How Central Is the Domestic Pig in the Epidemiological Cycle of Japanese Encephalitis Virus? A Review of Scientific Evidence and Implications for Disease Control


Abstract
1. Introduction
2. Materials and Methods
2.1. Protocol, Search Process, and Databases
2.2. Inclusion and Exclusion Criteria
3. Results
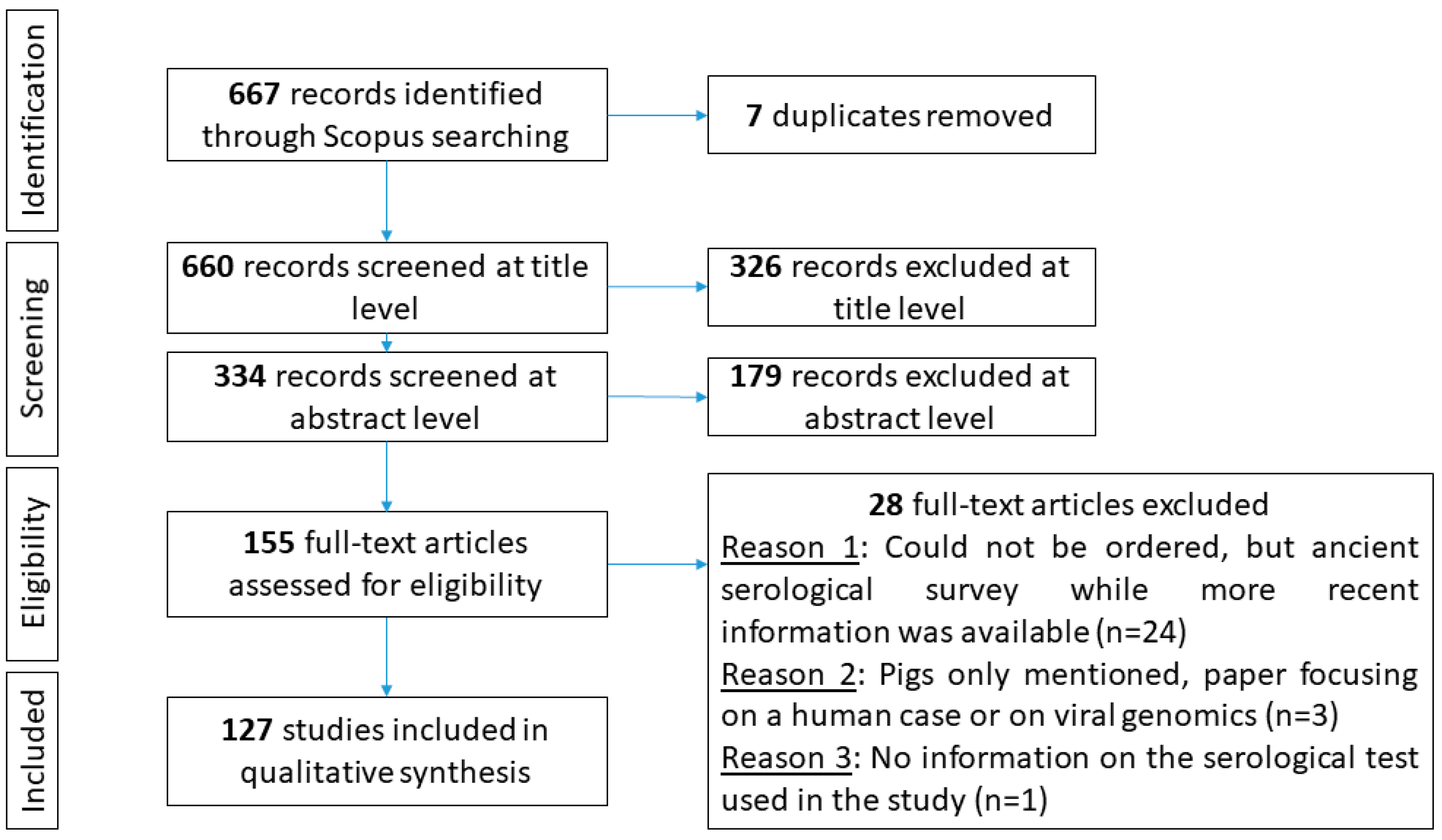
3.1. Study Selection
3.2. Individual Characteristics of JEV Infection in Swine
3.2.1. Viraemia in JEV-Infected Pigs
3.2.2. Humoral Response of Pigs after JEV Infection
3.2.3. Clinical Signs
3.2.4. Anti-JEV Maternal Antibodies
3.2.5. JEV Excretion
3.3. JEV Transmission Mechanisms and Patterns and Pig-Related Control
3.3.1. JEV Transmission Mechanisms
JEV Vector-Borne Transmission
JEV Direct Transmission
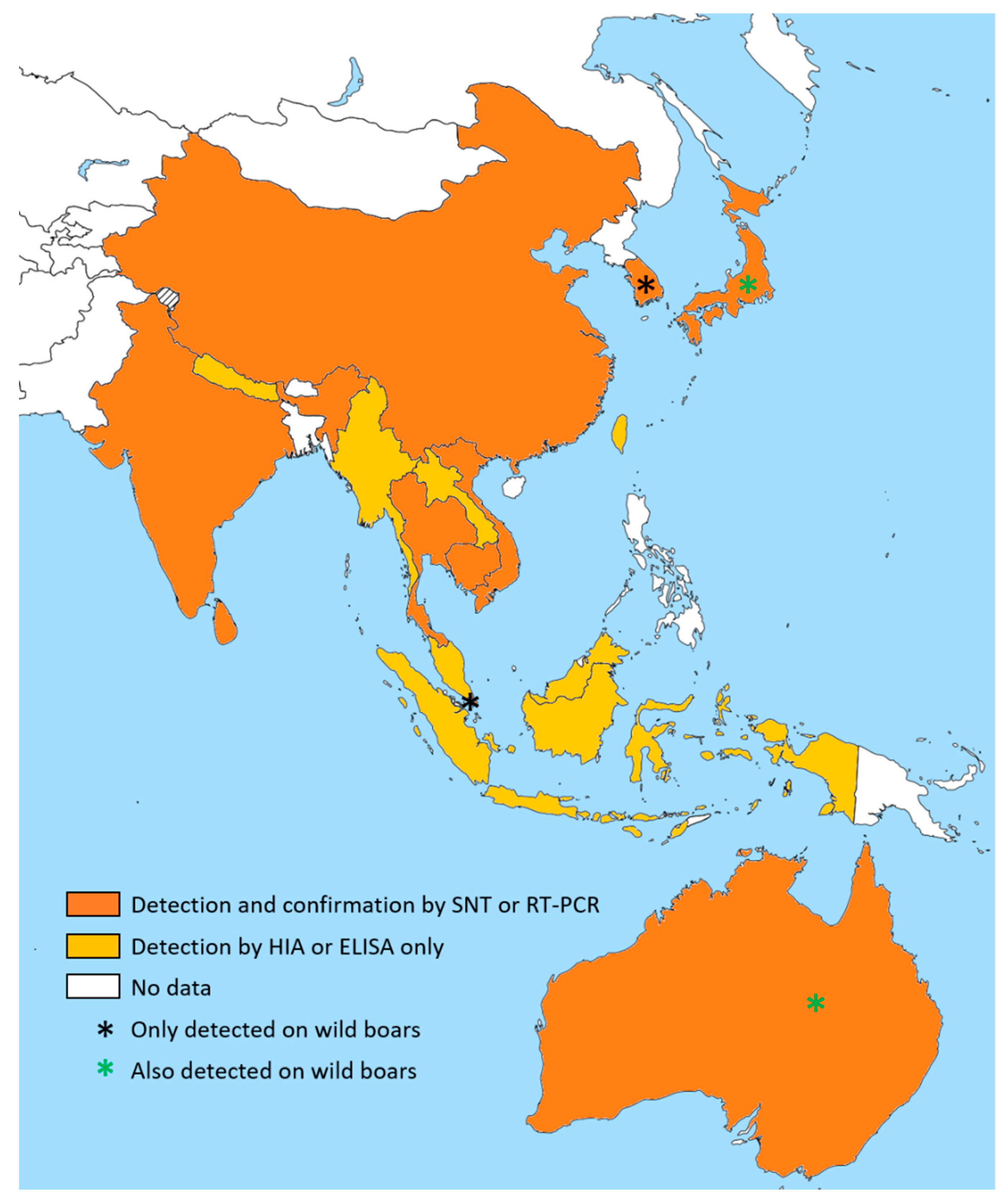
3.3.2. Geographical Distribution of JEV in Swine
3.3.3. Epidemiological Patterns
- (i)
- (ii)
- (iii)
- An epidemic pattern, with peaks of JEV transmission separated by periods of non-detection, as in north Australia [83], north India [117], Sri Lanka [82], the Chang Mai district in Thailand [124], and Japan [50,118,120,121,127,128]. In Japan, Konno et al. detailed cyclic outbreaks of JE among swine and human populations linked to vector abundances, which was reviewed in 2009 [120,122].
