Evidence for West Nile Virus and Usutu Virus Infections in Wild and Resident Birds in Germany, 2017 and 2018

 , , , , , , ,
, , , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
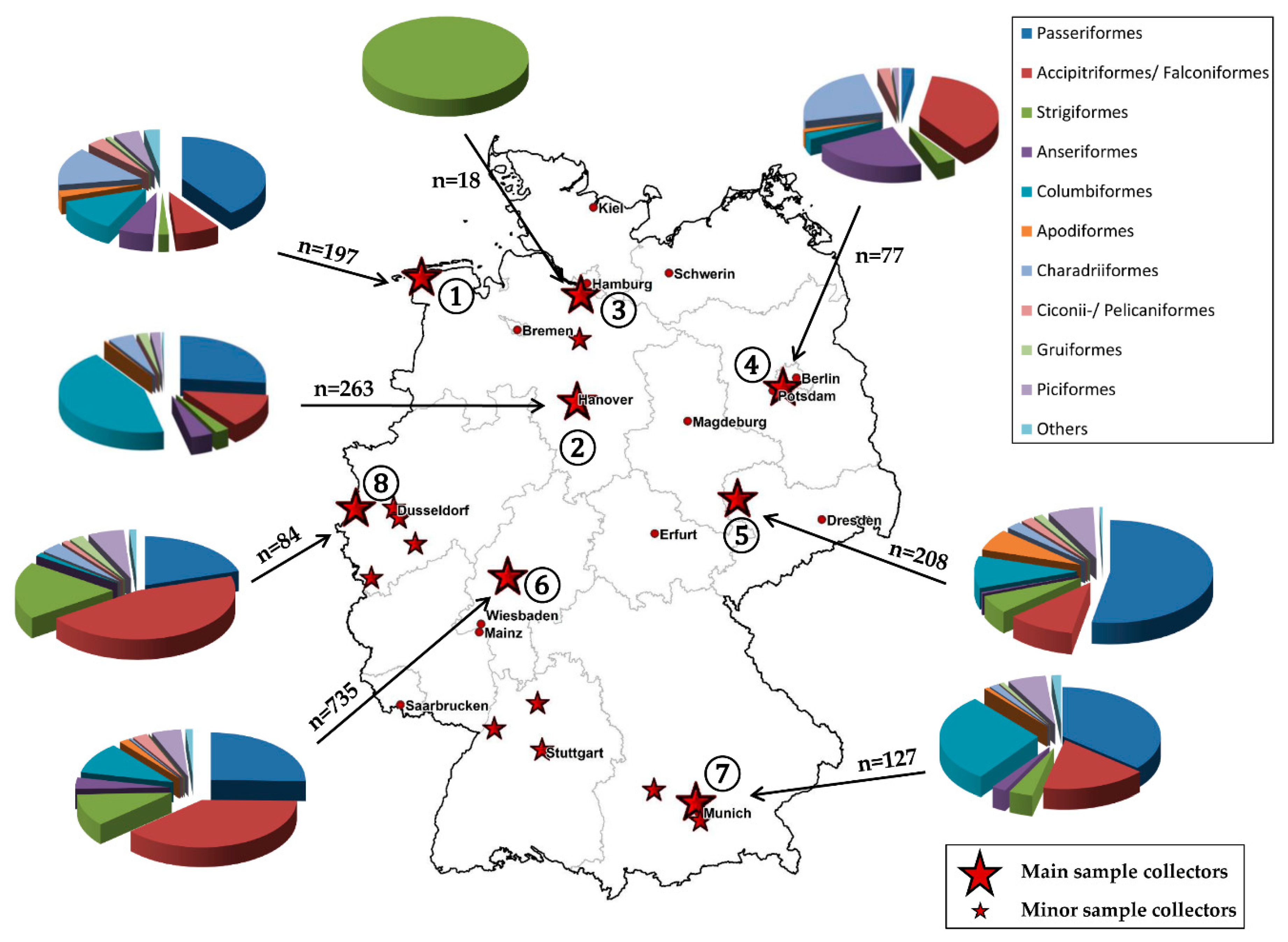
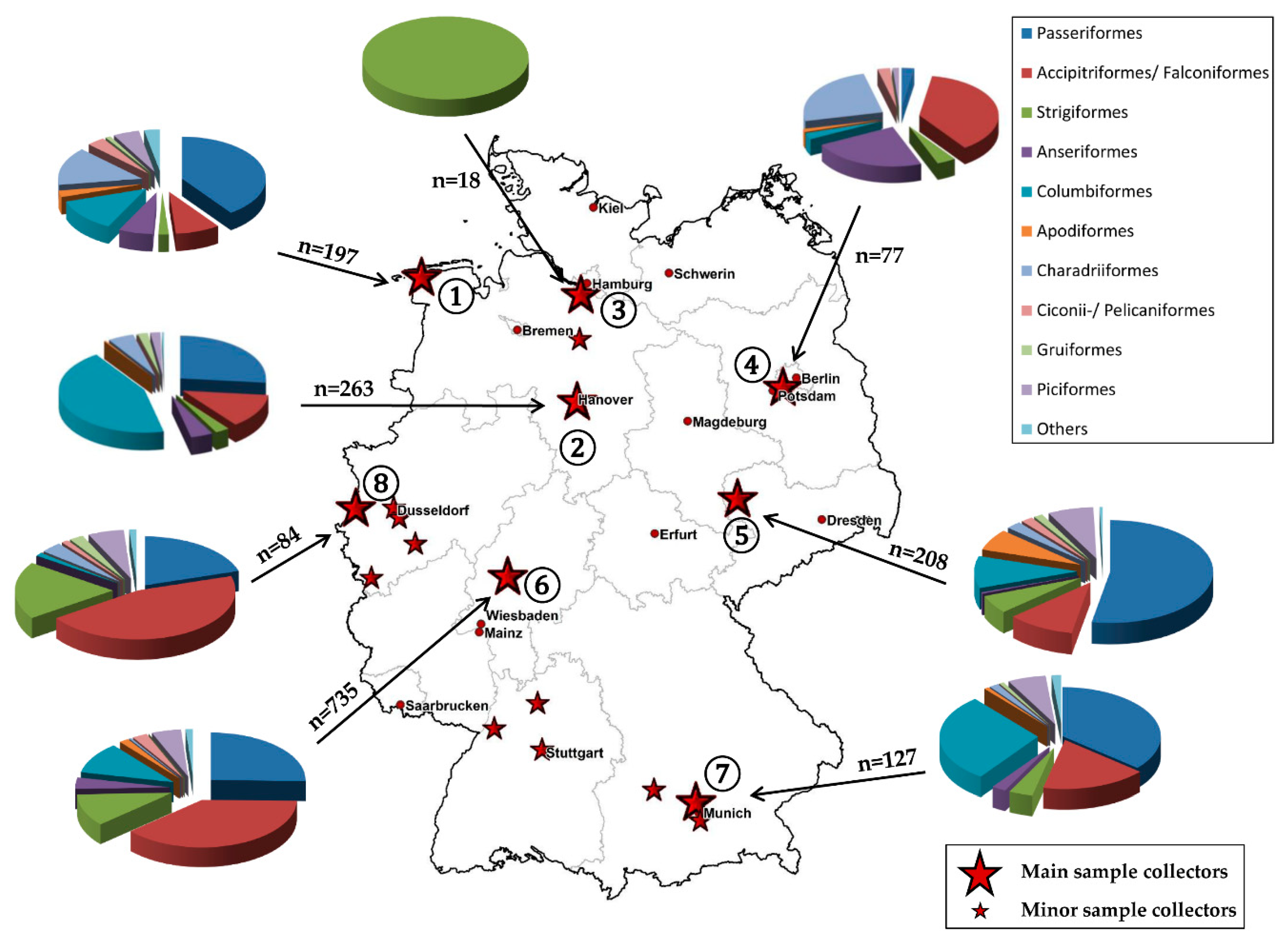
2.1. Sample Collection
2.2. Ethical Statement
2.3. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) and Phylogenetic Analysis
2.4. Serological Investigations
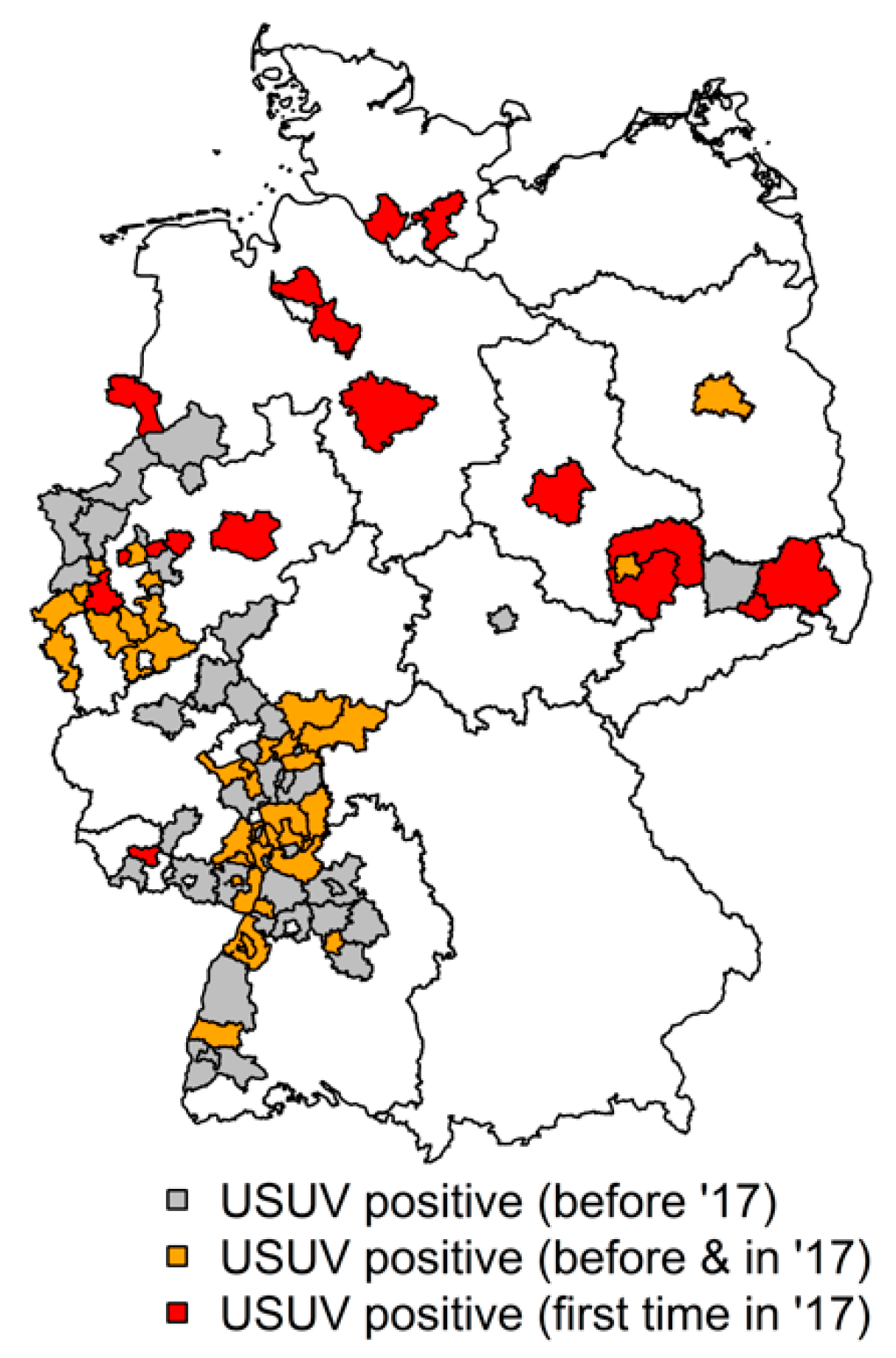
2.5. Maps
3. Results
3.1. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) Results
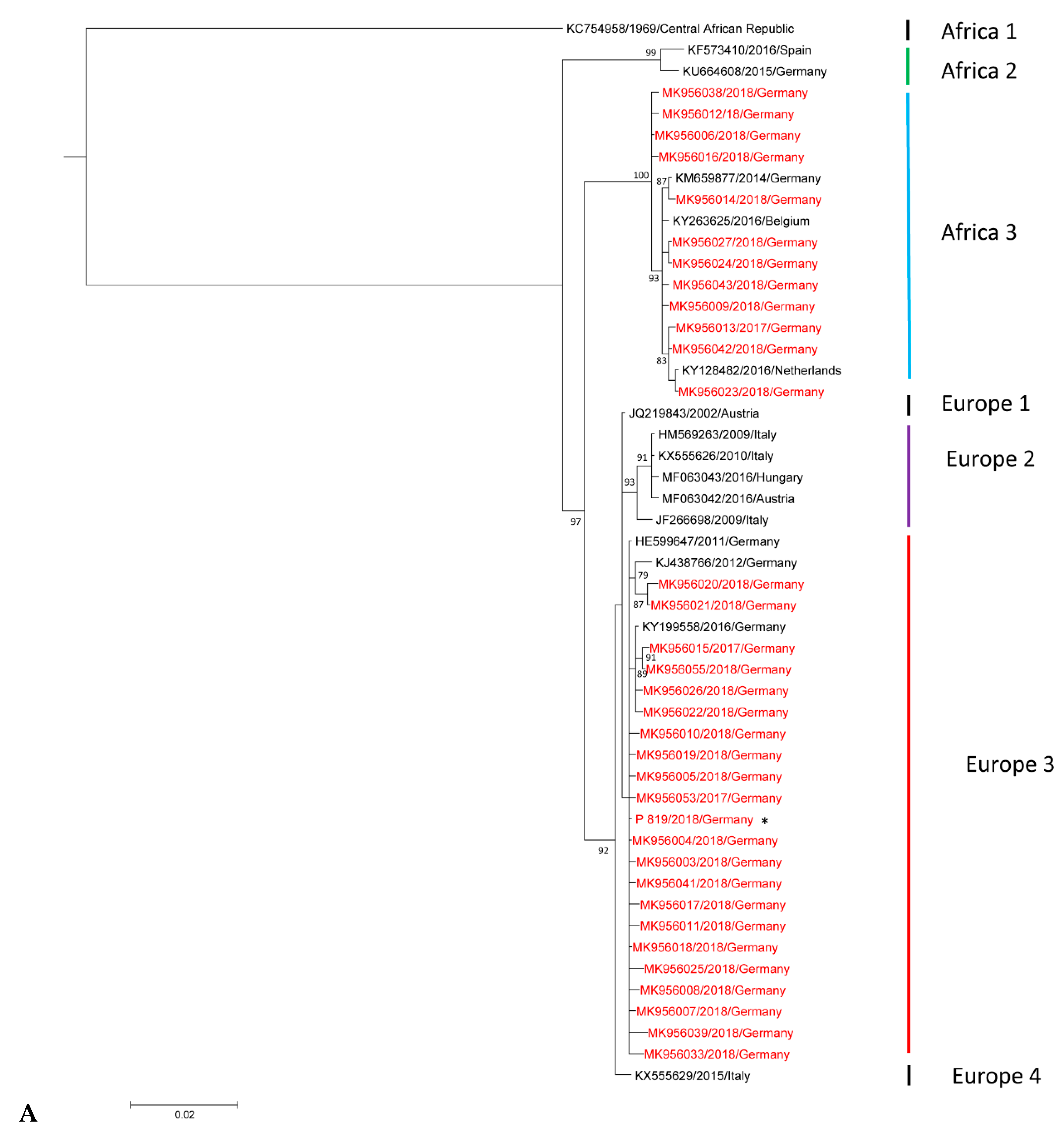
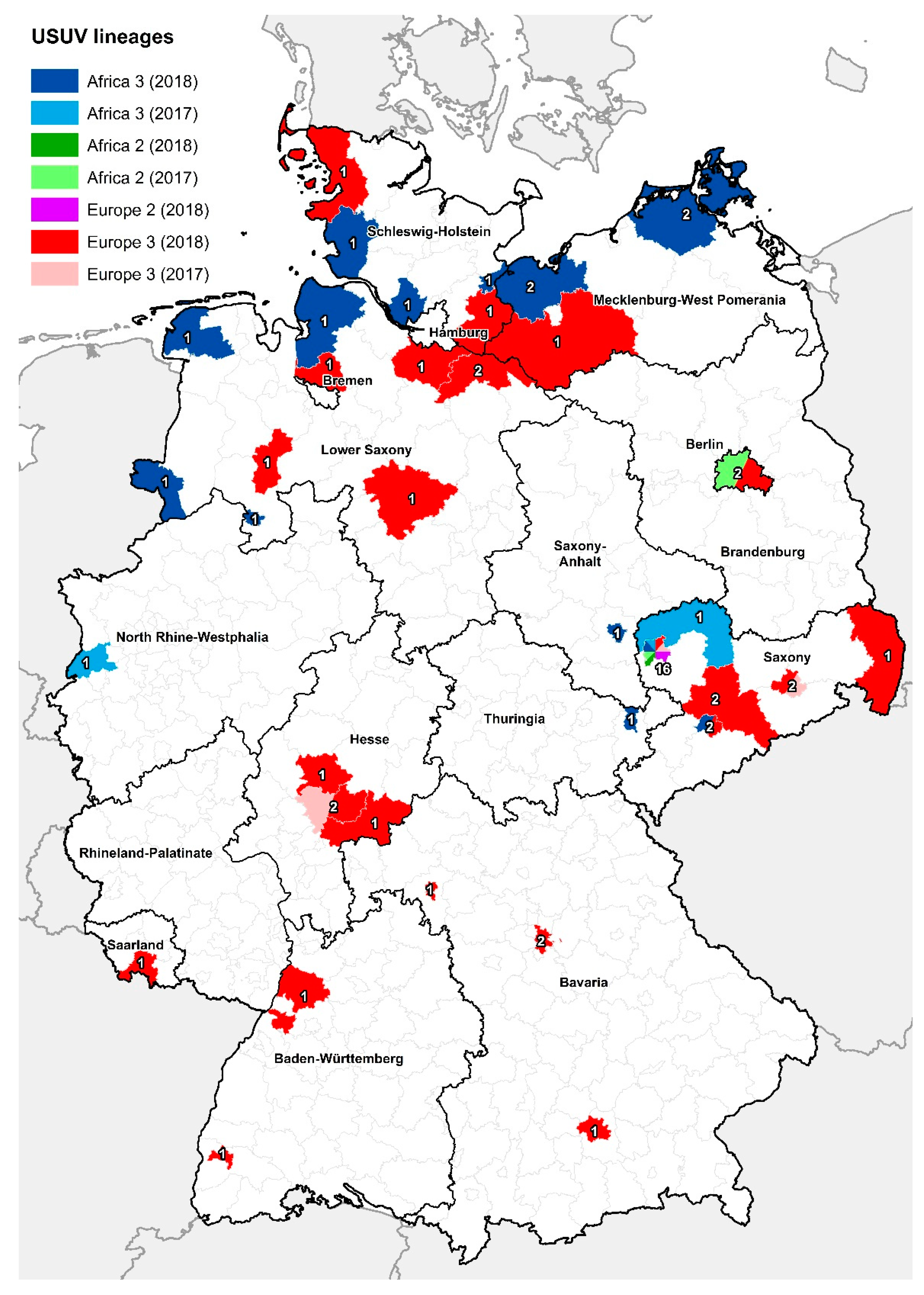
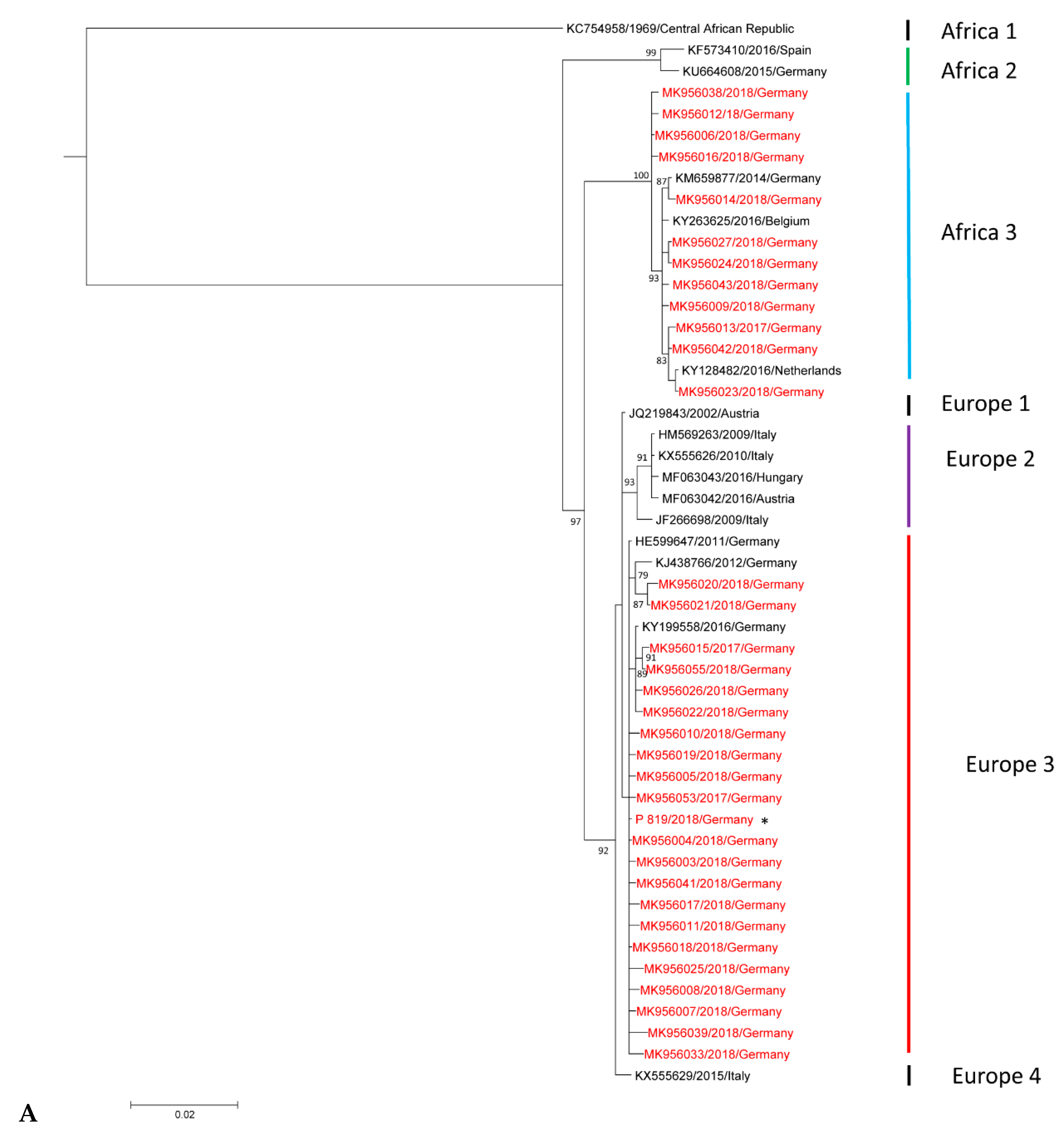
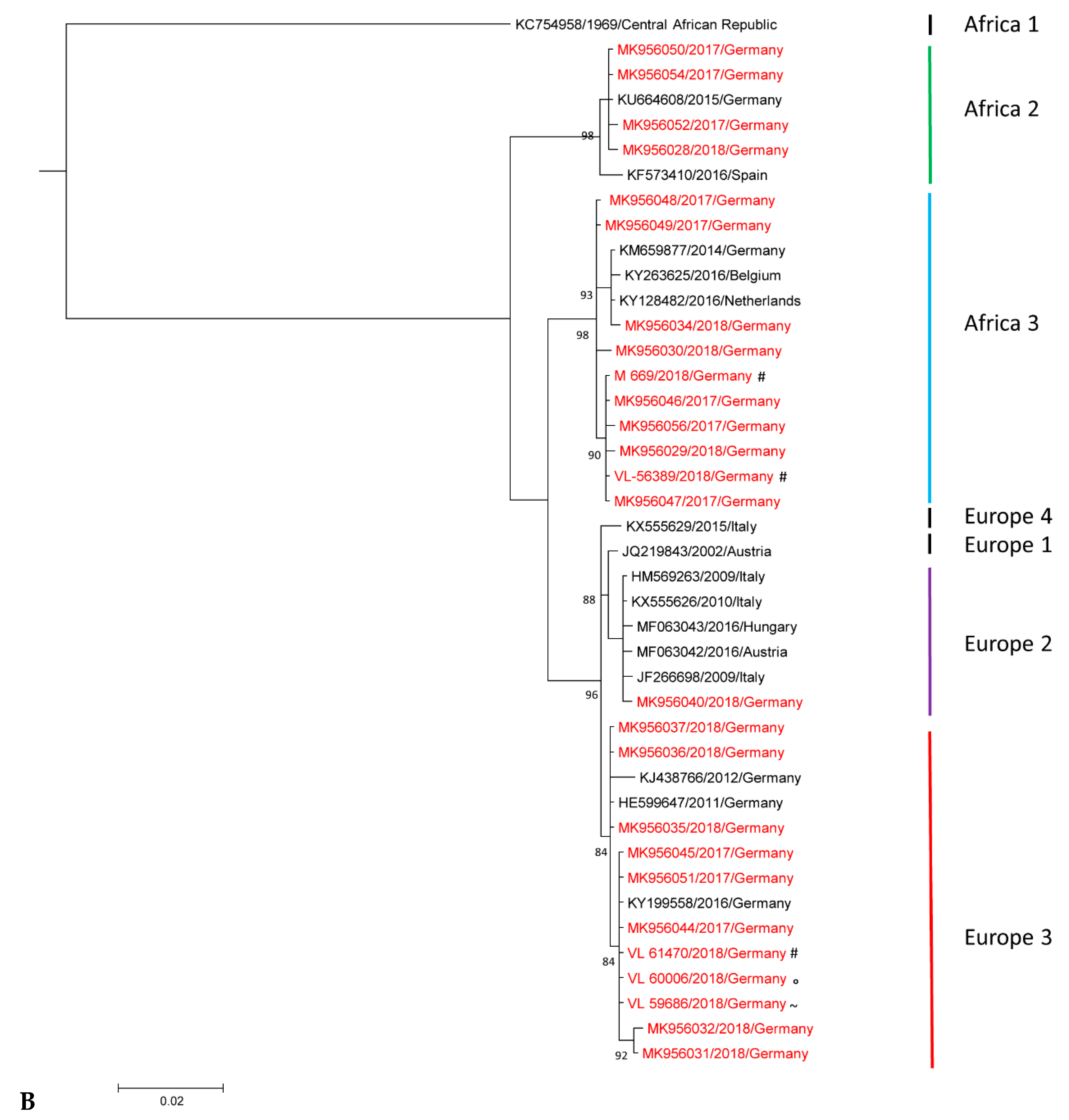
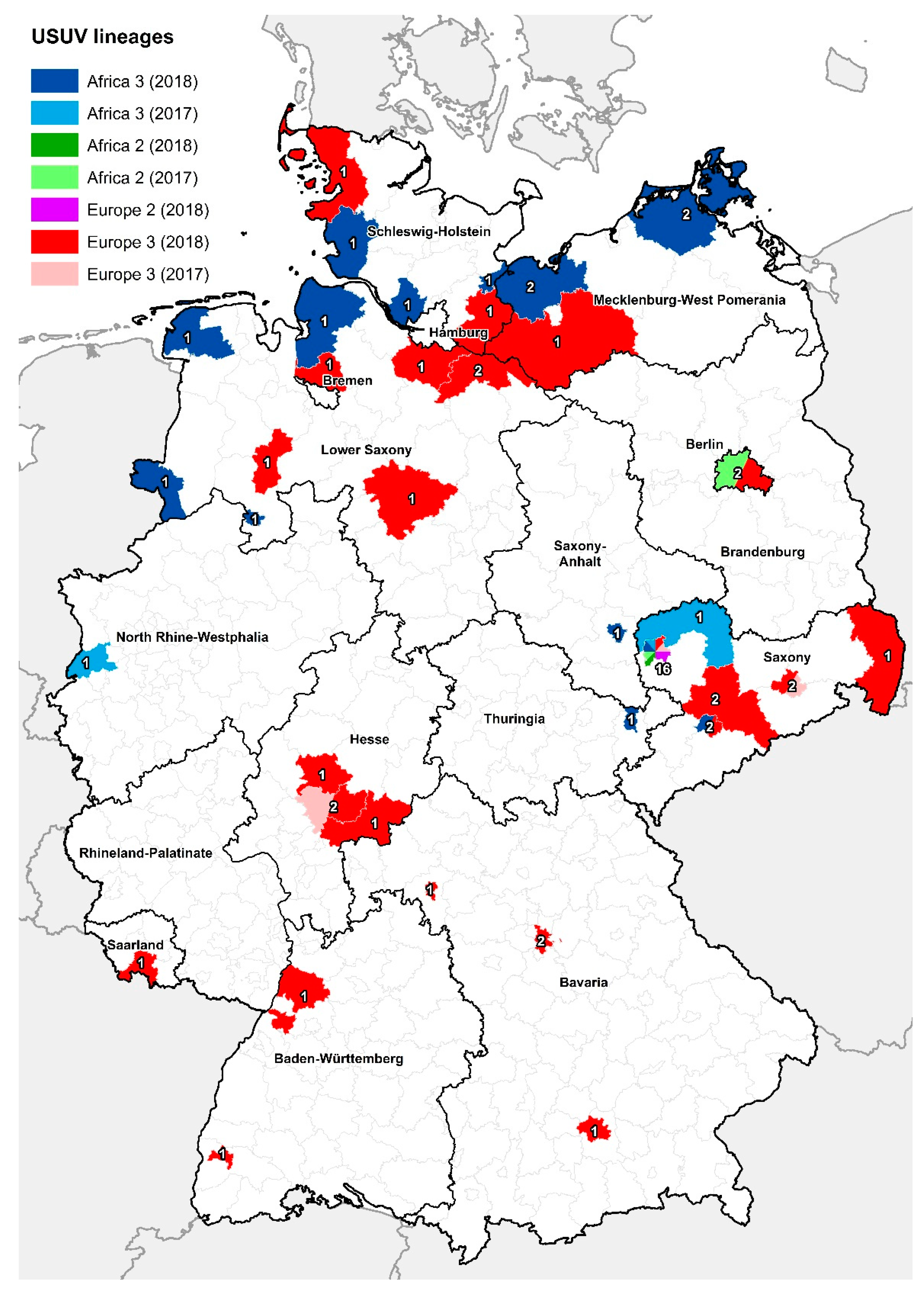
3.2. Phylogenetic Analysis of Usutu virus (USUV)-Positive Samples
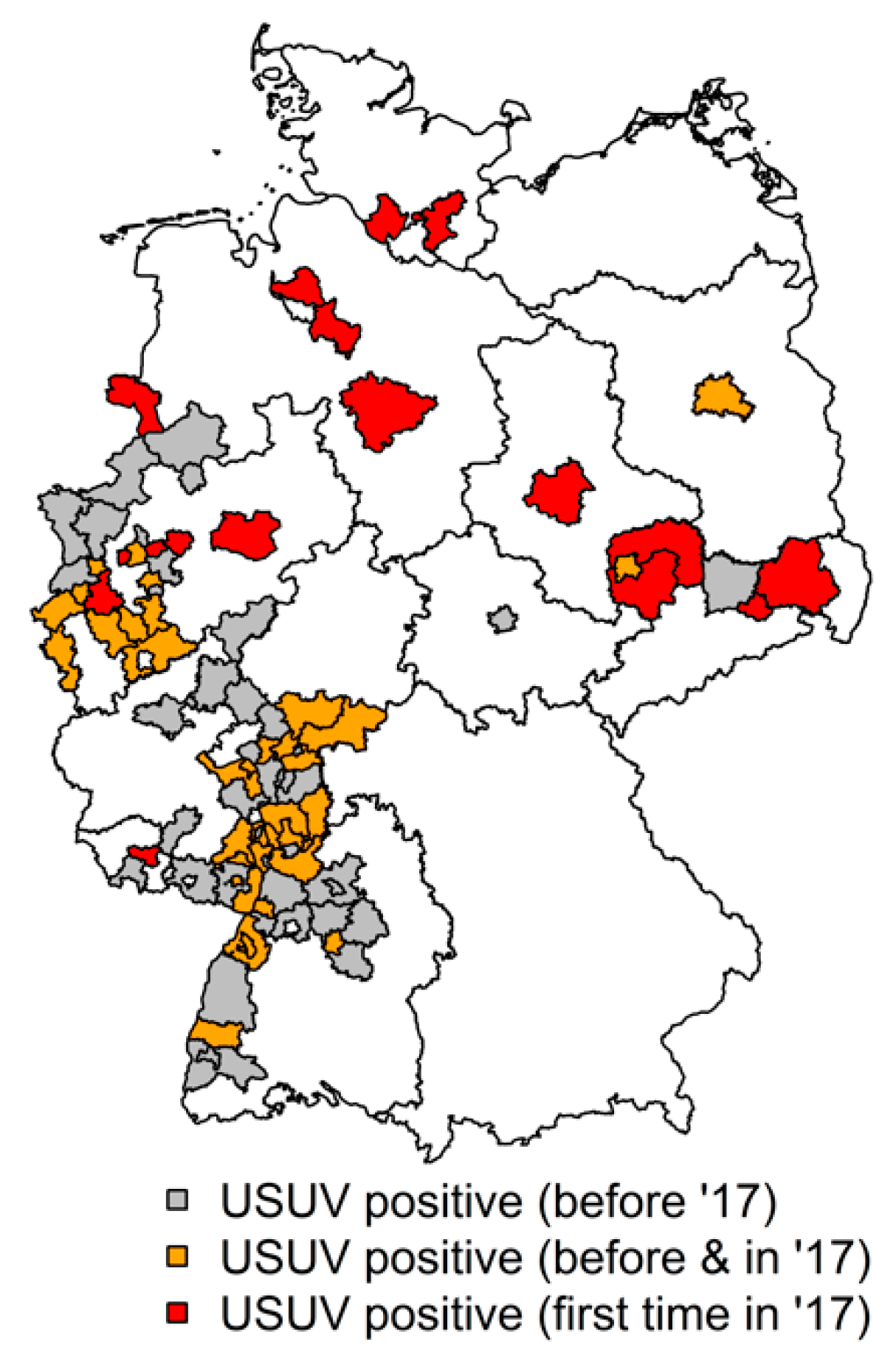
3.3. Serological Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hubalek, Z. An annotated checklist of pathogenic microorganisms associated with migratory birds. J. Wildl Dis 2004, 40, 639–659. [Google Scholar] [CrossRef] [PubMed]
- Rappole, J.H.; Hubalek, Z. Migratory birds and West Nile virus. J. Appl. Microbiol. 2003, 94, 47–58. [Google Scholar] [CrossRef]
- Seidowski, D.; Ziegler, U.; von Rönn, J.A.; Müller, K.; Hüppop, K.; Müller, T.; Freuling, C.; Mühle, R.U.; Nowotny, N.; Ulrich, R.G.; et al. West Nile virus monitoring of migratory and resident birds in Germany. Vector Borne Zoonotic Dis. 2010, 10, 639–647. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, U.; Seidowski, D.; Angenvoort, J.; Eiden, M.; Müller, K.; Nowotny, N.; Groschup, M.H. Monitoring of West Nile virus infections in Germany. Zoonoses Public Health 2012, 59, 95–101. [Google Scholar] [CrossRef]
