Silent Circulation of the Saint Louis Encephalitis Virus among Humans and Equids, Southeast Brazil

 , , , ,
, , , ,  , and
, and 
Abstract
:1. Introduction
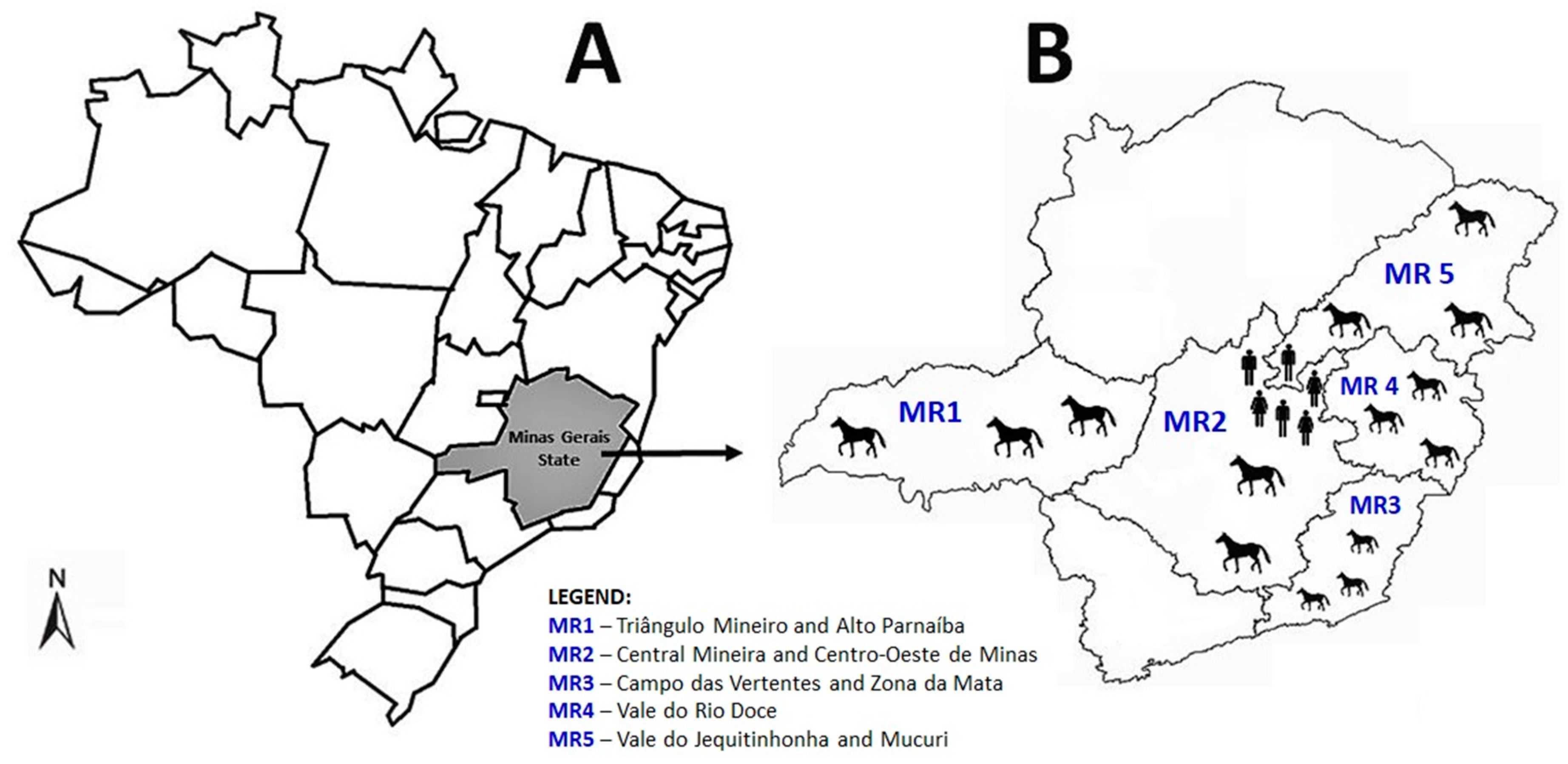
