Identification, Pathogenicity of Novel Fowl Adenovirus Serotype 4 SDJN0105 in Shandong, China and Immunoprotective Evaluation of the Newly Developed Inactivated Oil-emulsion FAdV-4 Vaccine


Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus Preparation
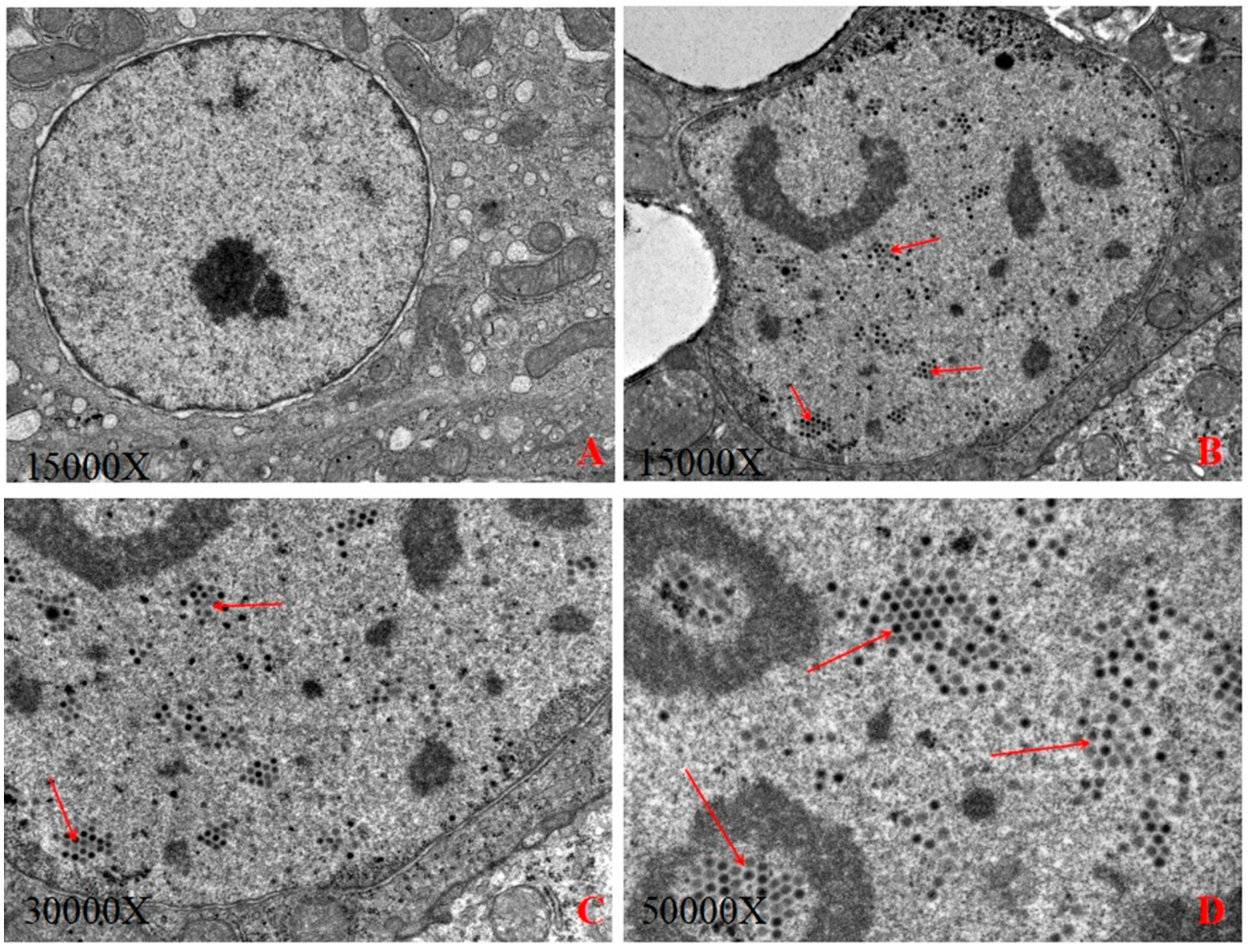
2.2. Electron Microscopic Examination
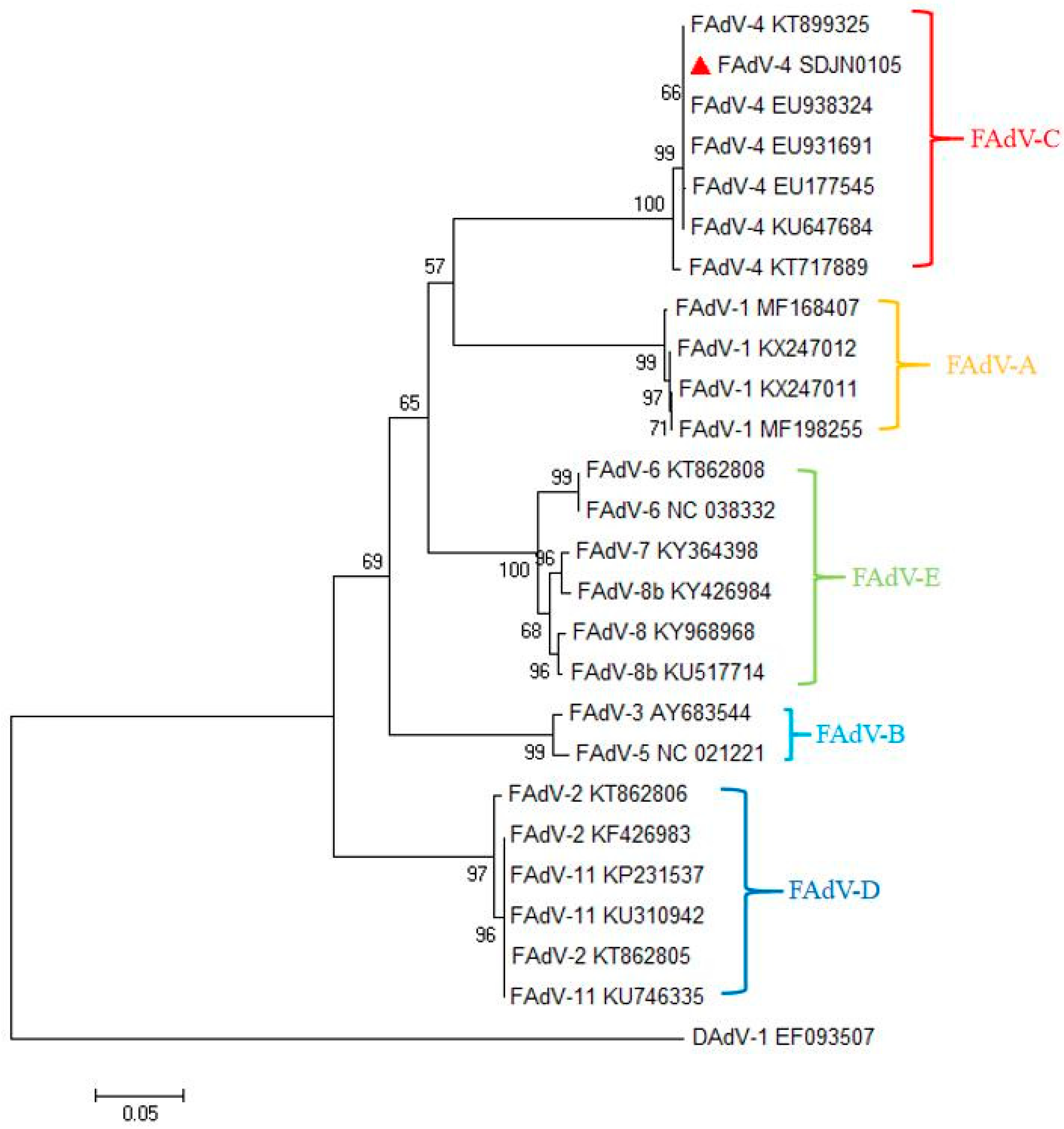
2.3. PCR and Phylogenetic Tree Analysis
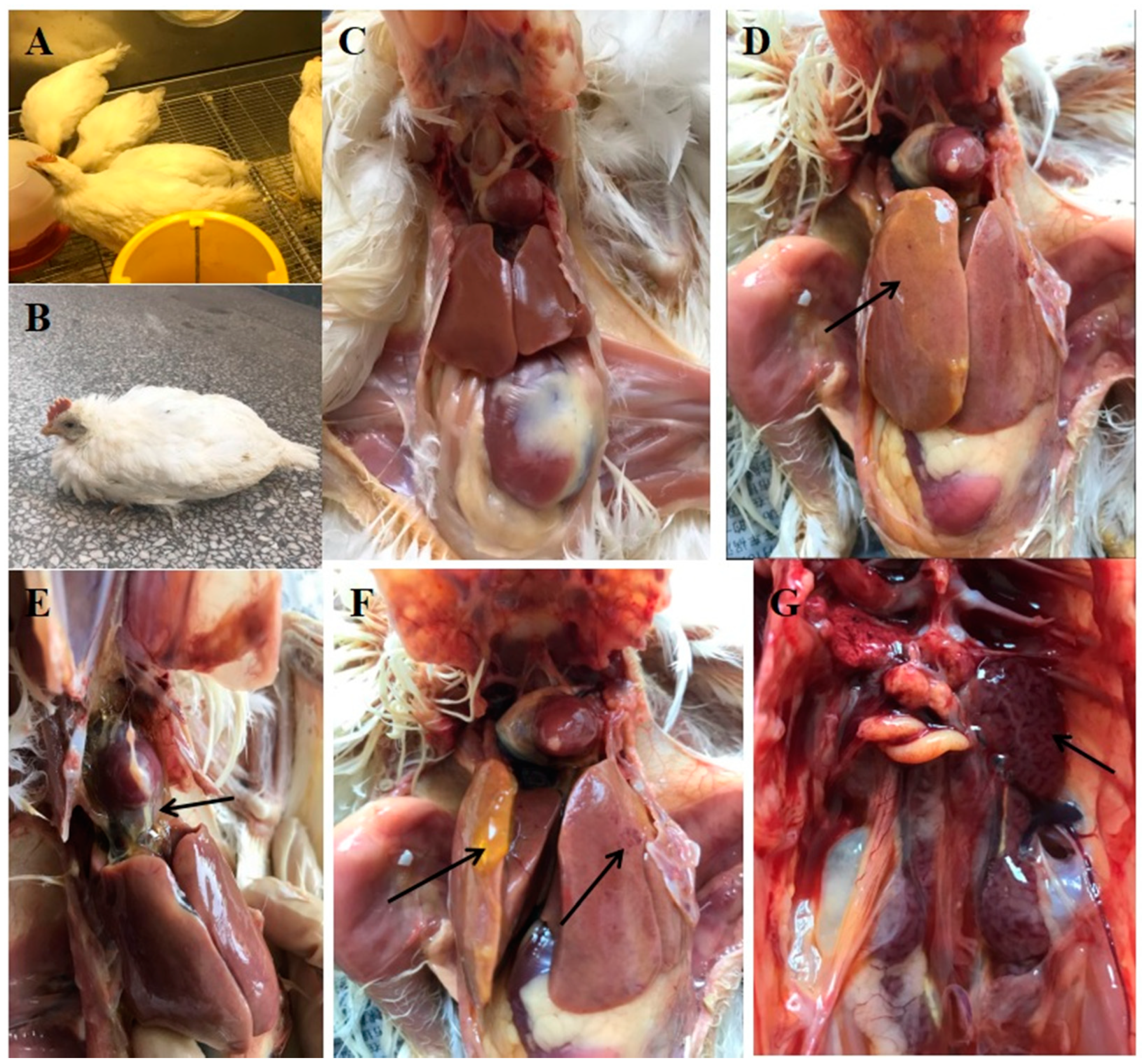
2.4. Replication and Pathogenicity of FAdV-4-SDJN0105 in Chickens
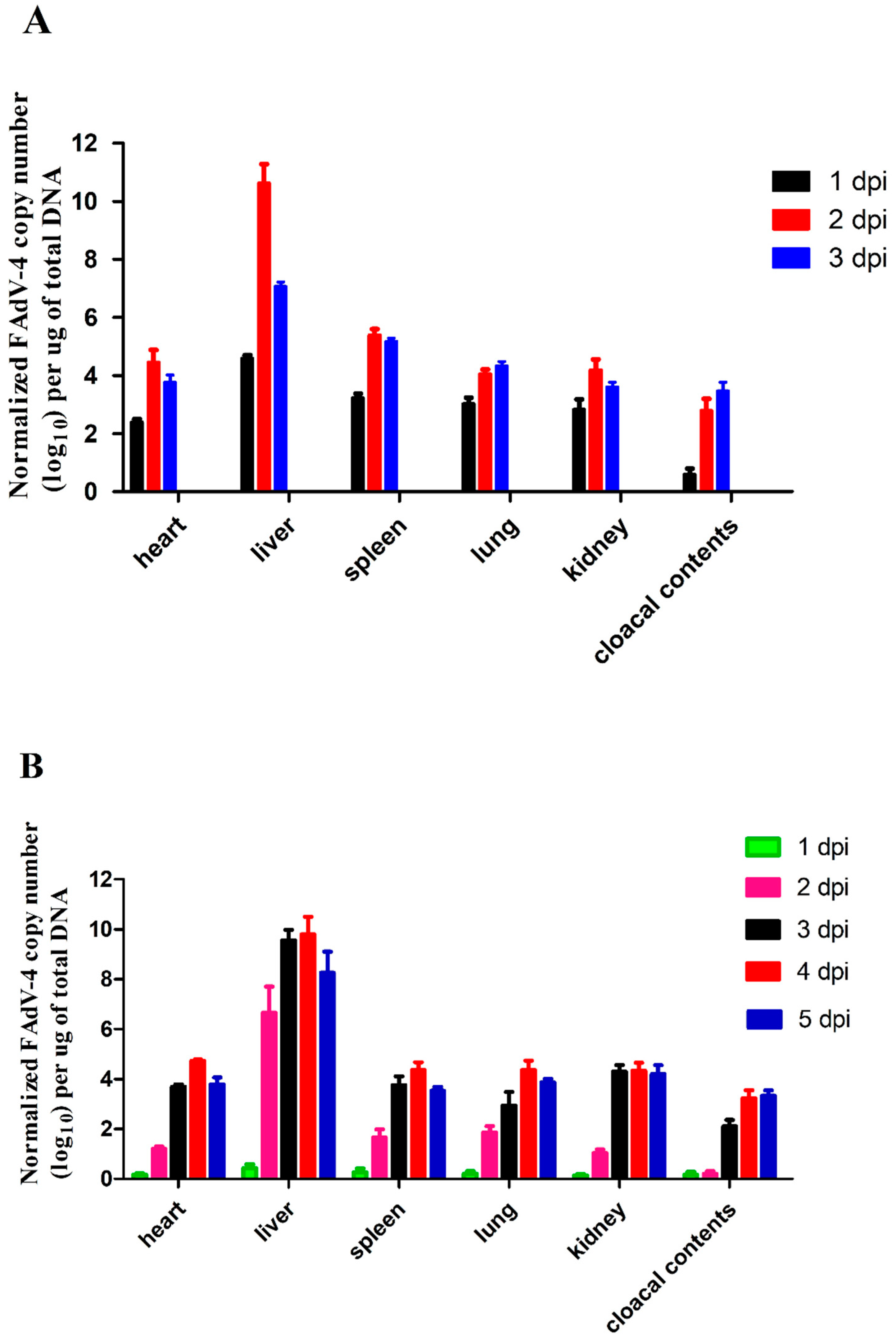
2.5. Viral Titers in Infected Chickens
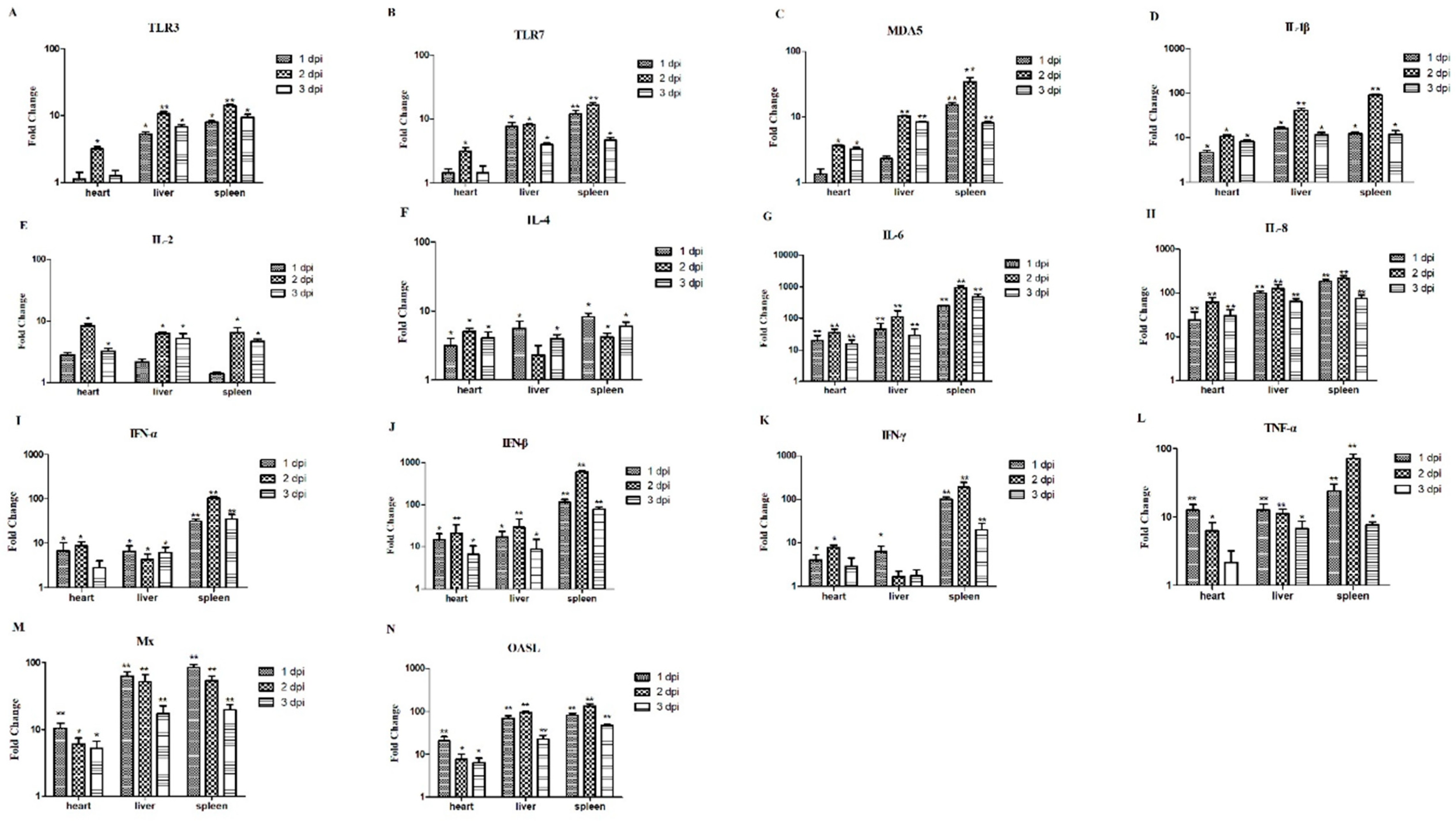
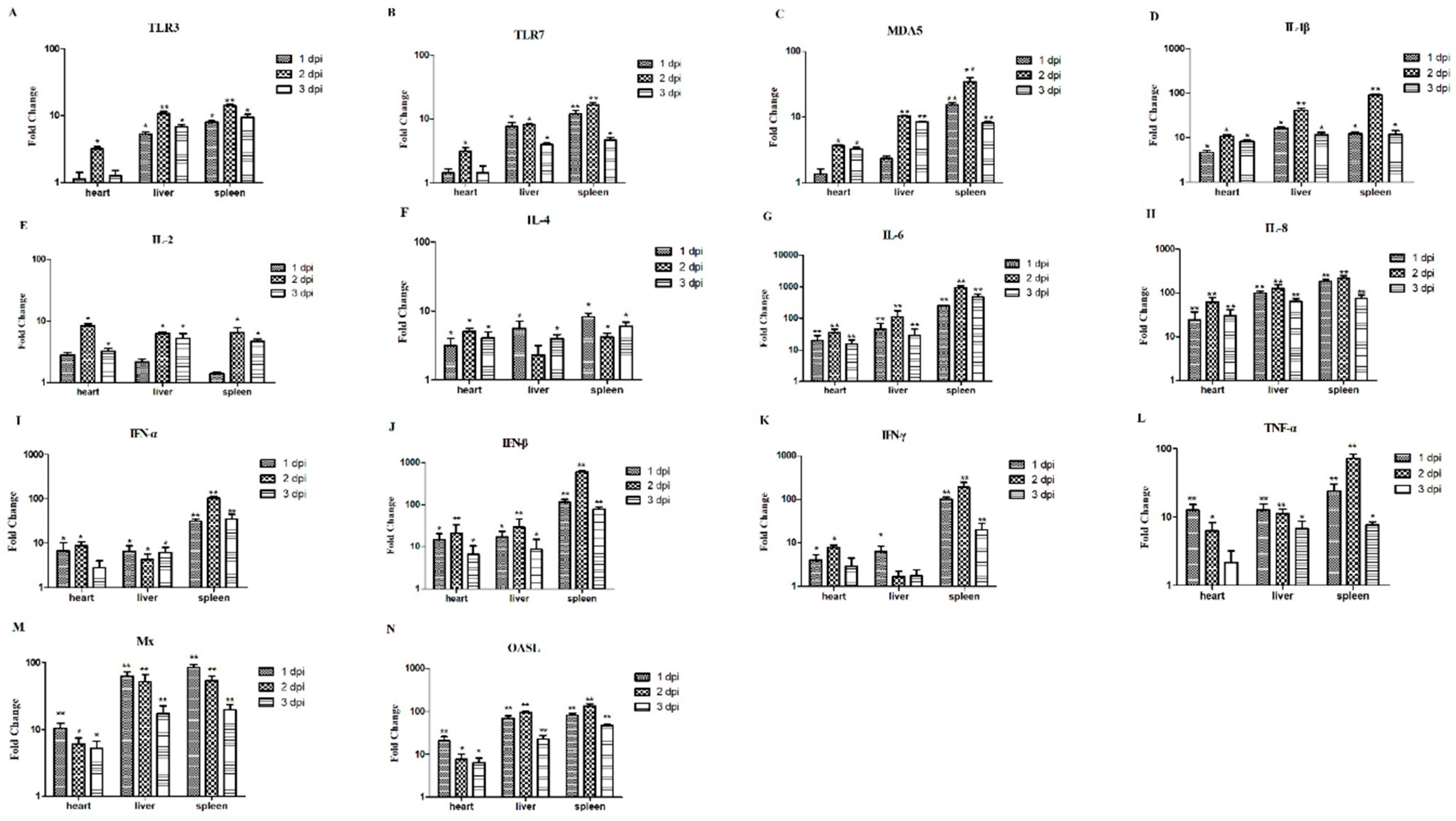
2.6. The Relative mRNA Expressions of the Hearts, Livers, and Spleens from the Infected Chickens
2.7. Preparation of Oil-Adjuvant Inactivated FAdV-4 Vaccine
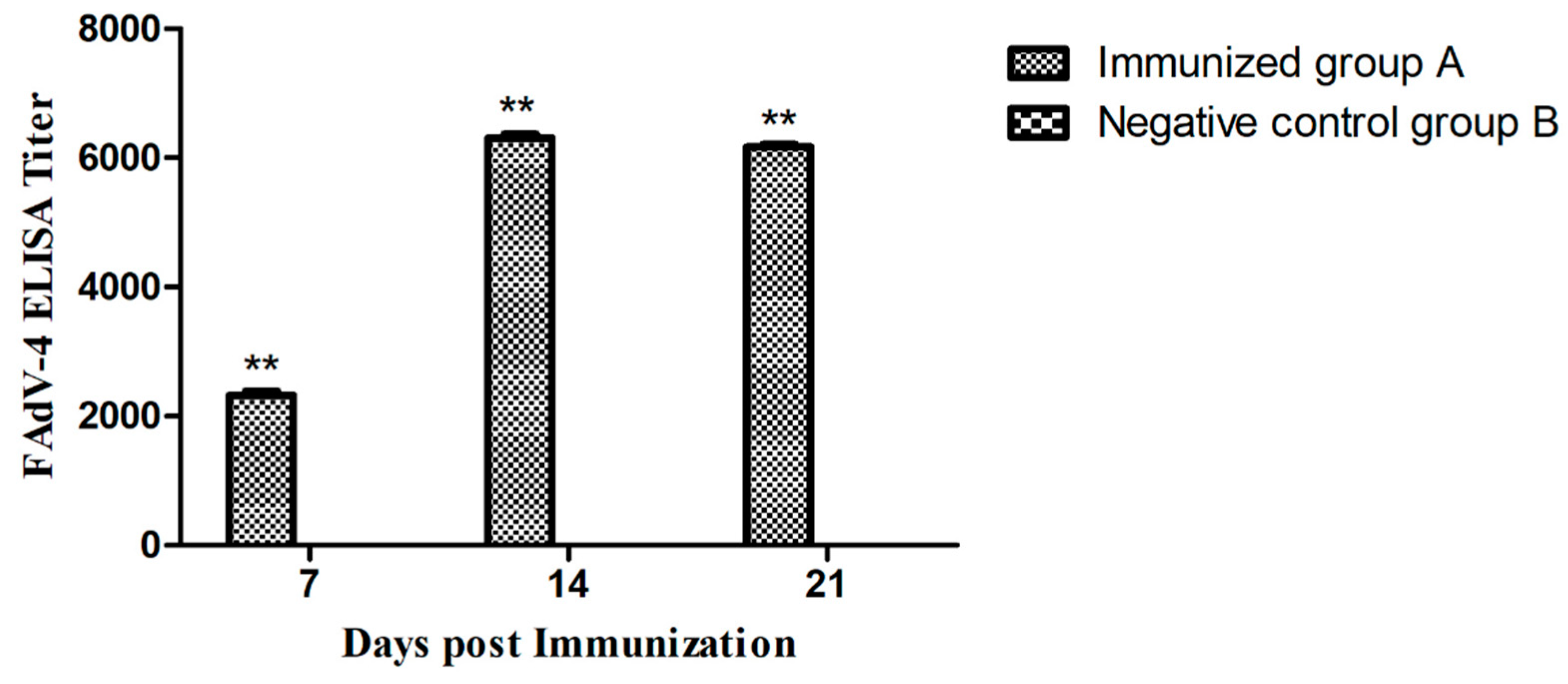
2.8. Antibody Assay
2.9. Virus Challenge Assay
2.10. Statistical Analysis
2.11. Ethics Statement
3. Results
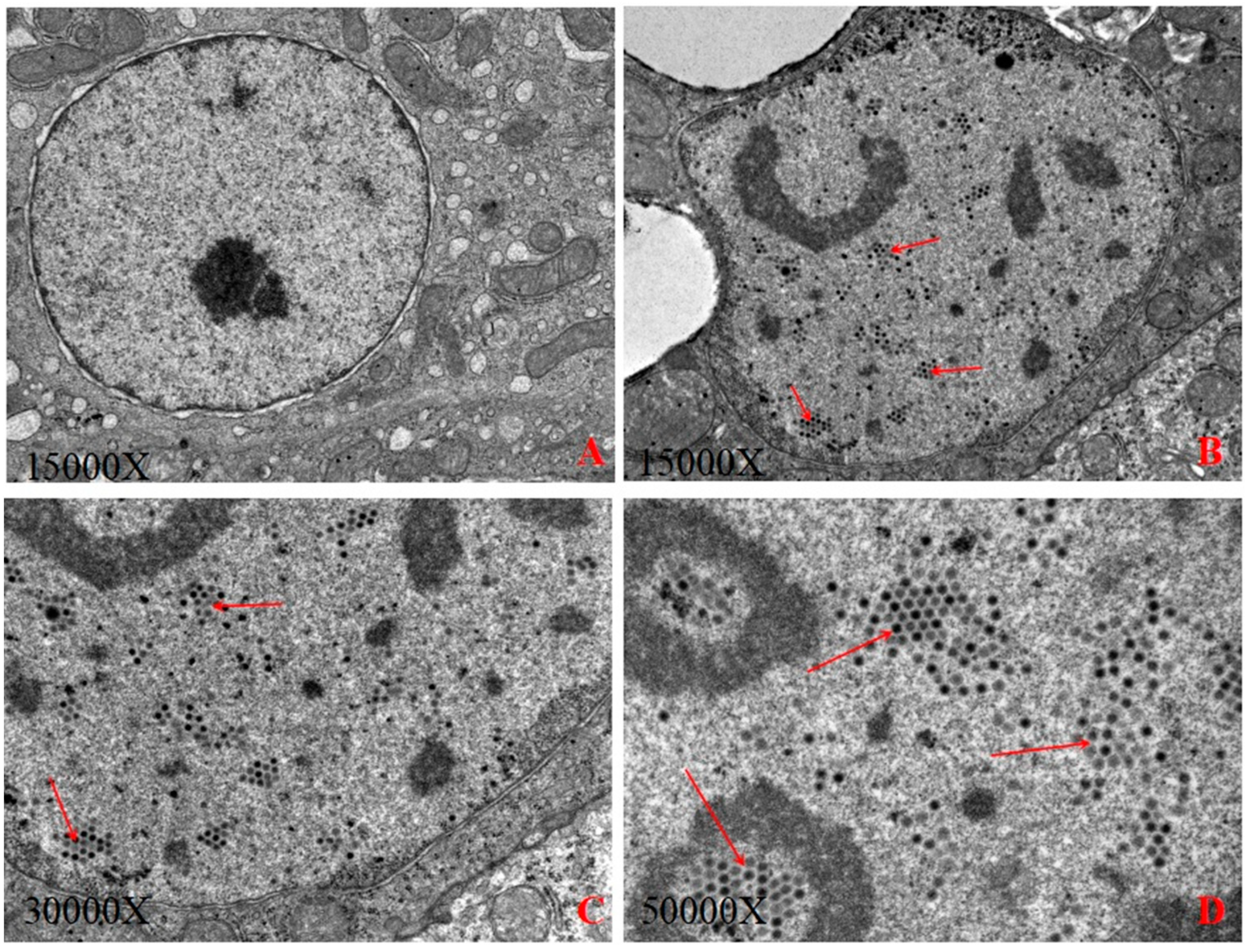
3.1. Electron Microscopic Examination
3.2. Phylogenetic and Genetic Analysis of the SDJN0105 Strains
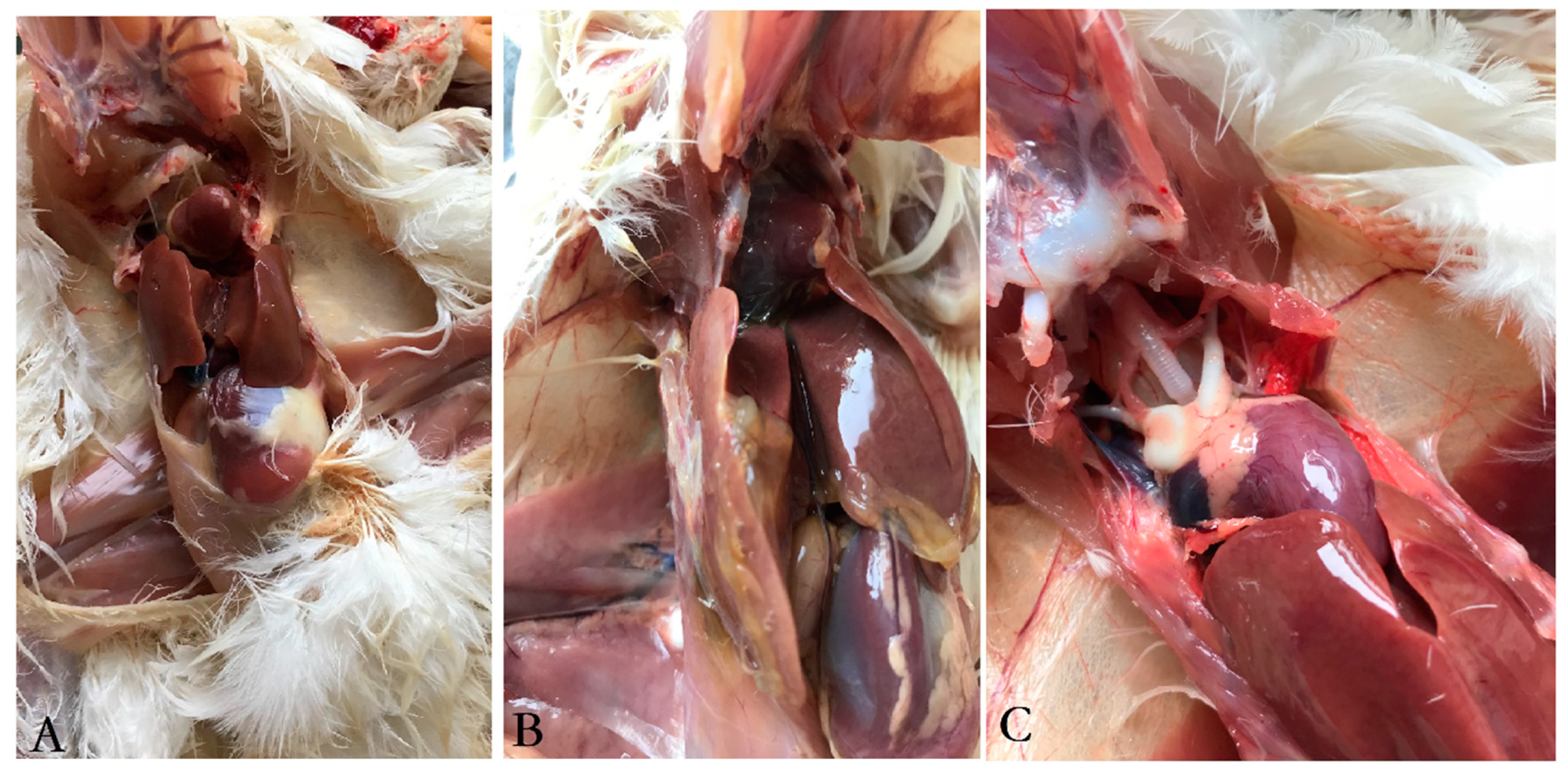
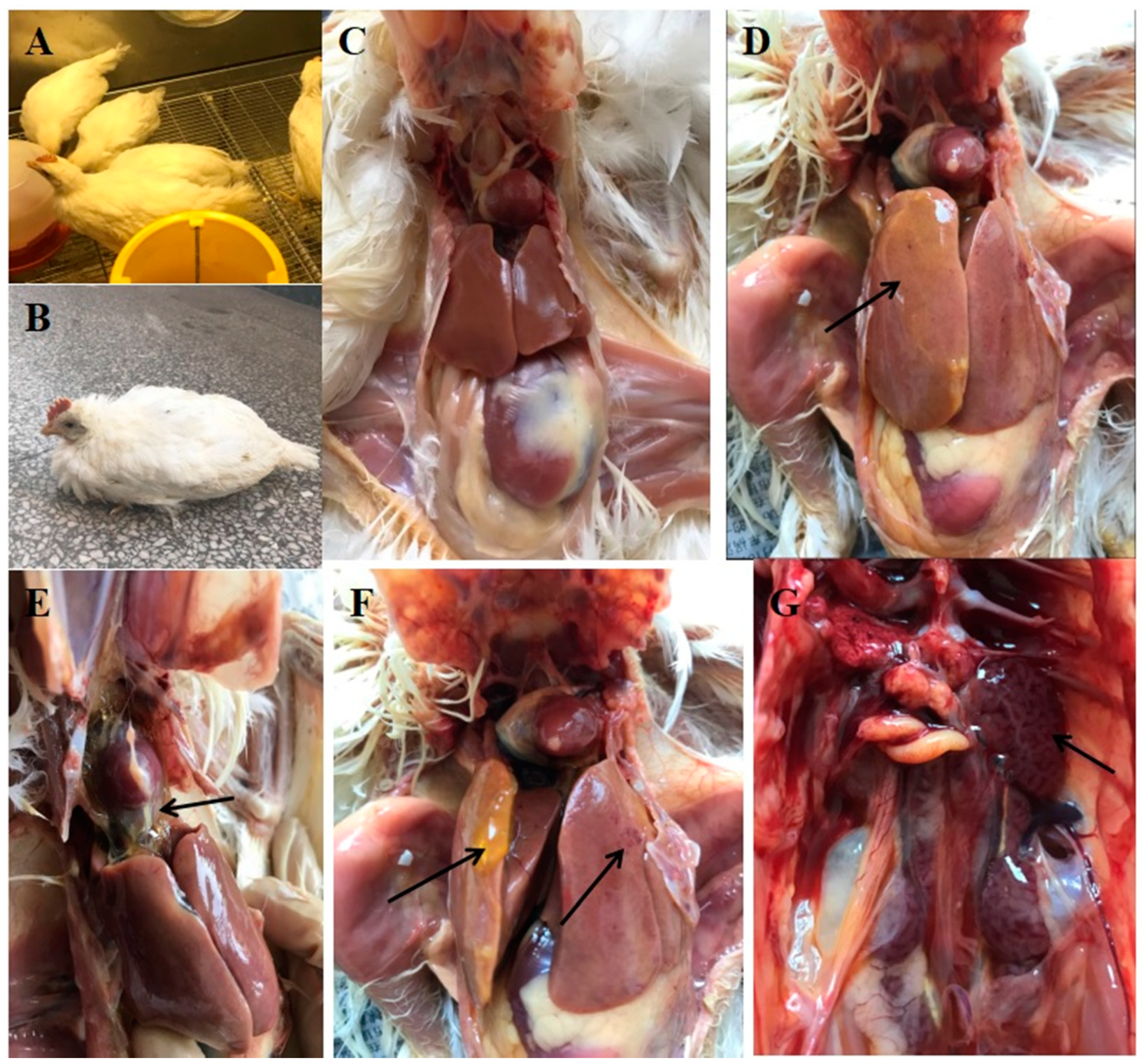
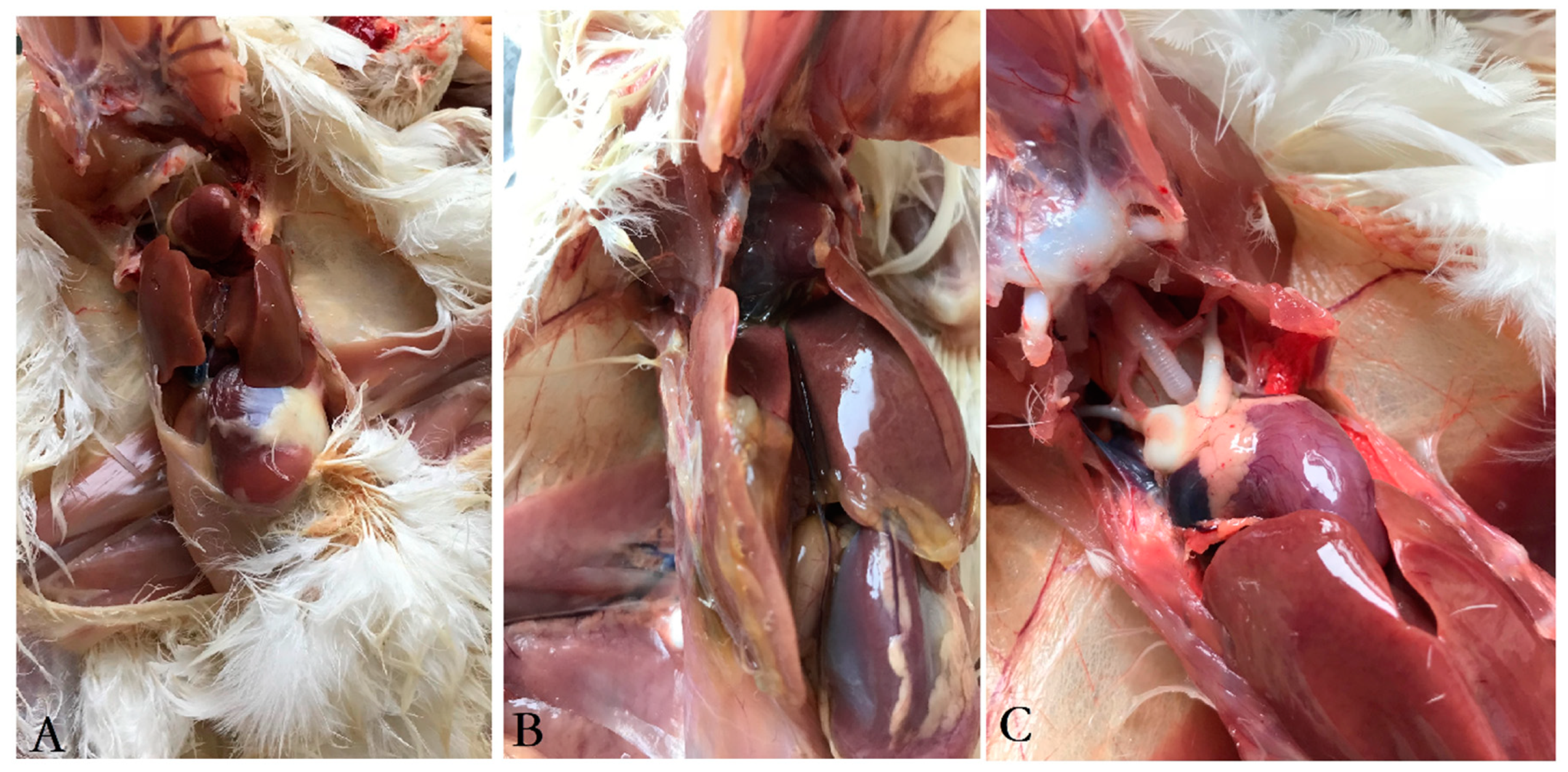
3.3. Clinical Symptoms and Gross Pathology
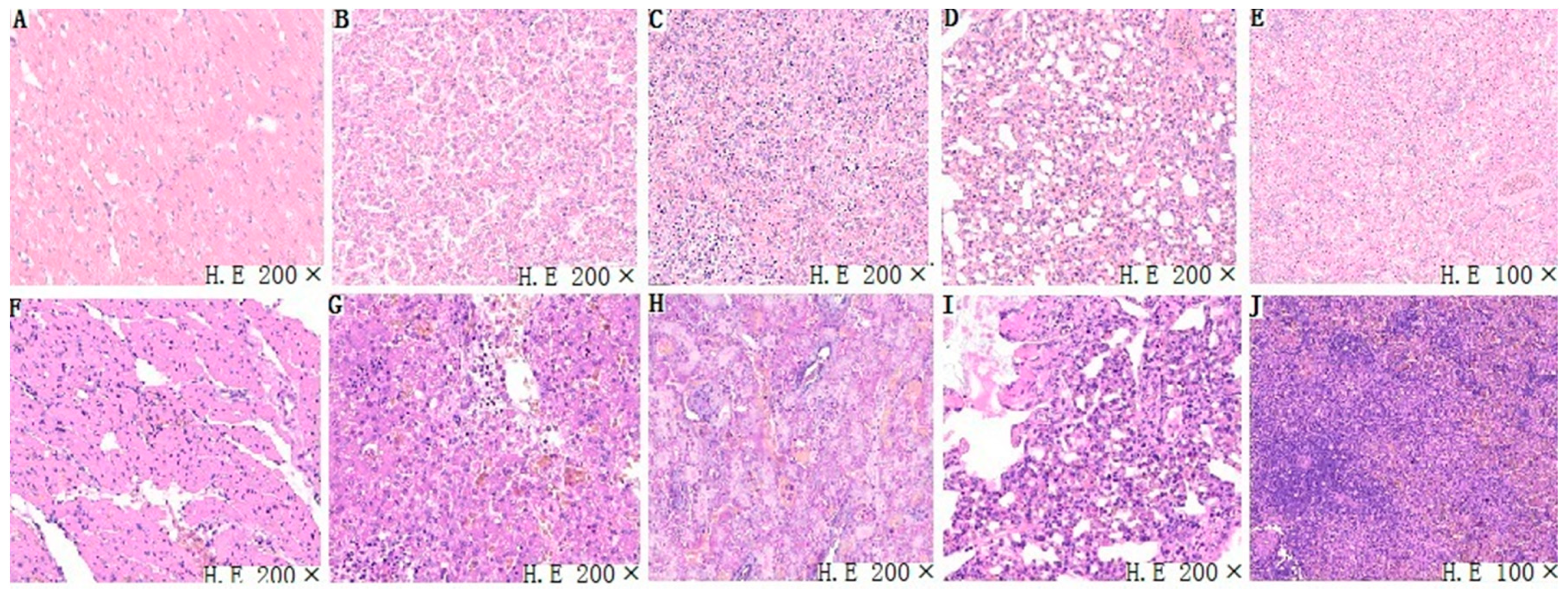
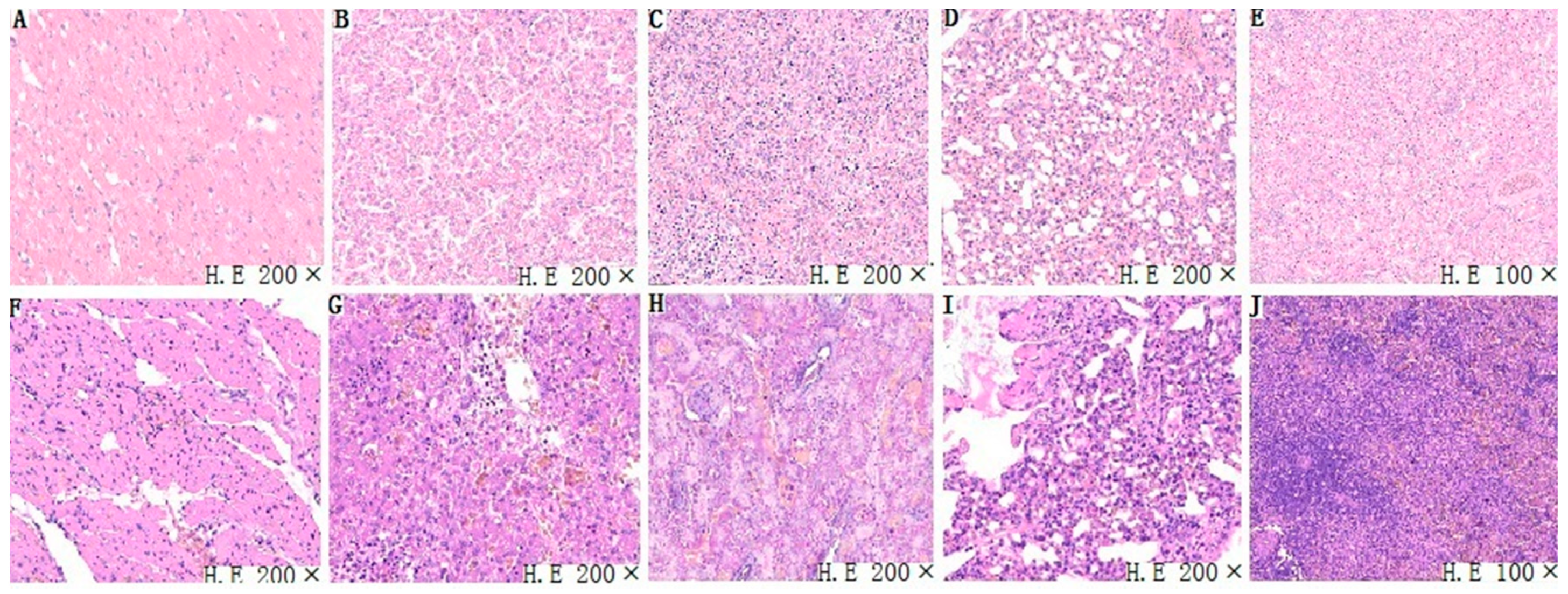
3.4. Histopathological Analysis
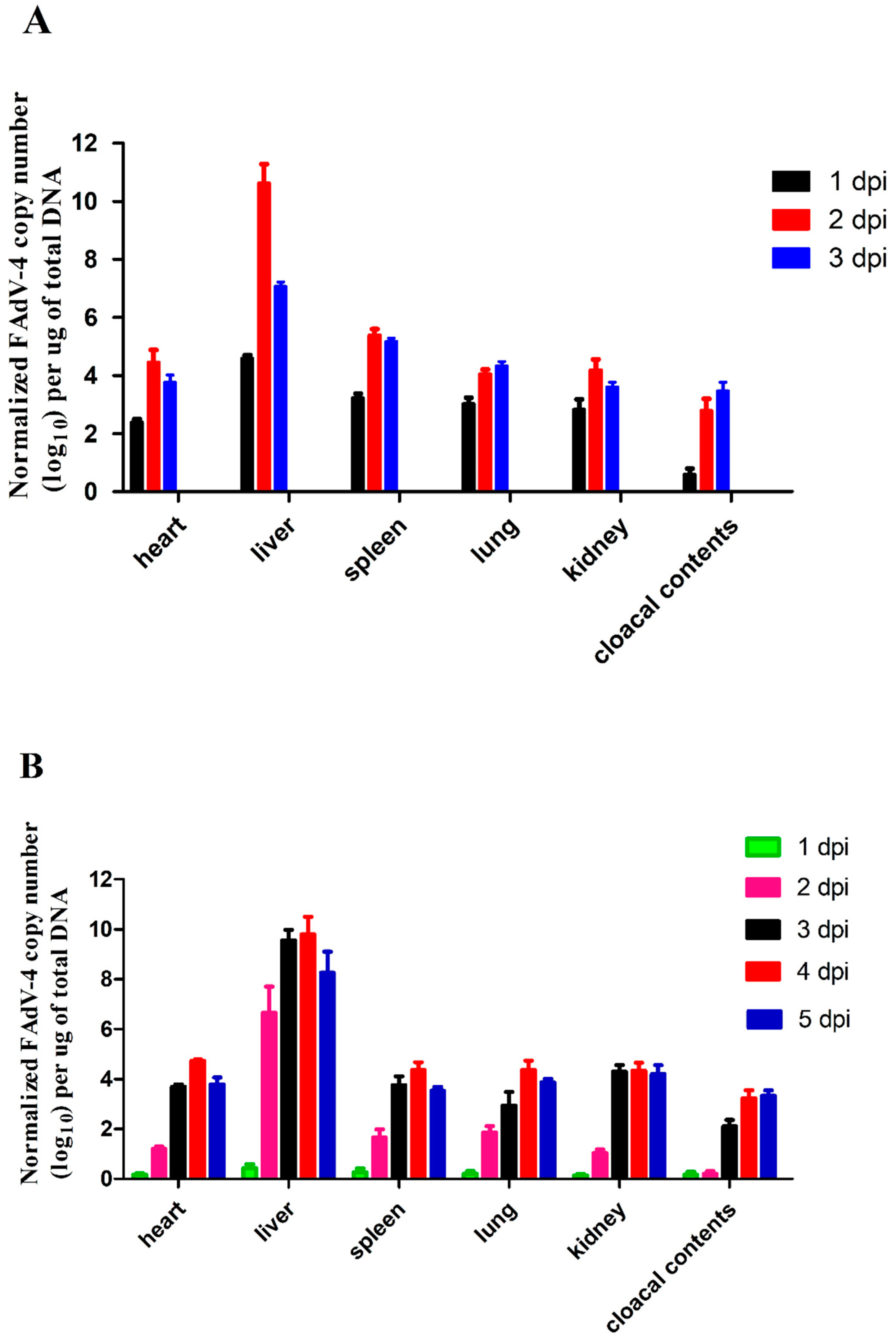
3.5. Viral Titers in Infected Chickens
3.6. The Relative mRNA Expressions of Hearts, Livers, and Spleens of the Infected Chickens
3.7. Antibody Responses of Vaccinated Chickens
3.8. Protective Efficacy of Inactivated Vaccine