- Michel, F.; Fischer, D.; Eiden, M.; Fast, C.; Reuschel, M.; Müller, K.; Rinder, M.; Urbaniak, S.; Brandes, F.; Schwehn, R.; et al. West Nile Virus and Usutu Virus Monitoring of Wild Birds in Germany. Int. J. Environ. Res. Public Health 2018, 15, 171. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, U.; Jöst, H.; Müller, K.; Fischer, D.; Rinder, M.; Tietze, D.T.; Danner, K.J.; Becker, N.; Skuballa, J.; Hamann, H.P.; et al. Epidemic Spread of Usutu Virus in Southwest Germany in 2011 to 2013 and Monitoring of Wild Birds for Usutu and West Nile Viruses. Vector Borne Zoonotic Dis. 2015, 15, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Calisher, C.H.; Gould, E.A. Taxonomy of the virus family Flaviviridae. Adv. Virus Res. 2003, 59, 1–19. [Google Scholar]
- McVey, D.S.; Wilson, W.C.; Gay, C.G. West Nile virus. Rev. Sci. Tech. 2015, 34, 431–439. [Google Scholar] [CrossRef]
- Chancey, C.; Grinev, A.; Volkova, E.; Rios, M. The global ecology and epidemiology of West Nile virus. Biomed. Res. Int. 2015, 2015, 376230. [Google Scholar] [CrossRef]
- Ciota, A.T. West Nile virus and its vectors. Curr. Opin. Insect. Sci. 2017, 22, 28–36. [Google Scholar] [CrossRef]
- Perez-Ramirez, E.; Llorente, F.; Jimenez-Clavero, M.A. Experimental infections of wild birds with West Nile virus. Viruses 2014, 6, 752–781. [Google Scholar] [CrossRef] [PubMed]
- David, S.; Abraham, A.M. Epidemiological and clinical aspects on West Nile virus, a globally emerging pathogen. Infect. Dis. 2016, 48, 571–586. [Google Scholar] [CrossRef] [PubMed]