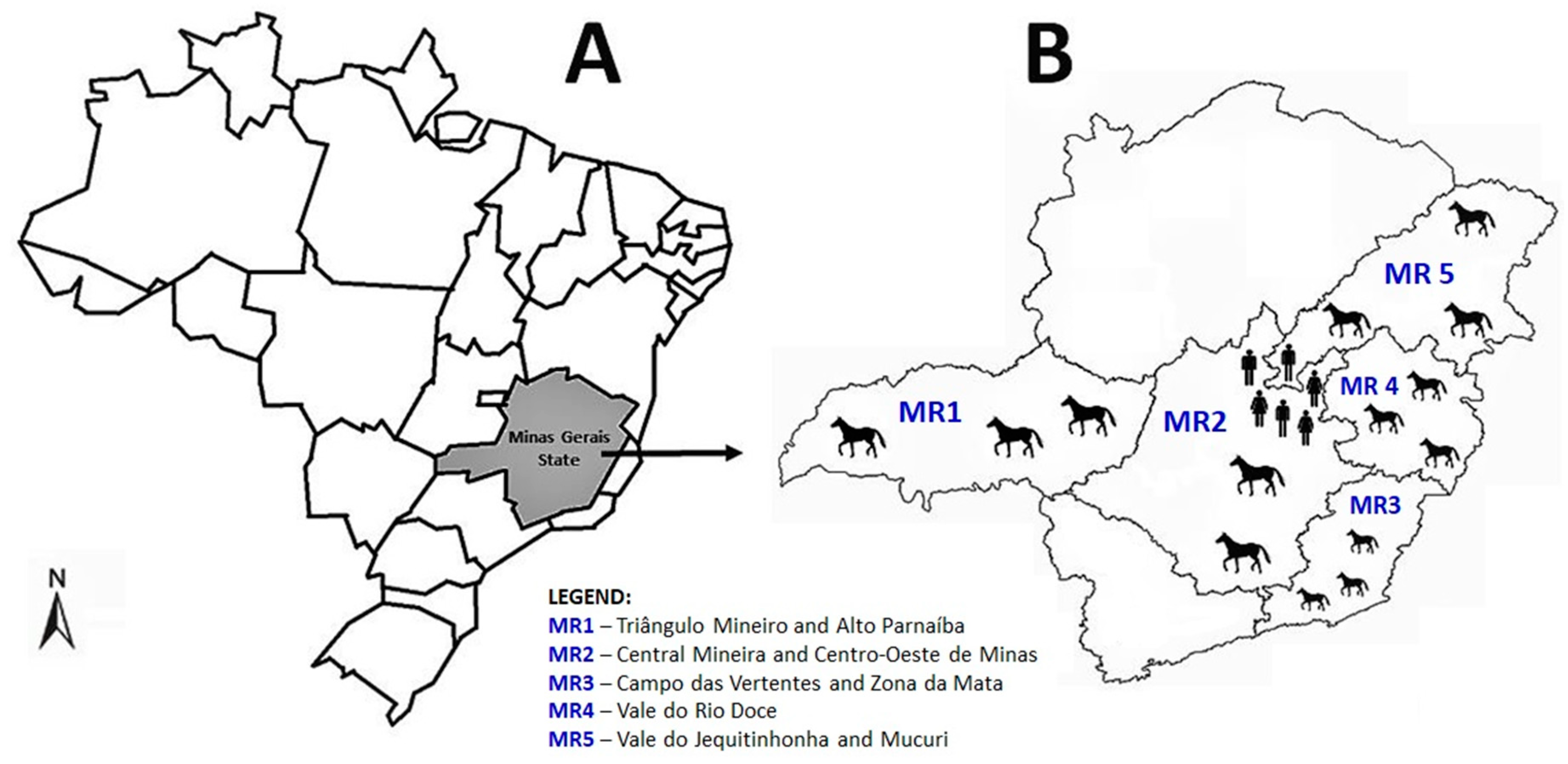
2. Materials and Methods
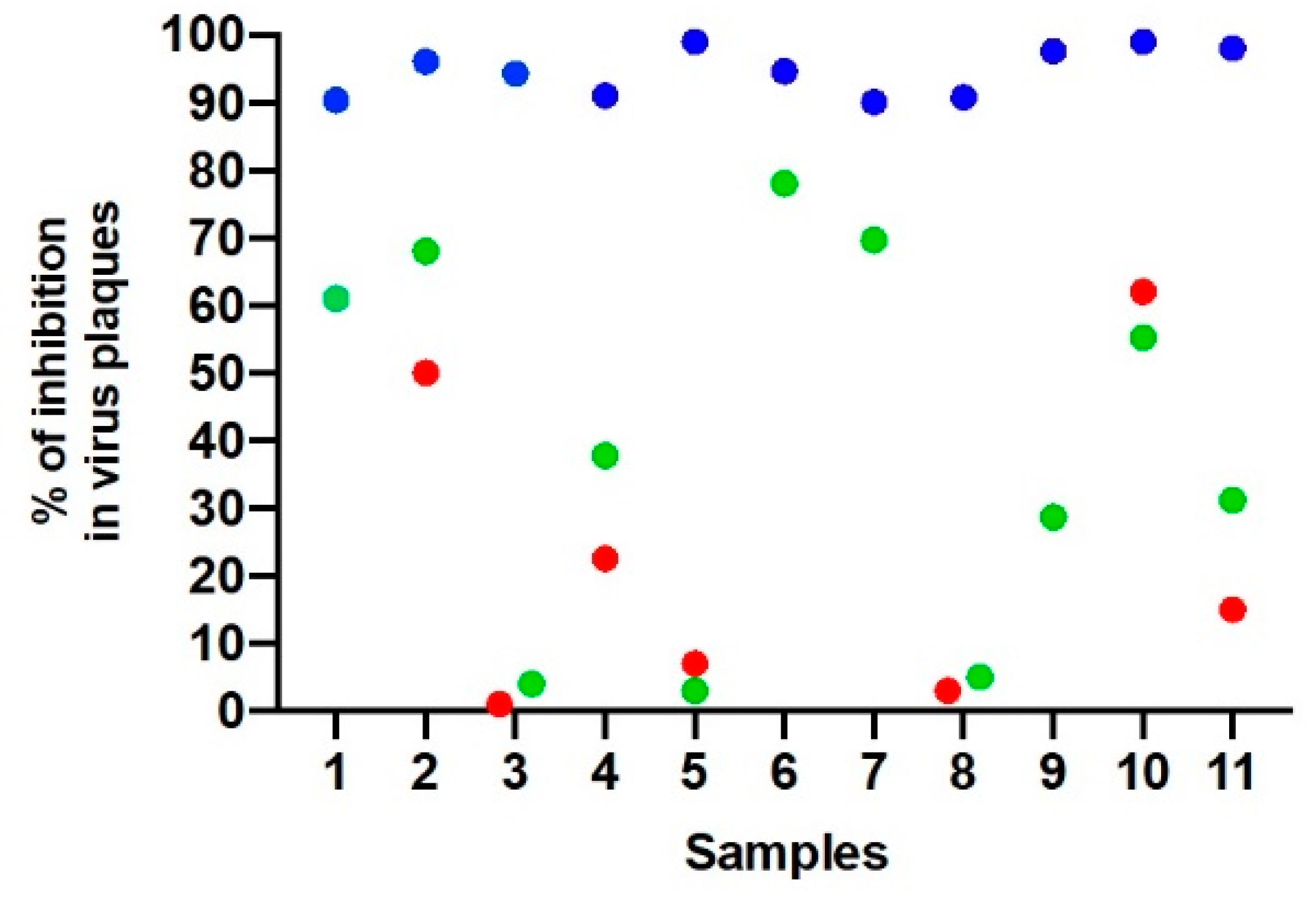
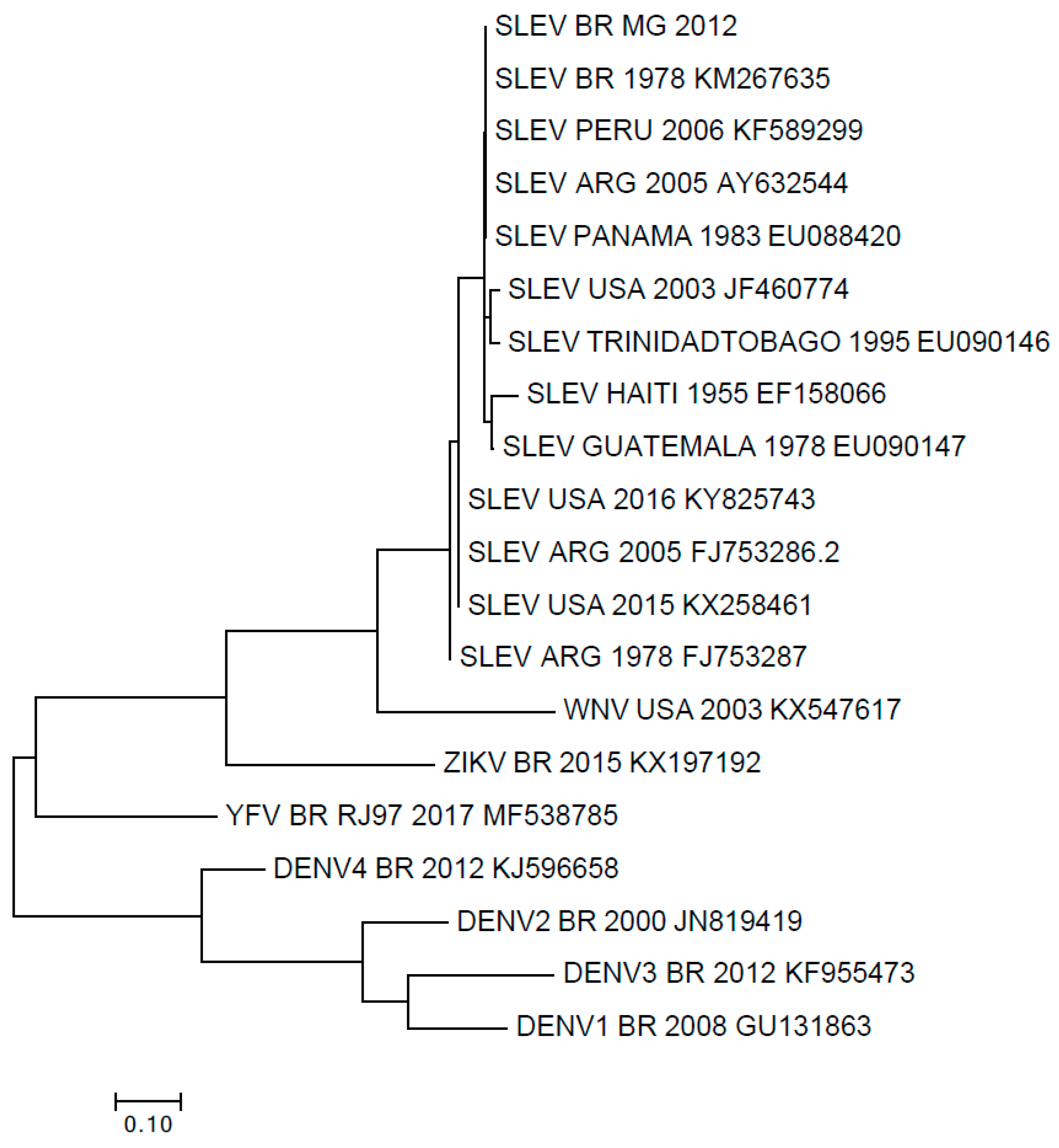
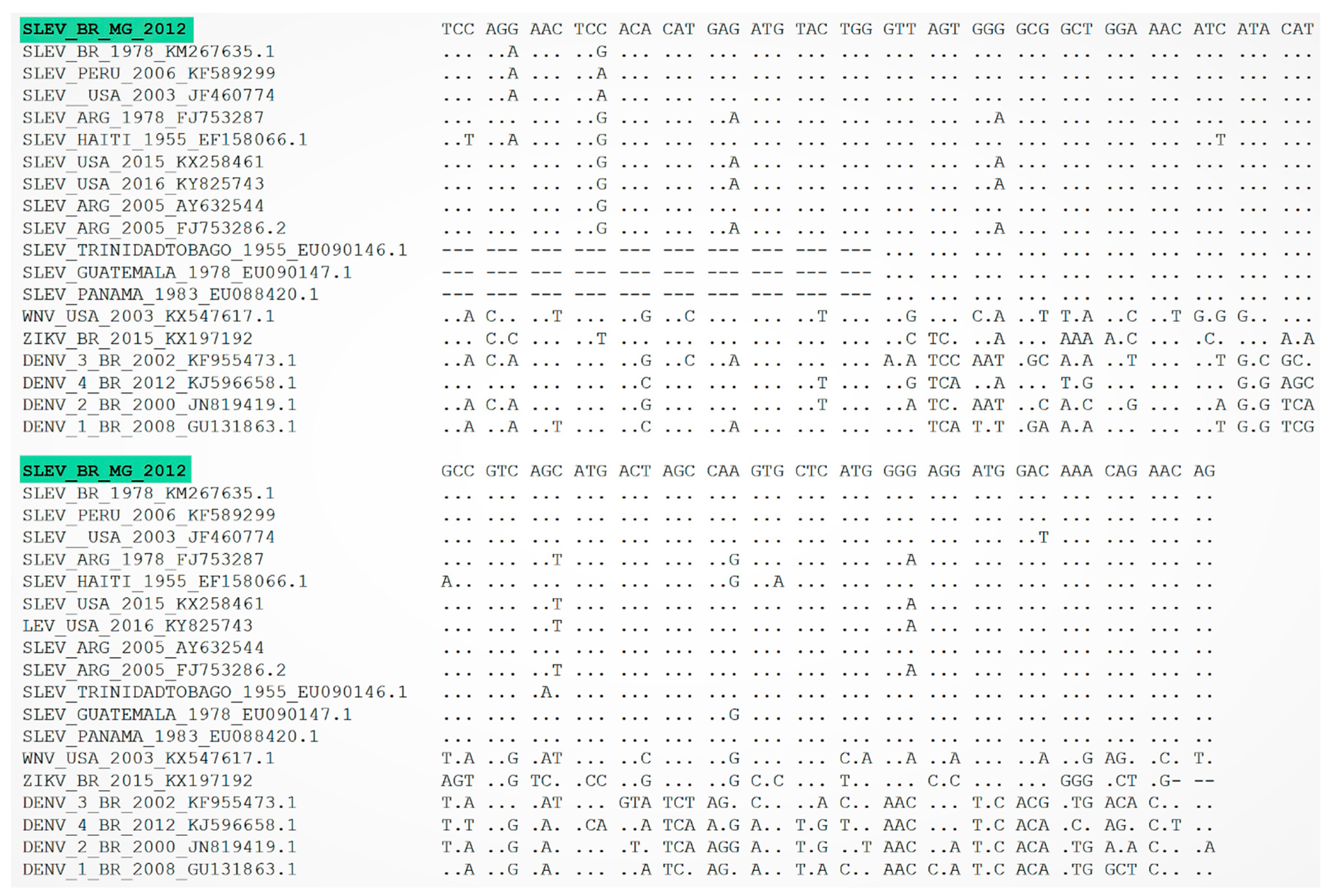
