Potential Viroporin Candidates From Pathogenic Viruses Using Bacteria-Based Bioassays



Abstract
1. Introduction
2. Materials and Methods
2.1. Protein Search
2.2. Bacterial Strains
2.3. Plasmids
2.4. Chemicals
2.5. Growth Media
2.6. Bacterial Growth
2.7. Maltose Complementation Assay
2.8. pHlux Assay
2.9. Experimental Controls
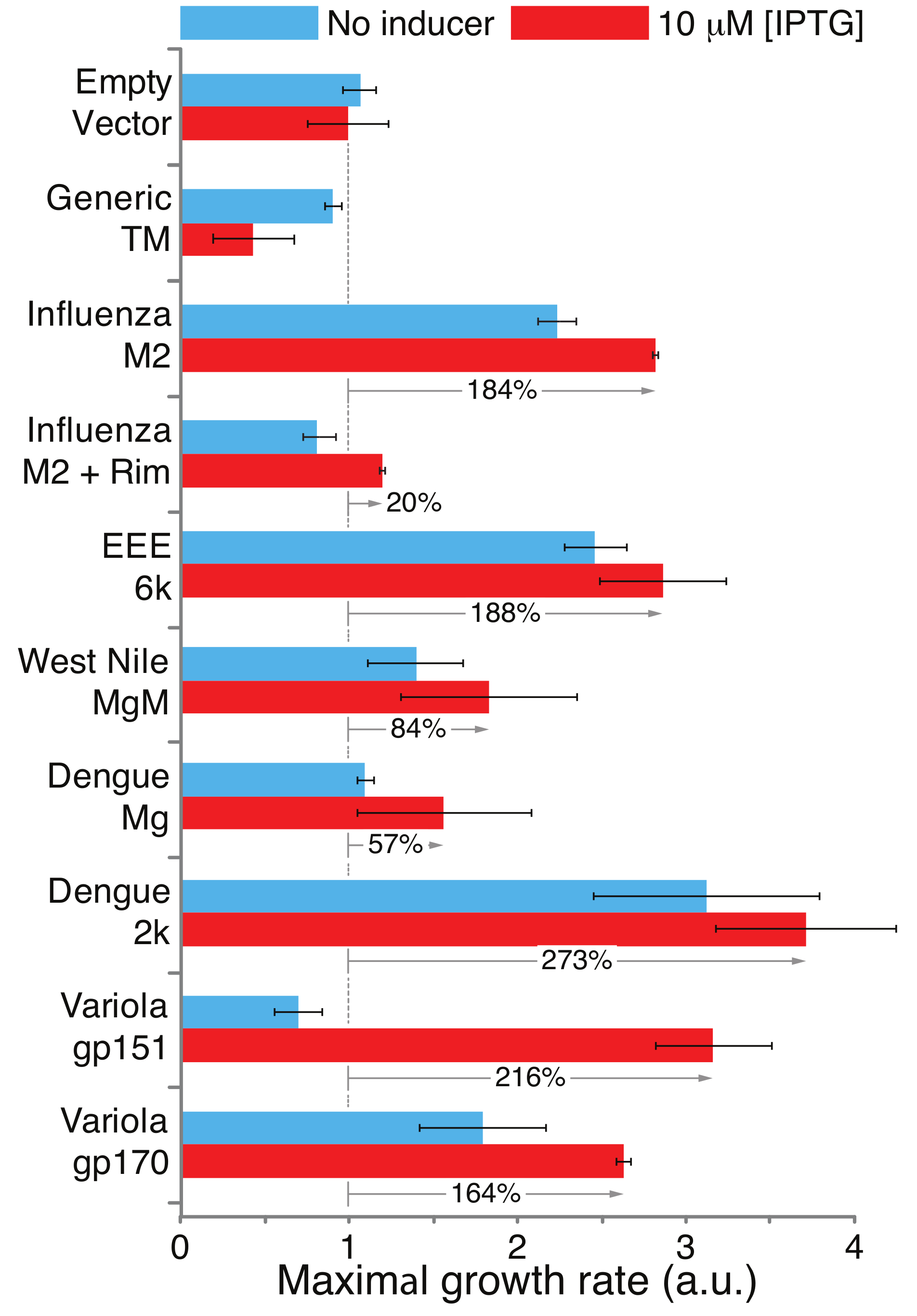
3. Results
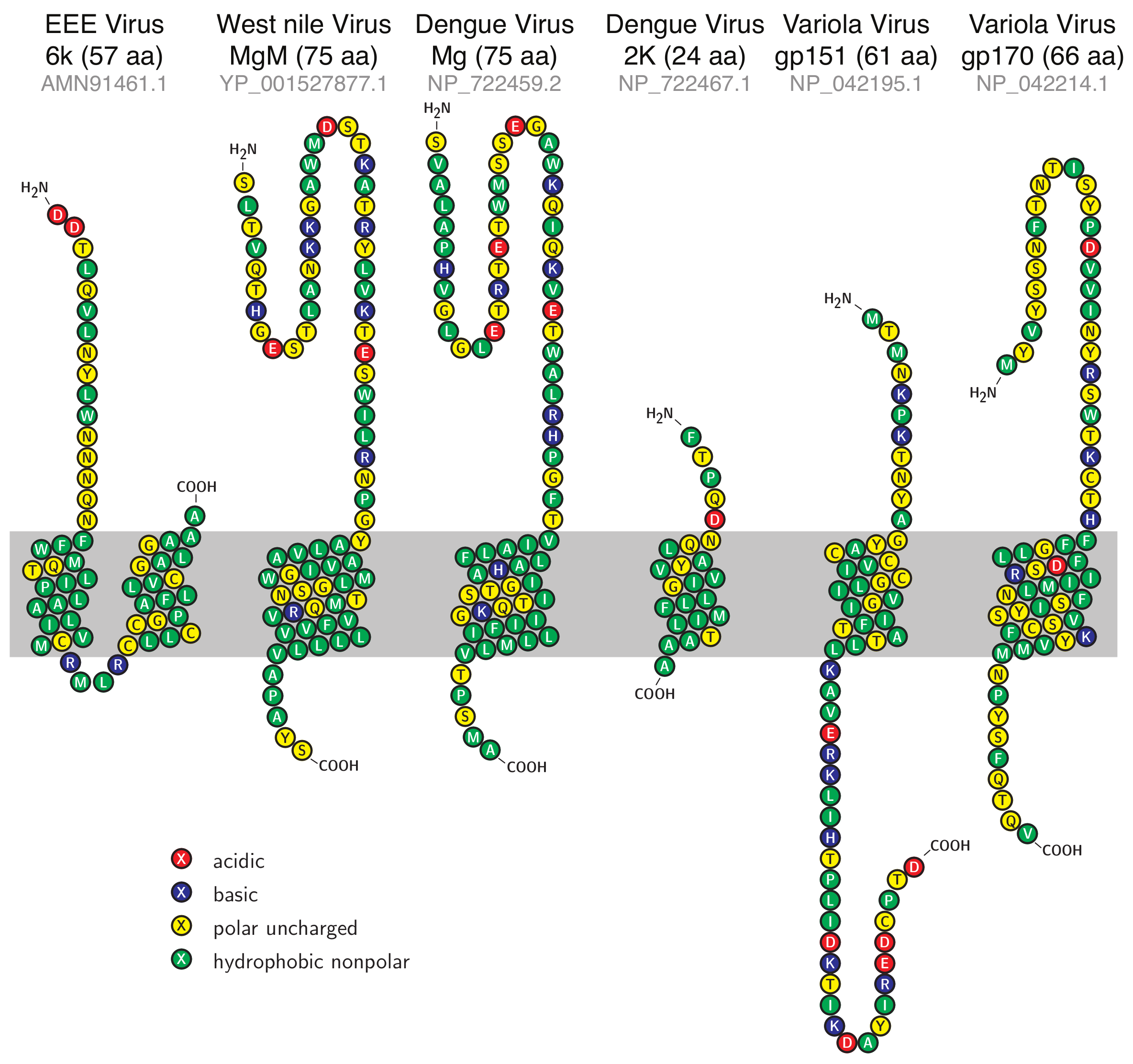
3.1. Sequence Analysis
3.2. Expression and Membrane Incorporation
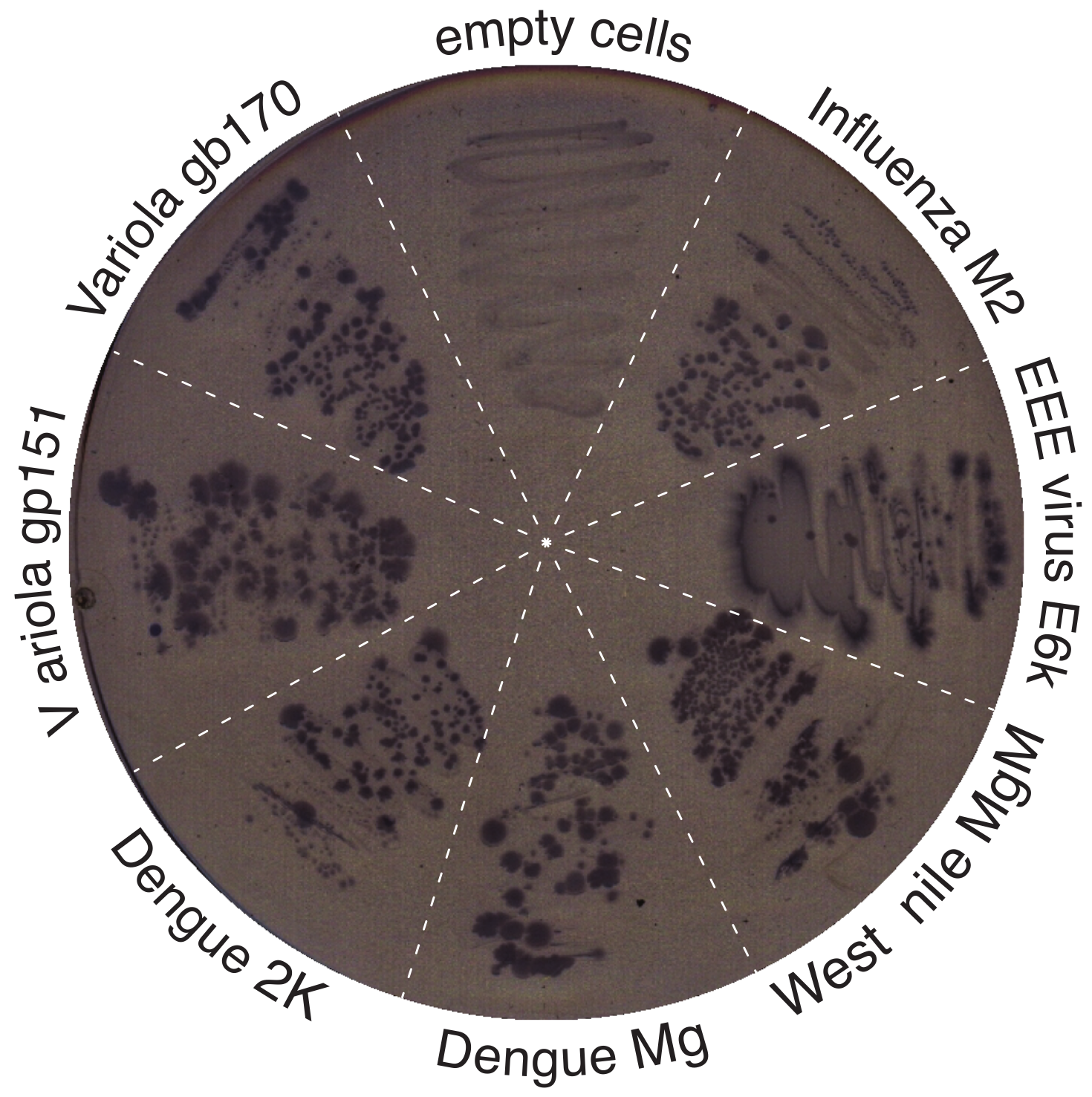
3.3. Positive Genetic Test
3.4. Negative Genetic Test
3.5. Proton Flux
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| EEE | Eastern equine encephalitis |
| MBP | Maltose binding protein |
| HIV | Human immunodeficiency virus |
| IPTG | isopropyl β-d-1-thiogalactopyranoside |
| LB | Lysogeny Broth |
| LBK | Lysogeny Broth in which Na+ is substituted by K+ |
| TM | Transmembrane |
References
- Gonzalez, M.E.; Carrasco, L. Viroporins. FEBS Lett. 2003, 552, 28–34. [Google Scholar] [CrossRef]
- DiMaio, D. Viral Miniproteins. Annu. Rev. Microbiol. 2014, 68, 21–43. [Google Scholar] [CrossRef] [PubMed]
- Fung, T.S.; Torres, J.; Liu, D.X. The Emerging Roles of Viroporins in ER Stress Response and Autophagy Induction During Virus Infection. Viruses 2015, 7, 2834–2857. [Google Scholar] [CrossRef] [PubMed]
- Sze, C.W.; Tan, Y.J. Viral Membrane Channels: Role and Function in the Virus Life Cycle. Viruses 2015, 7, 3261–3284. [Google Scholar] [CrossRef] [PubMed]
- Royle, J.; Dobson, S.J.; Müller, M.; Macdonald, A. Emerging Roles of Viroporins Encoded by DNA Viruses: Novel Targets for Antivirals? Viruses 2015, 7, 5375–5387. [Google Scholar] [CrossRef] [PubMed]
- To, J.; Surya, W.; Torres, J. Targeting the Channel Activity of Viroporins. Adv. Protein Chem. Struct. Biol. 2016, 104, 307–355. [Google Scholar] [PubMed]
- Lamb, R.A.; Lai, C.J. Conservation of the Influenza Virus Membrane Protein (M1) Amino Acid Sequence and an Open Reading Frame of RNA Segment 7 Encoding a Second Protein (M2) in H1N1 and H3N2 Strains. Virology 1981, 112, 746–751. [Google Scholar] [CrossRef]
