Cui, Y.; Zhang, H.; et al. Neutralization effects of antibody elicited by chimeric HBV S antigen viral-like particles presenting HCV neutralization epitopes. Vaccine 2018, 36, 2273–2281. [Google Scholar] [CrossRef]
- Kotiw, M.; Johnson, M.; Pandey, M.; Fry, S.; Hazell, S.L.; Netter, H.J.; Good, M.F.; Olive, C. Immunological response to parenteral vaccination with recombinant hepatitis B virus surface antigen virus-like particles expressing Helicobacter pylori KatA epitopes in a murine H. pylori challenge model. Clin. Vaccine Immunol. 2012, 19, 268–276. [Google Scholar] [CrossRef]
- Vietheer, P.T.K.; Boo, I.; Drummer, H.E.; Netter, H.J. Immunizations with chimeric hepatitis B virus-like particles to induce potential anti-hepatitis C virus neutralizing antibodies. Antivir. Ther. 2007, 12, 477–487. [Google Scholar]
- Pumpens, P.; Razanskas, R.; Pushko, P.; Renhof, R.; Gusars, I.; Skrastina, D.; Ose, V.; Borisova, G.; Sominskaya, I.; Petrovskis, I.; et al. Evaluation of HBs, HBc, and frCP virus-like particles for expression of human papillomavirus 16 E7 oncoprotein epitopes. Intervirology 2002, 45, 24–32. [Google Scholar] [CrossRef]
- von Brunn, A.; Früh, K.; Müller, H.M.; Zentgraf, H.W.; Bujard, H. Epitopes of the human malaria parasite P. falciparum carried on the surface of HBsAg particles elicit an immune response against the parasite. Vaccine 1991, 9, 477–484. [Google Scholar] [CrossRef]
- Delpeyroux, F.; Chenciner, N.; Lim, A.; Malpièce, Y.; Blondel, B.; Crainic, R.; van der Werf, S.; Streeck, R.E. A poliovirus neutralization epitope expressed on hybrid hepatitis B surface antigen particles. Science 1986, 233, 472–475. [Google Scholar] [CrossRef] [PubMed]
- Cheong, W.S.; Reiseger, J.; Turner, S.J.; Boyd, R.; Netter, H.J. Chimeric virus-like particles for the delivery of an inserted influenza A-specific CTL epitope. Antivir. Res. 2009, 81, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Delpeyroux, F.; Peillon, N.; Blondel, B.; Crainic, R.; Streeck, R.E. Presentation and immunogenicity of the hepatitis B surface antigen and a poliovirus neutralization antigen on mixed empty envelope particles. J. Virol. 1988, 62, 1836–1839. [Google Scholar] [CrossRef]
- Netter, H.J.; Macnaughton, T.B.; Woo, W.P.; Tindle, R.; Gowans, E.J. Antigenicity and immunogenicity of novel chimeric hepatitis B surface antigen particles with exposed hepatitis C virus epitopes. J. Virol. 2001, 75, 2130–2141. [Google Scholar] [CrossRef]
- Casares, S.; Brumeanu, T.D.; Richie, T.L. The RTS, S malaria vaccine. Vaccine 2010, 28, 4880–4894. [Google Scholar] [CrossRef]
- RTS,S Clinical Trials Partnership. Efficacy and safety of RTS,S/AS01 malaria vaccine with or without a booster dose in infants and children in Africa: Final results of a phase 3, individually randomised, controlled trial. Lancet 2015, 386, 31–45. [Google Scholar] [CrossRef]
- Kester, K.E.; Cummings, J.F.; Ofori-Anyinam, O.; Ockenhouse, C.F.; Krzych, U.; Moris, P.; Schwenk, R.; Nielsen, R.A.; Debebe, Z.; Pinelis, E.; et al. Randomized, double-blind, phase 2a trial of falciparum malaria vaccines RTS,S/AS01B and RTS,S/AS02A in malaria-naive adults: Safety, efficacy, and immunologic associates of protection. J. Infect. Dis. 2009, 200, 337–346. [Google Scholar] [CrossRef]
