Methylation Status of the Adeno-Associated Virus Type 2 (AAV2)

 ,
,
Abstract
Funding
Acknowledgments
Conflicts of Interest
References
- Tattersall, P. The Evolution of Parvovirus Taxonomy. In Parvoviruses; Kerr, J.R., Cotmore, S.F., Bloom, M.E., Linden, R.M., Parrish, C.R., Eds.; Hodder Arnold: London, UK, 2006. [Google Scholar]
- Tóth, R.; Mészáros, I.; Stefancsik, R.; Bartha, D.; Bálint, Á.; Zádori, Z. CpG distribution and methylation pattern in porcine parvovirus. PLoS ONE 2013, 8, e85986. [Google Scholar] [CrossRef]
- Carter, B.J. Adeno-Associated Virus Helper Functions. In Handbook of Parvoviruses; Tijssen, P., Ed.; CRC-Press: Boca Raton, FL, USA, 1989. [Google Scholar]
- Hüser, D.; Gogol-Döring, A.; Lutter, T.; Weger, S.; Winter, K.; Hammer, E.M.; Cathomen, T.; Reinert, K.; Heilbronn, R. Integration preferences of wildtype AAV-2 for consensus rep-binding sites at numerous loci in the human genome. PLoS Pathog. 2010, 6, e1000985. [Google Scholar] [CrossRef]
- Rabinowitz, J.E.; Samulski, J. Adeno-associated virus expression systems for gene transfer. Curr. Opin. Biotechnol. 1998, 9, 470–475. [Google Scholar] [CrossRef]
- Favre, D.; Blouin, V.; Provost, N.; Spisek, R.; Porrot, F.; Bohl, D.; Marme, F.; Cherel, Y.; Salvetti, A.; Hurtrel, B.; et al. Lack of an Immune Response against the Tetracycline-Dependent Transactivator Correlates with Long-Term Doxycycline-Regulated Transgene Expression in Nonhuman Primates after Intramuscular Injection of Recombinant Adeno-Associated Virus. J. Virol. 2002, 76, 11605–11611. [Google Scholar] [CrossRef]
- Rivera, V.M.; Gao, G.P.; Grant, R.L.; Schnell, M.A.; Zoltick, P.W.; Rozamus, L.W.; Clackson, T.; Wilson, J.M. Long-term pharmacologically regulated expression of erythropoietin in primates following AAV-mediated gene transfer. Blood 2005, 105, 1424–1430. [Google Scholar] [CrossRef]
- Attwood, J.T.; Yung, R.L.; Richardson, B.C. DNA methylation and the regulation of gene transcription. Cell. Mol. Life Sci. 2002, 59, 241–257. [Google Scholar] [CrossRef]