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Li, L.; Ling, L.; Luo, Q.; Zhang, T.; Zhao, K.; Wang, H.; Zhang, R.; Lu, Q.; Pan, Z.; Shao, H.; et al. Genome sequence of a fowl adenovirus serotype 4 strain lethal to chickens, isolated from china. Genome Announc. 2016, 4, e00140-16. [Google Scholar] [CrossRef] [PubMed]
- Guan, R.; Tian, Y.; Han, X.; Yang, X.; Wang, H. Complete genome sequence and pathogenicity of fowl adenovirus serotype 4 involved in hydropericardium syndrome in southwest china. Microb. Pathog. 2018, 117, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Du, D.; Zhang, P.; Li, X.; Tian, H.; Cheng, Y.; Sheng, D.; Han, X.; Shan, Y.; Li, X.; Yuan, Y.; et al. Cell-culture derived fowl adenovirus serotype 4 inactivated vaccine provides complete protection for virus infection on SPF chickens. Virusdisease 2017, 28, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Jaffery, M.S. A treatise on Angara disease (hydropericardium-pulmonary oedema-hepatonephritis syndrome). J. Pak. Vet. Med. Assoc. 1988, 34, 1–33. [Google Scholar]
- Khawaja, D.A.; Ahmed, S.; Rauf, A.M.; Zulfgar, M.; Mahmood, S.M.; Hassan, M. Isolation of an adenovirus from hydropericardium syndrome in broiler chicks. Pak. J. Vet. Res. 1988, 1, 2–17. [Google Scholar]
- Ganesh, K.; Raghavan, R. Hydropericardium hepatitis syndrome of broiler poultry: Current status of research. Res. Vet. Sci. 2000, 68, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Sun, W.; Zhang, G.; Qu, Y.; Wang, P.; Sun, H.; Xiao, Y.; Liu, S. Hydropericardium syndrome outbreak caused by fowl adenovirus serotype 4 in China in 2015. J. Gen. Virol. 2016, 97, 2684–2690. [Google Scholar]