3.3.4. Pig-Related JEV Control
4. Discussion
- (i)
- The large South Korean vaccination program has not prevented outbreaks in the human population in recent years [145];
- (ii)
- Whatever the pig production system and the socio-economical context, the turnover of pig populations is always rapid and therefore the cost of vaccination is high;
- (iii)
- JEV may still circulate within vaccinated pig populations [144];
- (iv)
- Pig vaccines are based on GIII viruses, the dominant circulating genotype in Asia. However, there are now several studies showing the replacement of GIII by GI that could negatively modify the effectiveness of current vaccines [113,189,190]. JEV was indeed detected in vaccinated populations in aborted fetuses and stillborn piglets in China [68,70]. The authors warned against a potential lack of effectiveness of the vaccine and suggested that the safety of the SA14-14-2 strain belonging to GIII, which is used for vaccine development in pigs, should be reassessed;
- (v)
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Campbell, G.L.; Hills, S.L.; Fischer, M.; Jacobson, J.A.; Hoke, C.H.; Hombach, J.M.; AMarfin, A.; Solomon, T.; Tsai, T.F.; Tsu, V.D.; et al. Estimated global incidence of Japanese encephalitis: A systematic review. Bull. World Health Organ. 2011, 89, 766–774. [Google Scholar] [CrossRef] [PubMed]
- Fischer, M.; Hills, S.; Staples, E.; Johnson, B.; Yaich, M.; Solomon, T. Japanese Encephalitis Prevention and Control: Advances, Challenges, and New Initiatives. Emerg. Infect. 2008, 8, 93–124. [Google Scholar]
- Simon-Loriere, E.; Faye, O.; Prot, M. Autochthonous Japanese Encephalitis with Yellow Fever Coinfection in Africa. N. Engl. J. Med. 2017, 376, 1483–1485. [Google Scholar] [CrossRef] [PubMed]
- Le Flohic, G.; Gonzalez, J.-P. When Japanese Encephalitis Virus Invaded Eastern Hemisphere—The History of the Spread of Virus Genotypes. In Flavivirus Encephalitis; Ruzek, D., Ed.; InTech: London, UK, 2011. [Google Scholar] [CrossRef]
- Wang, H.; Liang, G. Epidemiology of Japanese encephalitis: Past, present, and future prospects. Ther. Clin. Risk Manag. 2015, 11, 435–448. [Google Scholar] [PubMed]
- Duong, V.; Choeung, R.; Gorman, C.; Laurent, D.; Crabol, Y.; Mey, C.; Peng, B.; Di Francesco, J.; Hul, V.; Sothy, H.; et al. Isolation and full-genome sequences of Japanese encephalitis virus genotype I strains from Cambodian human patients, mosquitoes and pigs. J. Gen. Virol. 2017, 98, 2287–2296. [Google Scholar] [CrossRef] [PubMed]
- De Wispelaere, M.; Desprès, P.; Choumet, V. European Aedes albopictus and Culex pipiens Are Competent Vectors for Japanese Encephalitis Virus. PLoS Negl. Trop. Dis. 2017, 11, e0005294. [Google Scholar] [CrossRef] [PubMed]
- Huber, K.; Jansen, S.; Leggewie, M.; Badusche, M.; Schmidt-Chanasit, J.; Becker, N.; Tannich, E.; Becker, S.C. Aedes japonicus japonicus (Diptera: Culicidae) from Germany have vector competence for Japan encephalitis virus but are refractory to infection with West Nile virus. Parasitol. Res. 2014, 113, 3195–3199. [Google Scholar] [CrossRef]
- Turell, M.J.; Mores, C.N.; Dohm, D.J.; Komilov, N.; Paragas, J.; Lee, J.S.; Shermuhemedova, D.; Endy, T.P.; Kodirov, A.; Khodjaev, S. Laboratory Transmission of Japanese Encephalitis and West Nile Viruses by Molestus Form ofCulex pipiens(Diptera: Culicidae) Collected in Uzbekistan in 2004. J. Med. Èntomol. 2006, 43, 296–300. [Google Scholar] [CrossRef]
- Olson, J.G.; Ksiazek, T.G.; Tan, R.; Atmosoedjono, S.; Lee, V.H.; Converse, J.D. Correlation of population indices of female Culex tritaeniorhynchus with Japanese encephalitis viral activity in Kapuk, Indonesia. Southeast Asian J. Trop. Med. Public Health 1985, 16, 337–342. [Google Scholar]
- Rajavel, A.R.; Kumar, N.P.; Natarajan, R.; Vanamail, P.; Rathinakumar, A.; Jambulingam, P. Morphological and molecular characterization of the ecological, biological and behavioural variants of the JE vector Culex tritaeniorhynchus: An assessment of its taxonomic status. J. Vector Borne Dis. 2015, 52, 40–51. [Google Scholar]
- Lord, J.S.; Al-Amin, H.M.; Chakma, S.; Alam, M.S.; Gurley, E.S.; Pulliam, J.R.C. Sampling Design Influences the Observed Dominance of Culex tritaeniorhynchus: Considerations for Future Studies of Japanese Encephalitis Virus Transmission. PLoS Negl. Trop. Dis. 2016, 10, e0004249. [Google Scholar] [CrossRef] [PubMed]
- Gajanana, A.; Rajendran, R.; Samuel, P.P.; Thenmozhi, V.; Tsai, T.F.; Kimura-Kuroda, J.; Reuben, R. Japanese Encephalitis in South Arcot District, Tamil Nadu, India: A Three-Year Longitudinal Study of Vector Abundance and Infection Frequency. J. Med. Èntomol. 1997, 34, 651–659. [Google Scholar] [CrossRef] [PubMed]
- Gingrich, J.B.; Nisalak, A.; Latendresse, J.R.; Pomsdhit, J.; Paisansilp, S.; Hoke, C.H.; Chantalakana, C.; Satayaphantha, C.; Uechiewcharnkit, K. A longitudinal study of Japanese encephalitis in suburban Bangkok, Thailand. Southeast Asian J. Trop. Med. Public Health 1987, 18, 558–566. [Google Scholar]
- Nyari, N.; Singh, D.; Kakkar, K.; Sharma, S.; Pandey, S.N.; Dhole, T.N. Entomological and serological investigation of Japanese encephalitis in endemic area of eastern Uttar Pradesh, India. J. Vector Borne Dis. 2015, 52, 321–328. [Google Scholar]
- Buescher, E.L.; Scherer, W.F.; Mccluse, H.; Moyer, J.T.; Rosenberg, M.Z.; Yoshii, Y.; Okada, Y. Ecologic Studies of Japanese Encephalitis Virus in Japan. Am. J. Trop. Med. Hyg. 1959, 8, 678–688. [Google Scholar] [CrossRef]
- Scherer, W.F.; Moyer, J.T.; Izumi, T.; Gresser, I.; McCown, J. Ecologic Studies of Japanese Encephalitis Virus in Japan. Am. J. Trop. Med. Hyg. 1959, 8, 644–650. [Google Scholar] [CrossRef]
- Soman, R.S.; Rodrigues, F.M.; Guttikar, S.N.; Guru, P.Y. Experimental viraemia and transmission of Japanese encephalitis virus by mosquitoes in ardeid birds. Indian J. Med. Res. 1977, 66, 709–718. [Google Scholar]
- Le Flohic, G.; Porphyre, V.; Barbazan, P.; Gonzalez, J.-P. Review of Climate, Landscape, and Viral Genetics as Drivers of the Japanese Encephalitis Virus Ecology. PLoS Negl. Trop. Dis. 2013, 7, e2208. [Google Scholar] [CrossRef]
- Impoinvil, D.E.; Baylis, M.; Solomon, T. Japanese encephalitis: On the One Health agenda. Curr. Top. Microbiol. Immunol. 2013, 365, 205–247. [Google Scholar]
- Ricklin, M.E.; García-Nicolás, O.; Brechbühl, D.; Python, S.; Zumkehr, B.; Nougairede, A.; Charrel, R.N.; Posthaus, H.; Oevermann, A.; Summerfield, A. Vector-free transmission and persistence of Japanese encephalitis virus in pigs. Nat. Commun. 2016, 7, 10832. [Google Scholar] [CrossRef]
- Ricklin, M.E.; García-Nicolás, O.; Brechbühl, D.; Python, S.; Zumkehr, B.; Posthaus, H.; Oevermann, A.; Summerfield, A. Japanese encephalitis virus tropism in experimentally infected pigs. Veter Res. 2016, 47, 34. [Google Scholar] [CrossRef] [PubMed]
- Lyons, A.C.; Huang, Y.-J.S.; Park, S.L.; Ayers, V.B.; Hettenbach, S.M.; Higgs, S.; McVey, D.S.; Noronha, L.; Hsu, W.-W.; VanLandingham, D.L. Shedding of Japanese Encephalitis Virus in Oral Fluid of Infected Swine. Vector-Borne Zoonotic Dis. 2018, 18, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Cleton, N.B.; Page, M.J.; Bowen, R.A.; Bosco-Lauth, A. Age-Related Susceptibility to Japanese Encephalitis Virus in Domestic Ducklings and Chicks. Am. J. Trop. Med. Hyg. 2014, 90, 242–246. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, N.; Bosco-Lauth, A.; Oesterle, P.; Kohler, D.; Bowen, R. North American Birds as Potential Amplifying Hosts of Japanese Encephalitis Virus. Am. J. Trop. Med. Hyg. 2012, 87, 760–767. [Google Scholar] [CrossRef]
- Bae, W.; Kim, J.H.; Kim, J.; Lee, J.; Hwang, E.S. Changes of Epidemiological Characteristics of Japanese Encephalitis Viral Infection and Birds as a Potential Viral Transmitter in Korea. J. Korean Med. Sci. 2018, 33, 33. [Google Scholar] [CrossRef]
- Adi, A.A.A.M.; Astawa, N.M.; Damayanti, P.A.A.; Kardena, I.M.; Erawan, I.G.M.K.; Suardana, I.W.; Putra, I.G.A.A.; Matsumoto, Y. Seroepidemiological Evidence for the Presence of Japanese Encephalitis Virus Infection in Ducks, Chickens, and Pigs, Bali-Indonesia. Bali Med. J. 2016, 5, 189. [Google Scholar]
- Shimoda, H.; Ohno, Y.; Mochizuki, M.; Iwata, H.; Okuda, M.; Maeda, K. Dogs as Sentinels for Human Infection with Japanese Encephalitis Virus. Emerg. Infect. Dis. 2010, 16, 1137–1139. [Google Scholar] [CrossRef]
- Shimoda, H.; Tamaru, S.; Morimoto, M.; Hayashi, T.; Shimojima, M.; Maeda, K. Experimental infection of Japanese encephalitis virus in dogs. J. Veter Med. Sci. 2011, 73, 1241–1242. [Google Scholar] [CrossRef][Green Version]
- Wang, J.-L.; Pan, X.-L.; Zhang, H.-L.; Fu, S.-H.; Wang, H.-Y.; Tang, Q.; Wang, L.-F.; Liang, G.-D. Japanese Encephalitis Viruses from Bats in Yunnan, China. Emerg. Infect. Dis. 2009, 15, 939–942. [Google Scholar] [CrossRef]
- Kumar, K.; Arshad, S.S.; Selvarajah, G.T.; Abu, J.; Toung, O.P.; Abba, Y.; Bande, F.; Yasmin, A.R.; Sharma, R.; Ong, B.L.; et al. Prevalence and risk factors of Japanese encephalitis virus (JEV) in livestock and companion animal in high-risk areas in Malaysia. Trop. Anim. Health Prod. 2018, 50, 741–752. [Google Scholar] [CrossRef]