- Léger, A.; Guiner, C.; Nickerson, M.L.; Im, K.; Ferry, N.; Moullier, P.; Snyder, R.O.; Penaud-Budloo, M. Adeno-associated viral vector-mediated transgene expression is independent of dna methylation in primate liver and skeletal muscle. PLoS ONE 2011, 6, e20881. [Google Scholar] [CrossRef]
- Hüser, D.; Weger, S.; Heilbronn, R. Kinetics and frequency of adeno-associated virus site-specific integration into human chromosome 19 monitored by quantitative real-time PCR. J. Virol. 2002, 76, 7554–7559. [Google Scholar] [CrossRef]
- Heilbronn, R.; Bürkle, A.; Stephan, S.; zur Hausen, H. The adeno-associated virus rep gene suppresses herpes simplex virus-induced DNA amplification. J. Virol. 1990, 64, 3012–3018. [Google Scholar]
- Grimm, D.; Kern, A.; Rittner, K.; Kleinschmidt, J.A. Novel Tools for Production and Purification of Recombinant Adenoassociated Virus Vectors. Hum. Gene Ther. 1998, 9, 2745–2760. [Google Scholar] [CrossRef]
- Berns, K.I.; Pinkerton, T.C.; Thomas, G.F.; Hoggan, M.D. Detection of adeno-associated virus (AAV)-specific nucleotide sequences in DNA isolated from latently infected Detroit 6 cells. Virology 1975, 68, 556–560. [Google Scholar] [CrossRef]
- Li, L.C.; Dahiya, R. MethPrimer: Designing primers for methylation PCRs. Bioinformatics 2002, 11, 1427–1431. [Google Scholar] [CrossRef]
- Ruffing, M.; Heid, H.; Kleinschmidt, J.A. Mutations in the carboxy terminus of adeno-associated virus 2 capsid proteins affect viral infectivity: Lack of an RGD integrin-binding motif. J. Gen. Virol. 1994, 75, 3385–3392. [Google Scholar] [CrossRef]
- Cao, M.; You, H.; Hermonat, P.L. The X gene of Adeno-Associated Virus 2 (AAV2) is involved in viral DNA replication. PLoS ONE 2014, 9, e104596. [Google Scholar] [CrossRef]
- Stutika, C.; Gogol-Döring, A.; Botschen, L.; Mietzsch, M.; Weger, S.; Feldkamp, M.; Chen, W.; Heilbronn, R. A Comprehensive RNA Sequencing Analysis of the Adeno-Associated Virus (AAV) Type 2 Transcriptome Reveals Novel AAV Transcripts, Splice Variants, and Derived Proteins. J. Virol. 2016, 90, 1278–1289. [Google Scholar] [CrossRef]