- Niu, Y.; Sun, Q.; Zhang, G.; Sun, W.; Liu, X.; Xiao, Y.; Shang, Y.; Liu, S. Pathogenicity and immunosuppressive potential of fowl adenovirus in specific pathogen free chickens. Poult. Sci. 2017, 96, 3885–3892. [Google Scholar] [CrossRef]
- Mazaheri, A.; Prusas, C.; Voss, M.; Hess, M. Some strains of serotype 4 fowl adenoviruses cause inclusion body hepatitis and hydropericardium syndrome in chickens. Avian Pathol. 1998, 27, 269–276. [Google Scholar] [CrossRef]
- Zhang, T.; Jin, Q.; Ding, P.; Wang, Y.; Chai, Y.; Li, Y.; Luo, J.; Zhang, G. Molecular epidemiology of hydropericardium syndrome outbreak-associated serotype 4 fowl adenovirus isolates in central China. Virol. J. 2016, 13, 188. [Google Scholar] [CrossRef]
- Li, C.; Li, H.; Wang, D.; Wang, J.; Wang, Y.; Wang, S.; Li, J.; Liu, P.; Wang, J.; Xu, S.; et al. Characterization of fowl adenoviruses isolated between 2007 and 2014 in China. Vet. Microbiol. 2016, 197, 62–67. [Google Scholar]
- Tidona, C.; Darai, G. The Springer Index of Viruses; Springer Science & Business Media: Berlin, Germany, 2011. [Google Scholar]
- Adair, B.M.; Fitzgerald, S.D. Group I adenovirus infections. Dis. Poult. 2008, 12, 252–266. [Google Scholar]
- Benkö, M.; Harrach, B.; Both, G.W.; Russell, W.C.; Adair, B.M.; Adám, E.; de Jong, J.C.; Hess, M.; Johnson, M.; Kajon, A.; et al. Family Adenoviridae. Virus Taxonomy: Classification and Nomenclature of Viruses; Elsevier Academic Press: Cambridge, MA, USA, 2005; pp. 95–111. [Google Scholar]
- Echavarria, M. Adenoviruses in immunocompromised hosts. Clin. Microbiol. Rev. 2008, 21, 704–715. [Google Scholar] [CrossRef] [PubMed]