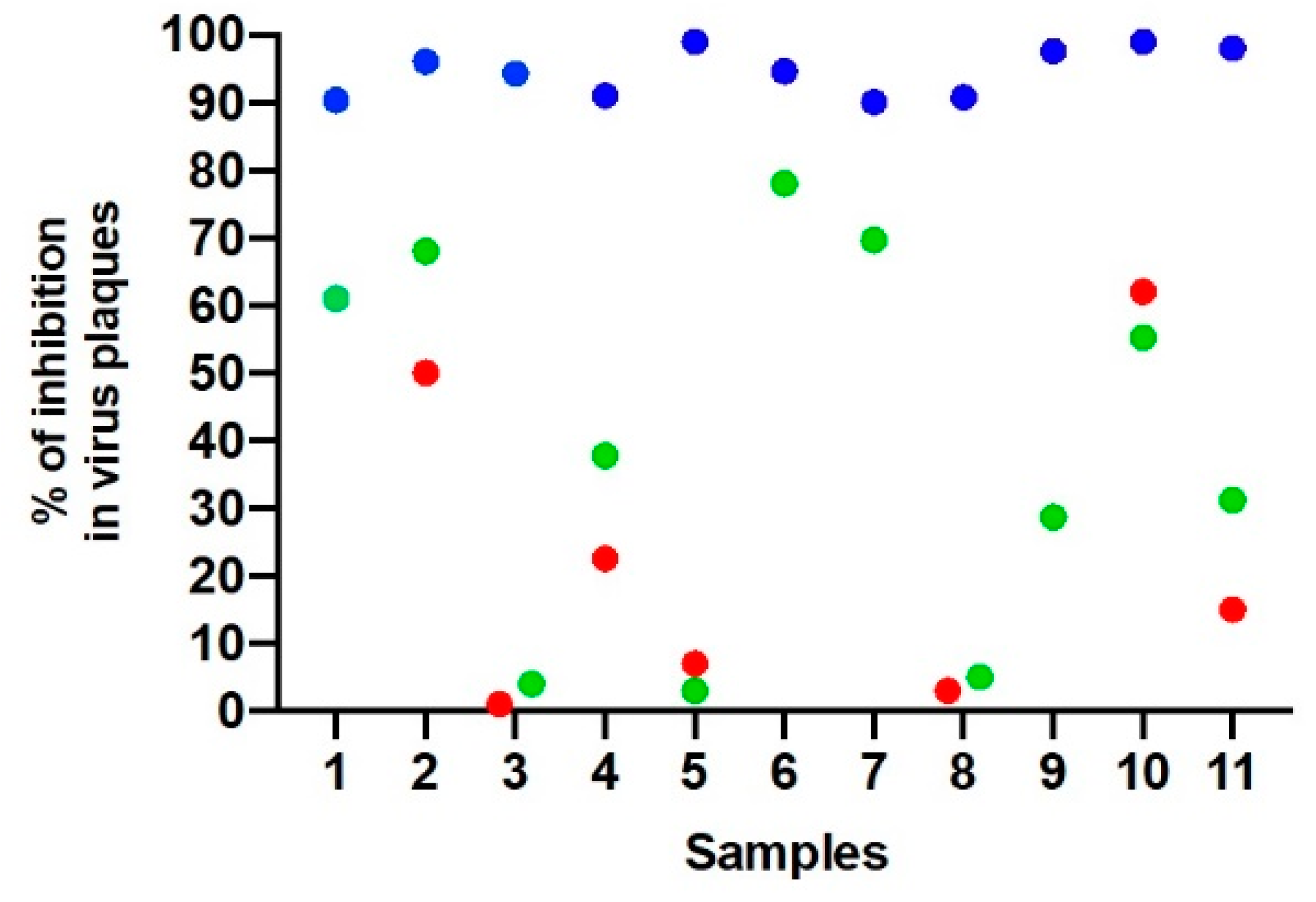
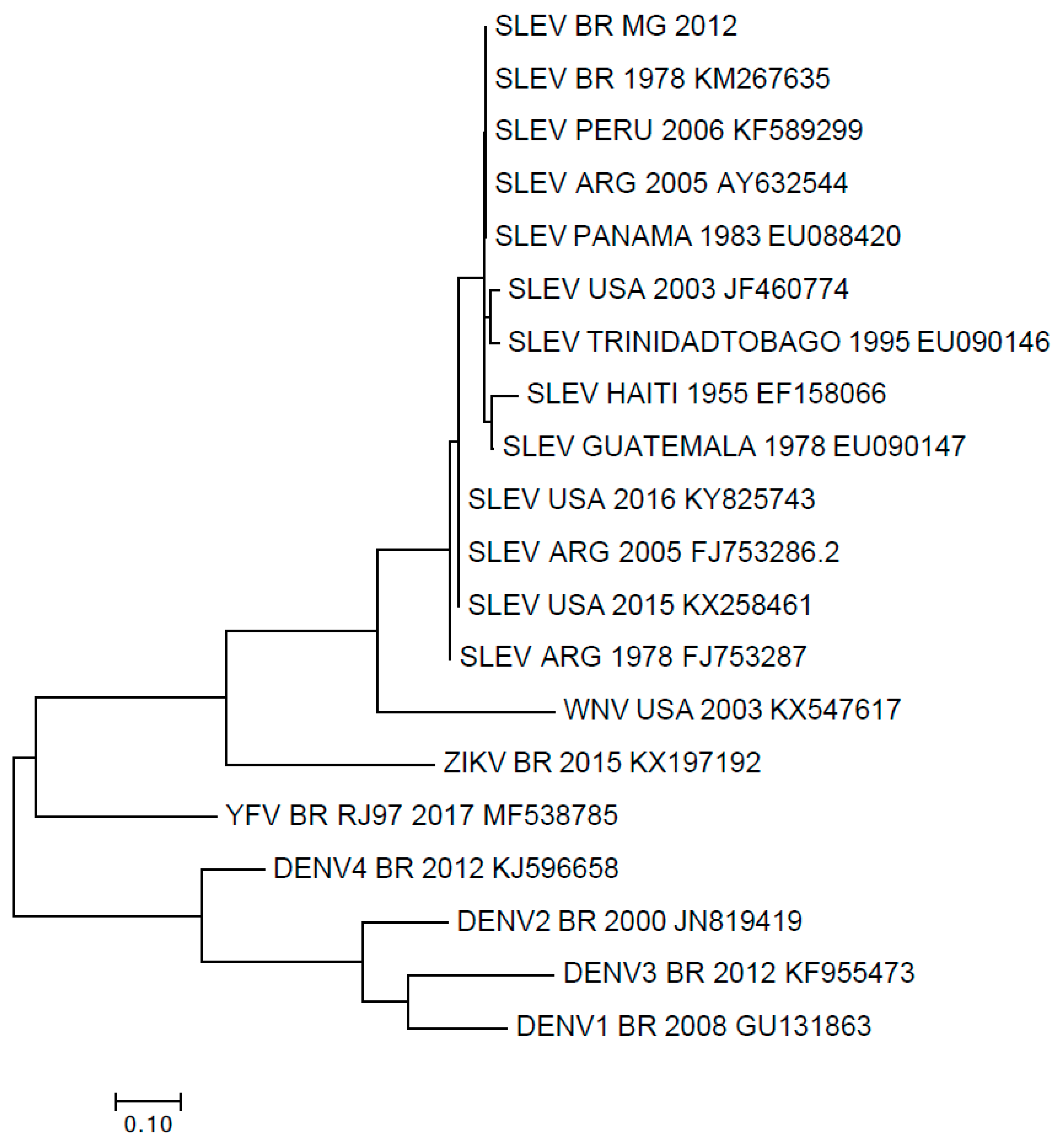
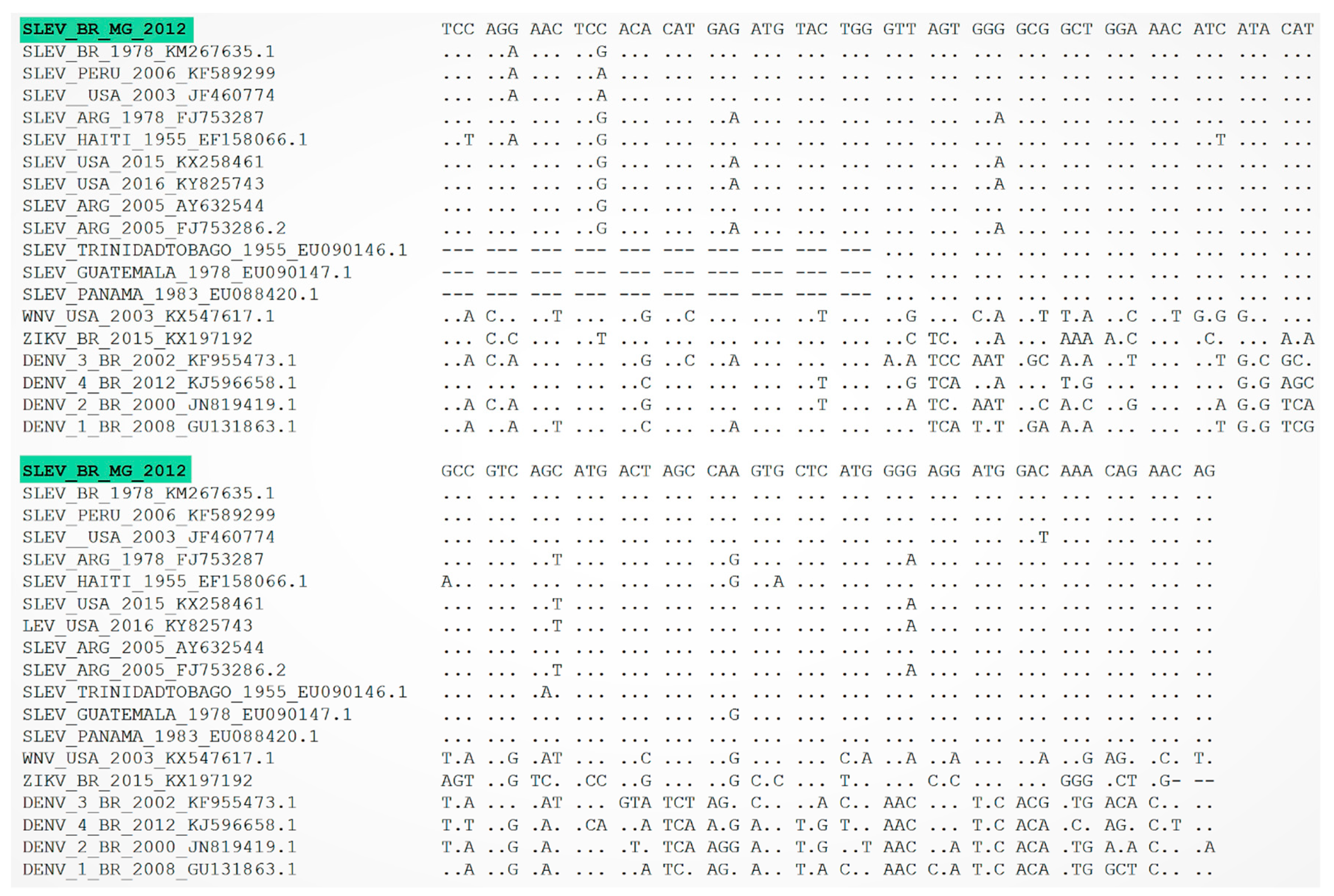
3. Results
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Diaz, A.; Coffey, L.L.; Burkett-Cadena, N.; Day, J.F. Reemergence of St. Louis Encephalitis Virus in the Americas. Emerg. Infect. Dis. 2018, 24, 2150–2157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Centers for Disease Control and Prevention. Saint Louis Encephalitis. Available online: https://www.cdc.gov/sle/index.html (accessed on 23 August 2019).
