Application of a Phage Cocktail for Control of Salmonella in Foods and Reducing Biofilms

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Enrichment, Isolation, Purification, and Preparation of Phages
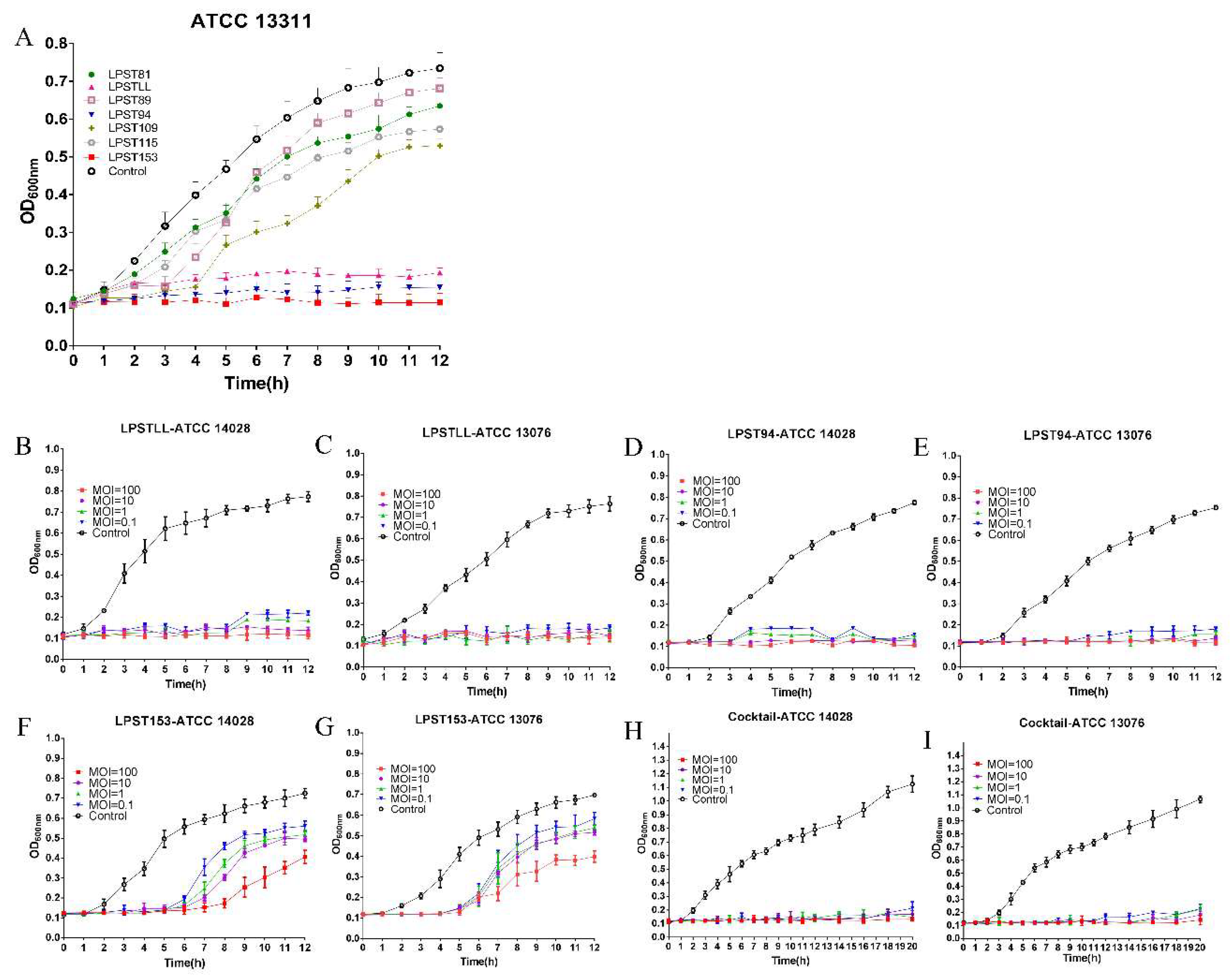
2.3. Screening of Phages Based on Spot Test and Lytic Capacity
2.3.1. Spot Test
2.3.2. Lytic Activity
2.4. Determination of Host Range by Efficiency of Plating (EOP)
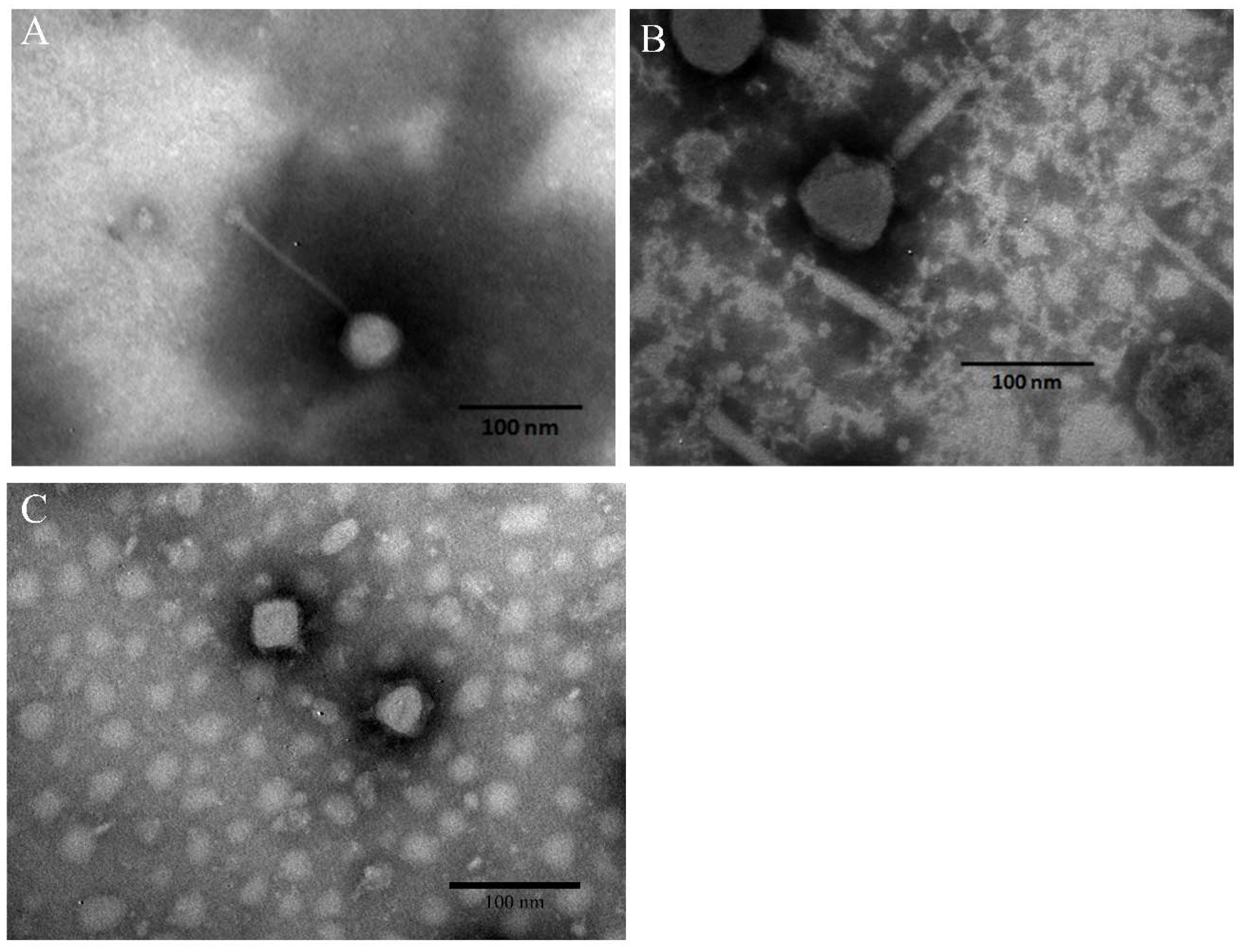
2.5. Transmission Electron Microscopy (TEM)
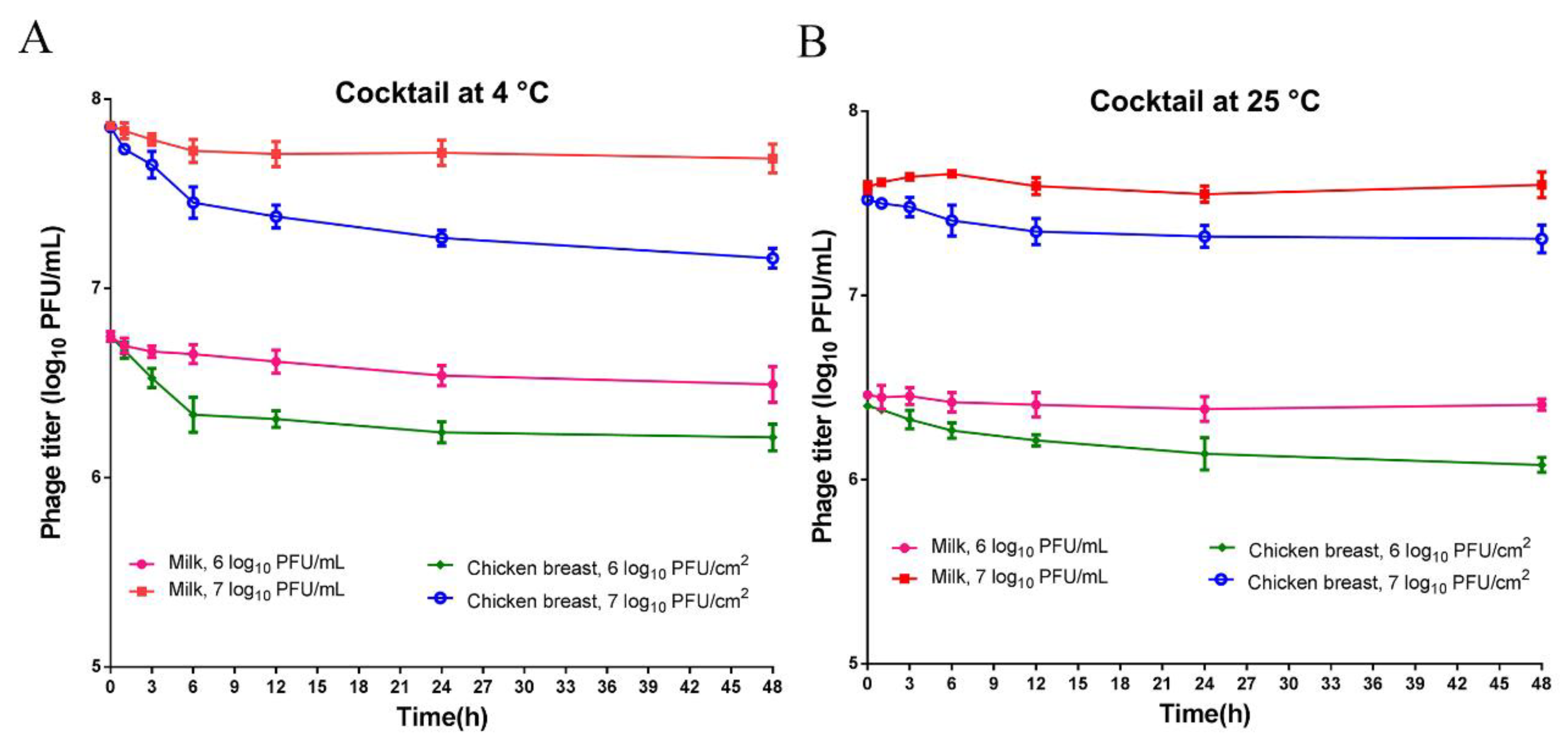
2.6. Phage Stability in Foods
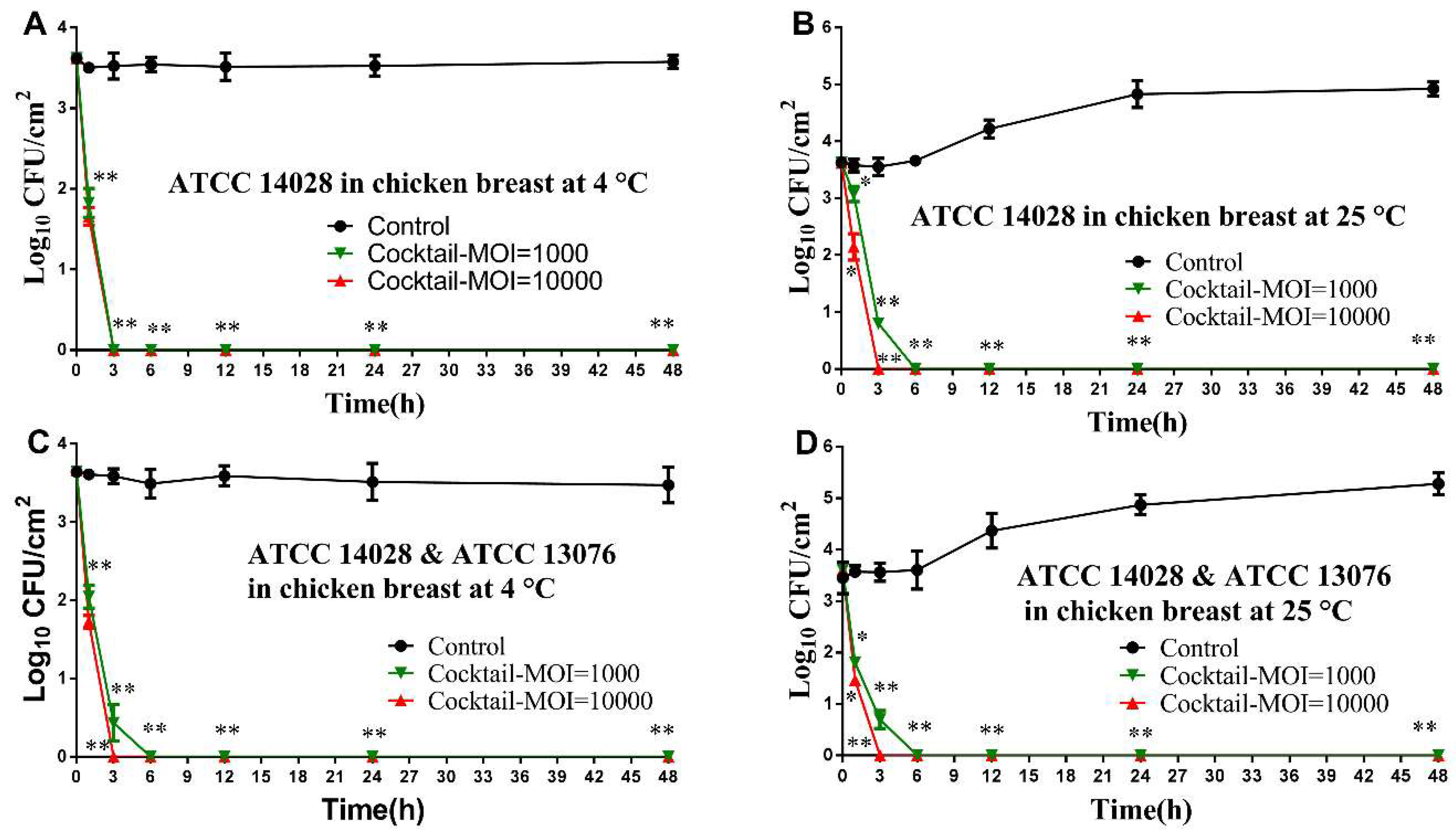
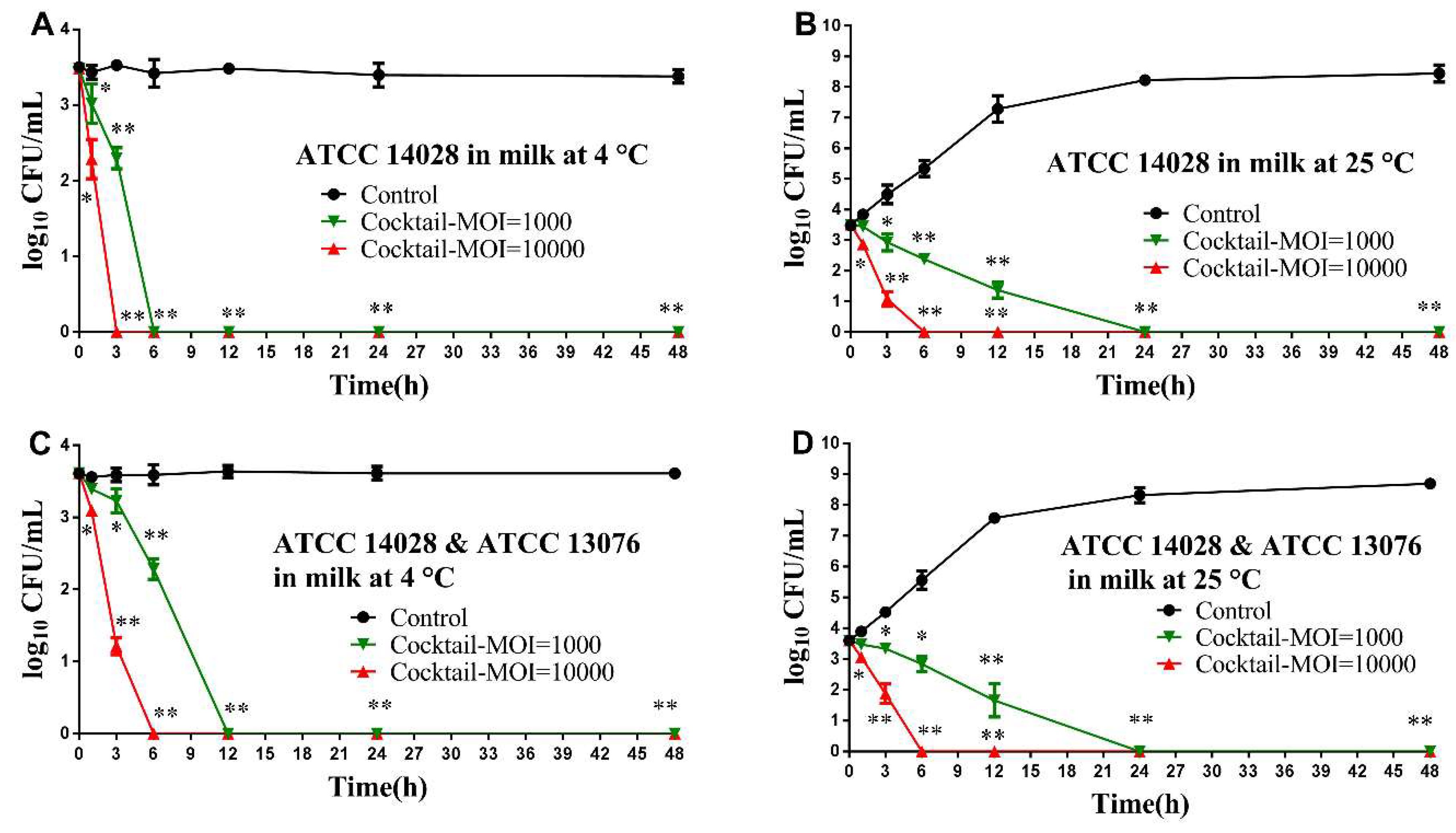
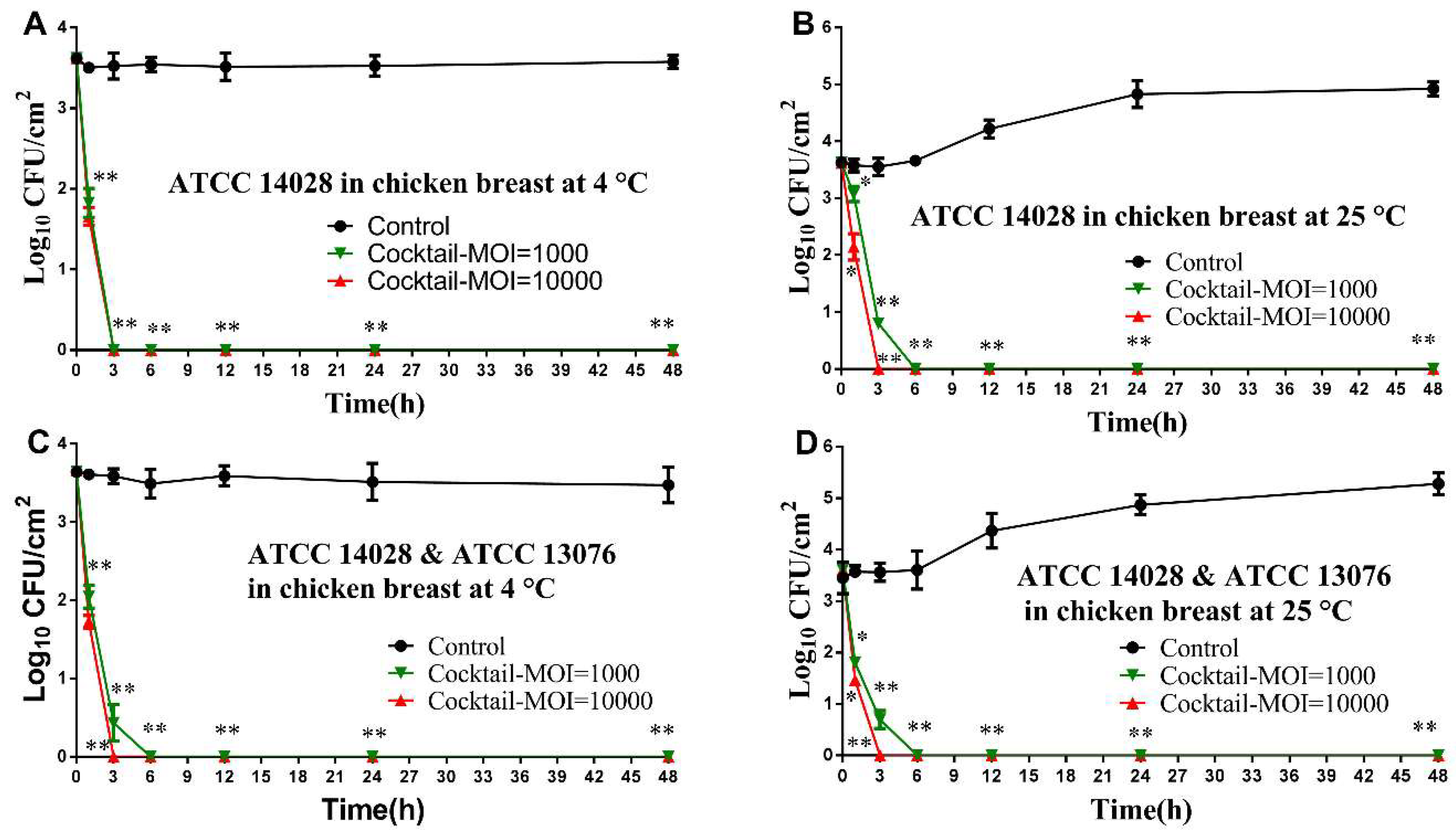
2.7. Biological Control of Salmonella in Foods Using Phage Cocktail
2.7.1. Food Samples
