Catch Me If You Can! RNA Silencing-Based Improvement of Antiviral Plant Immunity


Abstract
1. Introduction
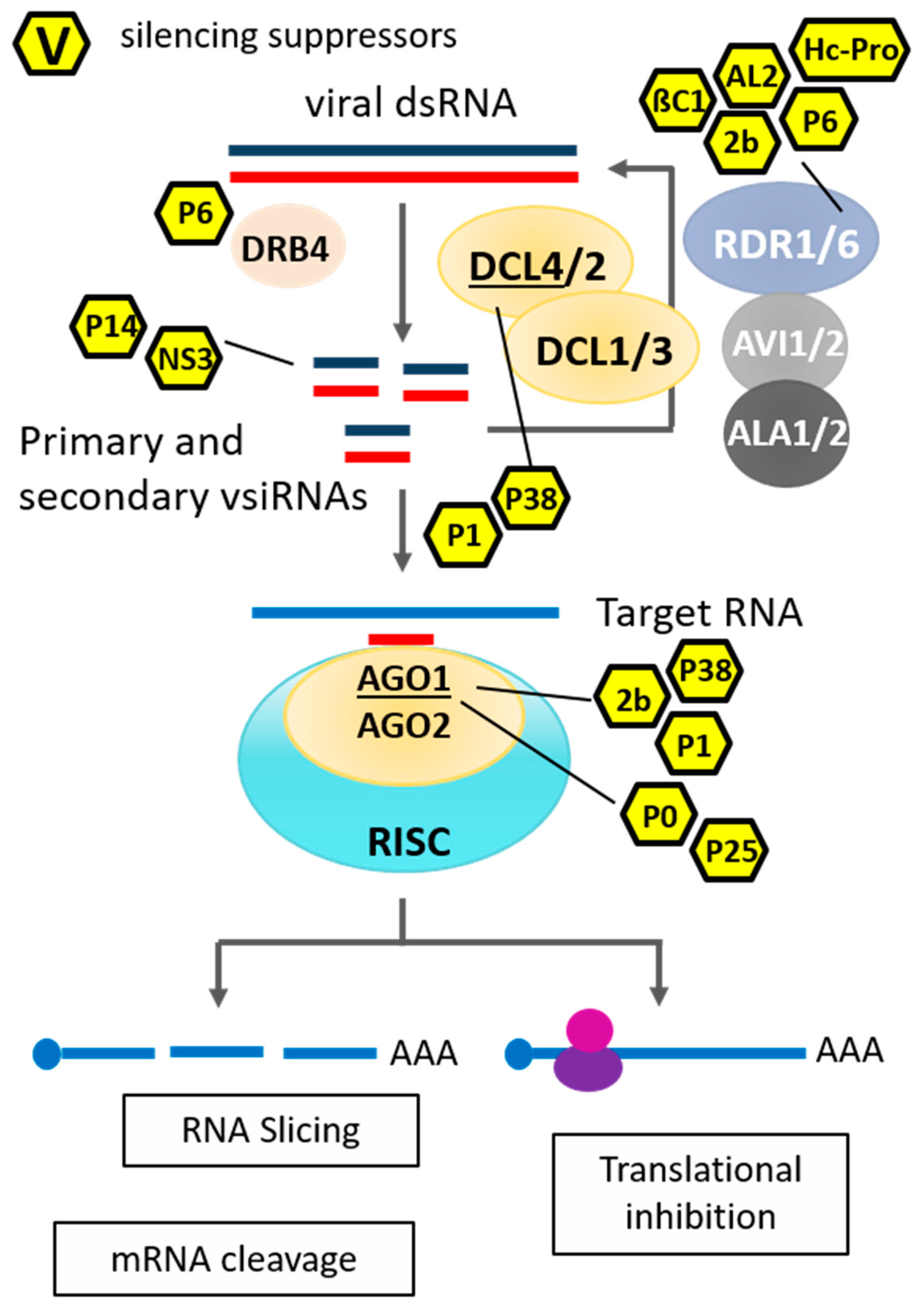
2. RNA Silencing-Mediated Antiviral Plant Immunity
3. Improvement of Plant Immunity Using RNA Silencing-Based Plant Protection Strategies
4. RNA Silencing-Based Crop Protection Against Viruses
5. RNA Silencing-Mediated Viral Resistance in Solanaceae—Tobacco, Tomato and Potato
6. RNA-Silencing-Mediated Viral Resistance in Cucurbitaceae—Cucumber, Melon and Watermelon
7. RNA-Silencing-Mediated Viral Resistance in Fabaceae—Soybean
8. RNA-Silencing-Mediated Viral Resistance in Poaceae—Rice
9. RNA-Silencing-Mediated Viral Resistance in Poaceae—Wheat and Barley
10. RNA-Silencing-Mediated VIRAL Resistance in Euphorbiacea—Cassava
11. RNA-Silencing-Mediated Viral Resistance in Fruits—Citrus
12. Outlook: Non-GMO RNA Spray Confer Virus Resistance
Acknowledgments
Conflicts of Interest
References
- Hull, R. Matthews’ Plant Virology, 4th ed.; Academic Press: San Diego, CA, USA, 2002. [Google Scholar]
- Dall’Ara, M.; Ratti, C.; Bouzoubaa, S.E.; Gilmer, D. Ins and Outs of Multipartite Positive-Strand RNA Plant Viruses: Packaging versus Systemic Spread. Viruses 2016, 8, 228. [Google Scholar] [CrossRef] [PubMed]
- Hull, R. Plant Virology, 5th ed.; Academic Press: Amsterdam, The Netherlands, 2013; p. 1118. [Google Scholar]
- Andika, I.B.; Kondo, H.; Sun, L. Interplays between Soil-Borne Plant Viruses and RNA Silencing-Mediated Antiviral Defense in Roots. Front. Microbiol. 2016, 7, 1458. [Google Scholar] [CrossRef] [PubMed]
- Simmons, H.E.; Holmes, E.C.; Gildow, F.E.; Bothe-Goralczyk, M.A.; Stephenson, A.G. Experimental Verification of Seed Transmission of zucchini yellow mosaic virus. Plant Dis. 2011, 95, 751–754. [Google Scholar] [CrossRef] [PubMed]
- Simmons, H.E.; Dunham, J.P.; Zinn, K.E.; Munkvold, G.P.; Holmes, E.C.; Stephenson, A.G. Zucchini yellow mosaic virus (ZYMV, Potyvirus): Vertical transmission, seed infection and cryptic infections. Virus Res. 2013, 176, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Pitzalis, N.; Heinlein, M. The roles of membranes and associated cytoskeleton in plant virus replication and cell-to-cell movement. J. Exp. Bot. 2017, 18, 117–132. [Google Scholar] [CrossRef]
- Zhou, J.S.; Drucker, M.; Ng, J.C. Direct and indirect influences of virus-insect vector-plant interactions on non-circulative, semi-persistent virus transmission. Curr. Opin. Virol. 2018, 33, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Gallet, R.; Michalakis, Y.; Blanc, S. Vector-transmission of plant viruses and constraints imposed by virus-vector interactions. Curr. Opin. Virol. 2018, 33, 144–150. [Google Scholar] [CrossRef]
- Gouveia, B.C.; Calil, I.P.; Machado, J.P.; Santos, A.A.; Fontes, E.P. Immune Receptors and Co-receptors in Antiviral Innate Immunity in Plants. Front. Microbiol. 2017, 5, 2139. [Google Scholar] [CrossRef]
- Carr, J.P.; Murphy, A.M.; Tungadi, T.; Yoon, J.-Y. Plant defense signals: Players and pawns in plant-virus-vector interactions. Plant Sci. 2019, 279, 87–95. [Google Scholar] [CrossRef]
- Moon, J.Y.; Park, J.M. Cross-Talk in Viral Defense Signaling in Plants. Front. Microbiol. 2016, 7, 904. [Google Scholar] [CrossRef]
- Wu, X.; Valli, A.; García, J.A.; Zhou, X.; Cheng, X. The Tug-of-War between Plants and Viruses: Great Progress and Many Remaining Questions. Viruses 2019, 11, 203. [Google Scholar] [CrossRef] [PubMed]
- Garcia, D.; Garcia, S.; Voinnet, O. Nonsense-Mediated Decay Serves as a General Viral Restriction Mechanism in Plants. Cell Host Microbe 2014, 16, 391–402. [Google Scholar] [CrossRef] [PubMed]
- Szádeczky-Kardoss, I.; Gál, L.; Auber, A.; Taller, J.; Silhavy, D. The No-go decay system degrades plant mRNAs that contain a long A-stretch in the coding region. Plant Sci. 2018, 275, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Szádeczky-Kardoss, I.; Csorba, T.; Auber, A.; Schamberger, A.; Nyikó, T.; Taller, J.; Orbán, T.I.; Burgyán, J.; Silhavy, D. The nonstop decay and the RNA silencing systems operate cooperatively in plants. Nucleic Acids Res. 2018, 46, 4632–4648. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 16, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Zipfel, C. Early molecular events in PAMP-triggered immunity. Curr. Opin. Plant Boil. 2009, 12, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Zipfel, C. Plant pattern-recognition receptors. Trends Immunol. 2014, 35, 345–351. [Google Scholar] [CrossRef]
- Tang, D.; Wang, G.; Zhou, J.-M. Receptor Kinases in Plant-Pathogen Interactions: More Than Pattern Recognition. Plant Cell 2017, 29, 618–637. [Google Scholar] [CrossRef]
- Weiberg, A.; Wang, M.; Lin, F.M.; Zhao, H.; Zhang, Z.; Kaloshian, I.; Huang, H.D.; Jin, H. Fungal small RNAs suppress plant immunity by hijacking host RNA interference pathways. Science 2013, 4, 118–123. [Google Scholar] [CrossRef]
- Weiberg, A.; Bellinger, M.; Jin, H. Conversations between kingdoms: Small RNAs. Curr. Opin. Biotechnol. 2015, 32, 207–215. [Google Scholar] [CrossRef]
- Weiberg, A.; Jin, H. Small RNAs--the secret agents in the plant-pathogen interactions. Curr. Opin. Plant Biol. 2015, 26, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Tsuda, K.; Parker, J.E. Effector-triggered immunity: From pathogen perception to robust defense. Annu. Rev. Plant Biol. 2015, 66, 487–511. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Kapos, P.; Zhang, Y. NLRs in plants. Curr. Opin. Immunol. 2015, 32, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Kapos, P.; Devendrakumar, K.T.; Li, X. Plant NLRs: From discovery to application. Plant Sci. 2019, 279, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Allan, A.C.; Lapidot, M.; Culver, J.N.; Fluhr, R. An early tobacco mosaic virus-induced oxidative burst in tobacco indicates extracellular perception of the virus coat protein. Plant Physiol. 2001, 126, 97–108. [Google Scholar] [CrossRef]
- Yang, H.; Gou, X.P.; He, K.; Xi, D.H.; Du, J.B.; Lin, H.H.; Li, J. BAK1 and BKK1 in Arabidopsis thaliana confer reduced susceptibility to turnip crinkle virus. Eur. J. Plant Pathol. 2010, 127, 149–156. [Google Scholar] [CrossRef]
- Kørner, C.J.; Klauser, D.; Niehl, A.; Domínguez-Ferreras, A.; Chinchilla, D.; Boller, T.; Heinlein, M.; Hann, D.R. The immunity regulator BAK1 contributes to resistance against diverse RNA viruses. Mol. Plant Microbe Interact. 2013, 26, 1271–1280. [Google Scholar] [CrossRef]
- Niehl, A.; Wyrsch, I.; Boller, T.; Heinlein, M. Double-stranded RNAs induce a pattern-triggered immune signaling pathway in plants. N. Phytol. 2016, 211, 1008–1019. [Google Scholar] [CrossRef]
- Perraki, A.; Gronnier, J.; Gouguet, P.; Boudsocq, M.; Deroubaix, A.F.; Simon, V.; German-Retana, S.; Legrand, A.; Habenstein, B.; Zipfel, C.; et al. REM1.3’s phospho-status defines its plasma membrane nanodomain organization and activity in restricting PVX cell-to-cell movement. PLoS Pathog. 2018, 12, e1007378. [Google Scholar] [CrossRef]
- Calil, I.P.; Fontes, E.P.B. Plant immunity against viruses: Antiviral immune receptors in focus. Ann. Bot. 2017, 119, 711–723. [Google Scholar] [CrossRef]
- Ramesh, S.V.; Sahu, P.P.; Prasad, M.; Praveen, S.; Pappu, H.R. Geminiviruses and Plant Hosts: A Closer Examination of the Molecular Arms Race. Viruses 2017, 9, 256. [Google Scholar] [CrossRef] [PubMed]
- Vaucheret, H.; Béclin, C.; Fagard, M. Post-transcriptional gene silencing in plants. J. Cell Sci. 2001, 114 Pt 17, 3083–3091. [Google Scholar] [PubMed]
- Vaucheret, H.; Fagard, M. Transcriptional gene silencing in plants: Targets, inducers and regulators. Trends Genet. 2001, 17, 29–35. [Google Scholar] [CrossRef]
- Castel, S.E.; Martienssen, R.A. RNA interference in the nucleus: Roles for small RNAs in transcription, epigenetics and beyond. Nat. Rev. Genet. 2013, 14, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.L.; Chekanova, J.A. Small RNAs: Essential regulators of gene expression and defenses against environmental stresses in plants. Wiley Interdiscip. Rev. RNA 2016, 7, 356–381. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Ferrer, V.; Voinnet, O. Roles of plant small RNAs in biotic stress responses. Annu. Rev. Plant Biol. 2009, 60, 485–510. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, A.J.; Baulcombe, D.C. A species of small antisense RNA in posttranscriptional gene silencing in plants. Science 1999, 286, 950–952. [Google Scholar] [CrossRef] [PubMed]
- Baulcombe, D. RNA silencing in plants. Nature 2004, 16, 356–363. [Google Scholar] [CrossRef]