- Kako, N.; Suzuki, S.; Sugie, N.; Kato, T.; Yanase, T.; Yamakawa, M.; Shirafuji, H. Japanese encephalitis in a 114-month-old cow: Pathological investigation of the affected cow and genetic characterization of Japanese encephalitis virus isolate. BMC Vet. Res. 2014, 10, 63. [Google Scholar] [CrossRef]
- Katayama, T.; Saito, S.; Horiuchi, S.; Maruta, T.; Kato, T.; Yanase, T.; Yamakawa, M.; Shirafuji, H. Nonsuppurative Encephalomyelitis in a Calf in Japan and Isolation of Japanese Encephalitis Virus Genotype 1 from the Affected Calf. J. Clin. Microbiol. 2013, 51, 3448–3453. [Google Scholar] [CrossRef] [PubMed]
- Hanna, J.N.; Ritchie, S.A.; Phillips, D.A.; Shield, J.; Bailey, M.C.; Mackenzie, J.S.; Poidinger, M.; McCall, B.J.; Mills, P.J. An outbreak of Japanese encephalitis in the Torres Strait, Australia, 1995. Med. J. Aust. 1996, 165, 256–260. [Google Scholar] [CrossRef] [PubMed]
- Ting, S.H.L.; Tan, H.C.; Wong, W.K.; Ng, M.L.; Chan, S.H.; Ooi, E.E. Seroepidemiology of neutralizing antibodies to Japanese encephalitis virus in Singapore: Continued transmission despite abolishment of pig farming? Acta Trop. 2004, 92, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Lian, W.-C.; Liau, M.-Y.; Mao, C.-L.; Lian, W.; Liau, M.; Mao, C. Diagnosis and Genetic Analysis of Japanese Encephalitis Virus Infected in Horses. J. Veter Med. Ser. B 2002, 49, 361–365. [Google Scholar] [CrossRef]
- Gulati, B.R.; Singha, H.; Singh, B.K.; Virmani, N.; Kumar, S.; Singh, R.K. Isolation and genetic characterization of Japanese encephalitis virus from equines in India. J. Veter Sci. 2012, 13, 111–118. [Google Scholar] [CrossRef]
- Baharuddin, A.; Hassan, A.A.; Sheng, G.C.; Nasir, S.B.; Othman, S.; Yusof, R.; Othman, R.; Rahman, N.A. Current approaches in antiviral drug discovery against the Flaviviridae family. Curr. Pharm. Des. 2014, 20, 3428–3444. [Google Scholar] [CrossRef]
- Liberati, A.; Altman, D.G.; Tetzlaff, J.; Mulrow, C.; Gøtzsche, P.C.; Ioannidis, J.P.; Clarke, M.; Devereaux, P.; Kleijnen, J.; Moher, D. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate health care interventions: Explanation and elaboration. J. Clin. Epidemiol. 2009, 62, e1–e34. [Google Scholar] [CrossRef]
- Yang, D.-K.; Kweon, C.-H.; Kim, B.-H.; Lim, S.-I.; Kim, S.-H.; Kwon, J.-H.; Han, H.-R. TaqMan reverse transcription polymerase chain reaction for the detection of Japanese encephalitis virus. J. Veter Sci. 2004, 5, 345–351. [Google Scholar] [CrossRef]
- Ogawa, H.; Taira, O.; Hirai, T.; Takeuchi, H.; Nagao, A.; Ishikawa, Y.; Tuchiya, K.; Nunoya, T.; Ueda, S. Multiplex PCR and multiplex RT-PCR for inclusive detection of major swine DNA and RNA viruses in pigs with multiple infections. J. Virol. Methods 2009, 160, 210–214. [Google Scholar] [CrossRef]
- Patel, P.; Landt, O.; Kaiser, M.; Faye, O.; Koppe, T.; Lass, U.; Sall, A.A.; Niedrig, M. Development of one-step quantitative reverse transcription PCR for the rapid detection of flaviviruses. Virol. J. 2013, 10, 58. [Google Scholar] [CrossRef] [PubMed]
- Eldadah, Z.A.; Asher, D.M.; Godec, M.S.; Pomeroy, K.L.; Goldfarb, L.G.; Feinstone, S.M.; Levitan, H.; Gibbs, C.J.; Gajdusek, D.C. Detection of flaviviruses by reverse-transcriptase polymerase chain reaction. J. Med. Virol. 1991, 33, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Hierholzer, J.C.; Killington, R.A. 2-Virus isolation and quantitation. In Virology Methods Manual; Mahy, B.W., Kangro, H.O., Eds.; Academic Press: London, UK, 1996; pp. 25–46. [Google Scholar]
- Chernesky, M.A. 6-Traditional serological tests. In Virology Methods Manual; Mahy, B.W., Kangro, H.O., Eds.; Academic Press: London, UK, 1996; pp. 107–122. [Google Scholar]
- Yonemitsu, K.; Minami, S.; Noguchi, K.; Kuwata, R.; Shimoda, H.; Maeda, K. Detection of anti-viral antibodies from meat juice of wild boars. J. Vet. Med Sci. 2019, 81, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Scherer, W.F.; Moyer, J.T.; Izumi, T. Immunologic studies of Japanese encephalitis virus in Japan. V. Maternal antibodies, antibody responses and viremia following infection of swine. J. Immunol. 1959, 83, 620–626. [Google Scholar]
- Sasaki, O.; Karoji, Y.; Kuroda, A.; Karaki, T.; Takenokuma, K.; Maeda, O. Protection of pigs against mosquito-borne japanese encephalitis virus by immunization with a live attenuated vaccine. Antivir. Res. 1982, 2, 355–360. [Google Scholar] [CrossRef]
- Imoto, J.-I.; Ishikawa, T.; Yamanaka, A.; Konishi, M.; Murakami, K.; Shibahara, T.; Kubo, M.; Lim, C.-K.; Hamano, M.; Takasaki, T.; et al. Needle-free jet injection of small doses of Japanese encephalitis DNA and inactivated vaccine mixture induces neutralizing antibodies in miniature pigs and protects against fetal death and mummification in pregnant sows. Vaccine 2010, 28, 7373–7380. [Google Scholar] [CrossRef] [PubMed]
- Takashima, I.; Hashimoto, N.; Watanabe, T.; Ouchi, N. Ecological Studies of Japanese Encephalitis Virus in Hokkaido: Interepidemic Outbreaks of Swine Abortion and Evidence for the Virus to Overwinter Locally. Am. J. Trop. Med. Hyg. 1988, 38, 420–427. [Google Scholar] [CrossRef]
- Shimizu, T.; Kawakami, Y.; Fukuhara, S.; Matumoto, M. Experimental stillbirth in pregnant swine infected with Japanese encephalitis virus. Jpn. J. Exp. Med. 1954, 24, 363–375. [Google Scholar]
- Carey, D.E.; Reuben, R.; Myers, R.M. Japanese encephalitis studies in Vellore, South India. V. Experimental infection and transmission. Indian J. Med Res. 1969, 57, 282–289. [Google Scholar]
- Williams, D.T.; ALunt, R.; Wang, L.F.; Daniels, P.W.; Newberry, K.M.; MacKenzie, J.S. Experimental infections of pigs with Japanese encephalitis virus and closely related Australian flaviviruses. Am. J. Trop. Med. Hyg. 2001, 65, 379–387. [Google Scholar] [CrossRef]
- Park, S.L.; Huang, Y.-J.S.; Lyons, A.C.; Ayers, V.B.; Hettenbach, S.M.; McVey, D.S.; Burton, K.R.; Higgs, S.; VanLandingham, D.L. North American domestic pigs are susceptible to experimental infection with Japanese encephalitis virus. Sci. Rep. 2018, 8, 7951. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, C.; Soler, A.; Nieto, R.; Sánchez, M.A.; Martins, C.; Pelayo, V.; Carrascosa, A.; Revilla, Y.; Simon, A.; Briones, V.; et al. Experimental Transmission of African Swine Fever (ASF) Low Virulent Isolate NH/P68 by Surviving Pigs. Transbound. Emerg. Dis. 2015, 62, 612–622. [Google Scholar] [CrossRef] [PubMed]
- Anderson, E. African swine fever: Current concept on its pathogenesis and immunology. Rev. Sci. Tech. 1986, 5, 477–486. [Google Scholar] [CrossRef]
- Beltran-Alcrudo, D.; Arias, M.; Gallardo, C.; Kramer, S.A.; Penrith, M.-L. African Swine Fever: Detection and Diagnosis: A Manual for Veterinarians; Food and Agriculture Organization of the United Nations: Rome, Italy, 2017. [Google Scholar]
- Gong, W.; Li, J.; Wang, Z.; Sun, J.; Mi, S.; Lu, Z.; Cao, J.; Dou, Z.; Sun, Y.; Wang, P.; et al. Virulence evaluation of classical swine fever virus subgenotype 2.1 and 2.2 isolates circulating in China. Veter Microbiol. 2019, 232, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Durand, B.; Davila, S.; Cariolet, R.; Mesplède, A.; Le Potier, M.-F. Comparison of viraemia- and clinical-based estimates of within- and between-pen transmission of classical swine fever virus from three transmission experiments. Veter Microbiol. 2009, 135, 196–204. [Google Scholar] [CrossRef]
- Ueba, N.; Maeda, A.; Otsu, K.; Mitsuda, B.; Kimoto, T. Natural infection of swine by Japanese encephalitis virus and its modification by vaccination. Biken J. J. Res. Inst. Microb. Dis. 1972, 15, 67–79. [Google Scholar]
- Geevarghese, G.; Shaikh, B.H.; Jacob, P.G.; Bhat, H.R. Persistence of haemagglutination-inhibition antibodies to JE and WN viruses in naturally infected domestic pigs in Karnataka State, India. Acta Virol. 1994, 38, 235–237. [Google Scholar]
- Nakamura, K.; Kaku, Y.; Yamada, M.; Yoshii, M. Nonsuppurative Encephalitis in Piglets after Experimental Inoculation of Japanese Encephalitis Flavivirus Isolated from Pigs. Veter Pathol. 2004, 41, 62–67. [Google Scholar]
- Yamada, M.; Nakamura, K.; Yoshii, M.; Kaku, Y.; Narita, M. Brain Lesions Induced by Experimental Intranasal Infection of Japanese Encephalitis Virus in Piglets. J. Comp. Pathol. 2009, 141, 156–162. [Google Scholar] [CrossRef]
- Wu, R.; Wang, Q.; Liu, H.; Chai, C.; He, B.; Huang, X.; Wen, Y.; Wen, X.; Yan, Q.; Ma, X.; et al. Phylogenetic analysis reveals that Japanese encephalitis virus genotype III is still prevalent in swine herds in Sichuan province in China. Arch. Virol. 2016, 161, 1719–1722. [Google Scholar] [CrossRef]