- Sejvar, J.J. West Nile Virus Infection. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef]
- Angenvoort, J.; Brault, A.C.; Bowen, R.A.; Groschup, M.H. West Nile viral infection of equids. Vet. Microbiol. 2013, 167, 168–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, N.; Fernandez de Marco, M.; Giovannini, A.; Ippoliti, C.; Danzetta, M.L.; Svartz, G.; Erster, O.; Groschup, M.H.; Ziegler, U.; Mirazimi, A.; et al. Emerging Mosquito-Borne Threats and the Response from European and Eastern Mediterranean Countries. Int. J. Environ. Res. Public Health 2018, 15, 2775. [Google Scholar] [CrossRef]
- Roehrig, J.T. West nile virus in the United States—A historical perspective. Viruses 2013, 5, 3088–3108. [Google Scholar] [CrossRef]
- Hubalek, Z.; Halouzka, J. West Nile fever—A reemerging mosquito-borne viral disease in Europe. Emerg. Infect. Dis. 1999, 5, 643–650. [Google Scholar] [CrossRef]
- Bakonyi, T.; Ferenczi, E.; Erdelyi, K.; Kutasi, O.; Csörgo, T.; Seidel, B.; Weissenböck, H.; Brugger, K.; Ban, E.; Nowotny, N. Explosive spread of a neuroinvasive lineage 2 West Nile virus in Central Europe, 2008/2009. Vet. Microbiol. 2013, 165, 61–70. [Google Scholar] [CrossRef]
- Wodak, E.; Richter, S.; Bago, Z.; Revilla-Fernandez, S.; Weissenbock, H.; Nowotny, N.; Winter, P. Detection and molecular analysis of West Nile virus infections in birds of prey in the eastern part of Austria in 2008 and 2009. Vet. Microbiol. 2011, 149, 358–366. [Google Scholar] [CrossRef] [Green Version]