- Bonvicini, F.; Manaresi, E.; Di Furio, F.; de Falco, L.; Gallinella, G. Parvovirus B19 DNA CpG dinucleotide methylation and epigenetic regulation of viral expression. PLoS ONE 2012, 7, e33316. [Google Scholar] [CrossRef]
- Hoelzer, K.; Shackelton, L.A.; Parrish, C.R. Presence and role of cytosine methylation in DNA viruses of animals. Nucleic Acids Res. 2008, 9, 2825–2837. [Google Scholar] [CrossRef]
- Shalginskikh, N.; Poleshko, A.; Skalka, A.M.; Katz, R.A. Retroviral DNA Methylation and Epigenetic Repression Are Mediated by the Antiviral Host Protein Daxx. J. Virol. 2013, 87, 2137–2150. [Google Scholar] [CrossRef]
- Hüser, D.; Gogol-Doring, A.; Chen, W.; Heilbronn, R. Adeno-Associated Virus Type 2 Wild-Type and Vector-Mediated Genomic Integration Profiles of Human Diploid Fibroblasts Analyzed by Third-Generation PacBio DNA Sequencing. J. Virol. 2014, 88, 11253–11263. [Google Scholar] [CrossRef]
- Deaton, A.M.; Bird, A. CpG islands and the regulation of transcription. Genes Dev. 2011, 25, 1010–1022. [Google Scholar] [CrossRef]
- Jones, P.A. Functions of DNA methylation: Islands, start sites, gene bodies and beyond. Nat. Rev. Genet. 2012, 13, 484–492. [Google Scholar] [CrossRef]
- Snyder, R.O.; Im, D.S.; Muzyczka, N. Evidence for covalent attachment of the adeno-associated virus (AAV) rep protein to the ends of the AAV genome. J. Virol. 1990, 64, 6204–6213. [Google Scholar]
- Ward, P.; Urcelay, E.; Kotin, R.; Safer, B.; Berns, K.I. Adeno-associated virus DNA replication in vitro: Activation by a maltose binding protein/Rep 68 fusion protein. J. Virol. 1994, 68, 6029–6037. [Google Scholar]
- Urcelay, E.; Ward, P.; Wiener, S.M.; Safer, B.; Kotin, R.M. Asymmetric replication in vitro from a human sequence element is dependent on adeno-associated virus Rep protein. J. Virol. 1995, 69, 2038–2046. [Google Scholar]