- Desingu, P.A.; Ray, P.K.; Patel, B.H.M.; Singh, R.; Saikumar, G. Pathogenic and Genotypic Characterization of a Japanese Encephalitis Virus Isolate Associated with Reproductive Failure in an Indian Pig Herd. PLoS ONE 2016, 11, 0147611. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Wu, R.; Liu, H.; Wen, X.; Huang, X.; Wen, Y.; Ma, X.; Yan, Q.; Huang, Y.; Zhao, Q.; et al. Tissue tropism and molecular characterization of a Japanese encephalitis virus strain isolated from pigs in southwest China. Virus Res. 2016, 215, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, J.; Boqvist, S.; Ståhl, K.; Thu, H.T.V.; Magnusson, U. Reproductive performance in sows in relation to Japanese Encephalitis Virus seropositivity in an endemic area. Trop. Anim. Health Prod. 2012, 44, 239–245. [Google Scholar] [CrossRef]
- Lu, B.; Qin, Y.; Li, B.; He, Y.; Duan, Q.; Liang, J.; Chen, Z.; Su, Q.; Bi, B.; Zhao, W. Full-Length Genome Sequence of Japanese Encephalitis Virus Strain FC792, Isolated from Guangxi, China. Genome Announc. 2017, 5, e01054-17. [Google Scholar] [CrossRef]
- Teng, M.; Luo, J.; Fan, J.M.; Chen, L.; Wang, X.T.; Yao, W.; Wang, C.Q.; Zhang, G.P. Molecular characterization of Japanese encephalitis viruses circulating in pigs and mosquitoes on pig farms in the Chinese province of Henan. Virus Genes 2013, 46, 170–174. [Google Scholar] [CrossRef]
- Liu, H.; Lu, H.-J.; Liu, Z.-J.; Jing, J.; Ren, J.-Q.; Liu, Y.-Y.; Lu, F.; Jin, N.-Y. Japanese Encephalitis Virus in Mosquitoes and Swine in Yunnan Province, China 2009–2010. Vector Borne Zoonotic Dis. 2013, 13, 41–49. [Google Scholar] [CrossRef]
- Cao, Q.S.; Li, X.M.; Zhu, Q.Y.; Wang, D.D.; Chen, H.C.; Qian, P. Isolation and molecular characterization of genotype 1 Japanese encephalitis virus, SX09S-01, from pigs in China. Virol. J. 2011, 8, 472. [Google Scholar] [CrossRef]
- Ogasa, A.; Yokoki, Y.; Fujisaki, Y.; Habu, A. Reproductive disorders in boars infected experimentally with Japanese encephalitis virus. Jpn. J. Anim. Reprod. 1977, 23, 171–175. [Google Scholar] [CrossRef]
- Okuno, T.; Mitchell, C.J.; Chen, P.S.; Wang, J.S.; Lin, S.Y. Seasonal infection of Culex mosquitos and swine with Japanese encephalitis virus. Bull. World Health Organ. 1973, 49, 347–352. [Google Scholar]
- Cappelle, J.; Duong, V.; Pring, L.; Kong, L.; Yakovleff, M.; Prasetyo, D.B.; Peng, B.; Choeung, R.; Duboz, R.; Ong, S.; et al. Intensive Circulation of Japanese Encephalitis Virus in Peri-urban Sentinel Pigs near Phnom Penh, Cambodia. PLoS Negl. Trop. Dis. 2016, 10, e0005149. [Google Scholar] [CrossRef]
- Di Francesco, J.; Choeung, R.; Peng, B.; Pring, L.; Pang, S.; Duboz, R.; Ong, S.; Sorn, S.; Tarantola, A.; Fontenille, D.; et al. Comparison of the dynamics of Japanese encephalitis virus circulation in sentinel pigs between a rural and a peri-urban setting in Cambodia. PLoS Negl. Trop. Dis. 2018, 12, e0006644. [Google Scholar] [CrossRef] [PubMed]
- Barzon, L.; Pacenti, M.; Franchin, E.; Pagni, S.; Martello, T.; Cattai, M.; Cusinato, R.; Palù, G. Excretion of West Nile Virus in Urine During Acute Infection. J. Infect. Dis. 2013, 208, 1086–1092. [Google Scholar] [CrossRef] [PubMed]
- Korhonen, E.M.; Huhtamo, E.; Virtala, A.-M.K.; Kantele, A.; Vapalahti, O. Approach to non-invasive sampling in dengue diagnostics: Exploring virus and NS1 antigen detection in saliva and urine of travelers with dengue. J. Clin. Virol. 2014, 61, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Hurlbut, H.S. The Pig-Mosquito Cycle of Japanese Encephalitis Virus in Taiwan. J. Med. Èntomol. 1964, 1, 301–307. [Google Scholar] [CrossRef]
- García-Nicolás, O.; Braun, R.O.; Milona, P.; Lewandowska, M.; Dijkman, R.; Alves, M.P.; Summerfield, A. Targeting of the Nasal Mucosa by Japanese Encephalitis Virus for Non-Vector-Borne Transmission. J. Virol. 2018, 92, e01091-18. [Google Scholar] [CrossRef]
- Duong, V.; Sorn, S.; Holl, D.; Rani, M.; Deubel, V.; Buchy, P. Evidence of Japanese encephalitis virus infections in swine populations in 8 provinces of Cambodia: Implications for national Japanese encephalitis vaccination policy. Acta Trop. 2011, 120, 146–150. [Google Scholar] [CrossRef]
- Burke, D.S.; Tingpalapong, M.; Ward, G.S.; Andre, R.; Leake, C.J. Intense transmission of Japanese encephalitis virus to pigs in a region free of epidemic encephalitis. Southeast Asian J. Trop. Med. Public Health 1985, 16, 199–206. [Google Scholar]
- Peiris, J.; Amerasinghe, F.; Arunagiri, C.; Perera, L.; Karunaratne, S.; Ratnayake, C.; Kulatilaka, T.; Abeysinghe, M. Japanese encephalitis in Sri Lanka: Comparison of vector and virus ecology in different agro-climatic areas. Trans. R. Soc. Trop. Med. Hyg. 1993, 87, 541–548. [Google Scholar] [CrossRef]
- Hanna, J.N.; ARitchie, S.; Hills, S.L.; Hurk, A.F.V.D.; APhillips, D.; Pyke, A.T.; Lee, J.M.; AJohansen, C.; MacKenzie, J.S. Japanese encephalitis in north Queensland, Australia, 1998. Med. J. Aust. 1999, 170, 533–536. [Google Scholar] [CrossRef]
- Higgins, D.A. A serological survey of pigs in Hong Kong for antibodies to Japanese encephalitis virus. Trop. Anim. Health Prod. 1970, 2, 23–27. [Google Scholar] [CrossRef]
- Kakkar, M.; Chaturvedi, S.; Saxena, V.K.; Dhole, T.N.; Kumar, A.; Rogawski, E.T.; Abbas, S.; Venkataramanan, V.V.; Chatterjee, P. Identifying sources, pathways and risk drivers in ecosystems of Japanese Encephalitis in an epidemic-prone north Indian district. PLoS ONE 2017, 12, e0175745. [Google Scholar] [CrossRef] [PubMed]
- Kumari, R.; Kumar, K.; Rawat, A.; Singh, G.; Yadav, N.K.; Chauhan, L.S. First indigenous transmission of Japanese Encephalitis in urban areas of National Capital Territory of Delhi, India. Trop. Med. Int. Health 2013, 18, 743–749. [Google Scholar] [CrossRef] [PubMed]
- Dutta, P.; Khan, S.A.; Khan, A.M.; Mahanta, J. Japanese encephalitis B-virus among the porcine population of Assam. Indian Vet. J. 2007, 84, 1125–1127. [Google Scholar]
- Kumanan, K.; Ramesh, A.; Velmurugan, R.; Jagannathan, S.; Padmanaban, V.D. Seroepidemiology of Japanese encephalitis among animals and birds in Tamil Nadu. Indian Vet. J. 2002, 79, 311–315. [Google Scholar]
- Damayanti, P.A.A.; Adi, A.A.A.M.; Astawa, I.N.M.; Sudarmaja, I.M.; Kardena, I.M.; Swastika, I.K. Incidence of Japanese Encephalitis among Children is associated with the Presence of Pigs in Bali, Indonesia. Biomed. Pharmacol. J. 2017, 10, 1333–1338. [Google Scholar] [CrossRef]
- Detha, A.; Wuri, D.A.; Santhia, K. Seroprevalence of Japanese encephalitis virus using competitive enzyme linked immunosorbent assay (C-ELISA) in pigs in East Sumba, Indonesia. J. Adv. Vet. Anim. Res. 2015, 2, 481–483. [Google Scholar] [CrossRef]
- Yamanaka, A.; Mulyatno, K.C.; Susilowati, H.; Hendrianto, E.; Utsumi, T.; Amin, M.; Lusida, M.I.; Soegijanto, S.; Konishi, E. Prevalence of antibodies to Japanese encephalitis virus among pigs in Bali and East Java, Indonesia, 2008. Jpn. J. Infect. Dis. 2010, 63, 58–60. [Google Scholar]
- Ohno, Y.; Sato, H.; Suzuki, K.; Yokoyama, M.; Uni, S.; Shibasaki, T.; Sashika, M.; Inokuma, H.; Kai, K.; Maeda, K. Detection of antibodies against Japanese encephalitis virus in raccoons, raccoon dogs and wild boars in Japan. J. Veter Med. Sci. 2009, 71, 1035–1039. [Google Scholar] [CrossRef]
- Hamano, M.; Lim, C.K.; Takagi, H.; Sawabe, K.; Kuwayama, M.; Kishi, N.; Kurane, I.; Takasaki, T. Detection of antibodies to Japanese encephalitis virus in the wild boars in Hiroshima prefecture, Japan. Epidemiol. Infect. 2007, 135, 974–977. [Google Scholar] [CrossRef]
- Sugiyama, I.; Shimizu, E.; Nogami, S.; Suzuki, K.; Miura, Y.; Sentsui, H. Serological survey of arthropod-borne viruses among wild boars in Japan. J. Veter Med. Sci. 2009, 71, 1059–1061. [Google Scholar] [CrossRef]
- Nidaira, M.; Kyan, H.; Taira, K.; Okano, S.; Oshiro, T.; Kato, T.; Kudo, N.; Azama, Y.; Mahoe, Y.; Kudaka, J.; et al. Survey of Japanese encephalitis virus in pigs and wild boars on Ishigaki and Iriomote Islands in Okinawa, Japan. Epidemiol. Infect. 2014, 142, 856–860. [Google Scholar] [CrossRef] [PubMed]
- Nidaira, M.; Taira, K.; Onodera, I.; Morikawa, T.; Itokazu, K.; Kudaka, J.; Ohno, A. Detection of Japanese encephalitis virus antibody in a pig on Yonaguni Island, where all pigs were slaughtered in 1997. Jpn. J. Infect. Dis. 2007, 60, 70–71. [Google Scholar] [PubMed]
- Nidaira, M.; Taira, K.; Okano, S.; Shinzato, T.; Morikawa, T.; Tokumine, M.; Asato, Y.; Tada, Y.; Miyagi, K.; Matsuda, S.; et al. Survey of Japanese encephalitis virus in pigs on Miyako, Ishigaki, Kume, and Yonaguni Islands in Okinawa, Japan. Jpn. J. Infect. Dis. 2009, 62, 220–224. [Google Scholar] [PubMed]
- Nidaira, M.; Taira, K.; Itokazu, K.; Kudaka, J.; Nakamura, M.; Ohno, A.; Takasaki, T. Survey of the antibody against japanese encephalitis virus in Ryukyu wild boars (Sus scrofa riukiuanus) in Okinawa, Japan. Jpn. J. Infect. Dis. 2007, 60, 309–311. [Google Scholar] [PubMed]
- Nidaira, M.; Taira, K.; Itokazu, K.; Okano, S.; Kudaka, J.; Nakamura, M.; Ohno, A.; Takasaki, T. Detection of Japanese encephalitis virus genome in Ryukyu wild boars (Sus scrofa riukiuanus) in Okinawa, Japan. Jpn. J. Infect. Dis. 2008, 61, 164–165. [Google Scholar]
- Yang, D.-K.; Kim, H.-H.; Hyun, B.-H.; Lim, S.-I.; Nam, Y.-K.; Nah, J.-J.; Song, J.-Y. Detection of Neutralizing Antibody Against Japanese Encephalitis Virus in Wild Boars of Korea. J. Bacteriol. Virol. 2012, 42, 353. [Google Scholar] [CrossRef]