- Papa, A.; Bakonyi, T.; Xanthopoulou, K.; Vazquez, A.; Tenorio, A.; Nowotny, N. Genetic characterization of West Nile virus lineage 2, Greece, 2010. Emerg. Infect. Dis. 2011, 17, 920–922. [Google Scholar] [CrossRef]
- Popovic, N.; Milosevic, B.; Urosevic, A.; Poluga, J.; Lavadinovic, L.; Nedelijkovic, J.; Jevtovic, D.; Dulovic, O. Outbreak of West Nile virus infection among humans in Serbia, August to October 2012. Eurosurveillance 2013, 18, 20613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barzon, L.; Papa, A.; Lavezzo, E.; Franchin, E.; Pacenti, M.; Sinigaglia, A.; Masi, G.; Trevisan, M.; Squarzon, L.; Toppo, S.; et al. Phylogenetic characterization of Central/Southern European lineage 2 West Nile virus: Analysis of human outbreaks in Italy and Greece, 2013–2014. Clin. Microbiol. Infect. 2015, 21. [Google Scholar] [CrossRef] [PubMed]
- Riccardo, F.; Monaco, F.; Bella, A.; Savini, G.; Russo, F.; Cagarelli, R.; Dottori, M.; Rizzo, C.; Venturi, G.; Di Luca, M.; et al. An early start of West Nile virus seasonal transmission: The added value of One Heath surveillance in detecting early circulation and triggering timely response in Italy. Eurosurveillance 2018, 23. [Google Scholar] [CrossRef] [PubMed]
- Haussig, J.M.; Young, J.J.; Gossner, C.M.; Mezei, E.; Bella, A.; Sirbu, A.; Pervanidou, D.; Drakulovic, M.B.; Sudre, B. Early start of the West Nile fever transmission season 2018 in Europe. Eurosurveillance 2018, 23. [Google Scholar] [CrossRef] [PubMed]
- ECDC European Center for Disease Prevention and Control (ECDC). Weekly Updates: 2018 West Nile Fever Transmission Season. Available online: https://ecdc.europa.eu/en/west-nile-fever/surveillance-and-disease-data/disease-data-ecdc (accessed on 1 June 2019).