- Reisen, W.K. Epidemiology of St. Louis encephalitis virus. Adv. Virus Res. 2003, 61, 139–183. [Google Scholar] [PubMed]
- Pierson, T.C.; Diamond, M.S. Flaviviruses. In Fields Virology; Knipe, D.M., Howley, P.P., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; pp. 747–794. [Google Scholar]
- Mondini, A.; Bronzoni, R.V.; Cardeal, I.L.; dos Santos, T.M.; Lázaro, E.; Nunes, S.H.; Nogueira, M.L. Simultaneous infection by DENV-3 and SLEV in Brazil. J. Clin. Virol. 2007, 40, 84–86. [Google Scholar] [CrossRef] [PubMed]
- Mondini, A.; Cardeal, I.L.; Lázaro, E.; Nunes, S.H.; Moreira, C.C.; Rahal, P.; Nogueira, M.L. Saint Louis encephalitis virus, Brazil. Emerg. Infect. Dis. 2007, 13, 176–178. [Google Scholar] [CrossRef] [PubMed]
- Terzian, C.; Mondini, A.; Bronzoni, R.V.; Drumond, B.P.; Ferro, B.P.; Cabrera, E.M.; Figueiredo, L.T.; Chiaravalloti-Neto, F.; Nogueira, M.L. Detection of Saint Louis encephalitis virus in dengue-suspected cases during a dengue 3 outbreak. Vector Borne Zoonotic Dis. 2011, 11, 291–300. [Google Scholar] [CrossRef]
- Heinen, L.B.; Zuchi, N.; Serra, O.P.; Cardoso, B.F.; Gondim, B.H.; Dos Santos, M.A.; Souto, F.J.; Paula, D.A.; Dutra, V.; Dezengrini-Slhessarenko, R. Saint Louis encephalitis virus in Mato Grosso, Central-Western, Brazil. Rev. Inst. Med. Trop. Sao Paulo. 2015, 57, 215–220. [Google Scholar] [CrossRef]
- Díaz, L.A.; Linás, G.A.; Vázques, A.; Tenorio, A.; Contigiani, M.S. Silent Circulation of St. Louis Encephalitis Virus Prior to an Encephalitis Outbreak in Cordoba, Argentina (2005). PLoS Negl. Trop. Dis. 2012, 6, e1489. [Google Scholar] [CrossRef]
- Figueiredo, L.T. Saint louis encephalitis virus and other arboviroses in the differential diagnosis for dengue. Rev. Soc. Bras. Med. Trop. 2014, 47, 541–542. [Google Scholar] [CrossRef]
- Silva, J.R.; Romeiro, M.F.; Souza, W.N.; Munhoz, T.D.; Borges, G.P.; Soares, O.A.; Campos, C.H.; Machado, R.Z.; Silva, M.L.; Faria, J.L.; et al. A Saint Louis encephalitis and Rocio virus serosurvey in Brazilian horses. Rev. Soc. Bras. Med. Trop. 2014, 47, 414–417. [Google Scholar] [CrossRef]
- Pauvolid-Corrêa, A.; Campos, Z.; Juliano, R.; Velez, J.; Nogueira, R.M.; Komar, N. Serological evidence of widespread circulation of West Nile and other flaviviruses in equines of the Pantanal, Brazil. PLoS Negl. Trop. Dis. 2014, 8, e2706. [Google Scholar] [CrossRef]
- Rosa, R.; Costa, E.A.; Marques, R.E.; Oliveira, T.S.; Furtini, R.; Bonfim, M.R.; Teixeira, M.M.; Paixão, T.A.; Santos, R.L. Isolation of saint louis encephalitis virus from a horse with neurological disease in Brazil. PLoS Negl. Trop. Dis. 2013, 7, e2537. [Google Scholar] [CrossRef] [PubMed]
- Borges, I.A.; Reynolds, M.G.; McCollum, A.M.; Figueiredo, P.O.; Ambrosio, L.L.D.; Vieira, F.N.; Costa, G.B.; Matos, A.C.D.; de Andrade Almeida, V.M.; Ferreira, P.C.P.; et al. Serological Evidence of Orthopoxvirus Circulation Among Equids, Southeast Brazil. Front. Microbiol. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed]
- Costa, G.B.; Augusto, L.T.; Leite, J.A.; Ferreira, P.C.; Bonjardim, C.A.; Abrahão, J.S.; Kroon, E.G.; Moreno, E.C.; Trindade, G.S. Seroprevalence of Orthopoxvirus in rural Brazil: Insights into anti-OPV immunity status and its implications for emergent zoonotic OPV. Virol. J. 2016, 13, 121. [Google Scholar] [CrossRef] [PubMed]
- Companhia de Tecnologia da Informação do Estado de Minas Gerais, Brasil. Clima, Vegetação, e Relevo. Available online: http://www.mg.gov.br/conteudo/conheca-minas/geografia/clima-vegetacao-e-relevo (accessed on 15 October 2019).