- Uusikerttula, H.; Hulincurtis, S.; Davies, J.; Parker, A.L. Oncolytic adenovirus: Strategies and insights for vector design and immuno-oncolytic applications. Viruses 2015, 7, 6009–6042. [Google Scholar] [CrossRef]
- Steer, P.A.; Kirkpatrick, N.C.; O’rourke, D.; Noormohammadi, A.H. Classifcation of fowl adenovirus serotypes by use of high-resolution melting-curve analysis of the hexon gene region. J. Clin. Microbiol. 2009, 47, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Sun, Q.; Zhu, M.; Zhao, J.; Zhang, G.; Liu, X.; Xiao, Y.; Liu, S. Molecular epidemiology and phylogenetic analysis of fowl adenoviruses caused hydropericardium outbreak in China during 2015. Poult. Sci. 2018, 97, 803–811. [Google Scholar] [CrossRef]
- Pålsson-McDermott, E.M.; O’Neill, L.A. Building an immune system from nine domains. Biochem. Soc. Trans. 2007, 35, 1437–1444. [Google Scholar] [CrossRef]
- Hornung, V.; Latz, E. Intracellular DNA recognition. Nat. Rev. Immunol. 2010, 10, 123–130. [Google Scholar] [CrossRef]
- Akira, S.; Uematsu, S.; Takechi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef]
- Pan, Q.; Yang, Y.; Shi, Z.; Liu, L.; Gao, Y.; Qi, X.; Liu, C.; Zhang, Y.; Cui, H.; Wang, X. Different dynamic distribution in chickens and ducks of the hypervirulent, novel genotype fowl adenovirus serotype 4 recently emerged in China. Front. Microbiol. 2017, 8, 1005. [Google Scholar] [CrossRef]
- Ye, J.; Liang, G.; Zhang, J.; Wang, W.; Song, N.; Wang, P.; Zheng, W.; Xie, Q.; Shao, H.; Wan, Z.; et al. Outbreaks of serotype 4 fowl adenovirus with novel genotype, China. Emerg. Microbes Infect. 2016, 5, e50. [Google Scholar] [CrossRef] [PubMed]