- Ketting, R.F. The many faces of RNAi. Dev. Cell 2011, 20, 148–161. [Google Scholar] [CrossRef]
- Vaucheret, H.; Vazquez, F.; Crété, P.; Bartel, D.P. The action of ARGONAUTE1 in the miRNA pathway and its regulation by the miRNA pathway are crucial for plant development. Genes Dev. 2004, 18, 1187–1197. [Google Scholar] [CrossRef]
- Brodersen, P.; Voinnet, O. The diversity of RNA silencing pathways in plants. Trends Genet. 2006, 22, 268–280. [Google Scholar] [CrossRef]
- Ghildiyal, M.; Zamore, P.D. Small silencing RNAs: An expanding universe. Nat. Rev. Genet. 2009, 10, 94–108. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Paroo, Z. Biochemical principles of small RNA pathways. Annu. Rev. Biochem. 2010, 79, 295–319. [Google Scholar] [CrossRef] [PubMed]
- Vaucheret, H. Post-transcriptional small RNA pathways in plants: Mechanisms and regulations. Genes Dev. 2006, 20, 759–771. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.W.; Voinnet, O. Antiviral immunity directed by small RNAs. Cell 2007, 130, 413–426. [Google Scholar] [CrossRef] [PubMed]
- Burgess, D.J. Small RNAs: Antiviral RNAi in mammals. Nat. Rev. Genet. 2013, 14, 821. [Google Scholar] [CrossRef] [PubMed]
- Cullen, B.R.; Cherry, S.; Tenoever, B.R. Is RNA interference a physiologically relevant innate antiviral immune response in mammals? Cell Host Microbe 2013, 14, 374–378. [Google Scholar] [CrossRef]
- Li, Y.; Lu, J.; Han, Y.; Fan, X.; Ding, S.W. RNA interference functions as an antiviral immunity mechanism in mammals. Science 2013, 342, 231–234. [Google Scholar] [CrossRef]
- Maillard, P.V.; Ciaudo, C.; Marchais, A.; Li, Y.; Jay, F.; Ding, S.W.; Voinnet, O. Antiviral RNA interference in mammalian cells. Science 2013, 342, 235–238. [Google Scholar] [CrossRef]
- Parent, J.S.; Bouteiller, N.; Elmayan, T.; Vaucheret, H. Respective contributions of Arabidopsis DCL2 and DCL4 to RNA silencing. Plant J. 2015, 81, 223–232. [Google Scholar] [CrossRef]
- Zhang, C.; Wu, Z.; Li, Y.; Wu, J. Biogenesis, Function, and Applications of Virus-Derived Small RNAs in Plants. Front. Microbiol. 2015, 9, 1237. [Google Scholar] [CrossRef] [PubMed]
- Deleris, A.; Gallego-Bartolome, J.; Bao, J.; Kasschau, K.D.; Carrington, J.C.; Voinnet, O. Hierarchical action and inhibition of plant Dicer-like proteins in antiviral defense. Science 2006, 313, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Qu, F.; Ye, X.; Morris, T.J. Arabidopsis DRB4, AGO1, AGO7, and RDR6 participate in a DCL4-initiated antiviral RNA silencing pathway negatively regulated by DCL1. Proc. Natl. Acad. Sci. USA 2008, 105, 14732–14737. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Ruiz, H.; Takeda, A.; Chapman, E.J.; Sullivan, C.M.; Fahlgren, N.; Brempelis, K.J.; Carrington, J.C. Arabidopsis RNA-dependent RNA polymerases and dicer-like proteins in antiviral defense and small interfering RNA biogenesis during Turnip Mosaic Virus infection. Plant Cell 2010, 22, 481–496. [Google Scholar] [CrossRef] [PubMed]
- Csorba, T.; Kontra, L.; Burgyán, J. viral silencing suppressors: Tools forged to fine-tune host-pathogen coexistence. Virology 2015, 479–480, 85–103. [Google Scholar] [CrossRef]
- Carbonell, A.; Carrington, J.C. Antiviral roles of plant Argonautes. Curr. Opin. Plant Biol. 2015, 27, 111–117. [Google Scholar] [CrossRef]
- Wang, X.B.; Wu, Q.; Ito, T.; Cillo, F.; Li, W.X.; Chen, X.; Yu, J.L.; Ding, S.W. RNAi-mediated viral immunity requires amplification of virus-derived siRNAs in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2010, 107, 484–489. [Google Scholar] [CrossRef]
- Garcia, D.; Garcia, S.; Pontier, D.; Marchais, A.; Renou, J.P.; Lagrange, T.; Voinnet, O. Ago hook and RNA helicase motifs underpin dual roles for SDE3 in antiviral defense and silencing of nonconserved intergenic regions. Mol. Cell 2012, 48, 109–120. [Google Scholar] [CrossRef]
- Taochy, C.; Gursanscky, N.R.; Cao, J.; Fletcher, S.J.; Dressel, U.; Mitter, N.; Tucker, M.R.; Koltunow, A.M.G.; Bowman, J.L.; Vaucheret, H.; et al. A Genetic Screen for Impaired Systemic RNAi Highlights the Crucial Role of DICER-LIKE 2. Plant Physiol. 2017, 175, 1424–1437. [Google Scholar] [CrossRef]
- Mourrain, P.; Beclin, C.; Elmayan, T.; Feuerbach, F.; Godon, C.; Morel, J.B.; Jouette, D.; Lacombe, A.M.; Nikic, S.; Picault, N.; et al. Arabidopsis SGS2 and SGS3 genes are required for posttranscriptional gene silencing and natural virus resistance. Cell 2000, 101, 533–542. [Google Scholar] [CrossRef]
- Melnyk, C.W.; Molnar, A.; Baulcombe, D.C. Intercellular and systemic movement of RNA silencing signals. EMBO J. 2011, 30, 3553–3563. [Google Scholar] [CrossRef] [PubMed]
- Brosnan, C.A.; Mitter, N.; Christie, M.; Smith, N.A.; Waterhouse, P.M.; Carroll, B.J. Nuclear gene silencing directs reception of long-distance mRNA silencing in Arabidopsis. Proc. Natl. Acad. Sci. USA 2007, 104, 14741–14746. [Google Scholar] [CrossRef] [PubMed]
- Yu, A.; Lepère, G.; Jay, F.; Wang, J.; Bapaume, L.; Wang, Y.; Abraham, A.L.; Penterman, J.; Fischer, R.L.; Voinnet, O.; et al. Dynamics and biological relevance of DNA demethylation in Arabidopsis antibacterial defense. Proc. Natl. Acad. Sci. USA 2013, 5, 2389–2394. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, A.; Stassen, J.H.M.; Furci, L.; Smith, L.M.; Ton, J. The role of DNA (de)methylation in immune responsiveness of Arabidopsis. Plant J. 2016, 88, 361–374. [Google Scholar] [CrossRef] [PubMed]
- Burgyán, J.; Havelda, Z. Viral suppressors of RNA silencing. Trends Plant Sci. 2011, 16, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Incarbone, M.; Zimmermann, A.; Hammann, P.; Erhardt, M.; Michel, F.; Dunoyer, P. Neutralization of mobile antiviral small RNA through peroxisomal import. Nat. Plants 2017, 3, 17094. [Google Scholar] [CrossRef] [PubMed]
- Murray, G.G.; Kosakovsky, P.S.L.; Obbard, D.J. Suppressors of RNAi from plant viruses are subject to episodic positive selection. Proc. Biol. Sci. 2013, 280, 20130965. [Google Scholar] [CrossRef]
- Daròs, J.A. Viral suppressors: Combatting RNA silencing. Nat. Plants 2017, 3, 17098. [Google Scholar] [CrossRef]
- Ghoshal, B.; Sanfaçon, H. Symptom recovery in virus-infected plants: Revisiting the role of RNA silencing mechanisms. Virology 2015, 479–480, 167–179. [Google Scholar] [CrossRef]
- Zhao, J.H.; Hua, C.L.; Fang, Y.Y.; Guo, H.S. The dual edge of RNA silencing suppressors in the virus-host interactions. Curr. Opin. Virol. 2016, 17, 39–44. [Google Scholar] [CrossRef]
- Yang, Z.; Li, Y. Dissection of RNAi-based antiviral immunity in plants. Curr. Opin. Virol. 2018, 32, 88–99. [Google Scholar] [CrossRef] [PubMed]
- Sidahmed, A.M.; Wilkie, B. Endogenous antiviral mechanisms of RNA interference: A comparative biology perspective. Methods Mol. Biol. 2010, 623, 3–19. [Google Scholar] [CrossRef]
- Voinnet, O. Induction and suppression of RNA silencing: Insights from viral infections. Nat. Rev. Genet. 2005, 6, 206–220. [Google Scholar] [CrossRef] [PubMed]
- Price, D.R.; Gatehouse, J.A. RNAi-mediated crop protection against insects. Trends Biotechnol. 2008, 26, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, H.C.; Miao, X.X. Feasibility, limitation and possible solutions of RNAi-based technology for insect pest control. Insect Sci. 2013, 20, 15–30. [Google Scholar] [CrossRef]
- Lilley, C.J.; Davies, L.J.; Urwin, P.E. RNA interference in plant parasitic nematodes: A summary of the current status. Parasitology 2012, 139, 630–640. [Google Scholar] [CrossRef]
- Nunes, C.C.; Dean, R.A. Host-induced gene silencing: A tool for understanding fungal host interaction and for developing novel disease control strategies. Mol. Plant Pathol. 2012, 13, 519–529. [Google Scholar] [CrossRef]
- Koch, A.; Kogel, K.H. New wind in the sails: Improving the agronomic value of crop plants through RNAi-mediated gene silencing. Plant Biotechnol. J. 2014, 12, 821–831. [Google Scholar] [CrossRef]
- Nowara, D.; Gay, A.; Lacomme, C.; Shaw, J.; Ridout, C.; Douchkov, D.; Hensel, G.; Kumlehn, J.; Schweizer, P. HIGS: Host-induced gene silencing in the obligate biotrophic fungal pathogen Blumeria graminis. Plant Cell 2010, 22, 3130–3141. [Google Scholar] [CrossRef]
- Koch, A.; Kumar, N.; Weber, L.; Keller, H.; Imani, J.; Kogel, K.H. Host-induced gene silencing of cytochrome P450 lanosterol C14α-demethylase-encoding genes confers strong resistance to Fusarium species. Proc. Natl. Acad. Sci. USA 2013, 110, 19324–19329. [Google Scholar] [CrossRef]