- Lamb, R.A.; Choppin, P.W. Identification of a Second Protein (M2) Encoded by RNA Segment 7 of Influenza Virus. Virology 1981, 112, 729–737. [Google Scholar] [CrossRef]
- Hay, A.J.; Wolstenholme, A.J.; Skehel, J.J.; Smith, M.H. The Molecular Basis of the Specific Anti-Influenza Action of Amantadine. EMBO J. 1985, 4, 3021–3024. [Google Scholar] [CrossRef] [PubMed]
- Pinto, L.H.; Holsinger, L.J.; Lamb, R.A. Influenza Virus M2 Protein Has Ion Channel Activity. Cell 1992, 69, 517–528. [Google Scholar] [CrossRef]
- Holsinger, L.J.; Lamb, R.A. Influenza Virus M2 Integral Membrane Protein Is a Homotetramer Stabilized by Formation of Disulfide Bonds. Virology 1991, 183, 32–43. [Google Scholar] [CrossRef]
- Stouffer, A.L.; Acharya, R.; Salom, D.; Levine, A.S.; Di Costanzo, L.; Soto, C.S.; Tereshko, V.; Nanda, V.; Stayrook, S.; DeGrado, W.F. Structural Basis for the Function and Inhibition of an Influenza Virus Proton Channel. Nature 2008, 451, 596–599. [Google Scholar] [CrossRef] [PubMed]
- Schnell, J.R.; Chou, J.J. Structure and Mechanism of the M2 Proton Channel of Influenza a Virus. Nature 2008, 451, 591–595. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Yi, M.; Dong, H.; Qin, H.; Peterson, E.; Busath, D.D.; Zhou, H.X.; Cross, T.A. Insight Into the Mechanism of the Influenza a Proton Channel From a Structure in a Lipid Bilayer. Science 2010, 330, 509–512. [Google Scholar] [CrossRef] [PubMed]
- Paterson, R.G.; Takeda, M.; Ohigashi, Y.; Pinto, L.H.; Lamb, R.A. Influenza B Virus BM2 Protein Is an Oligomeric Integral Membrane Protein Expressed at the Cell Surface. Virology 2003, 306, 7–17. [Google Scholar] [CrossRef]
- Horvath, C.M.; Williams, M.A.; Lamb, R.A. Eukaryotic Coupled Translation of Tandem Cistrons: Identification of the Influenza B Virus BM2 Polypeptide. EMBO J. 1990, 9, 2639–2647. [Google Scholar] [CrossRef] [PubMed]