- Ruan, S.; Zhao, J.; Ren, Y.; Feng, J.; Zhang, G. Phylogenetic analyses of Fowl Adenoviruses (FAdV) isolated in China and pathogenicity of a FAdV-8 isolate. Avian Dis. 2017, 61, 353–357. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Ruan, S.; Zhao, J.; Yang, H.; Zhang, G. Recombinant Fiber-2 protein-based indirect ELISA for antibody detection of fowl adenovirus serotype 4. Avian Dis. 2018, 62, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Li, R.; Li, G.; Lin, J.; Han, S.; Hou, X.; Weng, H.; Guo, M.; Lu, Z.; Li, N.; Shang, Y.; et al. Fowl adenovirus serotype 4 SD0828 infections causes high mortality rate and cytokine levels in specific pathogen-free chickens compared to ducks. Front. Immunol. 2018, 9, 49. [Google Scholar] [CrossRef] [PubMed]
- Hess, M. Detection and differentiation of avian adenoviruses: A review. Avian Pathol. 2000, 29, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Meulemans, G.; Couvreur, B.; Decaesstecker, M.; Boschmans, M.; van den Berg, T.P. Phylogenetic analysis of fowl adenoviruses. Avian Pathol. 2004, 33, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Lim, T.H.; Lee, D.H.; Youn, H.N.; Yuk, S.S.; Kim, B.Y.; Choi, S.W.; Jung, C.H.; Han, J.H.; Song, C.S. An inactivated oil-emulsion fowl adenovirus serotype 4 vaccine provides broad cross-protection against various serotypes of fowl adenovirus. Vaccine 2014, 32, 3564–3568. [Google Scholar] [CrossRef] [PubMed]
- Sevimli, A.; Mısırlıoğlu, D.; Yağcı, A.; Bülbül, A.; Yılmaztepe, A.; Altunbas, K. The role of chicken IL-1β, IL-6 and TNF-α in the occurrence of amyloid arthropathy. Vet. Res. Commun. 2008, 32, 499–508. [Google Scholar] [CrossRef]
- Feng, M.; Dai, M.; Cao, W.; Tan, Y.; Li, Z.; Shi, M.; Zhang, X. ALV-J strain SCAU-HN06 induces innate immune responses in chicken primary monocyte-derived macrophages. Poult. Sci. 2017, 96, 42–50. [Google Scholar] [CrossRef]