2.7.2. Adding Salmonella and Phage Cocktail for Treatment
2.7.3. Recovered Bacterial Load from Treated Foods
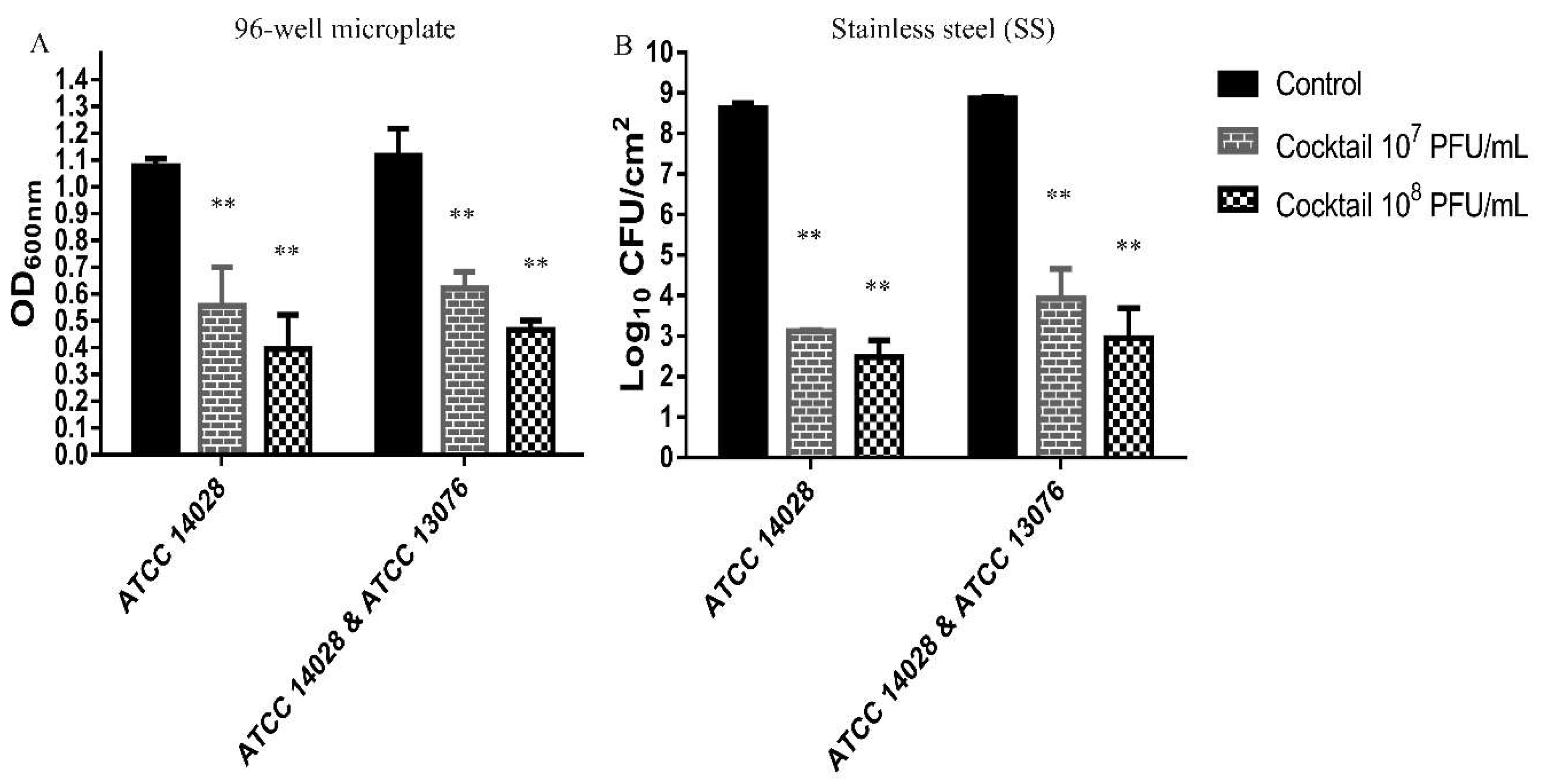
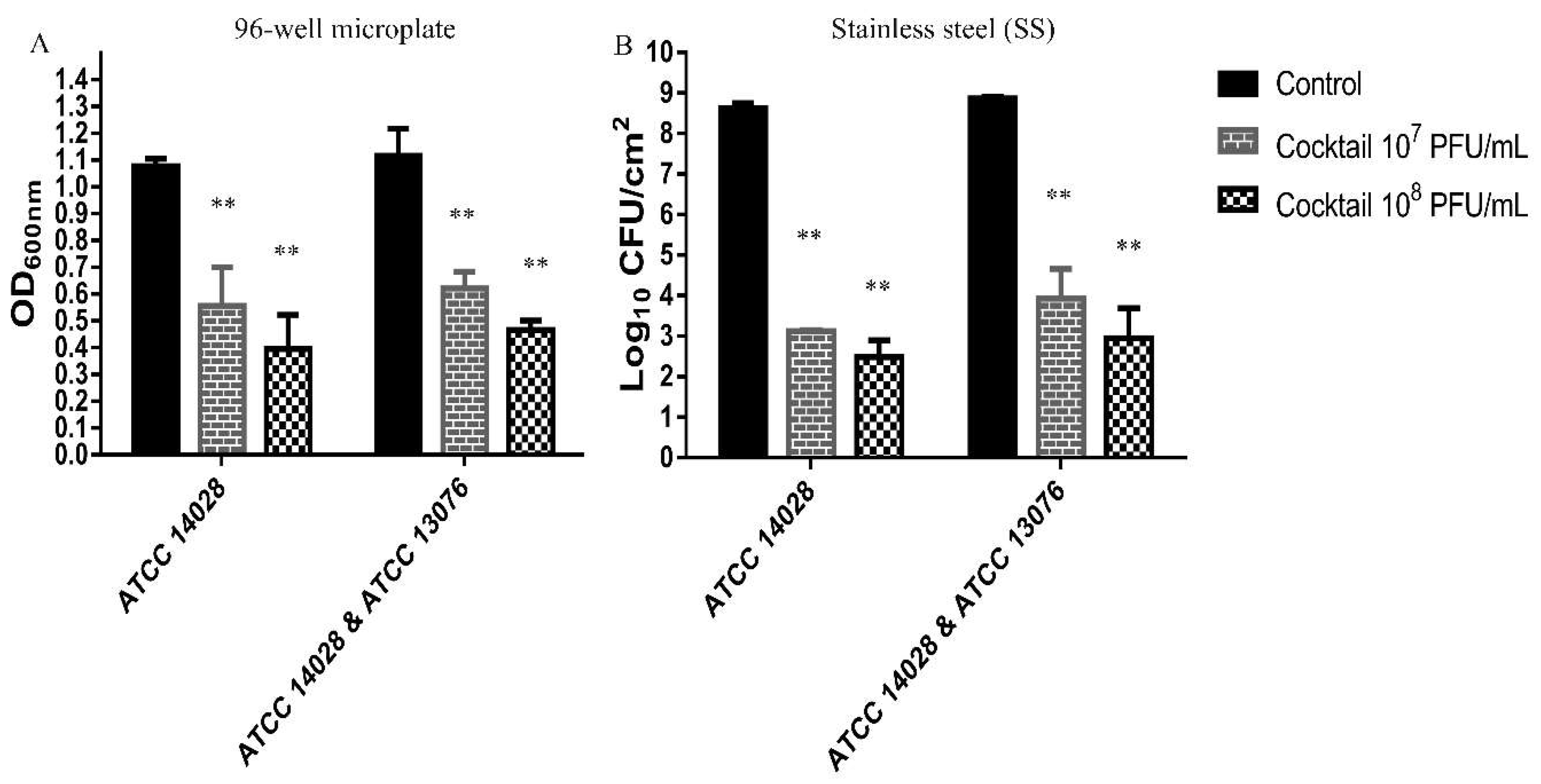
2.8. Effect of Phage Cocktail Against Biofilm of Salmonella in 96-Well Microplate and on Stainless Steel Surface
2.8.1. Biofilm Assay in 96-Well Microplate
2.8.2. Biofilm Assay on Stainless Steel Surface
2.9. Statistical Analysis
3. Results
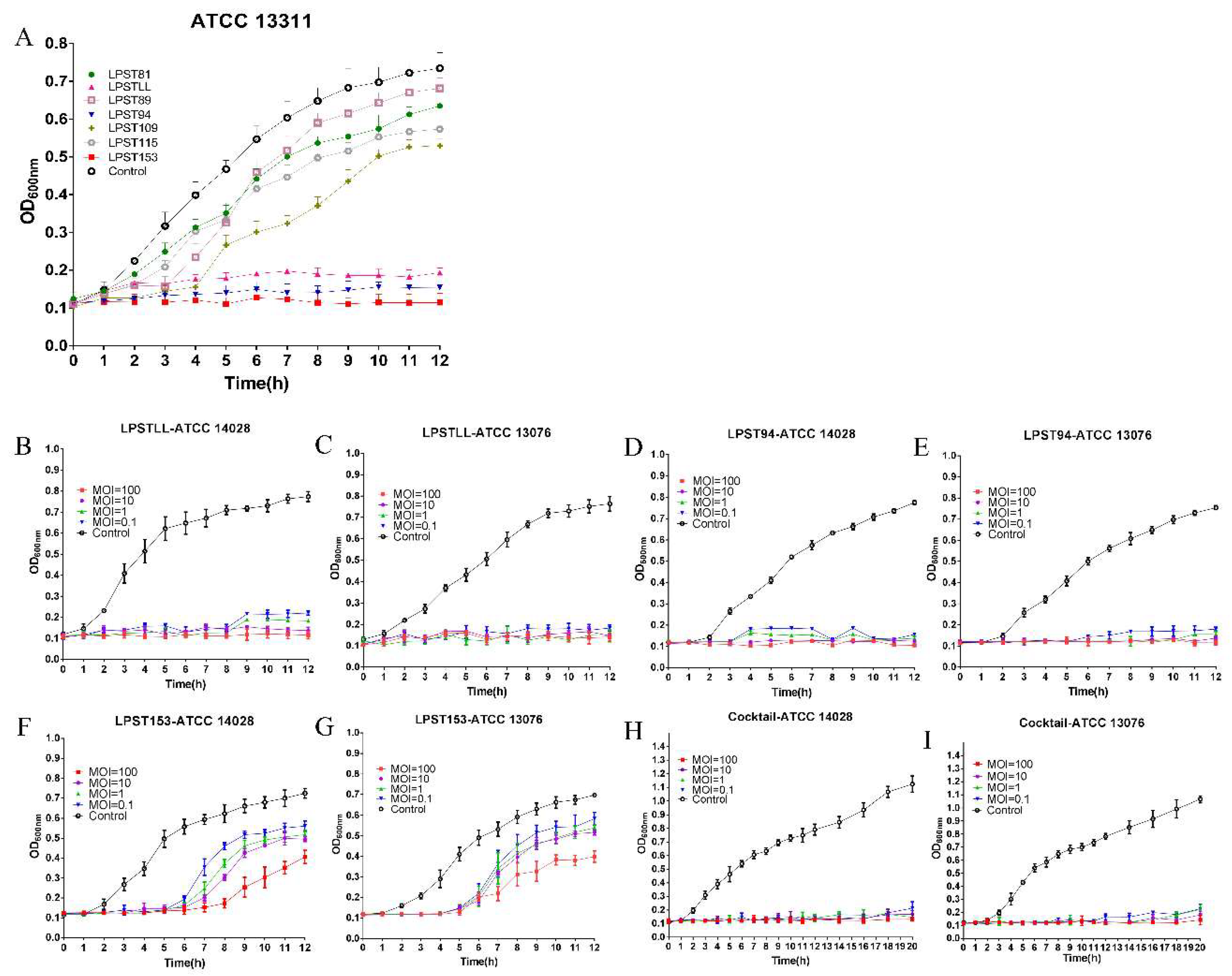
3.1. Isolation and Screening of Phages
3.2. Host Range of Phages by Efficiency of Plating (EOP)
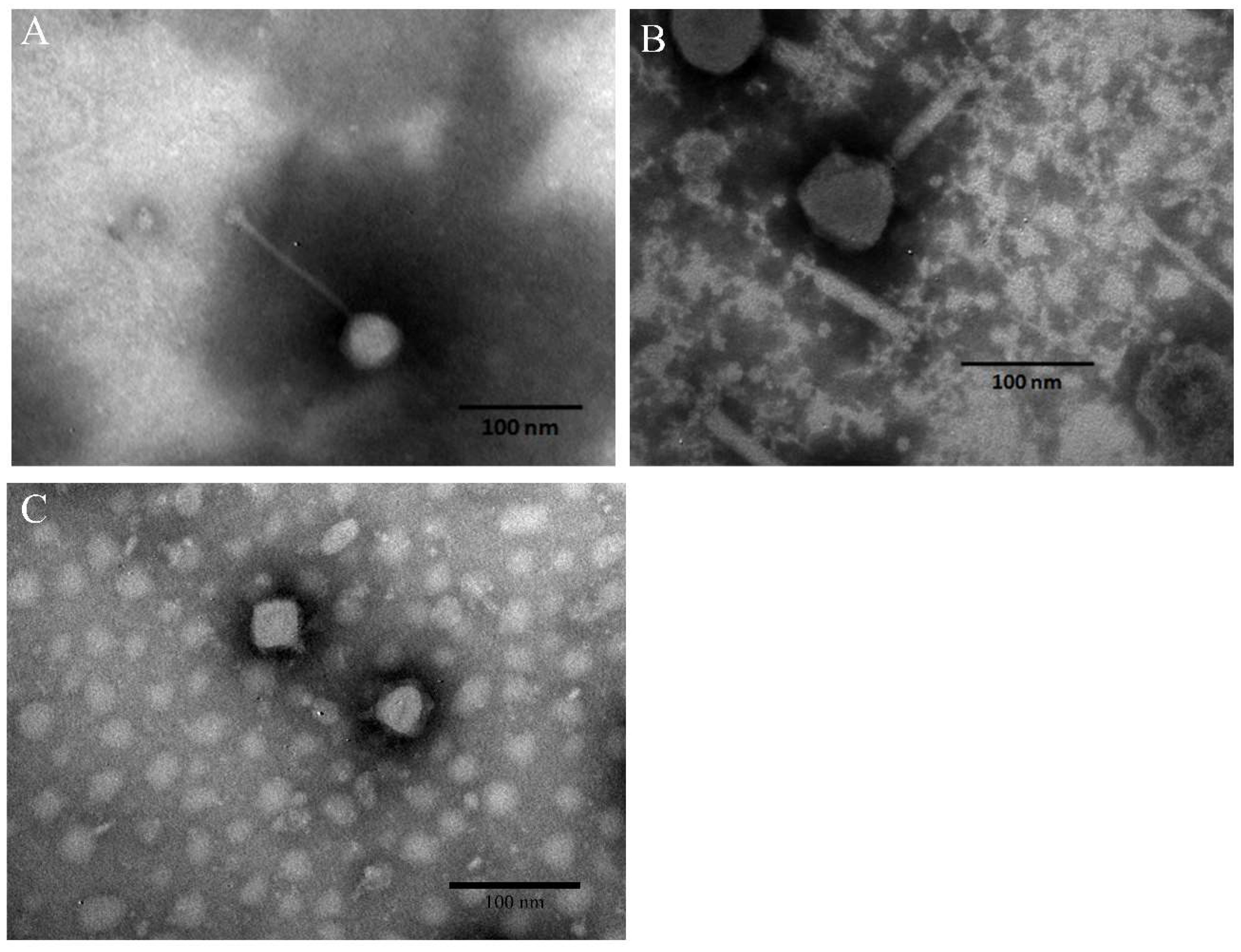
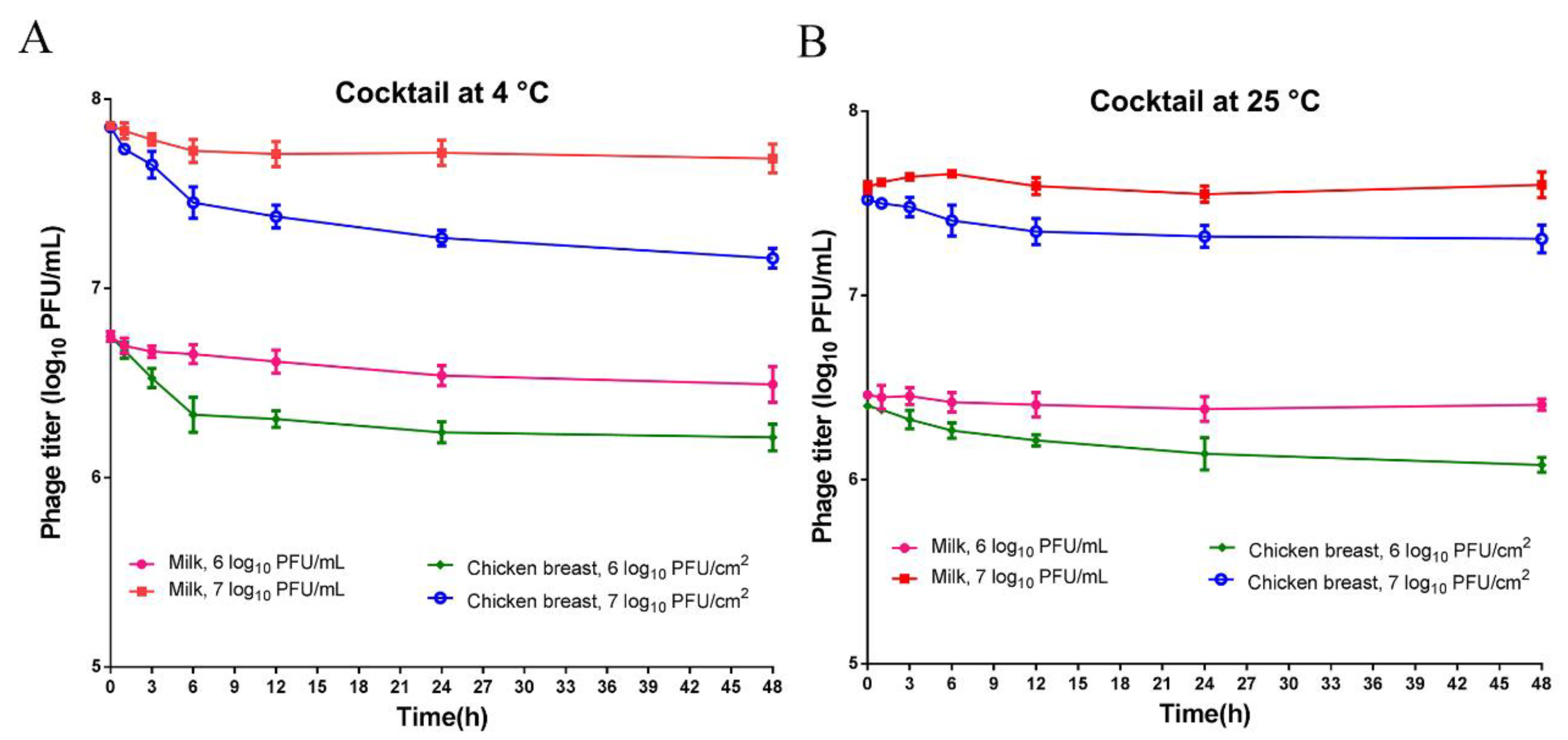
3.3. Morphology of Phages and Stability of Phage Cocktail in Foods
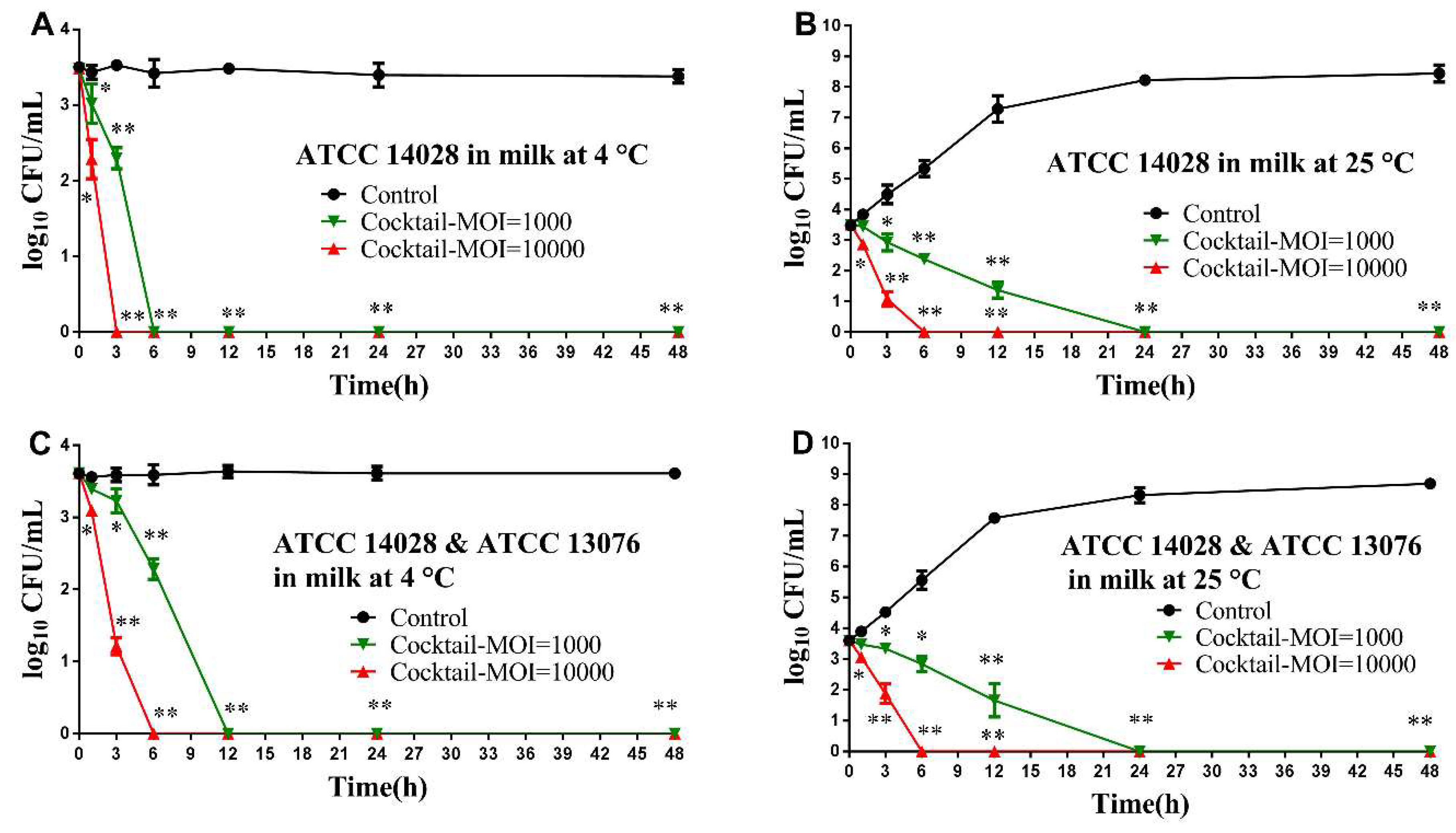