- Semenza, J.C.; Suk, J.E. Vector-borne diseases and climate change: A European perspective. FEMS Microbiol. Lett. 2018, 365. [Google Scholar] [CrossRef] [PubMed]
- Scheuch, D.E.; Schäfer, M.; Eiden, M.; Heym, E.C.; Ziegler, U.; Walther, D.; Schmidt-Chanasit, J.; Keller, M.; Groschup, M.H.; Kampen, H. Detection of Usutu, Sindbis, and Batai Viruses in Mosquitoes (Diptera: Culicidae) Collected in Germany, 2011–2016. Viruses 2018, 10, 389. [Google Scholar] [CrossRef]
- Leggewie, M.; Badusche, M.; Rudolf, M.; Jansen, S.; Börstler, J.; Krumkamp, R.; Huber, K.; Krüger, A.; Schmidt-Chanasit, J.; Tannich, E.; et al. Culex pipiens and Culex torrentium populations from Central Europe are susceptible to West Nile virus infection. One Health 2016, 2, 88–94. [Google Scholar] [CrossRef] [Green Version]
- Becker, N.; Krüger, A.; Kuhn, C.; Plenge-Bönig, A.; Thomas, S.M.; Schmidt-Chanasit, J.; Tannich, E. Mosquitoes as vectors for exotic pathogens in Germany. Bundesgesundheitsblatt Gesundh. Gesundh. 2014, 57, 531–540. [Google Scholar] [CrossRef]
- Jansen, S.; Heitmann, A.; Lühken, R.; Leggewie, M.; Helms, M.; Badusche, M.; Rossini, G.; Schmidt-Chanasit, J.; Tannich, E. Culex torrentium: A Potent Vector for the Transmission of West Nile Virus in Central Europe. Viruses 2019, 11, 492. [Google Scholar] [CrossRef]
- Ziegler, U.; Lühken, R.; Keller, M.; Cadar, D.; van der Grinten, E.; Michel, F.; Albrecht, K.; Eiden, M.; Rinder, M.; Lachmann, L.; et al. West Nile virus epizootic in Germany, 2018. Antiviral Res. 2019, 162, 39–43. [Google Scholar] [CrossRef]
- Gaibani, P.; Rossini, G. An overview of Usutu virus. Microbes Infect. 2017, 19, 382–387. [Google Scholar] [CrossRef] [PubMed]
- Weissenböck, H.; Bakonyi, T.; Rossi, G.; Mani, P.; Nowotny, N. Usutu virus, Italy, 1996. Emerg. Infect. Dis. 2013, 19, 274–277. [Google Scholar] [CrossRef] [PubMed]
- Weissenböck, H.; Kolodziejek, J.; Fragner, K.; Kuhn, R.; Pfeffer, M.; Nowotny, N. Usutu virus activity in Austria, 2001–2002. Microbes Infect. 2003, 5, 1132–1136. [Google Scholar] [CrossRef]
- Chvala, S.; Kolodziejek, J.; Nowotny, N.; Weissenbock, H. Pathology and viral distribution in fatal Usutu virus infections of birds from the 2001 and 2002 outbreaks in Austria. J. Comp. Pathol. 2004, 131, 176–185. [Google Scholar] [CrossRef]
- Weissenböck, H.; Kolodziejek, J.; Url, A.; Lussy, H.; Rebel-Bauder, B.; Nowotny, N. Emergence of Usutu virus, an African mosquito-borne flavivirus of the Japanese encephalitis virus group, central Europe. Emerg. Infect. Dis. 2002, 8, 652–656. [Google Scholar] [CrossRef] [PubMed]
- Jöst, H.; Bialonski, A.; Maus, D.; Sambri, V.; Eiden, M.; Groschup, M.H.; Günther, S.; Becker, N.; Schmidt-Chanasit, J. Isolation of usutu virus in Germany. Am. J. Trop. Med. Hyg. 2011, 85, 551–553. [Google Scholar] [CrossRef] [PubMed]
- Becker, N.; Jöst, H.; Ziegler, U.; Eiden, M.; Höper, D.; Emmerich, P.; Fichet-Calvet, E.; Ehichioya, D.U.; Czajka, C.; Gabriel, M.; et al. Epizootic emergence of Usutu virus in wild and captive birds in Germany. PLoS ONE 2012, 7, e32604. [Google Scholar] [CrossRef]
- Ziegler, U.; Fast, C.; Eiden, M.; Bock, S.; Schulze, C.; Hoeper, D.; Ochs, A.; Schlieben, P.; Keller, M.; Zielke, D.E.; et al. Evidence for an independent third Usutu virus introduction into Germany. Vet. Microbiol. 2016, 192, 60–66. [Google Scholar] [CrossRef]
- Cadar, D.; Bosch, S.; Jöst, H.; Börstler, J.; Garigliany, M.M.; Becker, N.; Schmidt-Chanasit, J. Putative Lineage of Novel African Usutu Virus, Central Europe. Emerg. Infect. Dis. 2015, 21, 1647–1650. [Google Scholar] [CrossRef] [Green Version]
- Cadar, D.; Lühken, R.; van der Jeugd, H.; Garigliany, M.; Ziegler, U.; Keller, M.; Lahoreau, J.; Lachmann, L.; Becker, N.; Kik, M.; et al. Widespread activity of multiple lineages of Usutu virus, western Europe, 2016. Eurosurveillance 2017, 22. [Google Scholar] [CrossRef]
- Sieg, M.; Schmidt, V.; Ziegler, U.; Keller, M.; Höper, D.; Heenemann, K.; Rückner, A.; Nieper, H.; Muluneh, A.; Groschup, M.H.; et al. Outbreak and Cocirculation of Three Different Usutu Virus Strains in Eastern Germany. Vector Borne Zoonotic Dis. 2017, 17, 662–664. [Google Scholar] [CrossRef] [PubMed]
- Grottola, A.; Marcacci, M.; Tagliazucchi, S.; Gennari, W.; Di Gennaro, A.; Orsini, M.; Monaco, F.; Marchegiano, P.; Marini, V.; Meacci, M.; et al. Usutu virus infections in humans: A retrospective analysis in the municipality of Modena, Italy. Clin. Microbiol. Infect. 2017, 23, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Cadar, D.; Maier, P.; Müller, S.; Kress, J.; Chudy, M.; Bialonski, A.; Schlaphof, A.; Jansen, S.; Jöst, H.; Tannich, E.; et al. Blood donor screening for West Nile virus (WNV) revealed acute Usutu virus (USUV) infection, Germany, September 2016. Eurosurveillance 2017, 22. [Google Scholar] [CrossRef] [PubMed]
- Allering, L.; Jöst, H.; Emmerich, P.; Günther, S.; Lattwein, E.; Schmidt, M.; Seifried, E.; Sambri, V.; Hourfar, K.; Schmidt-Chanasit, J. Detection of Usutu virus infection in a healthy blood donor from south-west Germany, 2012. Eurosurveillance 2012, 17, 20341. [Google Scholar] [PubMed]
- Carletti, F.; Colavita, F.; Rovida, F.; Percivalle, E.; Baldanti, F.; Ricci, I.; De Liberato, C.; Rosone, F.; Messina, F.; Lalle, E.; et al. Expanding Usutu virus circulation in Italy: Detection in the Lazio region, central Italy, 2017 to 2018. Eurosurveillance 2019, 24. [Google Scholar] [CrossRef]
- Eiden, M.; Vina-Rodriguez, A.; Hoffmann, B.; Ziegler, U.; Groschup, M.H. Two new real-time quantitative reverse transcription polymerase chain reaction assays with unique target sites for the specific and sensitive detection of lineages 1 and 2 West Nile virus strains. J. Vet. Diagn. Investig. 2010, 22, 748–753. [Google Scholar] [CrossRef] [PubMed]
- Cavrini, F.; Della Pepa, M.E.; Gaibani, P.; Pierro, A.M.; Rossini, G.; Landini, M.P.; Sambri, V. A rapid and specific real-time RT-PCR assay to identify Usutu virus in human plasma, serum, and cerebrospinal fluid. J. Clin. Virol. 2011, 50, 221–223. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Hijmans, R.J. Raster: Geographic Data Analysis and Modeling. R Package Version 2.8-4. Available online: https://CRAN.R-project.org/package=raster (accessed on 11 July 2019).