- Hongo, S.; Ishii, K.; Mori, K.; Takashita, E.; Muraki, Y.; Matsuzaki, Y.; Sugawara, K. Detection of ion channel activity in Xenopus laevis oocytes expressing Influenza C virus CM2 protein. Arch. Virol. 2004, 149, 35–50. [Google Scholar] [CrossRef]
- Hongo, S.; Sugawara, K.; Nishimura, H.; Muraki, Y.; Kitame, F.; Nakamura, K. Identification of a Second Protein Encoded by Influenza C Virus RNA Segment 6. J. Gen. Virol. 1994, 75 Pt 12, 3503–3510. [Google Scholar] [CrossRef]
- Schubert, U.; Ferrer-Montiel, A.V.; Oblatt-Montal, M.; Henklein, P.; Strebel, K.; Montal, M. Identification of an Ion Channel Activity of the Vpu Transmembrane Domain and Its Involvement in the Regulation of Virus Release From HIV-1-Infected Cells. FEBS Lett. 1996, 398, 12–18. [Google Scholar] [CrossRef]
- Strebel, K.; Klimkait, T.; Martin, M.A. A Novel Gene of HIV-1, Vpu, and Its 16-Kilodalton Product. Science 1988, 241, 1221–1223. [Google Scholar] [CrossRef]
- Cohen, E.A.; Terwilliger, E.F.; Sodroski, J.G.; Haseltine, W.A. Identification of a Protein Encoded by the Vpu Gene of HIV-1. Nature 1988, 334, 532–534. [Google Scholar] [CrossRef] [PubMed]
- Griffin, S.D.C.; Beales, L.P.; Clarke, D.S.; Worsfold, O.; Evans, S.D.; Jaeger, J.; Harris, M.P.G.; Rowlands, D.J. The p7 Protein of Hepatitis C Virus Forms an Ion Channel That Is Blocked by the Antiviral Drug, Amantadine. FEBS Lett. 2003, 535, 34–38. [Google Scholar] [CrossRef]
- Lin, C.; Lindenbach, B.D.; Prágai, B.M.; McCourt, D.W.; Rice, C.M. Processing in the Hepatitis C Virus E2-NS2 Region: Identification of p7 and Two Distinct E2-Specific Products with Different C Termini. J. Virol. 1994, 68, 5063–5073. [Google Scholar] [PubMed]
- He, J.; Melnik, L.I.; Komin, A.; Wiedman, G.; Fuselier, T.; Morris, C.F.; Starr, C.G.; Searson, P.C.; Gallaher, W.R.; Hristova, K.; et al. Ebola Virus Delta Peptide is a Viroporin. J. Virol. 2017, 91, e00438-17. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.S.; Liao, C.L.; Tsao, C.H.; Chen, M.C.; Liu, C.I.; Chen, L.K.; Lin, Y.L. Membrane permeabilization by small hydrophobic nonstructural proteins of Japanese encephalitis virus. J. Virol. 1999, 73, 6257–6264. [Google Scholar] [PubMed]