3.4. Application of Phage Cocktail in Controlling Food-borne S. Typhimurium and S. Enteritidis
3.5. Effect of Phage Cocktail Against biofilm of Salmonella
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- CDC. FoodNet Fast Home Page. Available online: https://wwwn.cdc.gov/foodnetfast/ (accessed on 22 March 2018).
- EFSA. The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2016. EFSA J. 2017, 15, 1–288. [Google Scholar]
- Haraga, A.; Ohlson, M.B.; Miller, S.I. Salmonellae interplay with host cells. Nat. Rev. Microbiol. 2008, 6, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Pui, C.F.; Wong, W.C.; Chai, L.C.; Tunung, R.; Jeyaletchumi, P.; Hidayah, M.S.N.; Ubong, A.; Farinazleen, M.G.; Cheah, Y.K. Salmonella: A foodborne pathogen. Int. Food Res. J. 2011, 18, 465–473. [Google Scholar]
- CDC. Salmonella Outbreak Investigations from 2018. Available online: https://www.cdc.gov/salmonella/outbreaks-2018.html (accessed on 7 February 2019).
- Paudyal, N.; Anihouvi, V.; Hounhouigan, J.; Matsheka, M.I.; Sekwati-Monang, B.; Amoa-Awua, W.; Atter, A.; Ackah, N.B.; Mbugua, S.; Asagbra, A. Prevalence of foodborne pathogens in food from selected African countries-A meta-analysis. Int. J. Food Microbiol. 2017, 249, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Majowicz, S.E.; Musto, J.; Scallan, E.; Angulo, F.J.; Kirk, M.; O’Brien, S.J.; Jones, T.F.; Fazil, A.; Hoekstra, R.M. The global burden of nontyphoidal Salmonella gastroenteritis. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2010, 50, 882. [Google Scholar] [CrossRef] [PubMed]
- Hanning, I.B.; Nutt, J.D.; Ricke, S.C. Salmonellosis outbreaks in the United States due to fresh produce: Sources and potential intervention measures. Foodborne Pathog. Dis. 2009, 6, 635–648. [Google Scholar] [CrossRef] [PubMed]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the United States-major pathogens. Emerging Infect. Dis. 2011, 17, 16–22. [Google Scholar] [CrossRef]
- Hooton, S.P.; Atterbury, R.J.; Connerton, I.F. Application of a bacteriophage cocktail to reduce Salmonella Typhimurium U288 contamination on pig skin. Int. J. Food. Microbiol. 2011, 151, 157–163. [Google Scholar] [CrossRef]
- Multistate Outbreak of Salmonella Typhimurium Linked to Chicken Salad. Centers for Disease Control and Prevention. 2018. Available online: https://www.cdc.gov/salmonella/typhimurium-02-8/index.html (accessed on 11 October 2018).