- Conlan, J.V.; Vongxay, K.; Jarman, R.G.; Gibbons, R.V.; Lunt, R.A.; Fenwick, S.; Thompson, R.C.A.; Blacksell, S.D. Serologic study of pig-associated viral zoonoses in Laos. Am. J. Trop. Med. Hyg. 2012, 86, 1077–1084. [Google Scholar] [CrossRef]
- Simpson, D.; Smith, C.; Marshall, T.D.C.; Platt, G.; Way, H.; Bowen, E.; Bright, W.; Day, J.; McMahon, D.; Hill, M.; et al. Arbovirus infections in Sarawak: The role of the domestic pig. Trans. R. Soc. Trop. Med. Hyg. 1976, 70, 66–72. [Google Scholar] [CrossRef]
- Myint, L.; Oo, K.; Hlaing, W.; Thu, H.M.; Thant, K.Z.; Than, S.M.; Aye, K.T.; Oo, S.W.; Ha, T.; Moe, K.; et al. Investigation of Japanese encephalitis virus infection in Bogalay Township, Myanmar in 1999. Trop. Med. 2000, 42, 47–52. [Google Scholar]
- Thakur, K.K.; Pant, G.R.; Wang, L.; Hill, C.A.; Pogranichniy, R.M.; Manandhar, S.; Johnson, A.J.; Thakur, K. Seroprevalence of Japanese Encephalitis Virus and Risk Factors Associated with Seropositivity in Pigs in Four Mountain Districts in Nepal. Zoonoses Public Health 2012, 59, 393–400. [Google Scholar] [CrossRef]
- Pant, G.R.; Lunt, R.A.; Rootes, C.L.; Daniels, P.W. Serological evidence for Japanese encephalitis and West Nile viruses in domestic animals of Nepal. Comp. Immunol. Microbiol. Infect. Dis. 2006, 29, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Pant, G.R. A Serological Survey of Pigs, Horses, and Ducks in Nepal for Evidence of Infection with Japanese Encephalitis Virus. Ann. N. Y. Acad. Sci. 2006, 1081, 124–129. [Google Scholar] [CrossRef]
- See, E.; Tan, H.C.; Wang, D.; Ooi, E.E.; Lee, M.-A. Presence of hemagglutination inhibition and neutralization antibodies to Japanese encephalitis virus in wild pigs on an offshore island in Singapore. Acta Trop. 2002, 81, 233–236. [Google Scholar] [CrossRef]
- Zhang, H.; Luo, H.; Rehman, M.U.; Nabi, F.; Li, K.; Lan, Y.; Huang, S.; Zhang, L.; Mehmood, K.; Shahzad, M.; et al. Evidence of JEV in Culex tritaeniorhynchus and pigs from high altitude regions of Tibet, China. J. Vector Borne Dis. 2017, 54, 69–73. [Google Scholar] [PubMed]
- Zhang, H.; Rehman, M.U.; Li, K.; Luo, H.; Lan, Y.; Nabi, F.; Zhang, L.; Iqbal, M.K.; Zhu, S.; Javed, M.T.; et al. Epidemiologic survey of Japanese encephalitis virus infection, Tibet, China, 2015. Emerg. Infect. Dis. 2017, 23, 1023–1024. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-X.; Li, M.-H.; Fu, S.-H.; Chen, W.-X.; Liu, Q.-Y.; Zhang, H.-L.; Da, W.; Hu, S.-L.; La Mu, S.D.; Bai, J.; et al. Japanese Encephalitis, Tibet, China. Emerg. Infect. Dis. 2011, 17, 934–936. [Google Scholar] [CrossRef]
- Ruget, A.S.; Beck, C.; Gabassi, A.; Trévennec, K.; Lecollinet, S.; Chevalier, V.; Cappelle, J. Japanese encephalitis circulation pattern in swine of northern Vietnam and consequences for swine’s vaccination recommendations. Transbound. Emerg. Dis. 2018, 65, 1485–1492. [Google Scholar] [CrossRef]
- Tien, T.V.; Nguyen, H.T.; Mam, V.S.; Nga, P.T.; Igarashi, A. Prevention of Japanese Encephalitis (JE) by “BIKEN” vaccine and epidemiological survey on JE in Dong Anh District, Hanoi, Vietnam. Trop. Med. 1991, 33, 83–91. [Google Scholar]
- Do, Q.H.; Vu, T.Q.H.; Huynh, T.K.L.; Dinh, Q.T.; Deubel, V. Current situation of Japanese encephalitis in the south of Vietnam, 1976–1992. Trop. Med. 1994, 36, 202–214. [Google Scholar]
- Nga, P.T.; Phuong, l.; Nam, V.S.; Yen, N.T.; van Tien, T.; Lien, H.P. Transmission of Japanese encephalitis (JE) virus in Gia Luong District, Ha Bac Province, Vietnam, after JE vaccination, 1993–1994. Trop. Med. 1995, 37, 129–134. [Google Scholar]
- Lindahl, J.F.; Ståhl, K.; Chirico, J.; Boqvist, S.; Thu, H.T.V.; Magnusson, U. Circulation of Japanese Encephalitis Virus in Pigs and Mosquito Vectors within Can Tho City, Vietnam. PLoS Negl. Trop. Dis. 2013, 7, e2153. [Google Scholar] [CrossRef] [PubMed]
- Chan, I.P. Investigation of the HI titre of Japanese encephalitis of swine on Taiwan, Republic of China. Bull. Off. Int. Epizoot. 1968, 69, 141–143. [Google Scholar] [PubMed]
- Borah, J.; Dutta, P.; Khan, S.A.; Mahanta, J. Epidemiological concordance of Japanese encephalitis virus infection among mosquito vectors, amplifying hosts and humans in India. Epidemiol. Infect. 2013, 141, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, H.; Ishida, N.; Akiyama, K.; Shiratori, T.; Konno, J. Epidemiological analyses of Japanese encephalitis virus spread from mosquitoes to pigs through 5 years. Jpn. J. Med. Sci. Boil. 1970, 23, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Maeda, O.; Karaki, T.; Kuroda, A.; Sasaki, O.; Karoji, Y.; Takenokuma, K. Epidemiological studies on japanese encephalitis in kyoto city area, japan iii. seasonal prevalence of virus infections in several pig populations shown by virus recovery from engorged culex tritaeniorhynchus summorosus. Jpn. J. Med. Sci. Boil. 1978, 31, 277–290. [Google Scholar] [CrossRef] [PubMed]
- Konno, J.; Endo, K.; Agatsuma, H.; Ishida, N. Cyclic outbreaks of japanese encephalitis among pigs and humans1. Am. J. Epidemiol. 1966, 84, 292–300. [Google Scholar] [CrossRef]
- Ura, M. Ecology of Japanese encephalitis virus in Okinawa, Japan. II. Regional particularity of the virus dissemination in Okinawa island from 1966 to 1976. Trop. Med. 1976, 18, 165–178. [Google Scholar]
- Van Den Hurk, A.F.; Ritchie, S.A.; Mackenzie, J.S. Ecology and geographical expansion of japanese encephalitis virus. Annu. Rev. Entomol. 2009, 54, 17–35. [Google Scholar] [CrossRef]
- Nitatpattana, N.; Le Flohic, G.; Thongchai, P.; Nakgoi, K.; Palaboodeewat, S.; Khin, M.; Barbazan, P.; Yoksan, S.; Gonzalez, J.P. Elevated Japanese encephalitis virus activity monitored by domestic sentinel piglets in Thailand. Vector Borne Zoonotic Dis. 2011, 11, 391–394. [Google Scholar] [CrossRef]
- Johnsen, D.O.; Edelman, R.; Grossman, R.A.; Muangman, D.; Pomsdhit, J.; Gould, D.J. Study of Japanese encephalitis virus in chiangmai valley, Thailand V. Animal infections. Am. J. Epidemiol. 1974, 100, 57–68. [Google Scholar] [CrossRef]
- Van Peenen, P.F.D.; Joseph, P.L.; Atmosoedjono, S.; Irsiana, R.; Saroso, J.S. Japanese encephalitis virus from pigs and mosquitoes in Jakarta, Indonesia. Trans. R. Soc. Trop. Med. Hyg. 1975, 69, 477–479. [Google Scholar] [CrossRef]
- Makino, Y.; Saito, M.; Phommasack, B.; Vongxay, P.; Kanemura, K.; Pothawan, T.; Insisiengmay, S.; Fukunaga, T. Arbovirus infections in pilot areas in Laos. Trop. Med. 1994, 36, 131–139. [Google Scholar]
- Yoshikawa, A.; Nabeshima, T.; Inoue, S.; Agoh, M.; Morita, K. Molecular and serological epidemiology of Japanese encephalitis virus (JEV) in a remote island of western Japan: An implication of JEV migration over the East China Sea. Trop. Med. Health 2016, 44, 8. [Google Scholar] [CrossRef] [PubMed]
- Ura, M. Ecology of Japanese encephalitis virus in Okinawa, Japan. I. The investigation on pig and mosquito infection of the virus in Okinawa island from 1966 to 1976. Trop. Med. 1976, 18, 151–163. [Google Scholar]
- Grossman, R.A.; Edelman, R.; Gould, D.J. Study of japanese encephalitis virus in chiangmai valley, thailand vi. summary and conclusions1. Am. J. Epidemiol. 1974, 100, 69–76. [Google Scholar] [CrossRef]
- Hasegawa, M.; Nam, V.S.; Tuno, N.; Takagi, M.; Yen, N.T. Influence of the distribution of host species on adult abundance of Japanese encephalitis vectors Culex vishnui subgroup and Culex gelidus in a rice-cultivating village in northern Vietnam. Am. J. Trop. Med. Hyg. 2008, 78, 159–168. [Google Scholar] [CrossRef]
- Ohba, S.-Y.; Huynh, T.T.T.; Le, L.L.; Ngoc, H.T.; Le Hoang, S.; Takagi, M. Mosquitoes and Their Potential Predators in Rice Agroecosystems of the Mekong Delta, Southern Vietnam. J. Am. Mosq. Control. Assoc. 2011, 27, 384–392. [Google Scholar] [CrossRef]
- Ohba, S.; Van Soai n Van Anh, D.T.; Nguyen, Y.T.; Takagi, M. Study of mosquito fauna in rice ecosystems around Hanoi, Northern Vietnam. Acta Trop. 2015, 142, 89–95. [Google Scholar] [CrossRef]
- Murty, U.S.; Rao, M.S.; Arunachalam, N. The effects of climatic factors on the distribution and abundance of Japanese encephalitis vectors in Kurnool district of Andhra Pradesh, India. J. Vector Borne Dis. 2010, 47, 26–32. [Google Scholar]
- Self, L.S.; Shin, H.K.; Kim, K.H.; Lee, K.W.; Chow, C.Y.; Hong, H.K. Ecological studies on Culex tritaeniorhynchus as a vector of Japanese encephalitis. Bull. World Health Organ. 1973, 49, 41–47. [Google Scholar]
- Sudeep, A.B. Culex gelidus: An emerging mosquito vector with potential to transmit multiple virus infections. J. Vector Borne Dis. 2014, 51, 251–258. [Google Scholar] [PubMed]
- Rosen, L. The Natural History of Japanese Encephalitis Virus. Annu. Rev. Microbiol. 1986, 40, 395–414. [Google Scholar] [CrossRef] [PubMed]
- Keiser, J.; Maltese, M.F.; Erlanger, T.E.; Bos, R.; Tanner, M.; Singer, B.H.; Utzinger, J. Effect of irrigated rice agriculture on Japanese encephalitis, including challenges and opportunities for integrated vector management. Acta Trop. 2005, 95, 40–57. [Google Scholar] [CrossRef] [PubMed]