- León-Juárez, M.; Martínez-Castillo, M.; Shrivastava, G.; García-Cordero, J.; Villegas-Sepulveda, N.; Mondragón-Castelán, M.; Mondragón-Flores, R.; Cedillo-Barrón, L. Recombinant Dengue virus protein NS2B alters membrane permeability in different membrane models. Virol. J. 2016, 13, 1. [Google Scholar] [CrossRef]
- Han, Z.; Harty, R.N. The NS3 protein of bluetongue virus exhibits viroporin-like properties. J. Biol. Chem. 2004, 279, 43092–43097. [Google Scholar] [CrossRef]
- Gan, S.W.; Ng, L.; Lin, X.; Gong, X.; Torres, J. Structure and Ion Channel Activity of the Human Respiratory Syncytial Virus (hRSV) Small Hydrophobic Protein Transmembrane Domain. Protein Sci. 2008, 17, 813–820. [Google Scholar] [CrossRef]
- Collins, P.L.; Olmsted, R.A.; Johnson, P.R. The Small Hydrophobic Protein of Human Respiratory Syncytial Virus: Comparison Between Antigenic Subgroups a and B. J. Gen. Virol. 1990, 71 Pt 7, 1571–1576. [Google Scholar] [CrossRef]
- Strebel, K. HIV-1 Vpu - an Ion Channel in Search of a Job. Biochim. Biophys. Acta 2014, 1838, 1074–1081. [Google Scholar] [CrossRef]
- Rosenberg, M.R.; Weaver, L.M.; Casarotto, M.G. Probing Interactions of Vpu From HIV-1 with Amiloride-Based Compounds. Biochim. Biophys. Acta 2016, 1858, 733–739. [Google Scholar] [CrossRef] [PubMed]
- Pavlović, D.; Neville, D.C.; Argaud, O.; Blumberg, B.; Dwek, R.A.; Fischer, W.B.; Zitzmann, N. The Hepatitis C Virus p7 Protein Forms an Ion Channel That Is Inhibited by Long-Alkyl-Chain Iminosugar Derivatives. Proc. Natl. Acad. Sci. USA 2003, 100, 6104–6108. [Google Scholar]
- Luscombe, C.A.; Huang, Z.; Murray, M.G.; Miller, M.; Wilkinson, J.; Ewart, G.D. A Novel Hepatitis C Virus p7 Ion Channel Inhibitor, BIT225, Inhibits Bovine Viral Diarrhea Virus in Vitro and Shows Synergism with Recombinant Interferon-Alpha-2b and Nucleoside Analogues. Antivir. Res. 2010, 86, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Rey-Carrizo, M.; Torres, E.; Ma, C.; Barniol-Xicota, M.; Wang, J.; Wu, Y.; Naesens, L.; DeGrado, W.F.; Lamb, R.A.; Pinto, L.H.; et al. 3-Azatetracyclo[5.2.1.1(5,8).0(1,5)]Undecane Derivatives: From Wild-Type Inhibitors of the M2 Ion Channel of Influenza a Virus to Derivatives with Potent Activity Against the V27A Mutant. J. Med. Chem. 2013, 56, 9265–9274. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Cady, S.D.; Balannik, V.; Pinto, L.H.; DeGrado, W.F.; Hong, M. Discovery of Spiro-Piperidine Inhibitors and Their Modulation of the Dynamics of the M2 Proton Channel From Influenza a Virus. J. Am. Chem. Soc. 2009, 131, 8066–8076. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Griffin, D.E. Fields Virology; Wolters Kluwer: Philadelphia, PA, USA, 2013; Chapter 23. [Google Scholar]
- Pierson, T.C.; Diamond, M.S. Fields Virology; Wolters Kluwer: Philadelphia, PA, USA, 2013; Chapter 26. [Google Scholar]
- Moss, B. Fields Virology; Wolters Kluwer: Philadelphia, PA, USA, 2013; Chapter 66. [Google Scholar]
- Käll, L.; Krogh, A.; Sonnhammer, E. L Advantages of Combined Transmembrane Topology and Signal Peptide Prediction—The Phobius Web Server. Nucleic Acids Res. 2007, 35, W429–W432. [Google Scholar] [CrossRef] [PubMed]
- Käll, L.; Krogh, A.; Sonnhammer, E.L. A Combined Transmembrane Topology and Signal Peptide Prediction Method. J. Mol. Biol. 2004, 338, 1027–1036. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, K.; Stoffel, W. TMbase—A database of membrane spanning proteins segments. Biol. Chem. Hoppe Seyler 1993, 374, 166. [Google Scholar]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. A model recognition approach to the prediction of all-helical membrane protein structure and topology. Biochemistry 1994, 33, 3038–3049. [Google Scholar] [CrossRef] [PubMed]
- Stumpe, S.; Bakker, E.P. Requirement of a Large K+-Uptake Capacity and of Extracytoplasmic Protease Activity for Protamine Resistance of Escherichia Coli. Arch. Microbiol. 1997, 167, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Miesenböck, G.; De Angelis, D.A.; Rothman, J.E. Visualizing Secretion and Synaptic Transmission with pH-Sensitive Green Fluorescent Proteins. Nature 1998, 394, 192–195. [Google Scholar] [CrossRef]