- Qi, T.; Guo, J.; Peng, H.; Liu, P.; Kang, Z.; Guo, J. Host-Induced Gene Silencing: A Powerful Strategy to Control Diseases of Wheat and Barley. Int. J. Mol. Sci. 2019, 20, 206. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Negrete, E.A.; Carrillo-Tripp, J.; Rivera-Bustamante, R.F. RNA silencing against geminivirus: Complementary action of posttranscriptional gene silencing and transcriptional gene silencing in host recovery. J. Virol. 2009, 83, 1332–1340. [Google Scholar] [CrossRef]
- Aregger, M.; Borah, B.K.; Seguin, J.; Rajeswaran, R.; Gubaeva, E.G.; Zvereva, A.S.; Windels, D.; Vazquez, F.; Blevins, T.; Farinelli, L.; et al. Primary and secondary siRNAs in geminivirus-induced gene silencing. PLoS Pathog. 2012, 8, e1002941. [Google Scholar] [CrossRef] [PubMed]
- Broadbent, L. The epidemiology of tomato mosaic VII. The effect of TMV on tomato fruit yield and quality under glass. Ann. Appl. Biol. 1964, 54, 209–224. [Google Scholar] [CrossRef]
- Abel, P.P.; Nelson, R.S.; De, B.; Hoffmann, N.; Rogers, S.G.; Fraley, R.T.; Beachy, R.N. Delay of disease development in transgenic plants that express the tobacco mosaic virus coat protein gene. Science 1986, 232, 738–743. [Google Scholar] [CrossRef] [PubMed]
- Day, A.G.; Bejarano, E.R.; Buck, K.W.; Burrell, M.; Lichtenstein, C.P. Expression of an antisense viral gene in transgenic tobacco confers resistance to the DNA virus tomato golden mosaic virus. Proc. Natl. Acad. Sci. USA 1991, 88, 6721–6725. [Google Scholar] [CrossRef] [PubMed]
- Arif, M.; Azhar, U.; Arshad, M.; Zafar, Y.; Mansoor, S.; Asad, S. Engineering broad-spectrum resistance against RNA viruses in potato. Transgenic Res. 2012, 21, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Hernández, A.M.; Gosalvez, B.; Sempere, R.N.; Burgos, L.; Aranda, M.A.; Truniger, V. Melon RNA interference (RNAi) lines silenced for Cm-eIF4E show broad virus resistance. Mol. Plant Pathol. 2012, 13, 755–763. [Google Scholar] [CrossRef]
- Jelly, N.S.; Schellenbaum, P.; Walter, B.; Maillot, P. Transient expression of artificial microRNAs targeting Grapevine fanleaf virus and evidence for RNA silencing in grapevine somatic embryos. Transgenic Res. 2012, 21, 1319–1327. [Google Scholar] [CrossRef]
- Shekhawat, U.K.; Ganapathi, T.R.; Hadapad, A.B. Transgenic banana plants expressing small interfering RNAs targeted against viral replication initiation gene display high-level resistance to banana bunchy top virus infection. J. Gen. Virol. 2012, 93 Pt 8, 1804–1813. [Google Scholar] [CrossRef]
- Shimizu, T.; Nakazono-Nagaoka, E.; Akita, F.; Wei, T.; Sasaya, T.; Omura, T.; Uehara-Ichiki, T. Hairpin RNA derived from the gene for Pns9, a viroplasm matrix protein of Rice gall dwarf virus, confers strong resistance to virus infection in transgenic rice plants. J. Biotechnol. 2012, 157, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Niu, Y.; Zhang, K.; Liu, Y.; Zhou, X. Virus-derived transgenes expressing hairpin RNA give immunity to Tobacco mosaic virus and Cucumber mosaic virus. Virol. J. 2011, 8, 41. [Google Scholar] [CrossRef] [PubMed]
- Kalantidis, K.; Psaradakis, S.; Tabler, M.; Tsagris, M. The occurrence of CMV-specific short Rnas in transgenic tobacco expressing virus-derived double-stranded RNA is indicative of resistance to the virus. Mol. Plant Microbe Interact. 2002, 15, 826–833. [Google Scholar] [CrossRef] [PubMed]
- Yan, P.Q.; Bai, X.Q.; Wan, X.Q.; Guo, Z.K.; Li, L.J.; Gong, H.Y.; Chu, C.C. Expression of TMV coat protein gene RNAi in transgenic tobacco plants confer immunity to tobacco mosaic virus infection. Yi Chuan 2007, 29, 1018–1022. [Google Scholar] [CrossRef] [PubMed]
- Qu, J.; Ye, J.; Fang, R. Artificial microRNA-mediated virus resistance in plants. J. Virol. 2007, 81, 6690–6699. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.K.; Lohuis, D.; Goldbach, R.; Prins, M. High frequency induction of RNA-mediated resistance against Cucumber mosaic virus using inverted repeat constructs. Mol. Breed. 2004, 14, 215–226. [Google Scholar] [CrossRef]
- Di Nicola-Negri, E.; Brunetti, A.; Tavazza, M.; Ilardi, V. Hairpin RNA-mediated silencing of Plum pox virus P1 and HC-Pro genes for efficient and predictable resistance to the virus. Transgenic Res. 2005, 14, 989–994. [Google Scholar] [CrossRef]
- Kamachi, S.; Mochizuki, A.; Nishiguchi, M.; Tabei, Y. Transgenic Nicotiana benthamiana plants resistant to cucumber green mottle mosaic virus based on RNA silencing. Plant Cell Rep. 2007, 26, 1283–1288. [Google Scholar] [CrossRef]
- Winterhagen, P.; Dubois, C.; Sinn, M.; Wetzel, T.; Reustle, G.M. Gene silencing and virus resistance based on defective interfering constructs in transgenic Nicotiana benthamiana is not linked to accumulation of siRNA. Plant Physiol. Biochem. 2009, 47, 739–742. [Google Scholar] [CrossRef] [PubMed]
- Chung, B.N.; Palukaitis, P. Resistance to multiple viruses in transgenic tobacco expressing fused, tandem repeat, virus-derived double-stranded RNAs. Virus Genes 2011, 43, 454–464. [Google Scholar] [CrossRef]
- Ai, T.; Zhang, L.; Gao, Z.; Zhu, C.X.; Guo, X. Highly efficient virus resistance mediated by artificial microRNAs that target the suppressor of PVX and PVY in plants. Plant Biol. 2011, 13, 304–316. [Google Scholar] [CrossRef] [PubMed]
- Wagaba, H.; Patil, B.L.; Mukasa, S.; Alicai, T.; Fauquet, C.M. Taylor NJ6. Artificial microRNA-derived resistance to Cassava brown streak disease. J. Virol. Methods 2016, 231, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Patil, B.L.; Ogwok, E.; Wagaba, H.; Mohammed, I.U.; Yadav, J.S.; Bagewadi, B.; Taylor, N.J.; Kreuze, J.F.; Maruthi, M.N.; Alicai, T.; et al. RNAi-mediated resistance to diverse isolates belonging to two virus species involved in Cassava brown streak disease. Mol. Plant Pathol. 2011, 12, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Patil, B.L.; Bagewadi, B.; Yadav, J.S.; Fauquet, C.M. Mapping and identification of cassava mosaic geminivirus DNA-A and DNA-B genome sequences for efficient siRNA expression and RNAi based virus resistance by transient agro-infiltration studies. Virus Res. 2016, 213, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Thu, L.T.M.; Thuy, V.T.X.; Duc, L.H.; Son, L.V.; Ha, C.H.; Mau, C.H. RNAi-mediated resistance to SMV and BYMV in transgenic tobacco. Crop Breed. Appl. Biot. 2016, 16, 213–218. [Google Scholar] [CrossRef]
- Reyes, C.A.; De Francesco, A.; Peña, E.J.; Costa, N.; Plata, M.I.; Sendin, L.; Castagnaro, A.P.; García, M.L. Resistance to Citrus psorosis virus in transgenic sweet orange plants is triggered by coat protein-RNA silencing. J. Biotechnol. 2011, 151, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Batuman, O.; Mawassi, M.; Bar-Joseph, M. Transgenes consisting of a dsRNA of an RNAi suppressor plus the 3′ UTR provide resistance to Citrus tristeza virus sequences in Nicotiana benthamiana but not in citrus. Virus Genes 2006, 33, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Bendahmane, M.; Gronenborn, B. Engineering resistance against tomato yellow leaf curl virus (TYLCV) using antisense RNA. Plant Mol. Biol. 1997, 33, 351–357. [Google Scholar] [CrossRef]
- Mubin, M.; Mansoor, S.; Hussain, M.; Zafar, Y. Silencing of the AV2 gene by antisense RNA protects transgenic plants against a bipartite begomovirus. Virol. J. 2007, 19, 10. [Google Scholar] [CrossRef]
- Asad, S.; Haris, W.A.; Bashir, A.; Zafar, Y.; Malik, K.A.; Malik, N.N.; Lichtenstein, C.P. Transgenic tobacco expressing geminiviral RNAs are resistant to the serious viral pathogen causing cotton leaf curl disease. Arch. Virol. 2003, 148, 2341–2352. [Google Scholar] [CrossRef]
- Nahid, N.; Amin, I.; Briddon, R.W.; Mansoor, S. RNA interference-based resistance against a legume mastrevirus. Virol. J. 2011, 8, 499. [Google Scholar] [CrossRef] [PubMed]
- Medina-Hernández, D.; Rivera-Bustamante, R.F.; Tenllado, F.; Holguín-Peña, R.J. Effects and effectiveness of two RNAi constructs for resistance to Pepper golden mosaic virus in Nicotiana benthamiana plants. Viruses 2013, 5, 2931–2945. [Google Scholar] [CrossRef] [PubMed]
- Schwind, N.; Zwiebel, M.; Itaya, A.; Ding, B.; Wang, M.B.; Krczal, G.; Wassenegger, M. RNAi-mediated resistance to Potato spindle tuber viroid in transgenic tomato expressing a viroid hairpin RNA construct. Mol. Plant Pathol. 2009, 10, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, H.; Zhang, J.; Zhang, C.; Gong, P.; Ziaf, K.; Xiao, F.; Ye, Z. Expression of artificial microRNAs in tomato confers efficient and stable virus resistance in a cell-autonomous manner. Transgenic Res. 2011, 20, 569–581. [Google Scholar] [CrossRef] [PubMed]