- Instituto Estadual de Florestas (IEF). Cobertura Vegetal de Minas Gerais. Available online: http://www.ief.mg.gov.br/florestas (accessed on 15 October 2019).
- Vasconcelos, P.F.C.; Travassos-da-Rosa, J.F.S.; Travassos-da-Rosa, A.P.A.; Dégallier, N.; Pinheiros, F.P.; Sá Filho, G.C. Epidemiologia das encefalites por arbovírus na Amazônia brasileira. Rev. Inst. Med. Trop. Sao Paulo 1991, 33, 465–476. [Google Scholar] [CrossRef]
- Vasconcelos, P.F.C.; Travassos-da-Rosa, A.P.A.; Pinheiro, F.P.; Shope, R.E.; Travassos-da-Rosa, J.F.S.; Rodrigues, S.G.; Dégallier, N.; Travassos-da-Rosa, E.S. Arboviruses pathogenic for man in Brazil. In An Overview of Arbovirology in Brazil and Neighbouring Countries; Travassos-da-Rosa, A.P.A., Vasconcelos, P.F.C., Travassos-da-Rosa, J.F.S., Eds.; Instituto Evandro Chagas: Belém, Brazil, 1998; pp. 72–99. [Google Scholar]
- Díaz, L.A.; Almiron, W.R.; Almeida, L.F.; Spinsanti, L.I.; Contigiani, M. Vigilancia del virus Encefalitis de San Luis y mosquitos (Diptera: Culicidae) en la Provincia de Córdoba, Argentina. Entomol Vectores. 2003, 10, 551–566. [Google Scholar]
- Díaz, L.A.; Ré, V.; Almirón, W.R.; Farías, A.; Vásquez, A.; Sanchez-Seco, M.P.; Aguilar, J.; Spinsanti, L.; Konigheim, B.; Visintin, A.; et al. Genotype III Saint Louis Encephalitis Virus Outbreak, Argentina, 2005. Emerg. Infect. Dis. 2006, 12, 1752–1754. [Google Scholar] [CrossRef]
- Maeda, A.; Maeda, J. Review of diagnostic plaque reduction neutralization tests for flavivirus infection. Vet. J. 2013, 195, 33–40. [Google Scholar] [CrossRef]
- De Morais Bronzoni, R.V.; Baleotti, F.G.; Ribeiro Nogueira, R.M.; Nunes, M.; Figueiredo, L.T.M. Duplex reverse transcription-PCR followed by nested PCR assays for detection and identification of Brazilian alphaviruses and flaviviruses. J. Clin. Microbiol. 2005, 43, 696–702. [Google Scholar] [CrossRef]
- Rezende, I.M.; Sacchetto, L.; Munhoz de Mello, É.; Alves, P.A.; Iani, F.C.M.; Adelino, T.É.R.; Duarte, M.M.; Cury, A.L.F.; Bernardes, A.F.L.; Santos, T.A.; et al. Persistence of Yellow fever virus outside the Amazon Basin, causing epidemics in Southeast Brazil, from 2016 to 2018. PLoS Negl. Trop. Dis. 2018, 12, e0006538. [Google Scholar] [CrossRef]
- Tavares-Neto, J.; Travassos da Rosa, A.P.A.; Vasconcelos, P.F.C.; Costa, J.M.L.; Travassos da Rosa, J.F.S.; Marsden, P.D. Research of antibodies to arbovirus in the serum of residents of the village of Corte da Pedra, Valença, Bahia. Mem. Inst. Oswaldo Cruz 1986, 81, 351–358. [Google Scholar] [CrossRef]
- Romano-Lieber, N.S.; Iversson, L.B. Serological survey on arbovirus infection in residents of ecological reserve. Rev. Saúde Pública. 2000, 34, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Spinsanti, L.I.; Farías, A.; Aguilar, J.; Díaz, M.P.; Ghisiglieri, S.; Bustos, M.A.; Vilches, N.; González, B.; Contigiani, M. Risk factors associated with St. Louis encephalitis seroprevalence in two populations from Córdoba, Argentina. Trans. R. Soc. Trop. Med. Hyg. 2007, 101, 1248–1252. [Google Scholar] [CrossRef] [PubMed]
- Spinsanti, L.I.; Ré, V.E.; Díaz, M.P.; Contigiani, M.S. Age-related seroprevalence study for St. Louis encephalitis in a population from Cordoba, Argentina. Rev. Inst. Med. Trop. Sao Paulo 2002, 44, 59–62. [Google Scholar] [CrossRef] [PubMed]
- Rivarola, M.E.; Albrieu-Llinás, G.; Pisano, M.B.; Tauro, L.B.; Gorosito-Serrán, M.; Beccaria, C.G.; Díaz, L.A.; Vásquez, A.; Quaglia, A.; López, C.; et al. Tissue tropism of Saint Louis encephalitis virus: Histopathology triggered by epidemic and non-epidemic strains isolated in Argentina. Virology 2017, 505, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Monath, T.P.; Cropp, C.B.; Bowen, G.S.; Kemp, G.E.; Mitchell, C.J.; Gardner, J.J. Variation in virulence for mice and rhesus monkeys among St. Louis encephalitis virus strains of different origin. Am. J. Trop. Med. Hyg. 1980, 29, 948–962. [Google Scholar] [CrossRef]
- Pupo, M.; Guzmán, M.G.; Fernández, R.; Llop, A.; Dickinson, F.O.; Pérez, D.; Cruz, R.; González, T.; Estévez, G.; González, H.; et al. West Nile Virus Infection in Humans and Horses, Cuba. Emerg. Infect. Dis. 2006, 12, 1022–1024. [Google Scholar] [CrossRef]