- Collins, K.A.; Snaith, R.; Cottingham, M.G.; Gilbert, S.C.; Hill, A.V.S. Enhancing protective immunity to malaria with a highly immunogenic virus-like particle vaccine. Sci. Rep. 2017, 7, 46621. [Google Scholar] [CrossRef]
- Patient, R.; Hourioux, C.; Vaudin, P.; Pagès, J.C.; Roingeard, P. Chimeric hepatitis B and C viruses envelope proteins can form subviral particles: Implications for the design of new vaccine strategies. New Biotechnol. 2009, 25, 226–234. [Google Scholar] [CrossRef] [PubMed]
- McCluskie, M.J.; Evans, D.M.; Zhang, N.; Benoit, M.; McElhiney, S.P.; Unnithan, M.; DeMarco, S.C.; Clay, B.; Huber, C.; Deora, A.; et al. The effect of pre-existing anti-carrier immunity on subsequent responses to CRM197 or Qb-VLP conjugate vaccines. Immunopharmacol. Immunotoxicol. 2016, 38, 184–196. [Google Scholar] [CrossRef] [PubMed]
- Saxena, M.; Van, T.T.H.; Baird, F.J.; Coloe, P.J.; Smooker, P.M. Pre-existing immunity against vaccine vectors—Friend or foe? Microbiology 2013, 159, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Jegerlehner, A.; Wiesel, M.; Dietmeier, K.; Zabel, F.; Gatto, D.; Saudan, P.; Bachmann, M.F. Carrier induced epitopic suppression of antibody responses induced by virus-like particles is a dynamic phenomenon caused by carrier-specific antibodies. Vaccine 2010, 28, 5503–5512. [Google Scholar] [CrossRef]
- Schutze, M.P.; Leclerc, C.; Jolivet, M.; Audibert, F.; Chedid, L. Carrier-induced epitopic suppression, a major issue for future synthetic vaccines. J. Immunol. 1985, 135, 2319–2322. [Google Scholar]
- Beaumont, E.; Roingeard, P. Chimeric hepatitis B virus (HBV)/hepatitis C virus (HCV) subviral envelope particles induce efficient anti-HCV antibody production in animals pre-immunized with HBV vaccine. Vaccine 2015, 33, 973–976. [Google Scholar] [CrossRef]
- Netter, H.J.; Woo, W.P.; Tindle, R.; Macfarlan, R.I.; Gowans, E.J. Immunogenicity of recombinant HBsAg/HCV particles in mice pre-immunised with hepatitis B virus-specific vaccine. Vaccine 2003, 21, 2692–2697. [Google Scholar] [CrossRef]
- Bojang, K.A.; Milligan, P.J.M.; Pinder, M.; Vigneron, L.; Alloueche, A.; Kester, K.E.; Ballou, W.R.; Conway, D.J.; Reece, W.H.H.; Gothard, P.; et al. Efficacy of RTS,S/AS02 malaria vaccine against Plasmodium falciparum infection in semi-immune adult men in The Gambia: A randomised trial. Lancet 2001, 358, 1927–1934. [Google Scholar] [CrossRef]
- Li, P.; Haque, M.A.; Blum, J.S. Role of disulfide bonds in regulating antigen processing and epitope selection. J. Immunol. 2002, 169, 2444–2450. [Google Scholar] [CrossRef]
- Carmicle, S.; Steede, N.K.; Landry, S.J. Antigen three-dimensional structure guides the processing and presentation of helper T-cell epitopes. Mol. Immunol. 2007, 44, 1159–1168. [Google Scholar] [CrossRef] [PubMed]
- Prato, S.; Fleming, J.; Schmidt, C.W.; Corradin, G.; Lopez, J.A. Cross-presentation of a human malaria CTL epitope is conformation dependent. Mol. Immunol. 2006, 43, 2031–2036. [Google Scholar] [CrossRef] [PubMed]