- Ke, B.; Sun, J.; He, D.; Li, X.; Liang, Z.; Ke, C.W. Serovar distribution, antimicrobial resistance profiles, and PFGE typing of Salmonella enterica strains isolated from 2007–2012 in Guangdong, China. BMC Infect. Dis. 2014, 14, 338. [Google Scholar] [CrossRef]
- Huang, X.; Huang, Q.; Shi, W.; Liang, J.H.; Ling-Ling, L.U.; Deng, X.L.; Zhang, Y.H. Epidemiological burden of nontyphoidal Salmonella infection in Guangzhou. Chin. J. Food Hyg. 2014, 26, 217–222. [Google Scholar]
- Luo, Q.; Li, S.; Liu, S.; Tan, H. Foodborne illness outbreaks in China, 2000–2014. Int. J. Clin. Exp. Med. 2017, 10, 5821–5831. [Google Scholar]
- Stapels, D.A.C.; Hill, P.W.S.; Westermann, A.J.; Fisher, R.A.; Thurston, T.L.; Saliba, A.-E.; Blommestein, I.; Vogel, J.; Helaine, S. Salmonella persisters undermine host immune defenses during antibiotic treatment. Science 2018, 362, 1156–1160. [Google Scholar] [CrossRef] [PubMed]
- White, A.P.; Gibson, D.L.; Kim, W.; Kay, W.W.; Surette, M.G. Thin aggregative fimbriae and cellulose enhance long-term survival and persistence of Salmonella. J. Bacteriol. 2006, 188, 3219–3227. [Google Scholar] [CrossRef] [PubMed]
- Steenackers, H.; Hermans, K.; Vanderleyden, J.; Keersmaecker, S.C.J.D.; Steenackers, H.; Hermans, K.; Vanderleyden, J.; Keersmaecker, S.C.J.D. Salmonella biofilms: An overview on occurrence, structure, regulation and eradication. Food Res. Int. 2012, 45, 502–531. [Google Scholar] [CrossRef]
- Cogan, T.A.; Bloomfield, S.F.; Humphrey, T.J. The effectiveness of hygiene procedures for prevention of cross-contamination from chicken carcases in the domestic kitchen. Lett. Appl. Microbiol. 2010, 29, 354–358. [Google Scholar] [CrossRef]
- Reij, M.W.; Den Aantrekker, E.D.; ILSI Europe Risk Analysis in Microbiology Task Force. Recontamination as a source of pathogens in processed foods. Int. J. Food Microbiol. 2004, 91, 1–11. [Google Scholar] [CrossRef]
- Rodrigues, D.; Teixeira, P.; Oliveira, R.; Azeredo, J. Salmonella enterica Enteritidis biofilm formation and viability on regular and triclosan-impregnated bench cover materials. J. Food Prot. 2011, 74, 32. [Google Scholar] [CrossRef]
- Corcoran, M.; Morris, D.; De Lappe, N.; O’Connor, J.; Lalor, P.; Dockery, P.; Cormican, M. Commonly used disinfectants fail to eradicate Salmonella enterica biofilms from food contact surface materials. Appl. Environ. Microbiol. 2014, 80, 1507. [Google Scholar] [CrossRef]
- Perez-Rodriguez, F.; Valero, A.; Carrasco, E.; García, R.M.A.; Zurera, G. Understanding and modelling bacterial transfer to foods: A review. Trends Food Sci. Technol. 2008, 19, 131–144. [Google Scholar] [CrossRef]
- Chylkova, T.; Cadena, M.; Ferreiro, A.; Pitesky, M. Susceptibility of Salmonella Biofilm and Planktonic Bacteria to Common Disinfectant Agents Used in Poultry Processing. J. Food Prot. 2017, 80, 1072. [Google Scholar] [CrossRef]
- Neetoo, H.; Mahomoodally, F. Use of antimicrobial films and edible coatings incorporating chemical and biological preservatives to control growth of Listeria monocytogenes on cold smoked salmon. BioMed Res. Int. 2014, 2014, 534915. [Google Scholar] [CrossRef] [PubMed]
- Musyoka, J.N.; Abong, G.O.; Mbogo, D.M.; Fuchs, R.; Low, J.; Heck, S.; Muzhingi, T. Effects of Acidification and Preservatives on Microbial Growth during Storage of Orange Fleshed Sweet Potato Puree. Int. J. Food Sci. 2018, 2018, 7435–7443. [Google Scholar] [CrossRef] [PubMed]
- Latou, E.; Mexis, S.F.; Badeka, A.V.; Kontominas, M.G. Shelf life extension of sliced wheat bread using either an ethanol emitter or an ethanol emitter combined with an oxygen absorber as alternatives to chemical preservatives. J. Cereal Sci. 2010, 52, 457–465. [Google Scholar] [CrossRef]
- Pawlowska, A.M.; Zannini, E.; Coffey, A.; Arendt, E.K. “Green preservatives”: Combating fungi in the food and feed industry by applying antifungal lactic acid bacteria. Adv. Food Nutr. Res. 2012, 66, 217. [Google Scholar] [PubMed]
- Khoshnoud, M.J.; Siavashpour, A.; Bakhshizadeh, M.; Rashedinia, M. Effects of sodium benzoate, a commonly used food preservative, on learning, memory, and oxidative stress in brain of mice. J. Biochem. Mol. Toxicol. 2017, 32, 2. [Google Scholar] [CrossRef] [PubMed]
- Hoiby, N.; Bjarnsholt, T.; Givskov, M.; Molin, S.; Ciofu, O. Antibiotic resistance of bacterial biofilms. Int. J. Antimicrob. Agents. 2010, 35, 322–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omer, M.T.; Syed, A.A.; Khalid, J.; Askari, B. Study to evaluate the impact of heat treatment on water soluble vitamins in milk. JPMA J. Pak. Med. Assoc. 2010, 60, 909. [Google Scholar]
- Leskova, E.; Kubíková, J.; Kováčiková, E.; Košická, M.; Porubská, J.; Holčíková, K. Vitamin losses: Retention during heat treatment and continual changes expressed by mathematical models. J. Food Compos. Anal. 2006, 19, 252–276. [Google Scholar] [CrossRef]
- Uribarri, J.; Woodruff, S.; Goodman, S.; Cai, W.; Chen, X.; Pyzik, R.; Yong, A.; Striker, G.E.; Vlassara, H. Advanced glycation end products in foods and a practical guide to their reduction in the diet. J. Am. Diet. Assoc. 2010, 110, 911–916. [Google Scholar] [CrossRef]
- Mark, C.; Christian, F.; Enric, M.; Paul, M.M.; Ian, P. The European ban on growth-promoting antibiotics and emerging consequences for human and animal health. J. Antimicrob. Chemother. 2003, 52, 159–161. [Google Scholar] [Green Version]
- Matsuzaki, S.; Uchiyama, J.; Takemurauchiyama, I.; Daibata, M. Perspective: The age of the phage. Nature 2014, 509, S9. [Google Scholar] [CrossRef] [PubMed]
- Goodridge, L.D.; Bisha, B. Phage-based biocontrol strategies to reduce foodborne pathogens in foods. Bacteriophage 2011, 1, 130–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woolston, J.; Parks, A.R.; Abuladze, T.; Anderson, B.; Li, M.; Carter, C.; Hanna, L.F.; Heyse, S.; Charbonneau, D.; Sulakvelidze, A. Bacteriophages lytic for rapidly reduce contamination on glass and stainless steel surfaces. Bacteriophage 2013, 3, e25697. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Shi, Y.; Ji, W.; Meng, X.; Zhang, J.; Wang, H.; Lu, C.; Sun, J.; Yan, Y. Application of a bacteriophage lysin to disrupt biofilms formed by the animal pathogen Streptococcus suis. Appl. Environ. Microbiol. 2011, 77, 8272. [Google Scholar]
- Keen, E.C.; Adhya, S.L. Phage Therapy: Current Research and Applications. Clin. Infect. Dis. 2015, 61, 141–142. [Google Scholar] [CrossRef] [Green Version]
- Kutter, E.M.; Gvasalia, G.; Alavidze, Z.; Brewster, E. Phage therapy in clinical practice: treatment of human infections. Curr. Pharm. Biotechnol. 2010, 11, 69–86. [Google Scholar] [CrossRef] [PubMed]
- Ruchi, T.; Kuldeep, D.; Amit, K.; Anu, R.; Sanjay, K. Bacteriophage therapy for safeguarding animal and human health: A review. Pak. J. Biol. Sci. 2014, 17, 301. [Google Scholar]
- Hyman, P.; Abedon, S.T.; Hyman, P.; Abedon, S.T. Bacteriophages in health and disease. In Bacteriophages in Health & Disease; CABI: Wallingford, UK, 2012. [Google Scholar]
- Mccallin, S.; Alam, S.S.; Barretto, C.; Sultana, S.; Berger, B.; Huq, S.; Krause, L.; Bibiloni, R.; Schmitt, B.; Reuteler, G. Safety analysis of a Russian phage cocktail: From metagenomic analysis to oral application in healthy human subjects. Virology 2013, 443, 187–196. [Google Scholar] [CrossRef]
- Carlton, R.M.; Noordman, W.H.; Biswas, B.; Meester, E.D.D.; Loessner, M.J. Bacteriophage P100 for control of Listeria monocytogenes in foods: Genome sequence, bioinformatic analyses, oral toxicity study, and application. Regul. Toxicol. Pharmacol. 2005, 43, 301–312. [Google Scholar] [CrossRef]
- Greer, G.G. Bacteriophage control of foodborne bacteriat. J. Food Prot. 2005, 68, 1102. [Google Scholar] [CrossRef]
- O’Flynn, G.; Coffey, A.; Fitzgerald, G.F.; Ross, R.P. The newly isolated lytic bacteriophages st104a and st104b are highly virulent against Salmonella enterica. J. Appl. Microbiol. 2006, 101, 251–259. [Google Scholar] [CrossRef]
- O’Flynn, G.; Ross, R.P.; Fitzgerald, G.F.; Coffey, A. Evaluation of a cocktail of three bacteriophages for biocontrol of Escherichia coli O157:H7. Appl. Environ. Microbiol. 2004, 70, 3417–3424. [Google Scholar] [CrossRef]
- Abuladze, T.; Li, M.; Menetrez, M.Y.; Dean, T.; Senecal, A.; Sulakvelidze, A. Bacteriophages reduce experimental contamination of hard surfaces, tomato, spinach, broccoli, and ground beef by Escherichia coli O157:H7. Appl. Environ. Microbiol. 2008, 74, 6230–6238. [Google Scholar] [CrossRef]
- Sparvero, L.J.; Asafuadjei, D.; Rui, K.; Tang, D.; Amin, N.; Im, J.; Rutledge, R.; Lin, B.; Amoscato, A.A.; Zeh, H.J. RAGE (Receptor for Advanced Glycation Endproducts), RAGE Ligands, and their role in Cancer and Inflammation. J. Transl. Med. 2009, 7, 17. [Google Scholar] [CrossRef]
- Carter, C.D.; Parks, A.; Abuladze, T.; Li, M.; Woolston, J.; Magnone, J.; Senecal, A.; Kropinski, A.M.; Sulakvelidze, A. Bacteriophage cocktail significantly reduces Escherichia coli O157: H7 contamination of lettuce and beef, but does not protect against recontamination. Bacteriophage 2012, 2, 178. [Google Scholar] [CrossRef]
- Spricigo, D.A.; Bardina, C.; Cortés, P.; Llagostera, M. Use of a bacteriophage cocktail to control Salmonella in food and the food industry. Int. J. Food Microbiol. 2013, 165, 169–174. [Google Scholar] [CrossRef]
- Albino, L.A.; Rostagno, M.H.; Húngaro, H.M.; Mendonça, R.C. Isolation, characterization, and application of bacteriophages for Salmonella spp. biocontrol in pigs. Foodborne Pathog. Dis. 2014, 11, 602–609. [Google Scholar] [CrossRef]
- Bao, H.; Zhang, P.; Zhang, H.; Zhou, Y.; Zhang, L.; Wang, R. Bio-Control of Salmonella Enteritidis in Foods Using Bacteriophages. Viruses 2015, 7, 4836–4853. [Google Scholar] [CrossRef]
- Galarce, N.; Escobar, B.; Rojas, V.; Navarro, C.; Turra, G.; Robeson, J.; Borie, C. Application of a virulent bacteriophage cocktail leads to reduction of serovar Enteritidis counts in processed meat products. Biocontrol Sci. Technol. 2016, 26, 1–26. [Google Scholar] [CrossRef]
- Huang, C.; Shi, J.; Ma, W.; Zhi, L.; Jia, W.; Li, J.; Wang, X. Isolation, characterization, and application of a novel specific Salmonella bacteriophage in different food matrices. Food Res. Int. 2018, 111, 631–641. [Google Scholar] [CrossRef]
- Srinivasan, S.; Harrington, G.W.; Xagoraraki, I.; Goel, R. Factors affecting bulk to total bacteria ratio in drinking water distribution systems. Water Res. 2008, 42, 3393–3404. [Google Scholar] [CrossRef]
- Akhtar, M.; Viazis, S.; Diez-Gonzalez, F. Isolation, identification and characterization of lytic, wide host range bacteriophages from waste effluents against Salmonella enterica serovars. Food Control 2014, 38, 67–74. [Google Scholar] [CrossRef]
- Klieve, A.V. Bacteriophages. In Methods in Gut Microbial Ecology for Ruminants; Springer: Dordrecht, The Netherlands, 2005; pp. 39–46. [Google Scholar]
- Mullan, W.M. Bacteriophage and Food Fermentations. Phage Assay and Enumeration. Available online: https://slideplayer.com/slide/13011905/ (accessed on 29 March 2018).