- Ntui, V.O.; Kong, K.; Azadi, P.; Khan, R.S.; Chin, D.P.; Igawa, T.; Mii, M.; Nakamura, I. RNAi-Mediated Resistance to Cucumber Mosaic Virus (CMV) in Genetically Engineered Tomato. AJPS 2014, 5, 554–572. [Google Scholar] [CrossRef]
- Fuentes, A.; Ramos, P.L.; Fiallo, E.; Callard, D.; Sánchez, Y.; Peral, R.; Rodríguez, R.; Pujol, M. Intron-hairpin RNA derived from replication associated protein C1 gene confers immunity to tomato yellow leaf curl virus infection in transgenic tomato plants. Transgenic Res. 2006, 15, 291–304. [Google Scholar] [CrossRef] [PubMed]
- Zrachya, A.; Kumar, P.P.; Ramakrishnan, U.; Levy, Y.; Loyter, A.; Arazi, T.; Lapidot, M.; Gafni, Y. Production of siRNA targeted against TYLCV coat protein transcripts leads to silencing of its expression and resistance to the virus. Transgenic Res. 2007, 16, 385–398. [Google Scholar] [CrossRef] [PubMed]
- Sano, T.; Nagayama, A.; Ogawa, T.; Ishida, I.; Okada, Y. Transgenic potato expressing a double-stranded RNA-specific ribonuclease is resistant to potato spindle tuber viroid. Nat. Biotechnol. 1997, 15, 1290–1294. [Google Scholar] [CrossRef] [PubMed]
- Kreuze, J.F.; Klein, I.S.; Lazaro, M.U.; Chuquiyuri, W.J.; Morgan, G.L.; Mejía, P.G.; Ghislain, M.; Valkonen, J.P. RNA silencing-mediated resistance to a crinivirus (Closteroviridae) in cultivated sweet potato (Ipomoea batatas L.) and development of sweet potato virus disease following co-infection with a potyvirus. Mol. Plant Pathol. 2008, 9, 589–598. [Google Scholar] [CrossRef] [PubMed]
- Hameed, A.; Tahir, M.N.; Asad, S.; Bilal, R.; Van Eck, J.; Jander, G.; Mansoor, S. RNAi-Mediated Simultaneous Resistance Against Three RNA Viruses in Potato. Mol. Biotechnol. 2017, 59, 73–83. [Google Scholar] [CrossRef]
- Missiou, A.; Kalantidis, K.; Boutla, A.; Tzortzakaki, S.; Tabler, M.; Tsagris, M. Generation of transgenic potato plants highly resistant to potato virus Y (PVY) through RNA silencing. Mol. Breed. 2004, 14, 185–197. [Google Scholar] [CrossRef]
- Fang, G.; Grumet, R. Genetic engineering of potyvirus resistance using constructs derived from the zucchini yellow mosaic virus coat protein gene. Mol. Plant Microbe Interact. 1993, 6, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Krubphachaya, P.; Jurícek, M.; Kertbundit, S. Induction of RNA-mediated resistance to papaya ringspot virus type W. J. Biochem. Mol. Biol. 2007, 31, 404–411. [Google Scholar] [CrossRef]
- Leibman, D.; Wolf, D.; Saharan, V.; Zelcer, A.; Arazi, T.; Yoel, S.; Gaba, V.; Gal-On, A. A high level of transgenic viral small RNA is associated with broad potyvirus resistance in cucurbits. Mol. Plant Microbe Interact. 2011, 24, 1220–1238. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.Y.; Ku, H.M.; Chiang, Y.H.; Ho, H.Y.; Yu, T.A.; Jan, F.J. Development of transgenic watermelon resistant to Cucumber mosaic virus and Watermelon mosaic virus by using a single chimeric transgene construct. Transgenic Res. 2012, 21, 983–993. [Google Scholar] [CrossRef] [PubMed]
- Tougou, M.; Furutani, N.; Yamagishi, N.; Shizukawa, Y.; Takahata, Y.; Hidaka, S. Development of resistant transgenic soybeans with inverted repeat-coat protein genes of soybean dwarf virus. Plant Cell Rep. 2006, 25, 1213–1218. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Sato, S.; Ye, X.; Dorrance, A.E.; Morris, T.J.; Clemente, T.E.; Qu, F. Robust RNAi-based resistance to mixed infection of three viruses in soybean plants expressing separate short hairpins from a single transgene. Phytopathology 2011, 101, 1264–1269. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Kim, M.J.; Pak, J.H.; Jung, H.W.; Choi, H.K.; Lee, Y.H.; Baek, I.Y.; Ko, J.M.; Jeong, S.C.; Pack, I.S.; et al. Characterization of SMV resistance of soybean produced by genetic transformation of SMV-CP gene in RNAi. Plant Biotechnol. Rep. 2013, 7, 425–433. [Google Scholar] [CrossRef]
- Kim, H.J.; Kim, M.J.; Pak, J.H.; Im, H.H.; Lee, D.H.; Kim, K.H.; Lee, J.H.; Kim, D.H.; Choi, H.K.; Jung, H.W.; et al. RNAi-mediated Soybean mosaic virus (SMV) resistance of a Korean Soybean cultivar. Plant Biotechnol. Rep. 2016, 10, 257. [Google Scholar] [CrossRef]
- Gao, L.; Ding, X.; Li, K.; Liao, W.; Zhong, Y.; Ren, R.; Liu, Z.; Adhimoolam, K.; Zhi, H. Characterization of Soybean mosaic virus resistance derived from inverted repeat-SMV-HC-Pro genes in multiple soybean cultivars. Theor. Appl. Genet. 2015, 128, 1489–1505. [Google Scholar] [CrossRef]
- Yang, X.; Niu, L.; Zhang, W.; Yang, J.; Xing, G.; He, H.; Guo, D.; Du, Q.; Qian, X.; Yao, Y.; et al. RNAi-mediated SMV P3 cistron silencing confers significantly enhanced resistance to multiple Potyvirus strains and isolates in transgenic soybean. Plant Cell Rep. 2018, 37, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Bonfim, K.; Faria, J.C.; Nogueira, E.O.; Mendes, E.A.; Aragão, F.J. RNAi-mediated resistance to Bean golden mosaic virus in genetically engineered common bean (Phaseolus vulgaris). Mol. Plant Microbe Interact. 2007, 20, 717–726. [Google Scholar] [CrossRef] [PubMed]
- Cruz, A.R.R.; Aragão, F.J.L. RNAi-based enhanced resistance to Cowpea severe mosaic virus and Cowpea aphid-borne mosaic virus in transgenic cowpea. Plant Pathol. 2014, 63, 831–837. [Google Scholar] [CrossRef]
- Ludlow, E.J.; Mouradov, A.; Spangenberg, G.C. Post-transcriptional gene silencing as an efficient tool for engineering resistance to white clover mosaic virus in white clover (Trifolium repens). J. Plant Physiol. 2009, 166, 1557–1567. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, H.; Rajasubramaniam, S.; Rajam, M.V.; Dasgupta, I. RNA-interference in rice against Rice tungro bacilliform virus results in its decreased accumulation in inoculated rice plants. Transgenic Res. 2008, 17, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Yoshii, M.; Wei, T.; Hirochika, H.; Omura, T. Silencing by RNAi of the gene for Pns12, a viroplasm matrix protein of Rice dwarf virus, results in strong resistance of transgenic rice plants to the virus. Plant Biotechnol. 2009, 7, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Song, Y.; Wu, B.; Jiang, M.; Li, K.; Zhu, C.; Wen, F. Production of transgenic rice new germplasm with strong resistance against two isolations of Rice stripe virus by RNA interference. Transgenic Res. 2011, 20, 1367–1377. [Google Scholar] [CrossRef]
- Shimizu, T.; Nakazono-Nagaoka, E.; Uehara-Ichiki, T.; Sasaya, T.; Omura, T. Targeting specific genes for RNA interference is crucial to the development of strong resistance to Rice stripe virus. Plant Biotechnol. J. 2011, 9, 503–512. [Google Scholar] [CrossRef]
- Shimizu, T.; Nakazono-Nagaoka, E.; Akita, F.; Uehara-Ichiki, T.; Sasaya, T.; Omura, T. Immunity to Rice black streaked dwarf virus, a plant reovirus, can be achieved in rice plants by RNA silencing against the gene for the viroplasm component protein. Virus Res. 2011, 160, 400–403. [Google Scholar] [CrossRef]
- Shimizu, T.; Ogamino, T.; Hiraguri, A.; Nakazono-Nagaoka, E.; Uehara-Ichiki, T.; Nakajima, M.; Akutsu, K.; Omura, T.; Sasaya, T. Strong resistance against Rice grassy stunt virus is induced in transgenic rice plants expressing double-stranded RNA of the viral genes for nucleocapsid or movement proteins as targets for RNA interference. Phytopathology 2013, 103, 513–519. [Google Scholar] [CrossRef]
- Fahim, M.; Ayala-Navarrete, L.; Millar, A.A.; Larkin, P.J. Hairpin RNA derived from viral NIa gene confers immunity to wheat streak mosaic virus infection in transgenic wheat plants. Plant Biotechnol. J. 2010, 8, 821–834. [Google Scholar] [CrossRef] [PubMed]
- Fahim, M.; Millar, A.A.; Wood, C.C.; Larkin, P.J. Resistance to Wheat streak mosaic virus generated by expression of an artificial polycistronic microRNA in wheat. Plant Biotechnol. J. 2012, 10, 150–163. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Y.; Yang, L.; Zhou, S.F.; Wang, H.G.; Li, W.C.; Fu, F.L. Improvement of resistance to maize dwarf mosaic virus mediated by transgenic RNA interference. J. Biotechnol. 2011, 153, 181–187. [Google Scholar] [CrossRef]
- Wang, M.-B.; Abbott, D.C.; Waterhouse, P.M. A single copy of a virus-derived transgene encoding hairpin RNA gives immunity to Barley yellow dwarf virus. Mol. Plant Pathol. 2000, 1, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Vanderschuren, H.; Fütterer, J.; Gruissem, W. Resistance to cassava mosaic disease in transgenic cassava expressing antisense RNAs targeting virus replication genes. Plant Biotechnol. J. 2005, 3, 385–397. [Google Scholar] [CrossRef] [PubMed]