- Johnson, B.W.; Kosoy, O.; Hunsperger, E.; Beltran, M.; Delorey, M.; Guirakhoo, F.; Monath, T. Evaluation of Chimeric Japanese Encephalitis and Dengue Viruses for Use in Diagnostic Plaque Reduction Neutralization Tests. Clin. Vaccine Immunol. 2009, 16, 1052–1059. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Equid Population n = 279 (%) | Seropositive n = 60 (%) | Seronegative n = 219 (%) | Odds Ratio 95% CI | p Value |
|---|---|---|---|---|---|
| Sex | |||||
| Female | 104 (37.3) | 28 (46.7) | 76 (34.7) | Reference | |
| Male | 175 (62.7) | 32 (53.3) | 143 (65.3) | 1.6 (0.9–3.0) | 0.1 |
| Age (years) | |||||
| ≤3 | 56 (20.1) | 6 (10.0) | 50 (22.8) | Reference | |
| 4–8 | 137 (49.1) | 35 (58.3) | 102 (46.6) | 2.9 (1.1–7.2) | 0.03 |
| >8 | 86 (30.8) | 19 (31.7) | 67 (30.6) | 0.4 (0.1–1.1) | 0.1 |
| Breed | |||||
| Equus cabalus * | 258 (92.5) | 54 (90.0) | 204 (93.2) | Reference | |
| Hibrids ** | 21 (7.5) | 6 (10.0) | 15 (6.8) | 0.7 (0.2–1.8) | 0.6 |
| Region † | |||||
| MR1 | 70 (25.1) | 13 (21.7) | 57 (26.0) | 5.8 (1.7–19.8) | 0.01 |
| MR2 | 71 (25.4) | 12 (20.0) | 59 (26.9) | 6.5 (1.9–22.4) | 0.006 |
| MR3 | 49 (17.6) | 18 (30.0) | 31 (14.1) | 2.3 (0.7–7.7) | 0.03 |
| MR4 | 14 (5.0) | 8 (13.3) | 6 (2.7) | 9.8 (2.7–34.7) | 0.001 |
| MR5 | 75 (26.9) | 9 (15.0) | 66 (30.1) | Reference |
| Variables | Human Population n = 240 (%) * | Seropositive n = 11 (%) | Seronegative n = 229 (%) | Odds Ratio 95% CI | P Value |
|---|---|---|---|---|---|
| Gender | |||||
| Female | 113 (47.1) | 4 (36.4) | 106 (46.3) | 0.67 | Reference |
| Male | 127 (52.9) | 7 (63.6) | 123 (53.7) | (0.19–2.32) | 0.7 |
| Age (years) | |||||
| ≤18 | 40 (16.7) | 2 (18.2) | 38 (16.6) | *** | Reference |
| 19–30 | 40 (16.7) | 0 | 40 (17.5) | 0.7 | |
| 31–50 | 97 (40.4) | 4 (36.4) | 93 (40.6) | 0.99 | |
| >50 | 63 (26.2) | 5 (45.4) | 58 (25.3) | 0.9 | |
| Years of schooling completed | |||||
| None | 24 (10.0) | 0 | 24 (10.5) | *** | Reference |
| ≤6 | 156 (65.0) | 9 (81.8) | 147 (64.2) | 0.99 | |
| 7–12 | 57 (23.8) | 2 (18.2) | 55 (24.0) | 0.99 | |
| >12 | 3 (1.2) | 0 | 3 (1.3) | 0.99 | |
| Income ** | |||||
| ≤1 min wage | 178 (74.2) | 11 (100.0) | 167 (72.9) | 2.5 | Reference |
| >1 min wage | 19 (7.9) | 0 | 19 (8.3) | (0.14–44.3) | 0.99 |
| Contact with equids | |||||
| Yes | 141 (58.8) | 7 (63.6) | 134 (58.5) | 1.2 | 0.99 |
| No | 96 (41.2) | 4 (36.4) | 92 (41.5) | (0.3–4.2) | Reference |
| Contact with wild environment | |||||
| Yes | 117 (48.8) | 8 (72.7) | 109 (47.6) | 2.9 | 0.2 |
| No | 123 (51.2) | 3 (27.3) | 120 (52.4) | (0.75–11.3) | Reference |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barbosa Costa, G.; Marinho, P.E.S.; Vilela, A.P.P.; Saraiva-Silva, A.T.; Crispim, A.P.C.; Borges, I.A.; Dutra, A.G.S.; Lobato, Z.I.P.; dos Reis, J.K.P.; de Oliveira, D.B.; et al. Silent Circulation of the Saint Louis Encephalitis Virus among Humans and Equids, Southeast Brazil. Viruses 2019, 11, 1029. https://doi.org/10.3390/v11111029
Barbosa Costa G, Marinho PES, Vilela APP, Saraiva-Silva AT, Crispim APC, Borges IA, Dutra AGS, Lobato ZIP, dos Reis JKP, de Oliveira DB, et al. Silent Circulation of the Saint Louis Encephalitis Virus among Humans and Equids, Southeast Brazil. Viruses. 2019; 11(11):1029. https://doi.org/10.3390/v11111029
Chicago/Turabian StyleBarbosa Costa, Galileu, Paula Eillany Silva Marinho, Ana Paula Pessoa Vilela, Ana Teresa Saraiva-Silva, Ana Paula Correia Crispim, Iara Apolinário Borges, Ana Gabriella Stoffella Dutra, Zélia Inês Portela Lobato, Jenner Karlison Pimenta dos Reis, Danilo Bretas de Oliveira, and et al. 2019. "Silent Circulation of the Saint Louis Encephalitis Virus among Humans and Equids, Southeast Brazil" Viruses 11, no. 11: 1029. https://doi.org/10.3390/v11111029
APA StyleBarbosa Costa, G., Marinho, P. E. S., Vilela, A. P. P., Saraiva-Silva, A. T., Crispim, A. P. C., Borges, I. A., Dutra, A. G. S., Lobato, Z. I. P., dos Reis, J. K. P., de Oliveira, D. B., Drumond, B. P., Kroon, E. G., & Trindade, G. d. S. (2019). Silent Circulation of the Saint Louis Encephalitis Virus among Humans and Equids, Southeast Brazil. Viruses, 11(11), 1029. https://doi.org/10.3390/v11111029