- Ferro, P.J.; Swaggerty, C.L.; He, H.; Rothwell, L.; Kaiser, P.; Kogut, M.H. Recombinant chicken IL-6 does not activate heterophils isolated from day-old chickens in vitro. Dev. Comp. Immunol. 2005, 29, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.; Bo, L.; Wang, K.; Wang, J.; He, Y.; Wang, X.; Yang, Q.; Liu, T.; Chen, D.; Geng, Y.; et al. Interleukin-8 holds promise to serve as a molecular adjuvant in DNA vaccination model against Streptococcus iniae infection in fish. Oncotarget 2016, 7, 83938–83950. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Du, F.; Wang, X. TNF-α induces two distinct caspase-8 activation pathways. Cell 2008, 133, 693–703. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Danzi, M.C.; Choi, C.S.; Taherian, M.; Dalby-Hansen, C.; Ellman, D.G.; Madsen, P.M.; Bixby, J.L.; Lemmon, V.P.; Lambertsen, K.L.; et al. Opposing functions of microglial and macrophagic TNFR2 in the pathogenesis of experimental autoimmune Encephalomyelitis. Cell Rep. 2017, 18, 198–212. [Google Scholar] [CrossRef] [PubMed]
- Bhargava, R.; Janssen, W.; Altmann, C.; Andrés-Hernando, A.; Okamura, K.; Vandivier, R.W.; Ahuja, N.; Faubel, S. Intratracheal IL-6 protects against lung inflammation in direct, but not indirect, causes of acute lung injury in mice. PLoS ONE 2013, 8, e61405. [Google Scholar] [CrossRef] [PubMed]
- Pan, Q.; Yang, Y.; Gao, Y.; Qi, X.; Liu, C.; Zhang, Y.; Cui, H.; Wang, X. An inactivated novel genotype fowl adenovirus 4 protects chickens against the hydropericardium syndrome that recently emerged in China. Viruses 2017, 9, 216. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | GenBank Numbers | Serotype |