- Linden, R.M.; Winocour, E.; Berns, K.I. The recombination signals for adeno-associated virus site-specific integration. Proc. Natl. Acad. Sci. USA 1996, 93, 7966–7972. [Google Scholar] [CrossRef]
- Urabe, M.; Hasumi, Y.; Kume, A.; Surosky, R.T.; Kurtzman, G.J.; Tobita, K.; Ozawa, K. Charged-to-alanine scanning mutagenesis of the N-terminal half of adeno-associated virus type 2 Rep78 protein. J. Virol. 1999, 73, 2682–2693. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Xiao, X.; Li, J.; Samulski, R.J. Production of High-Titer Recombinant Adeno-Associated Virus Vectors in the Absence of Helper Adenovirus. J. Virol. 1998, 72, 2224–2232. [Google Scholar] [CrossRef]
- Cheung, A.K.M.; Hoggan, M.D.; Hauswirth, W.W.; Berns, K.I. Integration of the Adeno-Associated Virus Genome into Cellular DNA in Latently Infected Human Detroit 6 Cells. J. Virol. 1980, 33, 739–748. [Google Scholar] [CrossRef]
- Musatov, S.A.; Dudus, L.; Parrish, C.M.; Scully, T.A.; Fisher, K.J. Spontaneous mobilization of integrated recombinant adenoassociated virus in a cell culture model of virus latency. Virology 2002, 294, 151–169. [Google Scholar] [CrossRef]
- Smith-Moore, S.; Neil, S.J.D.; Fraefel, C.; Linden, R.M.; Bollen, M.; Rowe, H.M.; Henckaerts, E. Adeno-associated virus Rep proteins antagonize phosphatase PP1 to counteract KAP1 repression of the latent viral genome. Proc. Natl. Acad. Sci. USA 2018, 115, E3529–E3538. [Google Scholar] [CrossRef]
- Mazzio, E.A.; Soliman, K.F.A. Basic concepts of epigenetics impact of environmental signals on gene expression. Epigenetics 2012, 7, 119–130. [Google Scholar] [CrossRef]
- Penaud-Budloo, M.; Le Guiner, C.; Nowrouzi, A.; Toromanoff, A.; Cherel, Y.; Chenuaud, P.; Schmidt, M.; von Kalle, C.; Rolling, F.; Moullier, P.; et al. Adeno-Associated Virus Vector Genomes Persist as Episomal Chromatin in Primate Muscle. J. Virol. 2008, 16, 7875–7885. [Google Scholar] [CrossRef]
- Reeves, R.; Gorman, C.M.; Howard, B. Minichromosome assembly of non-integrated plasmid DNA transfected into mammalian cells. Nucleic Acids Res. 1985, 13, 3599–3615. [Google Scholar] [CrossRef]