- Bache, S.M.; Wickham, H. Magrittr: A Forward-Pipe Operator for R. R Package Version 1.5. Available online: https://CRAN.R-project.org/package=magrittr (accessed on 29 April 2019).
- Bivand, R.; Lewin-Koh, N. Maptools: Tools for Handling Spatial Objects. R Package Version 0.9-4. 2018. Available online: http://r-forge.r-project.org/projects/maptools/ (accessed on 18 February 2019).
- R Core Team: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018.
- Bahuon, C.; Marcillaud-Pitel, C.; Bournez, L.; Leblond, A.; Beck, C.; Hars, J.; Leparc-Goffart, I.; L’Ambert, G.; Paty, M.C.; Cavalerie, L.; et al. West Nile virus epizootics in the Camargue (France) in 2015 and reinforcement of surveillance and control networks. Rev. Sci. Tech. 2016, 35, 811–824. [Google Scholar] [CrossRef] [Green Version]
- Aberle, S.W.; Kolodziejek, J.; Jungbauer, C.; Stiasny, K.; Aberle, J.H.; Zoufaly, A.; Hourfar, M.K.; Weidner, L.; Nowotny, N. Increase in human West Nile and Usutu virus infections, Austria, 2018. Eurosurveillance 2018, 23. [Google Scholar] [CrossRef]
- Rudolf, I.; Bakonyi, T.; Sebesta, O.; Mendel, J.; Pesko, J.; Betasova, L.; Blazejova, H.; Venclikova, K.; Strakova, P.; Nowotny, N.; et al. West Nile virus lineage 2 isolated from Culex modestus mosquitoes in the Czech Republic, 2013: Expansion of the European WNV endemic area to the North? Eurosurveillance 2014, 19, 2–5. [Google Scholar] [CrossRef] [PubMed]
- Hubalek, Z.; Kosina, M.; Rudolf, I.; Mendel, J.; Strakova, P.; Tomesek, M. Mortality of Goshawks (Accipiter gentilis) Due to West Nile Virus Lineage 2. Vector Borne Zoonotic Dis. 2018, 18, 624–627. [Google Scholar] [CrossRef] [PubMed]
- Eldin, C.; Mailhe, M.; Zandotti, C.; Grard, G.; Galla, M.; Parola, P.; Brouqui, P.; Lagier, J.C. West Nile virus outbreak in the South of France: Implications for travel medicine. Travel Med. Infect. Dis. 2019, 28, 100–101. [Google Scholar] [CrossRef] [PubMed]
- Bazanow, B.; Jansen van Vuren, P.; Szymanski, P.; Stygar, D.; Fracka, A.; Twardon, J.; Kozdrowski, R.; Paweska, J.T. A Survey on West Nile and Usutu Viruses in Horses and Birds in Poland. Viruses 2018, 10, 87. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, N.; Gould, D.; Bowen, R.; Komar, N. Natural and experimental West Nile virus infection in five raptor species. J. Wildl. Dis. 2006, 42, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, U.; Angenvoort, J.; Fischer, D.; Fast, C.; Eiden, M.; Rodriguez, A.V.; Revilla-Fernandez, S.; Nowotny, N.; de la Fuente, J.G.; Lierz, M.; et al. Pathogenesis of West Nile virus lineage 1 and 2 in experimentally infected large falcons. Vet. Microbiol. 2013, 161, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Busquets, N.; Bertran, K.; Costa, T.P.; Rivas, R.; de la Fuente, J.G.; Villalba, R.; Solanes, D.; Bensaid, A.; Majo, N.; Pages, N. Experimental West Nile virus infection in Gyr-Saker hybrid falcons. Vector Borne Zoonotic Dis. 2012, 12, 482–489. [Google Scholar] [CrossRef]
- Nikolay, B. A review of West Nile and Usutu virus co-circulation in Europe: How much do transmission cycles overlap? Trans. R. Soc. Trop. Med. Hyg. 2015, 109, 609–618. [Google Scholar] [CrossRef]
- Gamino, V.; Höfle, U. Pathology and tissue tropism of natural West Nile virus infection in birds: A review. Vet. Res. 2013, 44, 39. [Google Scholar] [CrossRef]
- Wheeler, S.S.; Langevin, S.A.; Brault, A.C.; Woods, L.; Carroll, B.D.; Reisen, W.K. Detection of persistent west nile virus RNA in experimentally and naturally infected avian hosts. Am. J. Trop. Med. Hyg. 2012, 87, 559–564. [Google Scholar] [CrossRef]
- Ziegler, U.; Federal Friedrich-Loeffler Insitut (FLI), Federal Research Institute for Animal Health, Institute of Novel and Emerging Infectious Diseases, D-17493 Greifswald-Insel Riems, Germany; Lühken, R.; Bernhard-Nocht-Institute for Tropical Medicine, WHO Collaborating Centre for Arbovirus and Hemorrhagic Fever Reference and Research, D-20359 Hamburg, Germany. Personal communication, 2019.
- Lühken, R.; Jöst, H.; Cadar, D.; Thomas, S.M.; Bosch, S.; Tannich, E.; Becker, N.; Ziegler, U.; Lachmann, L.; Schmidt-Chanasit, J. Distribution of Usutu Virus in Germany and Its Effect on Breeding Bird Populations. Emerg. Infect. Dis. 2017, 23, 1994–2001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konrad, A. Usutu Virus Associated Population Decline of Common Blackbirds in the Northern Upper Rhine Plain in the Summer 2011. Avifauna-Nordbad, 2 November 2011. [Google Scholar]
- Bakonyi, T.; Jungbauer, C.; Aberle, S.W.; Kolodziejek, J.; Dimmel, K.; Stiasny, K.; Allerberger, F.; Nowotny, N. Usutu virus infections among blood donors, Austria, July and August 2017—Raising awareness for diagnostic challenges. Eurosurveillance 2017, 22. [Google Scholar] [CrossRef] [PubMed]
- Calzolari, M.; Bonilauri, P.; Bellini, R.; Albieri, A.; Defilippo, F.; Maioli, G.; Galletti, G.; Gelati, A.; Barbieri, I.; Tamba, M.; et al. Evidence of simultaneous circulation of West Nile and Usutu viruses in mosquitoes sampled in Emilia-Romagna region (Italy) in 2009. PLoS ONE 2010, 5, e14324. [Google Scholar] [CrossRef] [PubMed]
- Engel, D.; Jöst, H.; Wink, M.; Börstler, J.; Bosch, S.; Garigliany, M.M.; Jöst, A.; Czajka, C.; Lühken, R.; Ziegler, U.; et al. Reconstruction of the Evolutionary History and Dispersal of Usutu Virus, a Neglected Emerging Arbovirus in Europe and Africa. MBio 2016, 7, e01938-15. [Google Scholar] [CrossRef] [PubMed]
- Bakonyi, T.; Erdelyi, K.; Brunthaler, R.; Dan, A.; Weissenbock, H.; Nowotny, N. Usutu virus, Austria and Hungary, 2010–2016. Emerg. Microbes Infect. 2017, 6, e85. [Google Scholar] [CrossRef]
- Meister, T.; Lussy, H.; Bakonyi, T.; Sikutova, S.; Rudolf, I.; Vogl, W.; Winkler, H.; Frey, H.; Hubalek, Z.; Nowotny, N.; et al. Serological evidence of continuing high Usutu virus (Flaviviridae) activity and establishment of herd immunity in wild birds in Austria. Vet. Microbiol. 2008, 127, 237–248. [Google Scholar] [CrossRef]
- Bosch, J.; Schmidt-Chanasit, J.; Fiedler, W. Usutu Virus as cause of mass mortality in Blackbirds (Turdus merula) and other bird species in Europe: Experiences from five outbreaks between 2001 and 2011. Vogelwarte 2012, 50, 109–122. [Google Scholar]
- Ziegler, U.; Federal Friedrich-Loeffler Insitut (FLI), Federal Research Institute for Animal Health, Institute of Novel and Emerging Infectious Diseases, D-17493 Greifswald-Insel Riems, Germany. Personal communication, 2019.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Order | Year 2017 | Year 2018 | Total |
|---|---|---|---|
| Passeriformes | 291 | 219 | 510 |
| Accipitriformes/Falconiformes | 208 | 236 | 444 |
| Strigiformes | 66 | 79 | 145 |
| Anseriformes | 26 | 49 | 75 |
| Columbiformes | 115 | 166 | 281 |
| Apodiformes | 26 | 8 | 34 |
| Charadriiformes | 25 | 49 | 74 |
| Ciconiiformes/Pelicaniformes | 13 | 20 | 33 |
| Gruiformes | 5 | 8 | 13 |
| Piciformes | 47 | 34 | 81 |
| Suliformes | 3 | 0 | 3 |
| Galliformes | 2 | 0 | 2 |
| Podicipediformes | 3 | 2 | 5 |
| Coraciiformes | 5 | 0 | 5 |
| Procellariiformes | 0 | 1 | 1 |
| Cuculiformes | 0 | 1 | 1 |
| Psittaciformes | 0 | 2 | 2 |
| Total | 835 | 874 | 1709 |