|---|---|---|
| FAdV-A | MF168407 | FAdV-1 |
| KX247011 | FAdV-1 | |
| KX247012 | FAdV-1 | |
| MF198255 | FAdV-1 | |
| FAdV-B | AY683544 | FAdV-3 |
| NC_021221 | FAdV-5 | |
| FAdV-C | EU177545 | FAdV-4 |
| EU931691 | FAdV-4 | |
| EU938324 | FAdV-4 | |
| KU647684 | FAdV-4 | |
| KT899325 | FAdV-4 | |
| KT717889 | FAdV-10 | |
| FAdV-D | KT862805 | FAdV-2 |
| KT862806 | FAdV-2 | |
| KF426983 | FAdV-2 | |
| KU746335 | FAdV-11 | |
| KU310942 | FAdV-11 | |
| KP231537 | FAdV-11 | |
| FAdV-E | NC_038332 | FAdV-6 |
| KT862808 | FAdV-6 | |
| KY364398 | FAdV-7 | |
| KY968968 | FAdV-8 | |
| KU517714 | FAdV-8b | |
| KY426984 | FAdV-8b | |
| DAdV | EF093507 | DAdV-1 |
| Primer Name | Sequence of Oligonucleotide (5′–3′) | Size (bp) |
|---|---|---|
| q-FAdV-4-F | CGTCCCGACTACCCTAGAGA | 150 |
| q-FAdV-4-R | AGGTCCCGCAACTGAGACT | |
| β-actin-F | ATGAAGCCCAGAGCAAAAGA | 120 |
| β-actin-R | GGGGTGTTGAAGGTCTCAAA | |
| TLR3-F | ACAATGGCAGATTGTAGTCACCT | 123 |
| TLR3-R | GCACAATCCTGGTTTCAGTTTAG | |
| TLR7-F | TGTGATGTGGAAGCCTTTGA | 185 |
| TLR7-R | ATTATCTTTGGGCCCCAGTC | |
| MDA5-F | TGAAAGCCTTGCAGATGACTTA | 120 |
| MDA5-R | GCTGTTTCAAATCCTCCGTTAC | |
| TNF-α-F | CCGCCCAGTTCAGATGAGTT | 130 |
| TNF-α-R | GCAACAACCAGCTATGCACC | |
| IFN-α-F | CACCTTCCTCCAAGACAACGATT | 108 |
| IFN-α-R | GTGCGAGTGATAAATGTGAGGTTG | |
| IFN-β-F | CCTCAACCAGATCCAGCATT | 180 |
| IFN-β-R | GGATGAGGCTGTGAGAGGAG | |
| IFN-γ-F | TGAGCCAGATTGTTTCGATG | 186 |
| IFN-γ-R | CTTGGCCAGGTCCATGATA | |
| IL-1β-F | CCAGAAAGTGAGGCTCAACATTG | 114 |
| IL-1β-R | GACATGTAGAGCTTGTAGCCCTT | |
| IL-2-F | TACCTGGGAGAAGTGGTTACTCT | 117 |
| IL-2-R | TAGACCCGTAAGACTCTTGAGGT | |