- Chang, L.-S.; Shi, Y.; Shenk, T. Adeno-Associated Virus P5 Promoter Contains an Adenovirus ElA-Inducible Element and a Binding Site for the Major Late Transcription Factor. J. Virol. 1989, 63, 3479–3488. [Google Scholar]
- Shi, Y.; Seto, E.; Chang, L.S.; Shenk, T. Transcriptional repression by YY1, a human GLI-Krüppel-related protein, and relief of repression by adenovirus E1A protein. Cell 1991, 67, 377–388. [Google Scholar] [CrossRef]
- Kim, J.; Kollhoff, A.; Bergmann, A.; Stubbs, L. Methylation-sensitive binding of transcription factor YY1 to an insulator sequence within the paternally expressed imprinted gene, Peg3. Hum. Mol. Genet. 2003, 12, 233–245. [Google Scholar] [CrossRef]
- Molloy, P.L.; Watt, F. DNA methylation and specific protein-DNA interactions. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1990, 326, 267–275. [Google Scholar] [CrossRef]
- Bird, A. DNA methylation patterns and epigenetic memory. Genes Dev. 2002, 6–21. [Google Scholar] [CrossRef]
- Medvedeva, Y.A.; Khamis, A.M.; Kulakovskiy, I.V.; Ba-Alawi, W.; Bhuyan, M.S.I.; Kawaji, H.; Lassmann, T.; Harbers, M.; Forrest, A.R.R.; Bajic, V.B. Effects of cytosine methylation on transcription factor binding sites. BMC Genom. 2014, 15, 119. [Google Scholar] [CrossRef]



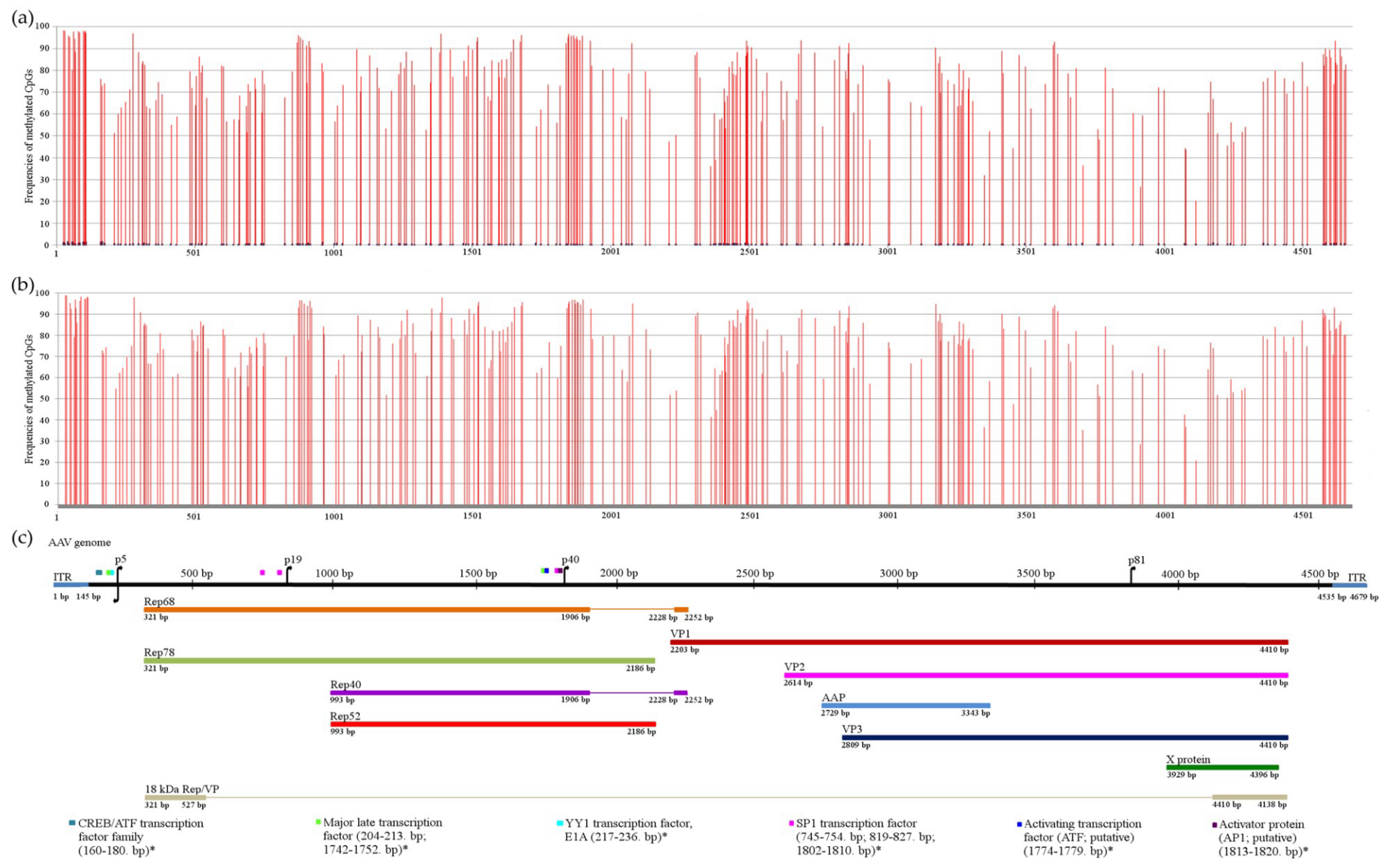{kind=link}
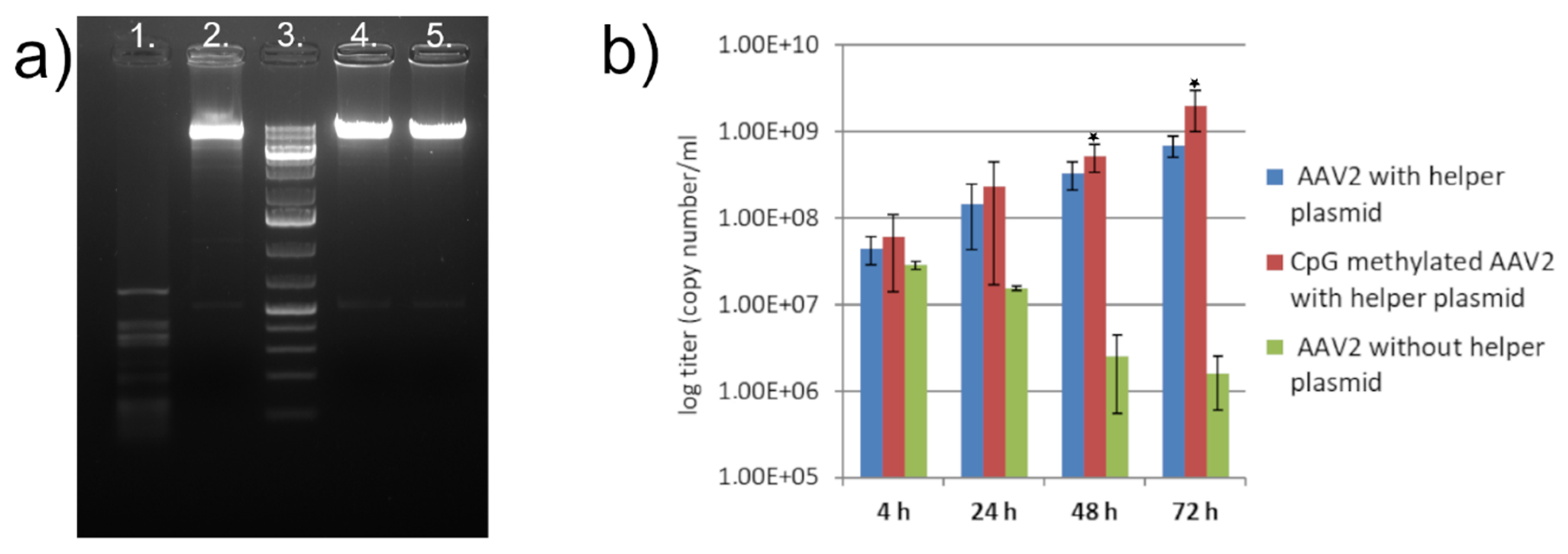{kind=link}
| Primer Name | Sequence | Product Size (bp) | CpGs in Product |
|---|---|---|---|
| AAV1F | 5′-TTGGTTATTTTTTTTTTGCGCGTT-3′ | 205 | 19 |
| AAV1R | 5′-CCTCTAATACAAAACCTCCCTA-3′ | ||
| AAV2F | 5′-GGGTTAGGGAGGTTTTGTATT-3′ | 279 | 17 |
| AAV2R | 5′-ATTCAAATCCATATCAAAATCTAAC-3′ | ||
| AAV3F | 5′-ATTTTGATATGGATTTGAATTTGATT-3′ | 343 | 23 |
| AAV3R | 5′-AAAATATAACACTCATCCACCACCT-3′ | ||
| AAV4F | 5′-AGGGAGAGAGTTATTTTTATATGTA-3′ | 372 | 25 |
| AAV4R | 5′-TCTAATTCTCTTTATTCTACTCCTAC-3′ | ||
| AAV5F | 5′-AAGGTGGTGGATGAGTGTTATATTT-3′ | 309 | 15 |
| AAV5R | 5′-AACCTAATCCTCCTAAATCCACTACTT-3′ | ||