- Dai, G.; Carmicle, S.; Steede, N.K.; Landry, S.J. Structural basis for helper T-cell and antibody epitope immunodominance in bacteriophage T4 Hsp10. J. Biol. Chem. 2002, 277, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Mirano-Bascos, D.; Steede, N.K.; Robinson, J.E.; Landry, S.J. Influence of disulfide-stabilized structure on the specificity of helper T-cell and antibody responses to HIV envelope glycoprotein gp120. J. Virol. 2010, 84, 3303–3311. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Landry, S.J. Three-dimensional structure determines the pattern of CD4+ T-cell epitope dominance in influenza virus hemagglutinin. J. Virol. 2008, 82, 1238–1248. [Google Scholar] [CrossRef]
- Wolfert, M.A.; Boons, G.J. Adaptive immune activation: Glycosylation does matter. Nat. Chem. Biol. 2013, 9, 776–784. [Google Scholar] [CrossRef]
- Dam, T.K.; Brewer, C.F. Lectins as pattern recognition molecules: The effects of epitope density in innate immunity. Glycobiology 2010, 20, 270–279. [Google Scholar] [CrossRef]
- Al-Barwani, F.; Young, S.L.; Baird, M.A.; Larsen, D.S.; Ward, V.K. Mannosylation of virus-like particles enhances internalization by antigen-presenting cells. PLoS ONE 2014, 9, e104523. [Google Scholar] [CrossRef]
- Freire, T.; Zhang, X.; Dériaud, E.; Ganneau, C.; Vichier-Guerre, S.; Azria, E.; Launay, O.; Lo-Man, R.; Bay, S.; Leclerc, C. Glycosidic Tn-based vaccines targeting dermal dendritic cells favor germinal center B-cell development and potent antibody response in the absence of adjuvant. Blood 2010, 116, 3526–3536. [Google Scholar] [CrossRef]
- Sheng, K.C.; Kalkanidis, M.; Pouniotis, D.S.; Esparon, S.; Tang, C.K.; Apostolopoulos, V.; Pietersz, G.A. Delivery of antigen using a novel mannosylated dendrimer potentiates immunogenicity in vitro and in vivo. Eur. J. Immunol. 2008, 38, 424–436. [Google Scholar] [CrossRef]
- Doe, B.; Steimer, K.S.; Walker, C.M. Induction of HIV-1 envelope (gp120)-specific cytotoxic T lymphocyte responses in mice by recombinant CHO cell-derived gp120 is enhanced by enzymatic removal of N-linked glycans. Eur. J. Immunol. 1994, 24, 2369–2376. [Google Scholar] [CrossRef]
- Mishra, H.; Mishra, D.; Mishra, P.K.; Nahar, M.; Dubey, V.; Jain, N.K. Evaluation of solid lipid nanoparticles as carriers for delivery of hepatitis B surface antigen for vaccination using subcutaneous route. J. Pharm. Pharm. Sci. 2010, 13, 495–509. [Google Scholar] [CrossRef] [PubMed]
- Vigerust, D.J.; Shepherd, V.L. Virus glycosylation: Role in virulence and immune interactions. Trends Microbiol. 2007, 15, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Hyakumura, M.; Walsh, R.; Thaysen-Andersen, M.; Kingston, N.J.; La, M.; Lu, L.; Lovrecz, G.; Packer, N.H.; Locarnini, S.; Netter, H.J. Modification of asparagine-linked glycan density for the design of hepatitis B virus virus-like particles with enhanced immunogenicity. J. Virol. 2015, 89, 11312–11322. [Google Scholar] [CrossRef] [PubMed]
- Lopez, M.; Rodriguez, E.N.; Lobaina, Y.; Musacchio, A.; Falcon, V.; Guillen, G.; Aguilar, J.C. Characterization of the size distribution and aggregation of virus-like nanoparticles used as active ingredients of the HeberNasvac therapeutic vaccine against chronic hepatitis B. Adv. Nat. Sci. Nanosci. Nanotechnol. 2017, 8, 025009. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Source | Cell type | Name | Antigen | Subtype | Manufacturer |
|---|---|---|---|---|---|
| Plasma-derived vaccines | - | Heptavax-B® | HBsAgS | ad | Merck [116] |
| - | Hevac B® | HBsAgS, -M | ad and ay | Pasteur [120,124,125] | |