- Morita, K.; Nabeshima, T.; Buerano, C.C. Japanese encephalitis. OIE Rev. Sci. Tech. 2015, 34, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Mansfield, K.L.; Hernández-Triana, L.M.; Banyard, A.C.; Fooks, A.R.; Johnson, N. Japanese encephalitis virus infection, diagnosis and control in domestic animals. Veter Microbiol. 2017, 201, 85–92. [Google Scholar] [CrossRef]
- Daniels, P.T. Arboviruses of Veterinary Significance in The Asia-Western Pacific Region, Such as Japanese Encephalitis Virus; Office International des Epizooties: Paris, France, 2003. [Google Scholar]
- McArthur, M.A.; Holbrook, M.R. Japanese Encephalitis Vaccines. J. Bioterrorism Biodefense 2011, 2. [Google Scholar] [CrossRef]
- Yun, S.-I.; Lee, Y.-M. Japanese encephalitis. Hum. Vaccines Immunother. 2014, 10, 263–279. [Google Scholar] [CrossRef]
- Konishi, E.; Yamaoka, M.; Kurane, I.; Mason, P.W. Japanese Encephalitis DNA Vaccine Candidates Expressing Premembrane and Envelope Genes Induce Virus-Specific Memory B Cells and Long-Lasting Antibodies in Swine. Virology 2000, 268, 49–55. [Google Scholar] [CrossRef]
- García-Nicolás, O.; Ricklin, M.E.; Liniger, M.; Vielle, N.J.; Python, S.; Souque, P.; Charneau, P.; Summerfield, A. A Japanese Encephalitis Virus Vaccine Inducing Antibodies Strongly Enhancing In Vitro Infection Is Protective in Pigs. Viruses 2017, 9, 124. [Google Scholar] [CrossRef]
- Seo, H.-J.; Kim, H.C.; Klein, T.A.; Ramey, A.M.; Lee, J.-H.; Kyung, S.-G.; Park, J.-Y.; Cho, Y.S.; Cho, I.-S.; Yeh, J.-Y. Molecular Detection and Genotyping of Japanese Encephalitis Virus in Mosquitoes during a 2010 Outbreak in the Republic of Korea. PLoS ONE 2013, 8, e55165. [Google Scholar] [CrossRef]
- Nah, J.-J.; Yang, D.-K.; Kim, H.-H.; Song, J.-Y. The present and future of veterinary vaccines for Japanese encephalitis in Korea. Clin. Exp. Vaccine Res. 2015, 4, 130–136. [Google Scholar] [CrossRef]
- Hsu, S.T.; Chang, L.C.; Lin, S.Y.; Chuang, T.Y.; Ma, C.H.; Inoue, Y.K.; Okuno, T. The effect of vaccination with a live attenuated strain of Japanese encephalitis virus on stillbirths in swine in Taiwan. Bull. World Health Organ. 1972, 46, 465–471. [Google Scholar] [PubMed]
- Ogata, M.; Nagao, Y.; Kikui, R.; Kitamura n Ariyasu, H. Effect of complete adjuvant on vaccination of live attenuated Japanese encephalitis vaccine to swine for preventing viremia. 39. Epidemiological studies of J.E. Acta Med. Okayama 1971, 25, 71–75. [Google Scholar]
- Clarke, D.H.; Casals, J. Techniques for Hemagglutination and Hemagglutination-Inhibition with Arthropod-Borne Viruses. Am. J. Trop. Med. Hyg. 1958, 7, 561–573. [Google Scholar] [CrossRef] [PubMed]
- Beck, C.; Lowenski, S.; Durand, B.; Bahuon, C.; Zientara, S.; Lecollinet, S. Improved reliability of serological tools for the diagnosis of West Nile fever in horses within Europe. PLoS Negl. Trop. Dis. 2017, 11, e0005936. [Google Scholar] [CrossRef]
- Kuno, G. Serodiagnosis of flaviviral infections and vaccinations in humans. Adv. Virus Res. 2003, 61, 3–65. [Google Scholar]
- Jain, P.; Jain, A.; Kumar, A.; Prakash, S.; Khan, D.N.; Singh, K.P.; Garg, R.K.; Kumar, R.; Kumar, G.A. Epidemiology and etiology of acute encephalitis syndrome in North India. Jpn. J. Infect. Dis. 2014, 67, 197–203. [Google Scholar] [CrossRef]
- Srey, V.H.; Sadones, H.; Ong, S.; Yim, C.; Reynes, J.-M.; Sor, S.; Mam, M.; Grosjean, P. Etiology of encephalitis syndrome among hospitalized children and adults in Takeo, Cambodia, 1999–2000. Am. J. Trop. Med. Hyg. 2002, 66, 200–207. [Google Scholar] [CrossRef]
- Cao, L.; Fu, S.; Lu, Z.; Tang, C.; Gao, X.; Li, X.; Lei, W.; He, Y.; Li, M.; Cao, Y.; et al. Detection of West Nile Virus Infection in Viral Encephalitis Cases, China. Vector Borne Zoonotic Dis. 2019, 19, 45–50. [Google Scholar] [CrossRef]
- Chang, K.J. Studies on the serological cross-reaction between dengue and Japanese encephalitis. Chin. J. Microbiol. Immunol. 1997, 30, 207–218. [Google Scholar]
- Calisher, C.H.; Karabatsos, N.; Dalrymple, J.M.; Shope, R.E.; Porterfield, J.S.; Westaway, E.G.; Brandt, W.E. Antigenic Relationships between Flaviviruses as Determined by Cross-neutralization Tests with Polyclonal Antisera. J. Gen. Virol. 1989, 70, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Mansfield, K.L.; Horton, D.L.; Johnson, N.; Li, L.; Barrett, A.D.T.; Smith, D.J.; Galbraith, S.E.; Solomon, T.; Fooks, A.R. Flavivirus-induced antibody cross-reactivity. J. Gen. Virol. 2011, 92, 2821–2829. [Google Scholar] [CrossRef] [PubMed]
- Goverdhan, M.K.; Kulkarni, A.B.; Gupta, A.K.; Tupe, C.D.; Rodrigues, J.J. Two-way cross-protection between West Nile and Japanese encephalitis viruses in bonnet macaques. Acta Virol. 1992, 36, 277–283. [Google Scholar] [PubMed]
- Nemeth, N.M.; Bosco-Lauth, A.M.; Bowen, R.A. Cross-Protection Between West Nile and Japanese Encephalitis Viruses in Red-Winged Blackbirds (Agelaius phoeniceus). Avian Dis. Dig. 2009, 4, 421–425. [Google Scholar] [CrossRef] [PubMed]
- Maeki, T.; Tajima, S.; Ikeda, M.; Kato, F.; Taniguchi, S.; Nakayama, E.; Takasaki, T.; Lim, C.-K.; Saijo, M. Analysis of cross-reactivity between flaviviruses with sera of patients with Japanese encephalitis showed the importance of neutralization tests for the diagnosis of Japanese encephalitis. J. Infect. Chemother. 2019, 25, 786–790. [Google Scholar] [CrossRef]
- Reuben, R.; Thenmozhi, V.; Samuel, P.P.; Gajanana, A.; Mani, T.R. Mosquito Blood Feeding Patterns as a Factor in the Epidemiology of Japanese Encephalitis in Southern India. Am. J. Trop. Med. Hyg. 1992, 46, 654–663. [Google Scholar] [CrossRef]
- Bhattacharyya, D.R.; Handique, R.; Dutta, L.P.; Dutta, P.; Doloi, P.; Goswami, B.K.; Sharma, C.K.; Mahanta, J. Host feeding patterns of Culex vishnui sub group of mosquitoes in Dibrugarh district of Assam. J. Commun. Dis. 1994, 26, 133–138. [Google Scholar]
- Arunachalam, N.; Samuel, P.P.; Hiriyan, J.; Rajendran, R.; Dash, A.P. Short report: Observations on the multiple feeding behavior of Culex tritaeniorhynchus (Diptera: Culicidae), the vector of Japanese encephalitis in Kerala in southern India. Am. J. Trop. Med. Hyg. 2005, 72, 198–200. [Google Scholar] [CrossRef]
- Wang, L.Y. Host preference of mosquito vectors of Japanese encephalitis. Chin. J. Microbiol. 1975, 8, 274–279. [Google Scholar]
- Samuel, P.P.; Arunachalam, N.; Hiriyan, J.; Tyagi, B.K. Host Feeding Pattern of Japanese Encephalitis Virus Vector Mosquitoes (Diptera: Culicidae) from Kuttanadu, Kerala, India. J. Med. Èntomol. 2008, 45, 927–932. [Google Scholar] [CrossRef]
- Tuno, N.; Tsuda, Y.; Takagi, M. How Zoophilic Japanese Encephalitis Vector Mosquitoes Feed on Humans. J. Med Entomol. 2017, 54, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Yap, G.; Lim, X.F.; Chan, S.; How, C.B.; Humaidi, M.; Yeo, G.; Mailepessov, D.; Kong, M.; Lai, Y.L.; Okumura, C.; et al. Serological evidence of continued Japanese encephalitis virus transmission in Singapore nearly three decades after end of pig farming. Parasites Vectors 2019, 12, 244. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, F.; Guttikar, S.; Pinto, B. Prevalence of antibodies to Japanese encephalitis and West Nile viruses among wild birds in the Krishna-Godavari Delta, Andhra Pradesh, India. Trans. R. Soc. Trop. Med. Hyg. 1981, 75, 258–262. [Google Scholar] [CrossRef]
- Saito, M.; Osa, Y.; Asakawa, M. Antibodies to Flaviviruses in Wild Ducks Captured in Hokkaido, Japan: Risk Assessment of Invasive Flaviviruses. Vector Borne Zoonotic Dis. 2009, 9, 253–258. [Google Scholar] [CrossRef]
- Buescher, E.L.; Scherer, W.F.; Rosenberg, M.Z.; Kutner, L.J.; McClure, H.E. Immunologic Studies of Japanese Encephalitis Virus in Japan: IV. Maternal Antibody in Birds. J. Immunol. 1959, 83, 614–619. [Google Scholar]
- Scherer, W.F.; Buescher, E.L.; McClure, H.E. Ecologic Studies of Japanese Encephalitis Virus in Japan. Am. J. Trop. Med. Hyg. 1959, 8, 689–697. [Google Scholar] [CrossRef]
- Jamgaonkar, A.V.; Yergolkar, P.N.; Geevarghese, G.; Joshi, G.D.; Joshi, M.V.; Mishra, A.C. Serological evidence for Japanese encephalitis virus and West Nile virus infections in water frequenting and terrestrial wild birds in Kolar District, Karnataka State, India. A retrospective study. Acta Virol. 2003, 47, 185–188. [Google Scholar]
- Yang, D.-K.; Oh, Y.-I.; Kim, H.-R.; Lee, Y.-J.; Moon, O.-K.; Yoon, H.; Kim, B.; Lee, K.-W.; Song, J.-Y. Serosurveillance for Japanese encephalitis virus in wild birds captured in Korea. J. Veter Sci. 2011, 12, 373–377. [Google Scholar] [CrossRef]
- Lord, J.S.; Gurley, E.S.; Pulliam, J.R.C. Rethinking Japanese Encephalitis Virus Transmission: A Framework for Implicating Host and Vector Species. PLoS Negl. Trop. Dis. 2015, 9, e0004074. [Google Scholar] [CrossRef]
- Gresser, I.; Hardy, J.L.; Hu, S.M.K.; Scherer, W.F. Factors Influencing Transmission of Japanese B Encephalitis Virus by a Colonized Strain of Culex Tritaeniorhynchus Giles, from Infected Pigs and Chicks to Susceptible Pigs and Birds. Am. J. Trop. Med. Hyg. 1958, 7, 365–373. [Google Scholar] [CrossRef]
- Hauerswald, H. Serological evidence for Japanese encephalitis and West Nile virus infections in domestic birds in Cambodia. In preparation.