| Order | Common Name | Scientific Name | Migration Pattern | Housing | WNV qRT-PCR no.pos. | USUV qRT-PCR no.pos. | Federal State |
|---|---|---|---|---|---|---|---|
| (a) qRT-PCR results in 2017 | |||||||
| Passeriformes | Eurasian Blackbird | Turdus merula | R, P | wild | 0 | 28 | SN, ST, BE, HE |
| Common Starling | Sturnus vulgaris | R, P, S | wild | 0 | 1 | HE | |
| Carrion Crow | Corvus corone | S | wild | 0 | 1 | NRW | |
| Western Jackdaw | Coloeus monedula | R, P | wild | 0 | 1 | NRW | |
| Piciformes | Great Spottet Woodpecker | Dendrocopos major | R, P, (S) | wild | 0 | 1 | LS |
| in Total in 2017 | 0/826 | 32/826 | |||||
| (b) qRT-PCR results in 2018 | |||||||
| Passeriformes | Eurasian Blackbird | Turdus merula | R, P | wild | 0 | 13 | SN, LS, HE |
| House Sparrow | Passer domesticus | R | wild | 0 | 1 | BW | |
| Eurasian Bullfinch | Pyrrhula pyrrhula | / | captive | 0 | 2 | SN, SH | |
| Domestic Canary | Serinus canaria forma domestica | / | captive | 0 | 1 | NRW | |
| Grey-Headed Bullfinch | Pyrrhula erythaca | / | captive | 0 | 1 | LS | |
| Song Thrush | Turdus philomelos | R, S | wild | 0 | 2 | SN | |
| Common Starling | Sturnus vulgaris | R, P, S | wild | 0 | 1 | SN | |
| Strigiformes | Great Grey Owl | Strix nebulosa | / | captive | 0 | 2 | BW, HE |
| Little Owl | Athene noctua | R | wild | 0 | 1 | HE | |
| Falconiformes | European Kestrel | Falco tinnunculus | R, P, S | wild | 0 | 1 | HE |
| Total | 0/781 | 25/781 | |||||
| Order | Common Name | Scientific Name | Migration Pattern | Housing | USUV RNA Positive Birds |
|---|---|---|---|---|---|
| Passeriformes | Eurasian Blackbird | Turdus merula | R, P | wild | 94 |
| Song Trush | Turdus philomelos | R, L | wild | 1 | |
| Golden-breasted Starling | Cosmopsarus regius | / | captive | 1 | |
| Strigiformes | Great Grey Owl | Strix nebulosa | / | captive | 2 |
| Northern Hawk-owl | Surnia ulula | / | captive | 2 | |
| in Total in 2017 | 100 | ||||
| Sample Collector | Order | Common Name | Scientific Name | Migration Pattern | No. Samples Tested | WNV Pos. (ND50) | USUV Pos. (ND50) |
|---|---|---|---|---|---|---|---|
| (a) WNV and USUV positive neutralization assay results (positives highlighted in red and bold) from wild bird serum in 2017. Cross-reacting antibody titers are also displayed in black | |||||||
| ① | Piciformes | Great Spottet Woodpecker | Dendrocopos major | R, P, (S) | 7 | 0 | 1 (10) |
| ② | Accipitriformes | Common Buzzard | Buteo buteo | R, P, S | 11 | 1 (10) | 0 |
| ② | Columbiformes | Common Wood Pigeon | Columba palumbus | R, P, S | 29 | 2 (10), 1 (15) | 0 |
| Feral Pigeon | Columba livia f. domestica | R, (P) | 14 | 1 (10) | 1 (20) | ||
| ② | Charadriiformes | Eurasian Woodcock | Scolopax rusticola | R, S | 3 | 1 (10) | 1 (20) |
| ② | Suliformes | Great Cormorant | Phalacrocorax carbo | R, S | 1 | 1 (10) | 0 |
| Total | 171 | 5 | 3 | ||||
| Seroprevalence | 2.92% | 1.75% | |||||
| (b) WNV- and USUV-positive neutralization assay results (positives highlighted in red and bold) from wild bird serum in 2018. Cross-reacting antibody titers are also displayed in black | |||||||
| ① | Passeriformes | Eurasian Blackbird | Turdus merula | R, P | 16 | 1 (10), 1 (20), 1 (60) | 2 (10), 1 (15), 1 (40), 1 (60), 1 (240), 1 (320) |
| ① | Carrion Crow | Corvus corone | R, P | 14 | 1 (10) | 0 | |
| ① | Spotted Flycatcher | Muscicapa striata | L | 2 | 1 (10) | 0 | |
| ① | Accipitriformes | Northern Goshawk | Accipiter gentilis | R, P | 1 | 0 | 1 (10) |
| ① | Common Buzzard | Buteo buteo | R, P, S | 6 | 0 | 1 (40) | |
| ① | Anseriformes | Northern Mallard Duck | Anas platyrhynchos | R, P, S | 5 | 1 (20) | 1 (10), 1 (480) |
| ① | Ciconiiformes/ Pelicaniformes | Northern Gannet | Morus bassanus | S, L | 6 | 1 (10) | 0 |
| ② | Passeriformes | Eurasian Blackbird | Turdus merula | R, P | 9 | 1 (120), 1 (240) | 1 (10), 1 (240), 1 (1280) |
| ② | Eurasian Jay | Garrulus glandarius | R, P | 3 | 1 (15), 1 (40) | 1 (1280) | |
| ② | Common Magpie | Pica pica | R | 5 | 1 (15) | 0 | |
| ② | Common Starling | Sturnus vulgaris | R, P, S | 2 | 1 (40) | 1 (240) | |
| ② | Accipitriformes | Common Buzzard | Buteo buteo | R, P, S | 11 | 2 (15) | 0 |
| ② | Columbiformes | Common Wood Pigeon | Columba palumbus | R, P, S | 48 | 1 (10), 1 (15), 1 (20), 1 (40) | 1 (40), 3 (80) |
| ② | Apodiformes | Common Swift | Apus apus | L | 1 | 0 | 1 (10) |
| ③ | Strigiformes | Eurasian Eagle Owl | Bubo bubo | zoo bird | 5 | 0 | 1 (30), 1 (20), 1 (15) |
| ③ | Snowy Owl | Bubo scandiacus | zoo bird | 2 | 0 | 1 (640) | |
| ③ | Ural Owl | Strix uralensis | zoo bird | 2 | 1 (30) | 1 (320) | |
| ③ | Northern Long-eared Owl | Asio otus | zoo bird | 3 | 1 (20) | 1 (80) | |
| ③ | Eurasian Tawny Owl | Strix aluco | zoo bird | 2 | 1 (10), 1 (60) | 1 (160), 1 (2560) | |
| Total | 304 | 7 | 29 | ||||
| Seroprevalence | 2.30% | 9.54% | |||||
| Sample Collector | Order | Common Name | Scientific Name | Migration Pattern | No. Samples Tested | WNV Pos. (ND50) | USUV Pos. (ND50) |
|---|---|---|---|---|---|---|---|
| (a) WNV- and USUV-positive neutralization assay results (positives highlighted in red and bold) from wild bird serum in 2017. Cross-reacting antibody titers are also displayed in black | |||||||
| ④ | Charadriiformes | Eurasian Woodcock | Scolopax rusticola | R, S | 7 | 2 (10), 1 (20) | 1 (10), 1 (25), 1 (30), 1 (40) |
| ④ | Ciconiiformes/Pelicaniformes | Grey Heron | Ardea cinerea | R, P, S | 1 | 0 | 1 (10) |
| ⑤ | Passeriformes | Eurasian Blackbird | Turdus merula | R, P | 11 | 1 (10), 1 (15), 1 (20) | 1 (20), 1 (40), 1 (50), 1 (80) |
| ⑤ | Falconiformes | European Kestrel | Falco tinnunculus | R, P, S | 5 | 1 (20), 1 (80) | 0 |
| ⑤ | Columbiformes | Common Wood Pigeon | Columba palumbus | R, P, S | 9 | 3 (10), 1 (60) | 2 (20), 1 (30), 1 (80), 1 (320), 1 (480) |
| ⑤ | Ciconiiformes/Pelicaniformes | White Stork | Ciconia ciconia | L | 1 | 1 (20) | 1 (40) |
| Total | 76 | 2 | 16 | ||||
| Seroprevalence | 2.63% | 21.05% | |||||
| (b) WNV- and USUV-positive neutralization assay results (positives highlighted in red and bold) from wild bird serum in 2018. Cross-reacting antibody titers are also displayed in black | |||||||
| ④ | Passeriformes | Common Magpie | Pica pica | R | 1 | 0 | 1 (10) |
| ④ | Accipitriformes | Northern Goshawk | Accipiter gentilis | R, P | 2 | 1 (320) | 0 |
| ④ | White-tailed Eagle | Haliaeetus albicilla | R, P | 5 | 1 (240) | 0 | |
| ④ | Common Buzzard | Buteo buteo | R, P, S | 7 | 1 (20) | 1 (10) | |
| ④ | Falconiformes | European Kestrel | Falco tinnunculus | R, P, S | 1 | 1 (160) | 1 (40) |
| ④ | Strigiformes | Northern Long-eared Owl | Asio otus | R, P, S | 2 | 1 (10) | 1 (60) |
| ④ | Columbiformes | Common Wood Pigeon | Columba palumbus | R, P, S | 2 | 0 | 1 (10) |
| ④ | Anseriformes | Northern Mallard Duck | Anas platyrhynchos | R, P, S | 2 | 0 | 1 (10) |
| ④ | Apodiformes | Common Swift | Apus apus | L | 1 | 1 (10) | 1 (30) |
| ④ | Charadriiformes | Eurasian Woodcock | Scolopax rusticola | R, S | 12 | 0 | 1 (10) |
| ⑤ | Passeriformes | Eurasian Blackbird | Turdus merula | R, P | 3 | 0 | 1 (60) |
| ⑤ | Carrion Crow | Corvus corone | P, S | 5 | 1 (240) | 1 (40) | |
| ⑤ | Accipitriformes | Common Buzzard | Buteo buteo | R, P, S | 2 | 0 | 1 (60), 1 (10) |