- Bertani, G. Studies on Lysogenesis. I. The Mode of Phage Liberation by Lysogenic Escherichia Coli. J. Bacteriol. 1951, 62, 293–300. [Google Scholar] [PubMed]
- Santner, P.; Martins, J.M.d.S.; Kampmeyer, C.; Hartmann-Petersen, R.; Laursen, J.S.; Stein, A.; Olsen, C.A.; Arkin, I.T.; Winther, J.R.; Willemoës, M.; et al. Random Mutagenesis Analysis of the Influenza A M2 Proton Channel Reveals Novel Resistance Mutants. Biochemistry 2018, 57, 5957–5968. [Google Scholar] [CrossRef] [PubMed]
- Santner, P.; Martins, J.M.d.S.; Laursen, J.S.; Behrendt, L.; Riber, L.; Olsen, C.A.; Arkin, I.T.; Winther, J.R.; Willemoës, M.; Lindorff-Larsen, K. A Robust Proton Flux (pHlux) Assay for Studying the Function and Inhibition of the Influenza A M2 Proton Channel. Biochemistry 2018, 57, 5949–5956. [Google Scholar] [CrossRef] [PubMed]
- Lemmon, M.A.; Flanagan, J.M.; Treutlein, H.R.; Zhang, J.; Engelman, D.M. Sequence specificity in the dimerization of transmembrane alpha-helices. Biochemistry 1992, 31, 12719–12725. [Google Scholar] [CrossRef] [PubMed]
- Lemmon, M.A.; Flanagan, J.M.; Hunt, J.F.; Adair, B.D.; Bormann, B.J.; Dempsey, C.E.; Engelman, D.M. Glycophorin A dimerization is driven by specific interactions between transmembrane alpha-helices. J. Biol. Chem. 1992, 267, 7683–7689. [Google Scholar] [PubMed]
- Mould, J.A.; Paterson, R.G.; Takeda, M.; Ohigashi, Y.; Venkataraman, P.; Lamb, R.A.; Pinto, L.H. Influenza B virus BM2 protein has ion channel activity that conducts protons across membranes. Dev. Cell 2003, 5, 175–184. [Google Scholar] [CrossRef]
- Antoine, A.F.; Montpellier, C.; Cailliau, K.; Browaeys-Poly, E.; Vilain, J.P.; Dubuisson, J. The Alphavirus 6K Protein Activates Endogenous Ionic Conductances When Expressed in Xenopus Oocytes. J. Membr. Biol. 2007, 215, 37–48. [Google Scholar] [CrossRef]