- Vanderschuren, H.; Akbergenov, R.; Pooggin, M.M.; Hohn, T.; Gruissem, W.; Zhang, P. Transgenic cassava resistance to African cassava mosaic virus is enhanced by viral DNA-A bidirectional promoter-derived siRNAs. Plant Mol. Biol. 2007, 64, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Vanderschuren, H.; Alder, A.; Zhang, P.; Gruissem, W. Dose-dependent RNAi-mediated geminivirus resistance in the tropical root crop cassava. Plant Mol. Biol. 2009, 70, 265–272. [Google Scholar] [CrossRef]
- Chauhan, R.D.; Beyene, G.; Kalyaeva, M.; Fauquet, C.M.; Taylor, N. Improvements in Agrobacterium-mediated transformation of cassava (Manihot esculenta Crantz) for large-scale production of transgenic plants. Plant Cell Tissue Organ Cult. 2015, 121, 591. [Google Scholar] [CrossRef]
- Wagaba, H.; Beyene, G.; Aleu, J.; Odipio, J.; Okao-Okuja, G.; Chauhan, R.D.; Munga, T.; Obiero, H.; Halsey, M.E.; Ilyas, M.; et al. Field Level RNAi-Mediated Resistance to Cassava Brown Streak Disease across Multiple Cropping Cycles and Diverse East African Agro-Ecological Locations. Front. Plant Sci. 2017, 7, 2060. [Google Scholar] [CrossRef]
- Vanderschuren, H.; Moreno, I.; Anjanappa, R.B.; Zainuddin, I.M.; Gruissem, W. Exploiting the combination of natural and genetically engineered resistance to cassava mosaic and cassava brown streak viruses impacting cassava production in Africa. PLoS ONE 2012, 7, e45277. [Google Scholar] [CrossRef]
- Yadav, J.S.; Ogwok, E.; Wagaba, H.; Patil, B.L.; Bagewadi, B.; Alicai, T.; Gaitan-Solis, E.; Taylor, N.J. Fauquet CM. RNAi-mediated resistance to Cassava brown streak Uganda virus in transgenic cassava. Mol. Plant Pathol. 2011, 12, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Ogwok, E.; Odipio, J.; Halsey, M.; Gaitán-Solís, E.; Bua, A.; Taylor, N.J.; Fauquet, C.M.; Alicai, T. Transgenic RNA interference (RNAi)-derived field resistance to cassava brown streak disease. Mol. Plant Pathol. 2012, 13, 1019–1031. [Google Scholar] [CrossRef] [PubMed]
- Odipio, J.; Ogwok, E.; Taylor, N.J.; Halsey, M.; Bua, A.; Fauquet, C.M.; Alicai, T. RNAi-derived field resistance to Cassava brown streak disease persists across the vegetative cropping cycle. GM Crops Food 2014, 5, 16–19. [Google Scholar] [CrossRef] [PubMed]
- Beyene, G.; Chauhan, R.D.; Ilyas, M.; Wagaba, H.; Fauquet, C.M.; Miano, D.; Alicai, T.; Taylor, N.J. A Virus-Derived Stacked RNAi Construct Confers Robust Resistance to Cassava Brown Streak Disease. Front. Plant Sci. 2017, 18, 2052. [Google Scholar] [CrossRef] [PubMed]
- Clarke, J.L.; Spetz, C.; Haugslien, S.; Xing, S.; Dees, M.W.; Moe, R.; Blystad, D.R. Agrobacterium tumefaciens-mediated transformation of poinsettia, Euphorbia pulcherrima, with virus-derived hairpin RNA constructs confers resistance to Poinsettia mosaic virus. Plant Cell Rep. 2008, 27, 1027–1038. [Google Scholar] [CrossRef][Green Version]
- López, C.; Cervera, M.; Fagoaga, C.; Moreno, P.; Navarro, L.; Flores, R.; Peña, L. Accumulation of transgene-derived siRNAs is not sufficient for RNAi-mediated protection against Citrus tristeza virus in transgenic Mexican lime. Mol. Plant Pathol. 2010, 11, 33–41. [Google Scholar] [CrossRef]
- Soler, N.; Plomer, M.; Fagoaga, C.; Moreno, P.; Navarro, L.; Flores, R.; Peña, L. Transformation of Mexican lime with an intron-hairpin construct expressing untranslatable versions of the genes coding for the three silencing suppressors of Citrus tristeza virus confers complete resistance to the virus. Plant Biotechnol. J. 2012, 10, 597–608. [Google Scholar] [CrossRef]
- Reyes, C.A.; Pena, E.J.; Zanek, M.C.; Sanchez, D.V.; Grau, O.; Garcia, M.L. Differential resistance to Citrus psorosis virus in transgenic Nicotiana benthamiana plants expressing hairpin RNA derived from the coat protein and 54K protein genes. Plant Cell Rep. 2009, 28, 1817–1825. [Google Scholar] [CrossRef]
- Niu, Q.W.; Lin, S.S.; Reyes, J.L.; Chen, K.C.; Wu, H.W.; Yeh, S.D.; Chua, N.H. Expression of artificial microRNAs in transgenic Arabidopsis thaliana confers virus resistance. Nat. Biotechnol. 2006, 24, 1420–1428. [Google Scholar] [CrossRef]
- Duan, C.G.; Wang, C.H.; Fang, R.X.; Guo, H.S. Artificial MicroRNAs highly accessible to targets confer efficient virus resistance in plants. J. Virol. 2008, 82, 11084–11095. [Google Scholar] [CrossRef]
- Mitiouchkina, T.Y.; Firsov, A.P.; Titova, S.M.; Pushin, A.S.; Shulga, O.A.; Dolgov, S.V. Different Approaches to Produce Transgenic Virus B Resistant Chrysanthemum. Agronomy 2018, 8, 28. [Google Scholar] [CrossRef]
- Pavli, O.I.; Panopoulos, N.J.; Goldbach, R.; Skaracis, G.N. BNYVV-derived dsRNA confers resistance to rhizomania disease of sugar beet as evidenced by a novel transgenic hairy root approach. Transgenic Res. 2010, 19, 915–922. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Guo, J.; Gao, S.; Lin, Q.; Wang, H.; Que, Y.; Xu, L. Transgenic Sugarcane Resistant to Sorghum mosaic virus Based on Coat Protein Gene Silencing by RNA Interference. BioMed Res. Int. 2015, 2015, 9. [Google Scholar] [CrossRef] [PubMed]
- Szittya, G.; Silhavy, D.; Molnár, A.; Havelda, Z.; Lovas, A.; Lakatos, L.; Bánfalvi, Z.; Burgyán, J. Low temperature inhibits RNA silencing-mediated defence by the control of siRNA generation. EMBO J. 2003, 22, 633–640. [Google Scholar] [CrossRef] [PubMed]
- Dalakouras, A.; Tzanopoulou, M.; Tsagris, M.; Wassenegger, M.; Kalantidis, K. Hairpin transcription does not necessarily lead to efficient triggering of the RNAi pathway. Transgenic Res. 2011, 20, 293–304. [Google Scholar] [CrossRef] [PubMed]
- Weising, K.; Schell, J.; Kahl, G. Foreign genes in plants: Transfer, structure, expression, and applications. Annu. Rev. Genet. 1988, 22, 421–477. [Google Scholar] [CrossRef]
- Han, Y.; Griffiths, A.; Li, H.; Grierson, D. The effect of endogenous mRNA levels on co-suppression in tomato. FEBS Lett. 2004, 563, 123–128. [Google Scholar] [CrossRef]
- Matsunaga, W.; Shimura, H.; Shirakawa, S.; Isoda, R.; Inukai, T.; Matsumura, T.; Masuta, C. Transcriptional silencing of 35S driven-transgene is differentially determined depending on promoter methylation heterogeneity at specific cytosines in both plus-and minus-sense strands. BMC Plant Biol. 2019, 19, 24. [Google Scholar] [CrossRef]
- Rajeevkumar, S.; Anunanthini, P.; Sathishkumar, R. Epigenetic silencing in transgenic plants. Front. Plant Sci. 2015, 6, 693. [Google Scholar] [CrossRef]
- Hosseinahli, N.; Aghapour, M.; Duijf, P.H.G.; Baradaran, B. Treating cancer with microRNA replacement therapy: A literature review. J. Cell Physiol. 2018, 233, 5574–5588. [Google Scholar] [CrossRef]
- Carbonell, A. Design and High-Throughput Generation of Artificial Small RNA Constructs for Plants. Methods Mol. Biol. 2019, 1932, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Chung, B.N.; Yoon, J.Y.; Palukaitis, P. Engineered resistance in potato against potato leafroll virus, potato virus A and potato virus Y. Virus Genes 2013, 47, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Lawson, C.; Kaniewski, W.; Haley, L.; Rozman, R.; Newell, C.; Sanders, P.; Tumer, N.E. Engineering resistance to mixed virus infection in a commercial potato cultivar: Resistance to potato virus X and potato virus Y in transgenic Russet Burbank. Biotechnology 1990, 8, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, A.J.; Brown, S.; Yuanhai, H.; Ishizuka, M.; Lowe, A.; Solis, A.A.; Grierson, D. A transgene with repeated DNA causes high frequency, post-transcriptional suppression of ACC-oxidase gene expression in tomato. Plant J. 1998, 15, 737–746. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, P.M.; Graham, M.W.; Wang, M.B. Virus resistance and gene silencing in plants can be induced by simultaneous expression of sense and antisense RNA. Proc. Natl. Acad. Sci. USA 1998, 95, 13959–13964. [Google Scholar] [CrossRef] [PubMed]
- Wesley, S.V.; Helliwell, C.A.; Smith, N.A.; Wang, M.B.; Rouse, D.T.; Liu, Q.; Gooding, P.S.; Singh, S.P.; Abbott, D.; Stoutjesdijk, P.A.; et al. Construct design for efficient, effective and high-throughput gene silencing in plants. Plant J. 2001, 27, 581–590. [Google Scholar] [CrossRef] [PubMed]
- Provvidenti, R. Diseases caused by viruses. In Compendium of Cucurbit Diseases; Zitter, T.A., Hopkins, D.L., Thomas, C.E., Eds.; APS Press: Minnesota, MN, USA, 1996; pp. 37–45. [Google Scholar]
- Gaba, V.; Zelcer, A.; Gal-On, A. cucurbit biotechnology—the importance of virus resistance. In Vitro Cell. Dev. Biol.-Plant 2004, 40, 346–358. [Google Scholar] [CrossRef]
- Truniger, V.; Aranda, M.A. Recessive resistance to plant viruses. Adv. Virus Res. 2009, 75, 119–159. [Google Scholar] [CrossRef]