| ⑤ | Columbiformes | Common Wood Pigeon | Columba palumbus | R, P, S | 7 | 1 (80), 1 (60), 1 (30) | 1 (320), 1 (80), 1 (30), 1 (10) |
| Total | 92 | 6 | 12 | ||||
| Seropevalence | 6.52% | 13.04% | |||||
| Sample Collector | Order | Common Name | Scientific Name | Migration Pattern | No. Samples Tested | WNV Pos. (ND50) | USUV Pos. (ND50) |
|---|---|---|---|---|---|---|---|
| (a) WNV- and USUV-positive neutralization assay results (positives highlighted in red and bold) from wild bird serum in 2017. Cross-reacting antibody titers are also displayed in black | |||||||
| ⑤ | Passeriformes | Eurasian Blackbird | Turdus merula | R, P | 4 | 1 (75), 1 (190) | 1 (160), 1 (3840) |
| ⑥ | Passeriformes | Eurasian Blackbird | Turdus merula | R, P | 23 | 1 (15) | 0 |
| ⑥ | Eurasian Blue Tit | Cyanistes caeruleus | R | 2 | 1 (10) | 0 | |
| ⑥ | Thrush | Turdus sp. | R, S | 10 | 1 (10) | 1 (15) | |
| ⑥ | Common Starling | Sturnus vulgaris | R, P, S | 5 | 1 (10) | 0 | |
| ⑥ | Accipitriformes | Long-legged Buzzard | Buteo rufinus | zoo bird | 2 | 0 | 1 (10) |
| ⑥ | Common Buzzard | Buteo buteo | R, P, S | 24 | 2 (10), 1 (10) | 1 (60), 1 (15) | |
| ⑥ | African Sea Eagle | Haliaeetus vocifer | zoo bird | 3 | 1 (30) | 0 | |
| ⑥ | Black Kite | Milvus migrans | L | 2 | 1 (10) | 0 | |
| ⑥ | Falconiformes | European Kestrel | Falco tinnunculus | R, P, S | 21 | 2 (15), 1 (30) | 0 |
| ⑥ | Columbiformes | Common Wood Pigeon | Columba palumbus | R, P, S | 16 | 0 | 2 (10), 1 (60) |
| ⑥ | Apodiformes | Common Swift | Apus apus | L | 10 | 1 (10) | 1 (15) |
| ⑥ | Ciconiiformes/Pelicaniformes | Grey Heron | Ardea cinerea | R, P, S | 7 | 1 (15) | 1 (40) |
| ⑥ | Stork sp. | Circonia sp. | L | 1 | 1 (10) | 0 | |
| ⑦ | Gruiformes | Eurasian coot | Fulica atra | P, S | 1 | 1 (15) | 1 (30) |
| ⑧ | Accipitriformes | Common Buzzard | Buteo buteo | R, P, S | 13 | 1 (20), 1 (10) | 2 (320), 1 (20), 1 (15), 1 (10) |
| ⑧ | Northern Goshawk | Accipiter gentilis | R, P | 2 | 0 | 1 (15) | |
| ⑧ | Strigiformes | Eurasian Tawny Owl | Strix aluco | R | 2 | 0 | 1 (80) |
| ⑧ | Gruiformes | Eurasian Coot | Fulica atra | P, S | 1 | 1 (10) | 0 |
| Total | 492 | 15 | 19 | ||||
| Seroprevalence | 3.05% | 3.86% | |||||
| (b) WNV- and USUV-positive neutralization assay results (positives highlighted in red and bold) from wild bird serum in 2018. Cross-reacting antibody titers are also displayed in black | |||||||
| ⑥ | Passeriformes | Eurasian Blackbird | Turdus merula | R, P, S | 19 | 2 (15), 1 (60), 1 (10) | 1 (320), 1 (30), 1 (20) |
| ⑥ | Common Magpie | Pica pica | R | 6 | 1 (15) | 0 | |
| ⑥ | Carrion Crow | Corvus corone | P, S | 9 | 1 (40) | 0 | |
| ⑥ | European Robin | Erithacus rubecula | P | 2 | 0 | 1 (15) | |
| ⑥ | Accipitriformes/Falconiformes | Black-chested Buzzard-eagle | Geranoaetus melanoleucus | zoo bird | 1 | 0 | 1 (10) |
| ⑥ | Griffon Vulture | Gyps fulvus | zoo bird | 4 | 0 | 1 (15) | |
| ⑥ | Northern Goshawk | Accipiter gentilis | R, P | 12 | 0 | 1 (10) | |
| ⑥ | Eastern Imperial Eagle | Aquila heliaca | zoo bird | 1 | 0 | 1 (15) | |
| ⑥ | Common Buzzard | Buteo buteo | R, P, S | 25 | 0 | 1 (15), 1 (20) | |
| ⑥ | Western Marsh Harrier | Circus aeruginosus | L | 6 | 1 (40), 1 (20) | 1 (10) | |
| ⑥ | Southern Crested Caracara | Caracara plancus | zoo bird | 1 (10) | 0 | ||
| ⑥ | Rüppell′s Vulture | Gyps rueppelli | zoo bird | 1 | 1 (15) | 1 (50) | |
| ⑥ | Golden Eagle | Aquila chrysaetos | zoo bird | 4 | 0 | 1 (60) | |
| ⑥ | White-backed Vulture | Gyps africanus | zoo bird | 1 | 1 (20) | 0 | |
| ⑥ | Eurasian Hobby | Falco subbuteo | L | 2 | 1 (30) | 0 | |
| ⑥ | European Kestrel | Falco tinnunculus | R, P, S | 14 | 1 (20) | 0 | |
| ⑥ | Strigiformes | Northern Long-eared Owl | Asio otus | R, P, S | 4 | 0 | 1 (30) |
| ⑥ | Columbiformes | Common Wood Pigeon | Columba palumbus | R, P, S | 21 | 1 (40) | 1 (80) |
| ⑥ | Feral Pigeon | Columba livia f. domestica | R, (P) | 13 | 0 | 2 (10), 1 (15) | |
| ⑥ | Anseriformes | Mute Swan | Cygnus olor | R, P, S | 9 | 1 (10) | 1 (15), 1 (40) |
| ⑥ | Egyptian Goose | Alopochen aegyptiacus | R (Neozoa) | 2 | 0 | 1 (10) | |
| ⑥ | Ciconiiformes/Pelicaniformes | Grey Heron | Ardea cinerea | R, P, S | 8 | 0 | 1 (10), 1 (40) |
| ⑥ | Cuculiformes | Common Cuckoo | Cuculus canorus | L | 1 | 1 (20) | 0 |
| ⑥ | Piciformes | Great Spottet Woodpecker | Dendrocopos major | R, P, (S) | 6 | 1 (10) | 0 |
| ⑦ | Columbiformes | Feral Pigeon | Columba livia f. domestica | R, (P) | 16 | 0 | 1 (30) |
| ⑧ | Accipitriformes | Common Buzzard | Buteo buteo | R, P, S | 3 | 0 | 1 (15) |
| ⑧ | Falconiformes | European Kestrel | Falco tinnunculus | R, P, S | 1 | 0 | 1 (30) |
| Total | 398 | 13 | 26 | ||||
| Seroprevalence | 3.27% | 6.53% | |||||
| Sample Collector | Year | Common Name | Scientific Name | WNV pos. (ND50) | USUV pos. (ND50) |
|---|---|---|---|---|---|
| ② | 2017 | Common Wood Pigeon | Columba palumbus | 1 (10) | 1 (15) |
| ② | 2017 | Common Wood Pigeon | Columba palumbus | 1 (10) | 1 (15) |
| ⑤ | 2017 | Eurasian Blackbird | Turdus merula | 1 (15) | 1 (20) |
| ⑤ | 2017 | Eurasian Blackbird | Turdus merula | 1 (160) | 1 (240) |
| ⑥ | 2017 | Northern Goshawk | Accipiter gentilis | 1 (15) | 1 (10) |
| ⑥ | 2017 | European Kestrel | Falco tinnunculus | 1 (15) | 1 (15) |
| ⑥ | 2017 | Snowy owl | Bubo scandiacus | 1 (10) | 1 (10) |
| ① | 2018 | Eurasian Blackbird | Turdus merula | 1 (30) | 1 (40) |
| ⑥ | 2018 | Eurasian Blackbird | Turdus merula | 1 (30) | 1 (30) |
| ⑥ | 2018 | Eurasian Blackbird | Turdus merula | 1 (10) | 1 (10) |
| ⑥ | 2018 | Eurasian Blackbird | Turdus merula | 1 (20) | 1, (15) |
| ⑥ | 2018 | House Sparrow | Passer domesticus | 1 (60) | 1 (80) |
| ⑥ | 2018 | Great Grey Owl | Strix nebulosa | 1 (30) | 1 (20) |
| ⑥ | 2018 | Stork sp. | Circonia sp. | 1 (10) | 1 (15) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Michel, F.; Sieg, M.; Fischer, D.; Keller, M.; Eiden, M.; Reuschel, M.; Schmidt, V.; Schwehn, R.; Rinder, M.; Urbaniak, S.; et al. Evidence for West Nile Virus and Usutu Virus Infections in Wild and Resident Birds in Germany, 2017 and 2018. Viruses 2019, 11, 674. https://doi.org/10.3390/v11070674
Michel F, Sieg M, Fischer D, Keller M, Eiden M, Reuschel M, Schmidt V, Schwehn R, Rinder M, Urbaniak S, et al. Evidence for West Nile Virus and Usutu Virus Infections in Wild and Resident Birds in Germany, 2017 and 2018. Viruses. 2019; 11(7):674. https://doi.org/10.3390/v11070674
Chicago/Turabian StyleMichel, Friederike, Michael Sieg, Dominik Fischer, Markus Keller, Martin Eiden, Maximilian Reuschel, Volker Schmidt, Rebekka Schwehn, Monika Rinder, Sylvia Urbaniak, and et al. 2019. "Evidence for West Nile Virus and Usutu Virus Infections in Wild and Resident Birds in Germany, 2017 and 2018" Viruses 11, no. 7: 674. https://doi.org/10.3390/v11070674
APA StyleMichel, F., Sieg, M., Fischer, D., Keller, M., Eiden, M., Reuschel, M., Schmidt, V., Schwehn, R., Rinder, M., Urbaniak, S., Müller, K., Schmoock, M., Lühken, R., Wysocki, P., Fast, C., Lierz, M., Korbel, R., Vahlenkamp, T. W., Groschup, M. H., & Ziegler, U. (2019). Evidence for West Nile Virus and Usutu Virus Infections in Wild and Resident Birds in Germany, 2017 and 2018. Viruses, 11(7), 674. https://doi.org/10.3390/v11070674