- Melton, J.V.; Ewart, G.D.; Weir, R.C.; Board, P.G.; Lee, E.; Gage, P.W. Alphavirus 6K Proteins Form Ion Channels. J. Biol. Chem. 2002, 277, 46923–46931. [Google Scholar] [CrossRef] [PubMed]
- Sanz, M.A.; Pérez, L.; Carrasco, L. Semliki Forest Virus 6K Protein Modifies Membrane Permeability After Inducible Expression in Escherichia Coli Cells. J. Biol. Chem. 1994, 269, 12106–12110. [Google Scholar]
- Premkumar, A.; Horan, C.R.; Gage, P.W. Dengue virus M protein C-terminal peptide (DVM-C) forms ion channels. J. Membr. Biol. 2005, 204, 33–38. [Google Scholar] [CrossRef]
- Johnson, M.; Zaretskaya, I.; Raytselis, Y.; Merezhuk, Y.; McGinnis, S.; Madden, T.L. NCBI BLAST: A better web interface. Nucleic Acids Res. 2008, 36, W5–W9. [Google Scholar] [CrossRef] [PubMed]
- Beitz, E. TEXtopo: Shaded Membrane Protein Topology Plots in LATEX2ε. Bioinformatics 2000, 16, 1050–1051. [Google Scholar] [CrossRef] [PubMed]
- Reuten, R.; Nikodemus, D.; Oliveira, M.B.; Patel, T.R.; Brachvogel, B.; Breloy, I.; Stetefeld, J.; Koch, M. Maltose-Binding Protein (MBP), a Secretion-Enhancing Tag for Mammalian Protein Expression Systems. PLoS ONE 2016, 11, e0152386. [Google Scholar] [CrossRef] [PubMed]
- Astrahan, P.; Flitman-Tene, R.; Bennett, E.R.; Krugliak, M.; Gilon, C.; Arkin, I.T. Quantitative Analysis of Influenza M2 Channel Blockers. Biochim. Biophys. Acta 2011, 1808, 394–398. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Alhadeff, R.; Assa, D.; Astrahan, P.; Krugliak, M.; Arkin, I.T. Computational and Experimental Analysis of Drug Binding to the Influenza M2 Channel. Biochim. Biophys. Acta 2014, 1838, 1068–1073. [Google Scholar] [CrossRef][Green Version]
- Assa, D.; Alhadeff, R.; Krugliak, M.; Arkin, I.T. Mapping the Resistance Potential of Influenza’s H+ Channel Against an Antiviral Blocker. J. Mol. Biol. 2016, 428, 4209–4217. [Google Scholar] [CrossRef] [PubMed]
- Taube, R.; Alhadeff, R.; Assa, D.; Krugliak, M.; Arkin, I.T. Bacteria-Based Analysis of HIV-1 Vpu Channel Activity. PLoS ONE 2014, 9, e105387. [Google Scholar] [CrossRef]