- Jan, F.J.; Fagoaga, C.; Pang, S.Z.; Gonsalves, D. A minimum length of N gene sequence in transgenic plants is required for RNA-mediated tospovirus resistance. J. Gen. Virol. 2000, 81 Pt 1, 235–242. [Google Scholar] [CrossRef]
- Pang, S.-Z.; Jan, F.-J.; Tricoli, D.M.; Russell, P.F.; Carney, K.J.; Hu, J.S.; Fuchs, M.; Quemada, H.D.; Gonsalves, D. Resistance tosquash mosaic comovirus in transgenic squash plants expressing its coatprotein genes. Mol. Breed. 2000, 6, 87–93. [Google Scholar] [CrossRef]
- Wu, H.W.; Yu, T.A.; Raja, J.A.; Wang, H.C.; Yeh, S.D. Generation of transgenic oriental melon resistant to Zucchini yellow mosaic virus by an improved cotyledon-cutting method. Plant Cell Rep. 2009, 28, 1053–1064. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.W.; Yu, T.A.; Raja, J.A.J.; Christopher, S.J.; Wang, S.L.; Yeh, S.D. Double-Virus Resistance of Transgenic Oriental Melon Conferred by Untranslatable Chimeric Construct Carrying Partial Coat Protein Genes of Two Viruses. Plant Dis. 2010, 94, 1341–1347. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.A.; Chiang, C.H.; Wu, H.W.; Li, C.M.; Yang, C.F.; Chen, J.H.; Chen, Y.W.; Yeh, S.D. Generation of transgenic watermelon resistant to Zucchini yellow mosaic virus and Papaya ringspot virus type W. Plant Cell Rep. 2011, 30, 359–371. [Google Scholar] [CrossRef] [PubMed]
- Pang, S.Z.; Jan, F.J.; Gonsalves, D. Nontarget DNA sequences reduce the transgene length necessary for RNA-mediated tospovirus resistance in transgenic plants. Proc. Natl. Acad. Sci. USA 1997, 94, 8261–8266. [Google Scholar] [CrossRef] [PubMed]
- Jan, F.J.; Fagoaga, C.; Pang, S.Z.; Gonsalves, D. A single chimeric transgene derived from two distinct viruses confers multi-virus resistance in transgenic plants through homology-dependent gene silencing. J. Gen. Virol. 2000, 81 Pt 8, 2103–2109. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, J.D.; Rice, M.E.; Hill, J.H. Evaluation of management strategies for bean leaf beetles (Coleoptera: Chrysomelidae) and Bean pod mottle virus (Comoviridae) in soybean. J. Econ. Entomol. 2008, 101, 1211–1227. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Lin, J.; Zheng, G.; Zhang, M.; Zhi, H. Recombinant soybean mosaic virus is prevalent in Chinese soybean fields. Arch. Virol. 2014, 159, 1793–1796. [Google Scholar] [CrossRef]
- Jenner, C.E.; Wang, X.; Tomimura, K.; Ohshima, K.; Ponz, F.; Walsh, J.A. The dual role of the potyvirus P3 protein of Turnip mosaic virus as a symptom and avirulence determinant in brassicas. Mol. Plant Microbe Interact. 2003, 16, 777–784. [Google Scholar] [CrossRef]
- Suehiro, N.; Natsuaki, T.; Watanabe, T.; Okuda, S. An important determinant of the ability of Turnip mosaic virus to infect Brassica spp. and/or Raphanus sativus is in its P3 protein. J. Gen. Virol. 2004, 85 Pt 7, 2087–2098. [Google Scholar] [CrossRef]
- Wang, Y.; Khatabi, B.; Hajimorad, M.R. Amino acid substitution in P3 of Soybean mosaic virus to convert avirulence to virulence on Rsv4-genotype soybean is influenced by the genetic composition of P3. Mol. Plant Pathol. 2015, 16, 301–307. [Google Scholar] [CrossRef]
- Wang, X.; Eggenberger, A.L.; Nutter, F.W.; John, H. Pathogen derived transgenic resistance to soybean mosaic virus in soybean. Mol. Breed. 2001, 8, 119. [Google Scholar] [CrossRef]
- Furutani, N.; Hidaka, H.; Kosaka, Y.; Shizukawa, Y.; Kanematsu, S. Coat protein gene-mediated resistance to soybean mosaic virus in transgenic soybean. Breed. Sci. 2006, 56, 119–124. [Google Scholar] [CrossRef][Green Version]
- Furutani, N.; Yamagishi, N.; Hidaka, S.; Shizukawa, Y.; Kanematsu, S.; Kosaka, Y. Soybean mosaic virus resistance in transgenic soybean caused by post-transcriptional gene silencing. Breed. Sci. 2007, 57, 123–128. [Google Scholar] [CrossRef]
- Lim, H.S.; Ko, T.S.; Hobbs, H.A.; Lambert, K.N.; Yu, J.M.; Mccoppin, N.K.; Korban, S.S.; Hartman, G.L.; Domier, L.L. Soybean mosaic virus helper component-protease alters leaf morphology and reduces seed production in transgenic soybean plants. Phytopathology 2007, 97, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Tougou, M.; Yamagishi, N.; Furutani, N.; Shizukawa, Y.; Takahata, Y.; Hidaka, S. Soybean dwarf virus-resistant transgenic soybeans with the sense coat protein gene. Plant Cell Rep. 2007, 26, 1967–1975. [Google Scholar] [CrossRef] [PubMed]
- FAO/WHO. Evaluation of Allergenicity of Genetically Modified Foods; Report of a joint FAO/WHO expert consultation on allergenicity of foods derived from biotechnology; Food and Agriculture Organization of the United Nations: Rome, Italy, 2001. [Google Scholar]
- Zhang, X.; Zhao, P.; Wu, K.; Zhang, Y.; Peng, M.; Liu, Z. Compositional equivalency of RNAi-mediated virus-resistant transgenic soybean and its nontransgenic counterpart. J. Agric. Food Chem. 2014, 14, 4475–4479. [Google Scholar] [CrossRef]
- OECD (Organisation for Economic Cooperation and Development). Concepts and Principles; International Energy Agency Publications: Paris, France, 1993. [Google Scholar]
- Omura, T.; Yan, J. Role of outer capsid proteins in transmission of Phytoreovirus by insect vectors. Adv. Virus Res. 1999, 54, 15–43. [Google Scholar]
- Wei, T.; Kikuchi, A.; Moriyasu, Y.; Suzuki, N.; Shimizu, T.; Hagiwara, K.; Chen, H.; Takahashi, M.; Ichiki-Uehara, T.; Omura, T. The spread of Rice dwarf virus among cells of its insect vector exploits virus-induced tubular structures. J. Virol. 2006, 80, 8593–8602. [Google Scholar] [CrossRef]
- Wei, T.; Kikuchi, A.; Suzuki, N.; Shimizu, T.; Hagiwara, K.; Chen, H.; Omura, T. Pns4 of rice dwarf virus is a phosphoprotein, is localized around the viroplasm matrix, and forms minitubules. Arch. Virol. 2006, 151, 1701–1712. [Google Scholar] [CrossRef]
- Ellis, M.H.; Rebetzke, G.J.; Chu, P. First report of Wheat streak mosaic virus in Australia. Plant Pathol. 2003, 52, 808. [Google Scholar] [CrossRef]
- Ellis, M.H.; Rebetzke, G.J.; Kelman, W.M.; Moore, C.S.; Hyles, J.E. Detection of Wheat streak mosaic virus in four pasture grass species in Australia. Plant Pathol. 2004, 53, 239. [Google Scholar] [CrossRef]
- Miller, W.A.; Rasochová, L. Barley yellow dwarf viruses. Annu. Rev. Phytopathol. 1997, 35, 167–190. [Google Scholar] [CrossRef] [PubMed]
- Voinnet, O.; Pinto, Y.M.; Baulcombe, D.C. Suppression of gene silencing: A general strategy used by diverse DNA and RNA viruses of plants. Proc. Natl. Acad. Sci. USA 1999, 23, 14147–14152. [Google Scholar] [CrossRef] [PubMed]
- Taylor, N.J.; Halsey, M.; Gaitán-Solís, E.; Anderson, P.; Gichuki, S.; Miano, D.; Bua, A.; Alicai, T.; Fauquet, C.M. The VIRCA Project: Virus resistant cassava for Africa. GM Crops Food 2012, 3, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Chellappan, P.; Masona, M.V.; Vanitharani, R.; Taylor, N.J.; Fauquet, C.M. Broad spectrum resistance to ssDNA viruses associated with transgene-induced gene silencing in cassava. Plant Mol. Biol. 2004, 56, 601–611. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.F.; Rocha-Pena, M.A. Citrus tristeza virus, Plant Disease of International Importance. In Disease of Fruit Crops; Kumar, J., Chaube, H.S., Singh, U.S., Mukhopdhyuy, A.N., Eds.; Prentice Hall: Englewood Cliffs, NJ, USA, 1992; Volume III, pp. 226–249. [Google Scholar]
- Fagoaga, C.; López, C.; de Mendoza, A.H.; Moreno, P.; Navarro, L.; Flores, R.; Peña, L. Post-transcriptional gene silencing of the p23 silencing suppressor of Citrus tristeza virus confers resistance to the virus in transgenic Mexican lime. Plant Mol. Biol. 2006, 60, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Ghorbel, R.; López, C.; Fagoaga, C.; Moreno, P.; Navarro, L.; Flores, R.; Peña, L. Transgenic citrus plants expressing the citrus tristeza virus p23 protein exhibit viral-like symptoms. Mol. Plant Pathol. 2001, 2, 27–36. [Google Scholar] [CrossRef]
- Fagoaga, C.; López, C.; Moreno, P.; Navarro, L.; Flores, R.; Peña, L. Viral-like symptoms induced by the ectopic expression of the p23 gene of Citrus tristeza virus are citrus specific and do not correlate with the pathogenicity of the virus strain. Mol. Plant Microbe Interact. 2005, 18, 435–445. [Google Scholar] [CrossRef]
- Noris, E.; Lucioli, A.; Tavazza, R.; Caciagli, P.; Accotto, G.P.; Tavazza, M. Tomato yellow leaf curl Sardinia virus can overcome transgene-mediated RNA silencing of two essential viral genes. J. Gen. Virol. 2004, 85 Pt 6, 1745–1749. [Google Scholar] [CrossRef]