- Dhanda, V.; Banerjee, K.; Deshmukh, P.K.; AIlkal, M. Experimental viraemia and transmission of Japanese encephalitis virus by mosquitoes in domestic ducks. Indian J. Med. Res. 1977, 66, 881–888. [Google Scholar] [PubMed]
- Frantz, L.; Meijaard, E.; Gongora, J.; Haile, J.; Groenen, M.A.M.; Larson, G. The Evolution of Suidae. Annu. Rev. Anim. Biosci. 2016, 4, 61–85. [Google Scholar] [CrossRef] [PubMed]
- Nitatpattana, N.; Apiwathnasorn, C.; Barbazan, P.; Leemingsawat, S.; Yoksan, S.; Gonzalez, J.L.Z. First isolation of Japanese encephalitis from Culex quinquefasciatus in Thailand. Southeast Asian J. Trop. Med. Public Health 2005, 36, 875–878. [Google Scholar]
- Thoa, N.T.K.; Vien, N.T.; Mai, T.T.; Xuan, N.T.N. Japanese encephalitis vectors: Isolation of virus from culicine mosquitoes in the Saigon area. Southeast Asian J. Trop. Med. Public Health 1974, 5, 408–412. [Google Scholar]
- Lindahl, J.; Chirico, J.; Boqvist, S.; Thu, H.T.V.; Magnusson, U. Occurrence of Japanese encephalitis virus mosquito vectors in relation to urban pig holdings. Am. J. Trop. Med. Hyg. 2012, 87, 1076–1082. [Google Scholar] [CrossRef]
- Huynh, T.T.T.; Aarnink, A.J.A.; Drucker, A.; Verstegen, M.W.A. Pig Production in Cambodia, Laos, Philippines, and Vietnam: A Review. Asian J. Agric. Dev. 2006, 3, 69–90. [Google Scholar]
- Diallo, A.O.I.; Chevalier, V.; Cappelle, J.; Duong, V.; Fontenille, D.; Duboz, R. How much does direct transmission between pigs contribute to Japanese Encephalitis virus circulation? A modelling approach in Cambodia. PLoS ONE 2018, 13, e0201209. [Google Scholar] [CrossRef]
- Zhao, S.; Lou, Y.; Chiu, A.P.; He, D. Modelling the skip-and-resurgence of Japanese encephalitis epidemics in Hong Kong. J. Theor. Boil. 2018, 454, 1–10. [Google Scholar] [CrossRef]
- Sunwoo, J.-S.; Jung, K.-H.; Lee, S.-T.; Lee, S.K.; Chu, K. Reemergence of Japanese Encephalitis in South Korea, 2010–2015. Emerg. Infect. Dis. 2016, 22, 1841–1843. [Google Scholar] [CrossRef]
- Tandale, B.V.; Khan, S.A.; Kushwaha, K.P.; Rahman, H.; Gore, M.M.; Sapkal, G.N.; Bondre, V.P.; Srivastava, D.; Laskar, B.; Mahanta, J. Effectiveness of Japanese encephalitis SA 14-14-2 live attenuated vaccine among Indian children: Retrospective 1:4 matched case-control study. J. Infect. Public Health 2018, 11, 713–719. [Google Scholar] [CrossRef]
- Khan, S.U.; Salje, H.; Hannan, A.; Islam, M.A.; Bhuyan, A.A.M.; Islam, M.A.; Rahman, M.Z.; Nahar, N.; Hossain, M.J.; Luby, S.P.; et al. Dynamics of Japanese Encephalitis Virus Transmission among Pigs in Northwest Bangladesh and the Potential Impact of Pig Vaccination. PLoS Negl. Trop. Dis. 2014, 8, e3166. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.J.; Kang, B.J.; Lim, Y.M.; Lee, C.K. Studies on Japanese encephalitis live vaccine. II. Development of an attenuated strain of virus (Anyang strain). Res. Rep. Off. Rural Dev. 1975, 17, 95–100. [Google Scholar]
- Chen, Y.-Y.; Fan, Y.-C.; Tu, W.-C.; Chang, R.-Y.; Shih, C.-C.; Lu, I.-H.; Chien, M.-S.; Lee, W.-C.; Chen, T.-H.; Chang, G.-J.; et al. Japanese Encephalitis Virus Genotype Replacement, Taiwan, 2009–2010. Emerg. Infect. Dis. 2011, 17, 2354–2356. [Google Scholar] [CrossRef] [PubMed]
- Schuh, A.J.; Ward, M.J.; Brown, A.J.L.; Barrett, A.D.T. Dynamics of the Emergence and Establishment of a Newly Dominant Genotype of Japanese Encephalitis Virus throughout Asia. J. Virol. 2014, 88, 4522–4532. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Fu, S.; Gao, X.; Li, M.; Cui, S.; Li, X.; Cao, Y.; Lei, W.; Lu, Z.; He, Y.; et al. Low Protective Efficacy of the Current Japanese Encephalitis Vaccine against the Emerging Genotype 5 Japanese Encephalitis Virus. PLoS Negl. Trop. Dis. 2016, 10, e0004686. [Google Scholar] [CrossRef]
- Tajima, S.; Moi, M.L.; Yagasaki, K.; Nakayama, E.; Takasaki, T.; Saijo, M.; Tomikawa, T.; Kurane, I.; Kotaki, A.; Lim, C.-K. In vitro growth, pathogenicity and serological characteristics of the Japanese encephalitis virus genotype V Muar strain. J. Gen. Virol. 2015, 96, 2661–2669. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.R.S.; Cohnstaedt, L.W.; Cernicchiaro, N. Japanese Encephalitis Virus: Placing Disease Vectors in the Epidemiologic Triad. Ann. Èntomol. Soc. Am. 2018, 111, 295–303. [Google Scholar] [CrossRef]
- Diallo, A.O.; Chevalier, V.; Cappelle, J.; Duboz, R.; Didier, F.; Benoit, D. Modelling and assessment of combining gilt vaccination, vector control and pig herd management to control Japanese Encephalitis virus transmission in Southeast Asia. BioRxiv 2018. [Google Scholar] [CrossRef]
- Ben-Dov, E. Bacillus thuringiensis subsp. israelensis and Its Dipteran-Specific Toxins. Toxins 2014, 6, 1222–1243. [Google Scholar] [CrossRef]
- Wiwattanawanichakun, P.; Ratwatthananon, A.; Poonsri, W.; Yooboon, T.; Pluempanupat, W.; Piyasaengthong, N.; Nobsathian, S.; Bullangpoti, V. The Possibility of Using Isolated Alkaloid Compounds and Crude Extracts of Piper retrofractum (Piperaceae) as Larvicidal Control Agents for Culex quinquefasciatus (Diptera: Culicidae) Larvae. J. Med. Èntomol. 2018, 55, 1231–1236. [Google Scholar] [CrossRef]
- Govindarajan, M.; Vaseeharan, B.; Alharbi, N.S.; Kadaikunnan, S.; Khaled, J.M.; Al-Anbr, M.N.; Alyahya, S.A.; Maggi, F.; Benelli, G. High efficacy of (Z)-γ-bisabolene from the essential oil of Galinsoga parviflora (Asteraceae) as larvicide and oviposition deterrent against six mosquito vectors. Environ. Sci. Pollut. Res. 2018, 25, 10555–10566. [Google Scholar] [CrossRef] [PubMed]
- Dutta, P.; Khan, S.A.; Khan, A.M.; Borah, J.; Sarmah, C.K.; Mahanta, J. The Effect of Insecticide-Treated Mosquito Nets (ITMNs) on Japanese Encephalitis Virus Seroconversion in Pigs and Humans. Am. J. Trop. Med. Hyg. 2011, 84, 466–472. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Protocol | Viraemia | Immune Response | Reference | |||
|---|---|---|---|---|---|---|
| Test | Detection Period | Peak Value | Test | Antibody Detection | ||
| 5 sows, IV inoculation, Kanagawa strain, bled daily | ICISM | 1–4 dpi | NA | VN in mice | >7 dpi | [51] |
| 4 piglets, SC inoculation, M5/596 and pig9 strains, bled daily | ICISM | For 4 days | 2.6 log LD50/0.03 mL | HIA | 7–35 * dpi | [47] |
| 2 piglets, SC inoculation, 9215 strain, bled daily | ICISM | 1–4 dpi | NA | HIA | All positive at 7 dpi | [52] |
| 6 piglets, SC inoculation, Nakayama strain, bled daily | ICISM + RT-PCR | 2–5 dpi | NA | SNT | All positive at 14 dpi | [53] |
| 12 piglets, IV + ID inoculation, Nakayama strain, bled daily | RT-PCR | 1–5 dpi | 10^4 U/mL ** | SNT | 3–11 * dpi | [22] |
| 10 piglets, IV inoculation, JE-91 strain, bled daily | RT-PCR | 3–5 dpi | 10^3.5 U/mL ** | SNT | All positive at 28 dpi | [54] |
| Method | Clinical Signs | Macroscopic Lesions | Microscopic Lesions | Reference |
|---|---|---|---|---|
| 5 pregnant sows, IV inoculation, Fuji strain, bled daily until farrow | No | / | / | [51] 1st experiment |
| 6 pregnant sows, IV inoculation, Kanagawa strain, bled daily until farrow | Mummified and hydrocephalic fetuses in 2/6 litters | / | / | [51] 2nd experiment |
| 14 piglets, IV inoculation, clinical signs monitoring and histopathology after euthanasia | Fever until 4 dpi, depression and hind limbs tremor | No | Non-suppurative encephalitis with perivascular cuffing of mononuclear cells and multifocal gliosis in grey and white matter cerebrum | [62] |
| 10 piglets, intranasal inoculation, clinical signs monitoring and histopathology after euthanasia | Fever until 4 dpi, depression and slight hind limbs tremor on 4 piglets until 10 dpi maximum | No | Non-suppurative encephalitis with perivascular cuffing of lymphocytes, multifocal gliosis, neuronal degeneration, and necrosis | [63] |