- Bassford, P.; Beckwith, J. Escherichia Coli Mutants Accumulating the Precursor of a Secreted Protein in the Cytoplasm. Nature 1979, 277, 538–541. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Protein | Accession No. | Length |
|---|---|---|---|
| EEE virus | 6k protein | AMN91461.1 | 57 |
| West Nile virus | Membrane glycoprotein M (MgM) | YP_001527877.1 | 75 |
| Dengue virus 1 | Membrane glycoprotein (Mg) | NP_722459.2 | 75 |
| Dengue virus 1 | 2k Protein | NP_722467.1 | 24 |
| Variola virus | gp151 | NP_042195.1 | 61 |
| Variola virus | gp170 | NP_042214.1 | 66 |
| Proteins | Phobious [39,40] | TMpred [41] | MEMSAT [42] |
|---|---|---|---|
| EEEV 6k protein | 17–33; 38–56 | 16–37; 38–57 | No TM detected |
| WNV MgM | 42–70 | 42–75 | 41–58 |
| DV Mg | 44–70 | 41–75 | 44–68 |
| DV 2k | 6–23 | 8–24 | Too short for prediction |
| VV gp151 | 12–32 | 12–32 | 12–32 |
| VV gp170 | 30–57 | 39–57 | 39–57 |
| 6k | MgM | Mg | 2k | gp151 | gp170 | |
|---|---|---|---|---|---|---|
| 6k | 0.077 | 2.9 | 4.5 | - | 0.75 | |
| MgM | 0.031 | 0.32 | - | - | ||
| Mg | 2.9 | 4.3 | - | - | ||
| 2k | - | - | - | - | - | |
| gp151 | - | - | - | - | - | |
| gp170 | 0.75 | - | - | 0.003 | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomar, P.P.S.; Oren, R.; Krugliak, M.; Arkin, I.T. Potential Viroporin Candidates From Pathogenic Viruses Using Bacteria-Based Bioassays. Viruses 2019, 11, 632. https://doi.org/10.3390/v11070632
Tomar PPS, Oren R, Krugliak M, Arkin IT. Potential Viroporin Candidates From Pathogenic Viruses Using Bacteria-Based Bioassays. Viruses. 2019; 11(7):632. https://doi.org/10.3390/v11070632
Chicago/Turabian StyleTomar, Prabhat Pratap Singh, Rivka Oren, Miriam Krugliak, and Isaiah T. Arkin. 2019. "Potential Viroporin Candidates From Pathogenic Viruses Using Bacteria-Based Bioassays" Viruses 11, no. 7: 632. https://doi.org/10.3390/v11070632
APA StyleTomar, P. P. S., Oren, R., Krugliak, M., & Arkin, I. T. (2019). Potential Viroporin Candidates From Pathogenic Viruses Using Bacteria-Based Bioassays. Viruses, 11(7), 632. https://doi.org/10.3390/v11070632