- Ribeiro, S.G.; Lohuis, H.; Goldbach, R.; Prins, M. Tomato chlorotic mottle virus is a target of RNA silencing but the presence of specific short interfering RNAs does not guarantee resistance in transgenic plants. J. Virol. 2007, 81, 1563–1573. [Google Scholar] [CrossRef]
- Tenllado, F.; Díaz-Ruíz, J.R. Double-stranded RNA-mediated interference with plant virus infection. J. Virol. 2001, 75, 12288–12297. [Google Scholar] [CrossRef] [PubMed]
- Holeva, M.; Sclavounos, A.P.; Kyriakopoulou, P.E.; Voloudakis, A.E. In Vitro produced dsRNA induces resistance against a sever Hellenic CMV isolate in tobacco and tomato. In Proceedings of the 8th International Congress of Plant Molecular Biology, Adelaide, Australia, 20–25 August 2006. [Google Scholar]
- Holeva, M.C.; Sclavounos, A.P.; Milla, S.P.; Kyriakopoulou, P.E.; Voloudakis, A.E. External application of dsRNA of the capsid protein (CP) or 2b gene of CMV reduces the severity of CMV-infection in tobacco. In Proceedings of the VIII International Congress on Molecular Plant–Microbe Interactions, Sorrento, Italy, 21–27 July 2007. [Google Scholar]
- Yin, G.; Sun, Z.; Liu, N.; Zhang, L.; Song, Y.; Zhu, C.; Wen, F. Production of double-stranded RNA for interference with TMV infection utilizing a bacterial prokaryotic expression system. Appl. Microbiol. Biotechnol. 2009, 84, 323–333. [Google Scholar] [CrossRef] [PubMed]
- Gan, D.; Zhang, J.; Jiang, H.; Jiang, T.; Zhu, S.; Cheng, B. Bacterially expressed dsRNA protects maize against SCMV infection. Plant Cell Rep. 2010, 29, 1261–1268. [Google Scholar] [CrossRef] [PubMed]
- Koch, A.; Biedenkopf, B.; Furch, A.C.U.; Abdellatef, E.; Weber, L.; Linicus, L.; Johannsmeier, J.; Jelonek, L.; Goesmann, A.; Cardoza, V.; et al. An RNAi-based control of Fusarium graminearum infections through spraying of long dsRNAs involves a plant passage and is controlled by the fungal silencing machinery. PLoS Pathog. 2016, 12, e1005901. [Google Scholar] [CrossRef]
- Mitter, N.; Worrall, E.A.; Robinson, K.E.; Xu, Z.P.; Carroll, B.J. Induction of virus resistance by exogenous application of double-stranded RNA. Curr. Opin. Virol. 2017, 26, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Konakalla, N.C.; Kaldis, A.; Berbati, M.; Masarapu, H.; Voloudakis, A.E. Exogenous application of double-stranded RNA molecules from TMV p126 and CP genes confers resistance against TMV in tobacco. Planta 2016, 244, 961–969. [Google Scholar] [CrossRef]
- Kaldis, A.; Berbati, M.; Melita, O.; Reppa, C.; Holeva, M.; Otten, P.; Voloudakis, A. Exogenously applied dsRNA molecules deriving from the Zucchini yellow mosaic virus (ZYMV) genome move systemically and protect cucurbits against ZYMV. Mol. Plant Pathol. 2018, 19, 883–895. [Google Scholar] [CrossRef] [PubMed]
- Niehl, A.; Soininen, M.; Poranen, M.M.; Heinlein, M. Synthetic biology approach for plant protection using dsRNA. Plant Biotechnol. J. 2018, 16, 1679–1687. [Google Scholar] [CrossRef]
- Ladewig, K.; Niebert, M.; Xu, Z.P.; Gray, P.P.; Lu, G.Q. Efficient siRNA delivery to mammalian cells using layered double hydroxide nanoparticles. Biomaterials 2010, 31, 1821–1829. [Google Scholar] [CrossRef]
- Ladewig, K.; Xu, Z.P.; Lu, G.Q. Layered double hydroxide nanoparticles in gene and drug delivery. Expert Opin. Drug Deliv. 2009, 6, 907–922. [Google Scholar] [CrossRef]
- Voloudakis, A.E.; Holeva, M.C.; Sarin, L.P.; Bamford, D.H.; Vargas, M.; Poranen, M.M.; Tenllado, F. Efficient double-stranded RNA production methods for utilization in plant virus control. Methods Mol. Biol. 2015, 1236, 255–274. [Google Scholar] [PubMed]


{kind=link}
| Solanaceae | Virus | Abbr. | Type | Target Gene | RNA Type | Construct Length | Reference |
| Tobacco | |||||||
| tobacco mosaic virus | (+) ssRNA | Movement protein | IR (hpRNA) | not mentioned | [94] | ||
| cucumber mosaic virus | CMV | (+) ssRNA | Replicase | IR (hpRNA) | not mentioned | [94] | |
| cucumber mosaic virus | CMV | (+) ssRNA | Coat protein | IR (hpRNA) | 718 bp | [95] | |
| tobacco mosaic virus | TMV | (+) ssRNA | Coat protein | dsRNA | not mentioned | [96] | |
| cucumber mosaic virus | CMV | (+) ssRNA | VSR 2b | amiRNAs | Precursor miR171a | [97] | |
| cucumber mosaic virus | CMV | (+) ssRNA | Coat protein | hpRNA | [98] | ||
| plum pox virus | PPV | (+) ssRNA | VSR P1 and HC-Pro | IR (hpRNA) | 733 bp (UTR/P1) | [99] | |
| 649 bp (P1/HC-Pro) | |||||||
| 706 bp (HC-Pro) | |||||||
| 678 bp (HC-Pro/P3) | |||||||
| cucumber green mottle mosaic virus | CGMMV | (+) ssRNA | Coat protein | IR (hpRNA) | not mentioned | [100] | |
| tomato bushy stunt virus grapevine leaf virus | TBSV | (+) ssRNA | Defective interfering (DI) Movement protein | Virus-derived (DI) RNA | 720 bp | [101] | |
| GFLV | (+) ssRNA | ||||||
| potato virus Y | PVY | (+) ssRNA | Coat protein | fused, tandem, IR; 200 bp (from each virus) | 600 bp fragment (PVY, PVA, PLRV) | [102] | |
| potato virus A | PVA | (+) ssRNA | cylindrical inclusion (CI) | 1000 bp fragment (PVY, PVA, PLRV, TRV, PMTV) | |||
| potato leafroll virus | PLRV | (+) ssRNA | Coat protein | ||||
| tobacco rattle virus | TRV | (+) ssRNA | Replicase | ||||
| potato mop-top virus | PMTV | (+) ssRNA | Replicase | ||||
| potato virus Y | PVY | (+) ssRNA | VSR HC-Pro | amiRNAs | Precursor miR159a, miR167b and miR171a | [103] | |
| potato virus X | PVX | (+) ssRNA | VSR TGBp1, p25 (p25) | ||||
| cassava brown streak Uganda virus | CBSUV CBSV | (+) ssRNA | conserved sequences of P1(CBSV and UCBSV), P3(CBSV and UCBSV), CI(UCBSV), NIb(CBSV and UCBSV), | amiRNA | Precursor miR159a | [104] | |
| cassava brown streak virus | (+) ssRNA | CP(UCBSV) and the 3′UTR | |||||
| cassava brown streak Uganda virus | CBSUV | (+) ssRNA | Coat protein (CBSUV) | hpRNA (pHELLSGATE) | Full-length; N-term. 397 bp; 491 C-term. | [105] | |
| cassava brown streak virus | CBSV | (+) ssRNA | |||||
| African cassava mosaic virus | ACMV | ssDNA | DNA-A and DNA-B genome | dsRNA | 12 constructs of different length | [106] | |
| soybean mosaic virus | SMV | (+) ssRNA | Coat protein | hpRNA (pK7GWIWG2) | not mentioned | [107] | |
| bean yellow mosaic virus | BYMV | (+) ssRNA | Coat protein | hpRNA (pK7GWIWG2) | not mentioned | [107] | |
| citrus psorosis virus | CPsV | (−) ssRNA | Coat protein and 54K gene | hpRNA (pHANNIBAL) | 372 bp (CP); 436 bp (54K) | [108] | |
| citrus tristeza virus | CTV | (+) ssRNA | 3′ p23 (VSR) and 3′ UTR | hpRNA | 900 bp | [109] | |
| tomato golden mosaic virus | TGMV | ssDNA | Replicase (AL1; AL2; AL3 gene) | antisense construct | 1258 bp | [88] | |
| tomato yellow leaf curl virus | TYLCV | ssDNA | Replicase (C1 gene) | antisense RNA | [110] | ||
| tomato leaf curl New Delhi virus | ToLCNDV | ssDNA | virion-sense gene (AV2) | antisense construct | [111] | ||
| cotton leaf curl disease | CLCuD | ssDNA | Replicase (AC1) | sense and antisense RNAs | 446 bp (AC1(5′half)+AC4(ORF)) | [112] | |
| Transcription activator (AC2) | 523 bp (AC1 (3′half)+AC2,AC4) | ||||||
| Rep enhancer (AC3) | 510 bp (AC1(97bp) +AC2+AC3) | ||||||
| chickpea chlorotic dwarf Pakistan virus | CpCDPKV | ssDNA | Rep gene, large intergenic region (LIR) and part of the MP gene | hpRNA | 730 bp | [113] | |
| 5′ Rep, LIR and 5′ MP | |||||||
| pepper golden mosaic virus | PepGMV | ssDNA | Replicase (AC1), intergenic region (IR) and coat protein (AV1) | hpRNA | [114] | ||
| Tomato | potato spindle tuber viroid | PSTVd | (+) ssRNA | Viral sequence | hpRNA | [115] | |
| cucumber mosaic virus | CMV | (+) ssRNA | 2a and 2b genes 3′ UTR | amiRNA | Precursor miR159a | [116] | |
| cucumber mosaic virus | CMV | (+) ssRNA | Replicase | hpRNA | 1138 bp | [117] | |
| tomato yellow leaf curl virus | TYLCV | ssDNA | Replicase (C1) | hpRNA | [118] | ||
| tomato yellow leaf curl virus | TYLCV | ssDNA | Coat protein | hpRNA | [119] | ||
| Potato | potato spindle tuber viroid | PSTVd | (+) ssRNA | Ribunuclease pac1 | dsRNA | [120] | |
| potato virus X | PVX | (+) ssRNA | ORF2 of PVX | hpRNA | 300 bo (ORF2-PVX) | [89] | |
| potato virus Y | PVY | (+) ssRNA | Protease gene PVY | 365 bp (HC-Pro-PVY) | |||
| potato leaf roll virus | PLRV | (+) ssRNA | CP gene PLRV | 300 bp (CP-PLRV) | |||
| sweet potato chlorotic stunt virus | SPCSV | (+) ssRNA | Replicase | hpRNA | [121] | ||
| sweet potato feathery mottle virus | SPFMV | (+) ssRNA | |||||
| potato virus X | PVX | (+) ssRNA | Coat protein | hpRNA | 600 bp: 180 bp (PVX), 240 bp (PVY), 180 bp (PVS) | [122] | |
| potato virus Y | PVY | (+) ssRNA | |||||
| potato virus S | PVS | (+) ssRNA | |||||
| potato virus Y | PVY | (+) ssRNA | CP gene PVY | hpRNA | 600 bp: 200 bp for each target | [102] | |