| 12 piglets, IV and ID inoculation, clinical signs monitoring, histopathology after euthanasia and RT-PCR on tissues | Fever until 5 dpi, reduce appetite, less manure, and reluctance to move until 6 to 9 dpi | No | Signs of viral meningoencephalomyelitis | [22] |
| 10 piglets, IV inoculation, clinical signs monitoring, histopathology after euthanasia and RT-PCR on tissues | Fever until 5 dpi, mild depression and lethargy until 5 dpi, mild ataxia between 10 and 13 dpi, 2 pigs with hind limb ataxia between 19 and 27 dpi | No | No | [54] |
| Lab Method | Material | Result | Differential Diagnosis Intention | Reproductive Failure | Country | Reference |
|---|---|---|---|---|---|---|
| RT-PCR | Aborted fetuses | Detection | No | No quantitative data | China | [68] |
| RT-PCR | Sample of 37 CSF of aborted piglets | 5/37 | No | No quantitative data | China | [69] |
| ICISM, virus isolation, RT-PCR | Sample of 108 brain tissues of stillborn piglets | 20/108 | No | No quantitative data | China | [70] |
| RT-PCR | Sample of 3 brain samples of stillborn piglets | 3/3 | CSFV, PRRSV, PRV, PPV not detected | 30 sows with RF/200 sows (15%) | China | [64] |
| RT-PCR | 31 brain samples of stillborn piglets (all stillborn piglets of the farm) | 7/31 | No | 10 sows with RF/28 sows (36%), 2–5 sb/sow, 31 sb in total | India | [65] |
| RT-PCR | Sample of 8 brain samples of stillborn piglets | 8/8 | CSFV, PRRSV, PRV, PPV not detected | 37 sows with RF/128 sows (29%) | China | [66] |
| HIA on body fluids and virus isolation (unspecified method) | Aborted fetuses | Isolation on “some” fetuses | No | 50 sows with RF/320 sows (3 farms) (16%) | Japan | [50] |
| Protocol | Test | Average Age of Waning of Anti-JEV Maternal Antibodies | Country | Reference |
|---|---|---|---|---|
| 80 piglets, 2–7 months old, from farms, bled monthly | HIA | >4 months old | Japan | [47] |
| 9 piglets, in mosquito traps, bled monthly | HIA | >1.5 months old | Japan | [47] |
| 2 cohorts of 15 piglets, 2 months old, bled every 10 days for 4 months | IgG ELISA | >3 months old | Cambodia, peri-urban | [74] |
| 2 cohorts of 15 piglets, 2 months old, bled every 10 days for 4 months | IgG ELISA | Peri-urban: >2 months oldRural: >3.5 months old | Cambodia, peri-urban and rural | [75] |
| 5 piglets, 2 months old, bled every month for 3 years | HIA | >2 months old | South India | [61] |
| Infection Route | Test for Viraemia and Oro-Nasal Fluids | Clinical Signs | Viraemia | Oro-Nasal Shedding | Reference |
|---|---|---|---|---|---|
| Contact with infected pigs in vector-free buildings | RT-PCR | Yes | Yes, 3–10 dac ~ 104 U/mL | Yes, 5–10 dac ~101.5 U/mL | [21] 1st experience |
| Oro-nasal inoculation | RT-PCR | Yes | Yes, 1–9 dpi ~103.5 U/mL | Yes, 3–9 dpi ~103 U/mL | [21] 2nd experience |
| Intranasal inoculation | / | Yes | / | / | [63] |
| Sampling Region | Sampling Year | Origin of Sampled Animals (A/F if Pigs) | Age of Sampled Animals | Tested Animals | Serological Test | Anti-JEV Antibodies Evidence | Confirmation Test and Result | Reference | Older References |
|---|---|---|---|---|---|---|---|---|---|
| Australia (T.) | 1995 | F | NS | 90 | HIA | 63/90 | SNT + | [34] | |
| Australia (T.) | 1998 | Feral pigs | NS | 113 | HIA | 90/113 | nd | [83] | |
| Cambodia | 2007 | A and F | ~4.3 m (20 d–12 m) | 505 | HIA and IgG ELISA | 65.7% and 63.5% | nd | [80] | [74,75]* |
| Hong Kong | 1968 | F | NS | 558 | HIA | 60,4% | nd | [84] | |
| India | 2014 | F | >3 m | 51 | IgG ELISA | 35/51 | RT-PCR + | [85] | [86,87,88] |
| Indonesia | 2015 | F | NS | 80 | IgG ELISA | 32/80 | nd | [89] | [90,91] |
| Japan (M.) | 2008 | Wild boars | NS | 36 | SNT | 30/36 | SNT + | [92] | [93,94] |
| Japan (Is.) | 2010 | F | NS | 128 | HIA | 3.1% | RT-PCR - | [95] | [96,97]** |
| Japan (Ir.) | 2010 | Wild boars | NS | 117 | HIA | 44.4% | RT-PCR - | [95] | [98,99] |
| South-Korea | 2011 | Wild boars | NS | 288 | SNT | 66% | SNT + | [100] | |
| Laos | 2009 | A | 4–12 m | 727 | HIA | 74.7% | nd | [101] | |
| Malaysia | 2016 | F | NS | 90 | IgG ELISA | 40/90 | nd | [31] | [102] |
| Myanmar | 1999 | F | NS | 36 | HIA | 12/36 | nd | [103] | |
| Nepal | 2010 | F | 4–48 m | 454 | IgG ELISA | 16.7% | nd | [104] | [105,106] |
| Singapore | 1999 | Wild boars | NS | 28 | HIA | 28/28 | SNT + | [107] | |
| Sri Lanka | 1988 | F | 1–24 m | 951 | SNT | 32.6% | SNT + | [82] | |
| Thailand | 1983 | A | 4–12 m | 100 | HIA | 74% | ICISM + | [81] | [13] |
| China (Tibet) | 2015 | A | 1–6 m | 102 | IgM ELISA | nr | RT-PCR + | [108] | [109,110] |
| Vietnam | 2010 | A | 4–8 m | 641 | IgG ELISA | 60.4% | SNT + | [111] | [67,112,113,114,115] |
| Taiwan | 1966 | F | 3–8 m | 6000 | HIA | 37.3% | nd | [116] |
| Country/Region | Sampling Protocol (Longitudinal or Cross-Sectional (Abattoir or Farm)) | Detected Seasonality in Pig Infection | Corresponding Climate | References |
|---|---|---|---|---|
| Cambodia | Longitudinal1 | All year | Not relevant ** | [74,75] |
| North Vietnam | Abattoir 2 | February–October * | Summer/rainy season | [114] |
| January–October * | [112] | |||
| South Vietnam | Farms 1 | All year | Not relevant ** | [67] |
| Laos | Abattoir 1 | June–July * | Summer/rainy season | [101] |
| August * | [126] | |||
| Malaysia | Abattoir 2,3 | November–January * | Summer/rainy season | [102] |
| Indonesia | Longitudinal 2,4 | October–March * | Summer/rainy season | [125] |
| Thailand | ||||
| Bangkok | Longitudinal 2,1 | February–May* | Hot and dry season | [14,123] |
| Chiang Mai | Longitudinal 2,3 | May–July | Hot/rainy season | [124,129] |
| Taiwan | Longitudinal 2,4 | March–October, peak in August/July* | Summer monsoon | [73,78,116] |
| Japan, main Island | Abattoir 1,5 | July–November, peak in August/September | Summer (rainfall) to fall (typhoons) | [127] |
| Abattoir 2 | June–December | [50] | ||
| Abattoir 2,4 | July–August | [118] | ||
| Abattoir 2 | May-March, peak in August | [120] | ||
| Japan, Okinawa | Abattoir 2 | August | Tropical climate | [121,128] |
| Abattoir 2,4 | April–October | |||
| North East India | Longitudinal 2 | June–August | Monsoon | [117] |
| Sri Lanka | ||||
| Dry zone | Longitudinal 3 | October–November | Hot/rainy season | [82] |
| Wet zone | March–April | |||
| North Australia | Longitudinal 2,3 | February–April | Hot/rainy season | [83] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ladreyt, H.; Durand, B.; Dussart, P.; Chevalier, V. How Central Is the Domestic Pig in the Epidemiological Cycle of Japanese Encephalitis Virus? A Review of Scientific Evidence and Implications for Disease Control. Viruses 2019, 11, 949. https://doi.org/10.3390/v11100949
Ladreyt H, Durand B, Dussart P, Chevalier V. How Central Is the Domestic Pig in the Epidemiological Cycle of Japanese Encephalitis Virus? A Review of Scientific Evidence and Implications for Disease Control. Viruses. 2019; 11(10):949. https://doi.org/10.3390/v11100949
Chicago/Turabian StyleLadreyt, Héléna, Benoit Durand, Philippe Dussart, and Véronique Chevalier. 2019. "How Central Is the Domestic Pig in the Epidemiological Cycle of Japanese Encephalitis Virus? A Review of Scientific Evidence and Implications for Disease Control" Viruses 11, no. 10: 949. https://doi.org/10.3390/v11100949
APA StyleLadreyt, H., Durand, B., Dussart, P., & Chevalier, V. (2019). How Central Is the Domestic Pig in the Epidemiological Cycle of Japanese Encephalitis Virus? A Review of Scientific Evidence and Implications for Disease Control. Viruses, 11(10), 949. https://doi.org/10.3390/v11100949