- Van, T.R.; Kropinski, A.M. Bacteriophage enrichment from water and soil. Methods Mol. Biol. 2009, 501, 15. [Google Scholar]
- Jin, W.J.; Ji, H.K.; Sang, P.S.; Han, J.E.; Ji, Y.C.; Park, S.C. Protective effects of the Aeromonas phages pAh1-C and pAh6-C against mass mortality of the cyprinid loach (Misgurnus anguillicaudatus) caused by Aeromonas hydrophila. Aquaculture 2013, 2, 289–295. [Google Scholar] [CrossRef]
- Huang, C.; Virk, S.M.; Shi, J.; Zhou, Y.; Willias, S.P.; Morsy, M.K.; Abdelnabby, H.E.; Liu, J.; Wang, X.; Li, J. Isolation, Characterization, and Application of Bacteriophage LPSE1 Against Salmonella enterica in Ready to Eat (RTE) Foods. Front. Microbiol 2018, 9, 1046. [Google Scholar] [CrossRef]
- Mohammadali Khan, M.; Nilsson, A.S. Correction: Isolation of phages for phage therapy: A comparison of spot tests and efficiency of plating analyses for determination of host range and efficacy. PLoS ONE 2015, 10, e0118557. [Google Scholar]
- Ackermann, H.W. Phage classification and characterization. Methods Mol. Biol. 2009, 501, 127. [Google Scholar]
- Laguerre, O.; Derens, E.; Palagos, B. Study of domestic refrigerator temperature and analysis of factors affecting temperature: A French survey. Int. J. Refrig. 2002, 25, 653–659. [Google Scholar] [CrossRef]
- Tomat, D.; Casabonne, C.; Aquili, V.; Balagué, C.; Quiberoni, A. Evaluation of a novel cocktail of six lytic bacteriophages against Shiga toxinproducing Escherichia coli in broth, milk and meat. Food Microbiol. 2018, 76, 434–442. [Google Scholar] [CrossRef]
- Kostaki, M.; Chorianopoulos, N.; Braxou, E.; Nychas, G.J.; Giaouris, E. Differential biofilm formation and chemical disinfection resistance of sessile cells of Listeria monocytogenes strains under monospecies and dual-species (with Salmonella enterica) conditions. Appl. Environ. Microbiol. 2012, 78, 2586–2595. [Google Scholar] [CrossRef]
- Else, T.A.; Pantle, C.R.; Amy, P.S. Boundaries for Biofilm Formation: Humidity and Temperature. Appl. Environ. Microbiol. 2003, 69, 5006–5010. [Google Scholar] [CrossRef] [Green Version]
- Shen, C.; Luo, Y.; Nou, X.; Bauchan, G.; Zhou, B.; Wang, Q.; Millner, P. Enhanced Inactivation of Salmonella and Pseudomonas Biofilms on Stainless Steel by Use of T-128, a Fresh-Produce Washing Aid, in Chlorinated Wash Solutions. Appl. Environ. Microbiol. 2012, 78, 6789–6798. [Google Scholar] [CrossRef]
- Tarabees, R.; Msa, E.; Shawish, R.; Basiouni, S.; Shehata, A.A. Isolation and characterization of Salmonella Enteritidis and Salmonella Typhimurium from chicken meat in Egypt. J. Infect. Dev. Ctries. 2017, 11, 314–319. [Google Scholar] [CrossRef]
- Rohwer, F.; Segall, A.M. In retrospect: A century of phage lessons. Nature 2015, 528, 46–48. [Google Scholar] [CrossRef]
- Careysmith, G.V.; Billington, C.; Cornelius, A.J.; Hudson, J.A.; Heinemann, J.A. Isolation and characterization of bacteriophages infecting Salmonella spp. FEMS Microbiol. Lett. 2006, 258, 182–186. [Google Scholar] [CrossRef]
- Bielke, L.; Higgins, S.; Donoghue, A.; Donoghue, D.; Hargis, B.M. Salmonella Host Range of Bacteriophages That Infect Multiple Genera. Poult. Sci. 2007, 86, 2536–2540. [Google Scholar] [CrossRef]
- Wongsuntornpoj, S.; Switt, A.I.M.; Bergholz, P.; Wiedmann, M.; Chaturongakul, S. Salmonella phages isolated from dairy farms in Thailand show wider host range than a comparable set of phages isolated from U.S. dairy farms. Vet. Microbiol. 2014, 172, 345–352. [Google Scholar] [CrossRef]
- Duc, H.M.; Son, H.M.; Honjoh, K.I.; Miyamoto, T. Isolation and application of bacteriophages to reduce Salmonella contamination in raw chicken meat. LWT 2018, 91, 353–360. [Google Scholar] [CrossRef]
- Mateus, L.; Costa, L.; Silva, Y.J.; Pereira, C.; Cunha, A.; Almeida, A. Efficiency of phage cocktails in the inactivation of Vibrio in aquaculture. Aquaculture 2014, 424–425, 167–173. [Google Scholar] [CrossRef]
- Pereira, C.; Silva, Y.J.; Santos, A.L.; Cunha, Â.; Gomes, N.C.M.; Almeida, A. Bacteriophages with Potential for Inactivation of Fish Pathogenic Bacteria: Survival, Host Specificity and Effect on Bacterial Community Structure. Mar. Drugs 2011, 9, 2236–2255. [Google Scholar] [CrossRef]
- Tanji, Y.; Shimada, T.; Yoichi, M.; Miyanaga, K.; Hori, K.; Unno, H. Toward rational control of Escherichia coli O157:H7 by a phage cocktail. Appl. Microbiol. Biotechnol. 2004, 64, 270–274. [Google Scholar] [CrossRef]
- Jaewoo, B.; Byeonghwa, J.; Sangryeol, R. Effective inhibition of Salmonella Typhimurium in fresh produce by a phage cocktail targeting multiple host receptors. Food Microbiol. 2019, 77, 52–60. [Google Scholar]
- Pereira, C.; Moreirinha, C.; Lewicka, M.; Almeida, P.; Clemente, C.; Romalde, J.L.; Nunes, M.L.; Almeida, A. Characterization and in vitro evaluation of new bacteriophages for the biocontrol of Escherichia coli. Virus Res. 2017, 227, 171–182. [Google Scholar] [CrossRef]
- Landsberger, M.; Gandon, S.; Meaden, S.; Rollie, C.; Chevallereau, A.; Chabas, H.; Buckling, A.; Westra, E.R.; Houte, S.V. Anti-CRISPR Phages Cooperate to Overcome CRISPR-Cas Immunity. Cell 2018, 174, 908–916. [Google Scholar] [CrossRef]
- Novacek, J.; Šiborová, M.; Benešík, M.; Pantůček, R.; Doškař, J.; Plevka, P. Structure and genome release of Twort-like Myoviridae phage with a double-layered baseplate. Proc. Natl. Acad. Sci. USA 2016, 113, 9351. [Google Scholar] [CrossRef]
- Hooton, S.P.; Timms, A.R.; Rowsell, J.; Wilson, R.; Connerton, I.F. Salmonella Typhimurium-specific bacteriophage ΦSH19 and the origins of species specificity in the Vi01-like phage family. Virol. J. 2011, 8, 1–14. [Google Scholar] [CrossRef]
- Sylwia, P.; Magdalena, K.; Romuald, G.; Lidia, M.; Anna, M. Bacteriophages as an alternative strategy for fighting biofilm development. Pol. J. Microbiol. 2014, 63, 137. [Google Scholar]
- Sunderland, K.; Yang, M.; Mao, C. Phage-Enabled Nanomedicine: From Probes to Therapeutics in Precision Medicine. Angew. Chem. 2016, 56, 1964. [Google Scholar] [CrossRef]
- Yele, A.B.; Thawal, N.D.; Sahu, P.K.; Chopade, B.A. Novel lytic bacteriophage AB7-IBB1 of Acinetobacter baumannii: Isolation, characterization and its effect on biofilm. Arch. Virol. 2012, 157, 1441–1450. [Google Scholar] [CrossRef]
- Islam, M.S.; Raz, A.; Liu, Y.; Elbassiony, K.R.A.; Dong, X.; Zhou, P.; Zhou, Y.; Li, J. Complete Genome Sequence of Aeromonas Phage ZPAH7 with Halo Zones, Isolated in China. Microbiol. Resour. Announc. 2019, 8, e01678-18. [Google Scholar] [CrossRef]
- Guenther, S.; Herzig, O.; Fieseler, L.; Klumpp, J.; Loessner, M.J. Biocontrol of Salmonella Typhimurium in RTE foods with the virulent bacteriophage FO1-E2. Int. J. Food Microbiol. 2012, 154, 66–72. [Google Scholar] [CrossRef]
- El-Shibiny, A.; El-Sahhar, S.; Adel, M. Phage applications for improving food safety and infection control in Egypt. J. Appl. Microbiol. 2017, 123, 556. [Google Scholar] [CrossRef]
- Moye, Z.; Woolston, J.; Sulakvelidze, A. Bacteriophage Applications for Food Production and Processing. Viruses 2018, 10, 205. [Google Scholar] [CrossRef]
- Sharma, C.S.; Dhakal, J.; Nannapaneni, R. Efficacy of Lytic Bacteriophage Preparation in Reducing Salmonella in Vitro, on Turkey Breast Cutlets, and on Ground Turkey. J. Food Prot. 2015, 78, 1357. [Google Scholar] [CrossRef]
- Sharma, M.; Patel, J.R.; Conway, W.S.; Ferguson, S.; Sulakvelidze, A. Effectiveness of bacteriophages in reducing Escherichia coli O157:H7 on fresh-cut cantaloupes and lettucet. J. Food Prot. 2009, 72, 1481. [Google Scholar] [CrossRef]