| potato virus A | PVA | (+) ssRNA | Cylindrical inclusion body PVA | ||||
| potato leaf roll virus | PLRV | (+) ssRNA | CP gene PLRV | ||||
| potato virus Y | PVY | (+) ssRNA | Coat protein | hpRNA | 605 bp | [123] | |
| Cucurbitacea | Virus | Type | Target gene | RNA Type | Construct Length | Reference | |
| Melon | zucchini yellow mosaic virus | ZYMV | (+) ssRNA | Coat protein | antisense | [124] | |
| papaya ringspot virus type W | PRSV-W | (+) ssRNA | Coat protein | hpRNA | [125] | ||
| zucchini yellow mosaic virus cucumber vein yellowing virus | ZYMV CVYV | (+) ssRNA | Cm-eIF4E translation initiation factors (eIF) | hpRNA (pHANNIBAL) | 175 bp | [90] | |
| melon necrotic spot virus | NSV | (+) ssRNA | |||||
| Moroccan watermelon mosaic virus | MWMV | (+) ssRNA | |||||
| (+) ssRNA | |||||||
| Cucumber and Melon | zucchini yellow mosaic virus | ZYMV | (+) ssRNA | VSR HC-Pro | hpRNA (pHANNIBAL) | 657 bp | [126] |
| Watermelon | watermelon silver mottle virus | WSMoV | (+) ssRNA | Partial N gene of WSMoV fused to partial CP gene sequences of CMV, CGMMV, WMV | silencer DNA (the middle half N gene of (WSMoV)) | 449 bp CP-CMV | [127] |
| cucumber mosaic virus | CMV | (+) ssRNA | 449 bp CP-CGMMV | ||||
| cucumber green mottle mosaic virus | CGMMV | (+) ssRNA | 449 bp CP-WMV | ||||
| watermelon mosaic virus | WMV | (+) ssRNA | |||||
| Fabaceae | Virus | Type | Target gene | RNA type | Construct length | Reference | |
| Soybean | soybean dwarf virus | SbDV | (+) ssRNA | Coat protein | hpRNA | 602 bp | [128] |
| alfalfa mosaic virus | AMV | (+) ssRNA | highly conserved sequences of one virus | Short dsRNA stems | 109 bp (AMV) | [129] | |
| bean pod mottle virus | BPMV | (+) ssRNA | 147 bp (BPMV) | ||||
| soybean mosaic virus | SMV | (+) ssRNA | 123 bp (SMV) | ||||
| soybean mosaic virus | SMV | (+) ssRNA | Coat protein | hpRNA | [130] | ||
| soybean mosaic virus | SMV | (+) ssRNA | VSR HC-Pro | hpRNA | [131] | ||
| soybean mosaic virus | SMV | (+) ssRNA | VSR HC-Pro | hpRNA | 268 bp | [132] | |
| soybean mosaic virus | SMV | (+) ssRNA | P3 cistron | hpRNA | 302 bp | [133] | |
| Common bean | bean golden mosaic virus | BGMV | (+) ssRNA | Replicase (AC1) | hpRNA | 411 bp | [134] |
| Cowpea | cowpea severe mosaic virus | CPSMV | (+) ssRNA | proteinase cofactor (CPSMV) Coat protein (CABMV) | hpRNA | 899 bp (415 bp RNA1- CPSMV and 462 bp CP-CABMV) | [135] |
| cowpea aphid-borne mosaic virus | CABMV | (+) ssRNA | |||||
| White clover | white clover mosaic virus | WCMV | (+) ssRNA | Replicase | sense, antisense and hpRNA | 790 bp | [136] |
| Poaceae | Virus | Type | Target gene | RNA type | Construct length | Reference | |
| Rice | rice tungro bacilliform virus | RTBV | dsDNA | ORF IV | dsRNA | 1326 bp | [137] |
| rice dwarf virus | RDV | dsRNA | non-structural protein Pns12 and Pns4 | hpRNA | 500 bp: Pns12-12N | [138] | |
| 499 bp: Pns12-12C | |||||||
| 500 bp: Pns4-4N | |||||||
| 504 bp: Pns4-4M | |||||||
| rice stripe virus | RSV | (−) ssRNA | Coat protein and special-disease protein (SP) | hpRNA | 300 bp (SP); 450 bp (CP) | [139] | |
| 700 bp (SP/CP; CP/SP) | |||||||
| rice stripe virus | RSV | (−) ssRNA | Nucleocapsid (pC3), MP (pC4), glycoprotein (pC2), non-structural protein (p4) | hpRNA | 500 bp (for each target) | [140] | |
| rice black streaked dwarf virus | RBSDV | dsRNA | nonstructural Pns9 protein | hpRNA | 500 bp | [141] | |
| rice grassy stunt virus | RGSV | (−) ssRNA | nucelocapsid protein pC5; movement protein pC6 | dsRNA | 500 bp (for each target) | [142] | |
| Wheat | wheat streak mosaic virus | WSMV | (−) ssRNA | nuclear inclusion protein ‘a’ (NIa) gene | hpRNA (pSTARGATE) | 696 bp | [143] |
| wheat streak mosaic virus | WSMV | (−) ssRNA | Conserved region of WSMV genome | amiRNA | Precursor miR395 | [144] | |
| Maize | maize dwarf mosaic virus | MDMV | (+) ssRNA | Coat protein | hpRNA | 404 bp | [145] |
| Barley | barley yellow dwarf virus | BYDV | (+) ssRNA | Polymerase gene | hpRNA | 1600 bp | [146] |
| Euphorbiaceae | Virus | Type | Target gene | RNA type | Construct length | Reference | |
| Cassava | African cassava mosaic virus | ACMV | ssDNA | Rep (AC1); TrAP (AC2), REn (AC3) | antisense construct | Full-length of each target gene | [147] |
| African cassava mosaic virus | ACMV | ssDNA | bidirectional promoter of ACMV DNA-A | intron-containing dsRNA | 256 bp | [148] | |
| African cassava mosaic virus | ACMV | ssDNA | Rep (AC1) | hpRNA | 154 bp | [149] | |
| Sir Lankan cassava mosaic virus | (SLCMV) | ssDNA | AV1 and AV2 | hpRNA | 527 bp | [117] | |
| cassava brown streak Uganda virus | CBSUV CBSV | (+) ssRNA | Coat protein | hpRNA | p5001 construct | [150] | |
| cassava brown streak virus | (+) ssRNA | ||||||
| cassava brown streak Uganda virus | CBSUV CBSV | (+) ssRNA | Coat protein | hpRNA | Field trials (Chauhan et al. 2015) | [151] | |
| cassava brown streak virus | (+) ssRNA | ||||||
| cassava brown streak Uganda virus | CBSUV CBSV | (+) ssRNA | Coat protein | hpRNA | 525 bp | [152] | |
| cassava brown streak virus | (+) ssRNA | ||||||
| cassava brown streak Uganda virus | CBSUV CBSV | (+) ssRNA | Coat protein | hpRNA | 894 bp (full-length CP) | [153] | |
| cassava brown streak virus | (+) ssRNA | 401 bp (CP-NT) | |||||
| cassava brown streak Uganda virus | CBSUV CBSV | (+) ssRNA | Coat protein | hpRNA | Field trials (Yadav et al. 2011) | [154] | |
| cassava brown streak virus | (+) ssRNA | ||||||
| cassava brown streak Uganda virus | CBSUV CBSV | (+) ssRNA | Coat protein | hpRNA | Field trials (Yadav et al. 2011) | [155] | |
| cassava brown streak virus | (+) ssRNA | ||||||
| cassava brown streak virus | CBSV | (+) ssRNA | Coat protein | hpRNA | 895 bp CP-CBSV | [156] | |
| cassava brown streak Uganda virus | CBSUV | 898 bp CP-CBSUV | |||||
| Poinsettia | poinsettia mosaic virus | PnMV | (+) ssRNA | Coat protein and RNA-dependent RNA-polymerase | hpRNA | 500 bp each target | [157] |
| Tropical fruits | Virus | Type | Target gene | RNA type | Construct length | Reference | |
| Citrus macrophylla | citrus tristeza virus | CTV | (+) ssRNA | 3′ p23 (VSR) and 3′ UTR | hpRNA | 900 bp | [109] |
| Mexican lime | citrus tristeza virus | CTV | (+) ssRNA | 3′ p23 (VSR) and 3′ UTR | sense, antisense and hpRNA | 549 bp | [158] |
| citrus tristeza virus | CTV | (+) ssRNA | VSR p20, p23, p25 | hpRNA | 548 bp (p20); 629 bp (p23); 670 bp (p25) | [159] | |
| citrus psorosis virus | CPsV | (−) ssRNA | Coat protein and 54K | hpRNA | 372 bp CP; 436 bp 54K | [160] | |
| Sweet orange | citrus psorosis virus | CPsV | (−) ssRNA | Coat protein, 54K or 24K genes | hpRNA | 372 bp CP; 436 bp 54K; 312 bp 24K | [108] |
| Banana | banana bunchy top virus | BBTV | ssDNA | Replicase | hpRNA | CP-full-length; 651 bp partial CP sequence | [92] |
| Brassicaceae | Virus | Type | Target gene | RNA type | Construct length | Reference | |
| Arabidopsis | turnip yellow mosaic virus | TYMV | (+) ssRNA | VSR P69 (TYMV) | amiRNAs | Precursor miR159a | [161] |
| turnip mosaic virus | TuMV | (+) ssRNA | VSR HC-Pro (TuMV) | ||||
| cucumber mosaic virus | CMV | (+) ssRNA | 3′ UTR | amiRNAs | Precursor miR159a | [162] | |
| Others | Virus species | Type | Target gene | RNA type | Construct length | Reference | |
| Asteraceae | chrysanthemum virus B | CVB | (+) ssRNA | Coat protein | sense, antisense and hpRNA | 273 bp | [163] |
| Amaranthaceae | beet necrotic yellow vein virus | BNYVV | (+) ssRNA | Replicase | hpRNA | 459 bp, 589 bp and 824 bp | [164] |
| Andropogoneae | sugarcane mosaic virus | SCMV | (+) ssRNA | Coat protein | hpRNA | Not mentioned | [165] |
| sorghum mosaic virus | SrMV | (+) ssRNA | |||||
| Vitaceae | grapevine fanleaf virus | GFLV | (+) ssRNA | Coat protein | amiRNA | Precursor miR319a | [91] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaffar, F.Y.; Koch, A. Catch Me If You Can! RNA Silencing-Based Improvement of Antiviral Plant Immunity. Viruses 2019, 11, 673. https://doi.org/10.3390/v11070673
Gaffar FY, Koch A. Catch Me If You Can! RNA Silencing-Based Improvement of Antiviral Plant Immunity. Viruses. 2019; 11(7):673. https://doi.org/10.3390/v11070673
Chicago/Turabian StyleGaffar, Fatima Yousif, and Aline Koch. 2019. "Catch Me If You Can! RNA Silencing-Based Improvement of Antiviral Plant Immunity" Viruses 11, no. 7: 673. https://doi.org/10.3390/v11070673
APA StyleGaffar, F. Y., & Koch, A. (2019). Catch Me If You Can! RNA Silencing-Based Improvement of Antiviral Plant Immunity. Viruses, 11(7), 673. https://doi.org/10.3390/v11070673