| IL-4-F | AGAGCATCCGGATAGTGAATGAC | 142 |
| IL-4-R | AGATCGATACCAGTCTGAGCAAC | |
| IL-6-F | CCTGTTCGCCTTTCAGACCT | 171 |
| IL-6-R | GGGATGACCACTTCATCGGG | |
| IL-8-F | ATTCAAGATGTGAAGCTGAC | 196 |
| IL-8-R | AGGATCTGCAATTAACATGAGG | |
| Mx-F | CAGCTCCAGAATGCATCAGA | 181 |
| Mx-R | GGCAATTCCAGGAAGATCAA | |
| OASL-F | GAGATGGAGGTCCTGGTGAA | 152 |
| OASL-R | CCAGCTCCTTGGTCTCGTAG |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meng, K.; Yuan, X.; Yu, J.; Zhang, Y.; Ai, W.; Wang, Y. Identification, Pathogenicity of Novel Fowl Adenovirus Serotype 4 SDJN0105 in Shandong, China and Immunoprotective Evaluation of the Newly Developed Inactivated Oil-emulsion FAdV-4 Vaccine. Viruses 2019, 11, 627. https://doi.org/10.3390/v11070627
Meng K, Yuan X, Yu J, Zhang Y, Ai W, Wang Y. Identification, Pathogenicity of Novel Fowl Adenovirus Serotype 4 SDJN0105 in Shandong, China and Immunoprotective Evaluation of the Newly Developed Inactivated Oil-emulsion FAdV-4 Vaccine. Viruses. 2019; 11(7):627. https://doi.org/10.3390/v11070627
Chicago/Turabian StyleMeng, Kai, Xiaoyuan Yuan, Jiang Yu, Yuxia Zhang, Wu Ai, and Youling Wang. 2019. "Identification, Pathogenicity of Novel Fowl Adenovirus Serotype 4 SDJN0105 in Shandong, China and Immunoprotective Evaluation of the Newly Developed Inactivated Oil-emulsion FAdV-4 Vaccine" Viruses 11, no. 7: 627. https://doi.org/10.3390/v11070627
APA StyleMeng, K., Yuan, X., Yu, J., Zhang, Y., Ai, W., & Wang, Y. (2019). Identification, Pathogenicity of Novel Fowl Adenovirus Serotype 4 SDJN0105 in Shandong, China and Immunoprotective Evaluation of the Newly Developed Inactivated Oil-emulsion FAdV-4 Vaccine. Viruses, 11(7), 627. https://doi.org/10.3390/v11070627