- Bigwood, T.; Hudson, J.A.; Billington, C. Influence of host and bacteriophage concentrations on the inactivation of food-borne pathogenic bacteria by two phages. FEMS Microbiol. Lett. 2010, 291, 59–64. [Google Scholar] [CrossRef]
- Abedon, S. Phage therapy pharmacology: Calculating phage dosing. Adv. Appl. Microbiol. 2011, 77, 1. [Google Scholar]
- Abedon, S.T. Lysis from without. Bacteriophage 2011, 1, 46–49. [Google Scholar] [CrossRef]
- Guenther, S.; Huwyler, D.; Richard, S.; Loessner, M. Virulent Bacteriophage for Efficient Biocontrol of Listeria monocytogenes in Ready-To-Eat Foods. Appl. Environ. Microbiol. 2009, 75, 93–100. [Google Scholar] [CrossRef]
- Sukumaran, A.T.; Nannapaneni, R.; Kiess, A.; Sharma, C.S. Reduction of Salmonella on chicken meat and chicken skin by combined or sequential application of lytic bacteriophage with chemical antimicrobials. Int. J. Food Microbiol. 2015, 207, 8–15. [Google Scholar] [CrossRef]
- Yeh, Y.; Purushothaman, P.; Gupta, N.; Ragnone, M.; Verma, S.C.; Mello, A.S.D. Bacteriophage application on red meats and poultry: Effects on Salmonella population in final ground products. Meat Sci. 2017, 127, 30–34. [Google Scholar] [CrossRef]
- Corbin, B.D.; Rjc, M.L.; Aron, G.M. Bacteriophage T4 multiplication in a glucose-limited Escherichia coli biofilm. Can. J. Microbiol. 2001, 47, 680–684. [Google Scholar] [CrossRef]
- Sharma, M.; Ryu, J.H.; Beuchat, L.R. Inactivation of Escherichia coli O157:H7 in biofilm on stainless steel by treatment with an alkaline cleaner and a bacteriophage. J. Appl. Microbiol. 2010, 99, 449–459. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strains | Strain ID Number | Numbers of Strains | Source of Strains |
|---|---|---|---|
| S.enterica serovar Typhimurium | ATCC 14028, ATCC 13311 | 2 | ATCC |
| UK-1, ST8, SGSC 4903, SL 1344, LT2 | 5 | LS | |
| S.enterica serovar Enteritidis | ATCC 13076 | 1 | ATCC |
| SJTUF 10978, SJTUF 10984 | 2 | SJTU | |
| LK5-3820, SGSC 4901 | 2 | LS | |
| S. enterica Serovar Pullorum | CVCC 519 | 1 | LS |
| S.enterica serovar Dublin | 3710,3723 | 2 | LS |
| S. enterica subsp. enterica serovar Anatum | ATCC 9270 | 1 | ATCC |
| S. enterica subsp. arizonae | CDC 346-86 | 1 | CDC |
| S. enterica subsp. enterica serovar Javiana | CVM 35943 | 1 | LS |
| S. enterica subsp. enterica serovar Kentucky | CVM 29188 | 1 | LS |
| S. enterica serovar Newport | E20002725 | 1 | CDC |
| S. enterica serovar Paratyphi B | CMCC 50094 | 1 | CMCC |
| S. enterica serovar Choleraesuls | ATCC 10708 | 1 | ATCC |
| Drug resistant S. enterica serovar Typhimurium | LST10, LST11, LST12, LST13, LST14, LST15, LST16, LST17, LST18, LST19 | 10 | LS |
| Drug resistant S. enterica serovar Enteritidis | LSE6, LSE7, LSE8, LSE9, LSE10, LSE11, LSE12, LSE13, LSE15 | 9 | LS |
| E. coli | BL21, DH5α | 2 | TB |
| ATCC 933 | 1 | ATCC | |
| F18AC, C83715, T10 | 3 | LS | |
| A. hydrophila | ZYAH72, ZYAH75, J1, D4 | 4 | LS |
| S. aureus | ATCC 6538, ATCC 8095, ATCC 29213 | 3 | ATCC |
| Listeria | ATCC 1914 | 1 | ATCC |
| Bacterial Strains | % of Positive Spot Test against Salmonella Serovars and Other Bacterial Strains | ||||||
|---|---|---|---|---|---|---|---|
| LPST81 | LPSTLL | LPST89 | LPST94 | LPST109 | LPST115 | LPST153 | |
| Salmonella serovars | |||||||
| Typhimurium (N = 7) | 71.4 | 100 | 100 | 100 | 100 | 71.4 | 85.7 |
| Enteritidis (N = 5) | 60 | 100 | 40 | 100 | 60 | 20 | 80 |
| Dublin (N = 2) | 0 | 100 | 0 | 100 | 50 | 0 | 50 |
| Choleraesuls (N = 1) | 0 | 100 | 0 | 100 | 0 | 0 | 100 |
| Newport (N = 1) | 0 | 100 | 100 | 100 | 0 | 0 | 0 |
| Paratyphi B (N = 1) | 100 | 100 | 0 | 100 | 100 | 0 | 100 |
| Anatum (N = 1) | 0 | 100 | 0 | 100 | 0 | 0 | 0 |
| Pullorum (N = 1) | 0 | 100 | 0 | 100 | 0 | 0 | 100 |
| Javiana (N = 1) | 0 | 100 | 100 | 100 | 0 | 0 | 100 |
| Kentucky (N = 1) | 0 | 100 | 0 | 100 | 0 | 0 | 0 |
| S. arizonae (N = 1) | 0 | 100 | 0 | 100 | 0 | 0 | 100 |
| Drug resistant Salmonella serovars | |||||||
| Typhimurium (N = 10) | 60 | 100 | 90 | 100 | 70 | 10 | 90 |
| Enteritidis (N = 9) | 11.1 | 100 | 33.3 | 100 | 22.2 | 11.1 | 88.9 |
| Other bacterial strains | |||||||
| E. coli (N = 6) | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A. hydrophila (N = 4) | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| S. aureus (N = 3) Listeria (N = 1) | 0 0 | 0 0 | 0 0 | 0 0 | 0 0 | 0 0 | 0 0 |
| Bacterial Strains | LPSTLL | LPST94 | LPST153 | Cocktail | Strains | LPSTLL | LPST94 | LPST153 | Cocktail |
|---|---|---|---|---|---|---|---|---|---|
| S.enterica serovar Typhimurium | Drug resistance S. enterica serovar Typhimurium | ||||||||
| ATCC 14028 | 1 | 1 | 0.18 | 1 | LST10 | 0 | 0.1 | 0.004 | 0.26 |
| ATCC 13311 | 1 | 1 | Host | 1 | LST11 | 0.19 | 0.4 | 0.003 | 0.48 |
| UK-1 | Host | Host | 0.17 | 1 | LST12 | 0.1 | 0.3 | 0.007 | 66 |
| ST8 | 1 | 1 | 0.1 | 1 | LST13 | 0.02 | 0.45 | 0.009 | 0.89 |
| SGSC 4903 | 1 | 1 | 1 | 1 | LST14 | 0.1 | 0.1 | 0.006 | 38 |
| SL 1344 | 1 | 1 | 1 | 1 | LST15 | 0.006 | 0.15 | 0.1 | 0.32 |
| LT2 | 1 | 1 | 1 | 1 | LST16 | 0.007 | 0.1 | 0.002 | 0.22 |
| S.enterica serovar Enteritidis | LST17 | 0 | 0.4 | 0.005 | 0.73 | ||||
| ATCC 13076 | 0.1 | 0.4 | 0.1 | 0.6 | LST18 | 0.009 | 0.1 | 0.010 | 0.24 |
| SJTUF 10978 | 0.02 | 0.24 | 0.19 | 0.4 | LST19 | 0 | 0.2 | 0.016 | 0.31 |
| SJTUF 10984 | 0.1 | 0.19 | 0.17 | 0.3 | Drug resistance S. enterica serovar Enteritidis | ||||
| LK5-3820 | 0.005 | 0.4 | 0.003 | 0.35 | LSE6 | 0 | 0.005 | 0 | 0.23 |
| SGSC 4901 | 0.18 | 0.3 | 0.12 | 0.5 | LSE7 | 0.017 | 0.1 | 0.004 | 0.37 |
| S.enterica serovar Dublin | LSE8 | 0.003 | 0.3 | 0.1 | 0.55 | ||||
| 3710 | 0 | 0.19 | 0 | 0.3 | LSE9 | 0.015 | 0.4 | 0 | 0.62 |
| 3723 | 0 | 0.2 | 0 | 0.25 | LSE10 | 0 | 0.1 | 0 | 0.23 |
| S. enterica serovar Choleraesuls | LSE11 | 0.1 | 0.006 | 0 | 0.29 | ||||
| ATCC 10708 | 0.009 | 0.16 | 0 | 0.3 | LSE12 | 0 | 0.2 | 0 | 0.27 |
| S. enterica serovar Newport | LSE13 | 0 | 0.007 | 0 | 0.24 | ||||
| E20002725 | 0 | 0.15 | 0 | 0.23 | LSE15 | 0.14 | 0.1 | 0.013 | 0.51 |
| S. enterica serovar Paratyphi B | |||||||||
| CMCC 50094 | 0.900 | 0.2 | 0.19 | 0.44 | |||||
| S. enterica Serovar Pullorum | |||||||||
| CVCC 519 | 0.017 | 0.2 | 0.1 | 0.67 | |||||
| S. enterica subsp. Enterica serovar Javiana | |||||||||
| CVM 35943 | 0.012 | 0.1 | 0.003 | 0.24 | |||||
| S. enterica subsp. enterica serovar Anatum | |||||||||
| ATCC 9270 | 0 | 0.007 | 0 | 0.21 | |||||
| S. enterica subsp. enterica serovar Kentucky | |||||||||
| CVM 29188 | 0.4 | 0.2 | 0.1 | 0.76 | |||||
| S. enterica subsp. arizonae | |||||||||
| CDC 346-86 | 0.004 | 0.2 | 0 | 0.27 | |||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Islam, M.S.; Zhou, Y.; Liang, L.; Nime, I.; Liu, K.; Yan, T.; Wang, X.; Li, J. Application of a Phage Cocktail for Control of Salmonella in Foods and Reducing Biofilms. Viruses 2019, 11, 841. https://doi.org/10.3390/v11090841
Islam MS, Zhou Y, Liang L, Nime I, Liu K, Yan T, Wang X, Li J. Application of a Phage Cocktail for Control of Salmonella in Foods and Reducing Biofilms. Viruses. 2019; 11(9):841. https://doi.org/10.3390/v11090841
Chicago/Turabian StyleIslam, Md. Sharifull, Yang Zhou, Lu Liang, Ishatur Nime, Kun Liu, Ting Yan, Xiaohong Wang, and Jinquan Li. 2019. "Application of a Phage Cocktail for Control of Salmonella in Foods and Reducing Biofilms" Viruses 11, no. 9: 841. https://doi.org/10.3390/v11090841
APA StyleIslam, M. S., Zhou, Y., Liang, L., Nime, I., Liu, K., Yan, T., Wang, X., & Li, J. (2019). Application of a Phage Cocktail for Control of Salmonella in Foods and Reducing Biofilms. Viruses, 11(9), 841. https://doi.org/10.3390/v11090841