| - | GCC VAC | HBsAgS | Green Cross Operation, Osaka [125] | ||
| - | Hepavax-B | HBsAgS | Korean Green Cross [126,127] | ||
| - | Hepaccine B | HBsAgS | Cheil Foods & Chemicals Company [128] | ||
| Recombinant (yeast-derived) | Saccharomyces cerevisiae | Recombivax® HB HB-Vax II® | HBsAgS | adw | Merck [129] |
| Saccharomyces cerevisiae | Engerix-B® | HBsAgS | adw | GlaxoSmithKline [129,130] | |
| Saccharomyces cerevisiae | TGP 943™ | HBsAgS, -M | adr | Takeda Chemical Industries [131] | |
| Saccharomyces cerevisiae | Euvax B® | HBsAgS | LG Chemical Ltd, [130,132] | ||
| Pichia pastoris | Shanvac B | HBsAgS | adw2 | Shantha Biotechnics [133] | |
| Pichia pastoris | Heberbiovac-HB® | HBsAgS | adw2 | Heber Biotech S.A., [130] | |
| Hansenula polymorpha | Heplisav-B® | HBsAgS | adw | Dynavax Technologies [134] | |
| Hansenula polymorpha | Hepavax-GeneTM | HBsAgS | adr | Janssen Pharma [130,135] | |
| Recombinant (mammalian cell-derived) | Chinese hamster ovary cells | Gen Hevac B® | HBsAgS, -M | ayw | Pasteur [120] |
| Chinese hamster ovary cells | Sci-B-Vac/Bio-Hep-B™/Hepimmune™ | HBsAgS, -M, -L | adw | VBI Vaccines [136] | |
| Mouse c127 clonal cell line | AG-3™ (Hepacare/Hepagene™) | HBsAgS, -M, -L | adw & ayw | Medeva [137] |
| Antigen | Target | Delivery Site | Expression System | Name | Study/Manufacturer |
|---|---|---|---|---|---|
| CS-HBsAg/HBsAgS | Malaria | N-terminal | Saccharomyces cerevisiae | RTS,S/AS01 Mosquirix™ | GlaxoSmithKline [5,6,190] |
| CS-HBsAgS | Malaria | N-terminal | Pichia pastoris | R21 | [208] |
| DENV-EDIII-HBsAgS/HBsAgS | Dengue virus | N-terminal | Pichia pastoris | DSV4 | [188] |
| HBsAgS-gp120 (HIV-1) | HIV-1 | C-terminal | CV-1 cell line | MR15, MR23 | [189] |
| Env1-, Env2-HBsAgS | Hepatitis C virus | Substitution/N-terminal extension | CHO cells | [192,209] | |
| HBsAgS-NANP repeats | Malaria | Insertion | HEK293F cell line | M-HBsAg-N4, -N9 | [195] |
| HBsAgS-HCV env epitopes | Hepatitis C virus | Insertion | HEK293T cell line | [196,198] | |
| HBsAgS-catalase epitope | Helicobacter pylori | Insertion | HuH-7 | VLP-KatA | [197] |
| HBsAgS-VP1 capsid epitope | Poliovirus | Insertion | Mouse L cells | HBsPolioAg | [201,203] |
| HBsAgS-matrix CTL epitope | Influenza A virus | Insertion/Substitution | HEK293T cell line | [202] | |
| Polyepitope-HBsAgS; | HIV | Substitution preS2 sequence | SW480 cells | [193] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ho, J.K.-T.; Jeevan-Raj, B.; Netter, H.-J. Hepatitis B Virus (HBV) Subviral Particles as Protective Vaccines and Vaccine Platforms. Viruses 2020, 12, 126. https://doi.org/10.3390/v12020126
Ho JK-T, Jeevan-Raj B, Netter H-J. Hepatitis B Virus (HBV) Subviral Particles as Protective Vaccines and Vaccine Platforms. Viruses. 2020; 12(2):126. https://doi.org/10.3390/v12020126
Chicago/Turabian StyleHo, Joan Kha-Tu, Beena Jeevan-Raj, and Hans-Jürgen Netter. 2020. "Hepatitis B Virus (HBV) Subviral Particles as Protective Vaccines and Vaccine Platforms" Viruses 12, no. 2: 126. https://doi.org/10.3390/v12020126
APA StyleHo, J. K.-T., Jeevan-Raj, B., & Netter, H.-J. (2020). Hepatitis B Virus (HBV) Subviral Particles as Protective Vaccines and Vaccine Platforms. Viruses, 12(2), 126. https://doi.org/10.3390/v12020126