| AAV6F | 5′-GGAGAAGTAGTGGATTTAGGAGGAT-3′ | 298 | 14 |
| AAV6R | 5′-AATTACAAACCCAAACAACCAAATA-3′ | ||
| AAV7F | 5′-GGAAAGATTATGAGTTTGATTAAAAT-3′ | 284 | 15 |
| AAV7R | 5′-AAAAAATTCTCATTAATCCAATTTAC-3′ | ||
| AAV8F | 5′-AATTGGATTAATGAGAATTTTTTTT-3′ | 315 | 21 |
| AAV8R | 5′-AATAACCTTCCCAAAATCATAATCC-3′ | ||
| AAV9F | 5′-TGATTTTGGGAAGGTTATTAAGTAG-3′ | 274 | 17 |
| AAV9R | 5′-ACAAAAAAACAACATCAAATTCATAC-3′ | ||
| AAV10F | 5′-TGATGTTGTTTTTTTGTAGATAATG-3′ | 345 | 10 |
| AAV10R | 5′-TAAACCAAATTTAAACTTCCACCAC-3′ | ||
| AAV11F | 5′-TGGTGGAAGTTTAAATTTGGTTTAT-3′ | 323 | 27 |
| AAV11R | 5′-AAAAATTCAAAAACCCTCTTTTTC-3′ | ||
| AAV12F | 5′-AAAAAGAGGGTTTTTGAATTTTTG-3′ | 152 | 6 |
| AAV12R | 5′-TTCAATCTTTTTCTTACAAACTACTAACC-3′ | ||
| AAV13F | 5′-TTTGGTTGAGGAATTTGTTAAGA-3′ | 369 | 18 |
| AAV13R | 5′-TTATAAATAAACAAAACCCAAATTC-3′ | ||
| AAV14F | 5′-GTTTTTTTTGGTTTGGGAATTAATA-3′ | 282 | 12 |
| AAV14R | 5′-AAATCTATTAAAATCAAAATACCCCC-3′ | ||
| AAV15F | 5′-TTGGGTTTTGTTTATTTATAATAATTATTT-3′ | 217 | 4 |
| AAV15R | 5′-AATATTAAAAAACTTAAAATTAAATCTCTT-3′ | ||
| AAV16F | 5′-AGATTTATTAATAATAATTGGGGATTT-3′ | 299 | 18 |
| AAV16R | 5′-TACTCCAAACAATAAAATAAAAAAC-3′ | ||
| AAV17F | 5′-AGTATGGATATTTTATTTTGAATAA-3′ | 316 | 12 |
| AAV17R | 5′-AAAAACCAATTCCTAAACTAATCCC-3′ | ||
| AAV18F | 5′-AGTTAAGGTTTTAGTTTTTTTAGGT-3′ | 340 | 12 |
| AAV18R | 5′-AAATTAATTATCCTAATTTCCTCTTC-3′ | ||
| AAV19F | 5′-AATGGTAGAGATTTTTTGGTGAATT-3′ | 317 | 9 |
| AAV19R | 5′-AACCCCTAAAAATACACATCTCTATC-3′ | ||
| AAV20F | 5′-AGGTATGGTTTGGTAGGATAGAGAT-3′ | 340 | 12 |
| AAV20R | 5′-ATCCACAATAAAATCCACATTAACAA-3′ | ||
| AAV21F | 5′-AGTGGGAGTTGTAGAAGGAAAATAGTA-3′ | 312 | 10 |
| AAV21R | 5′-TAACCAACTCCATCACTAAAAATTC-3′ | ||
| AAV22F | 5′-GTTTGTTAATGTGGATTTTATTGTGGAT-3′ | 360 | 22 |
| AAV22R | 5′-TAACCACTCCCTCTCTACGCGCT-3′ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tóth, R.; Mészáros, I.; Hüser, D.; Forró, B.; Marton, S.; Olasz, F.; Bányai, K.; Heilbronn, R.; Zádori, Z. Methylation Status of the Adeno-Associated Virus Type 2 (AAV2). Viruses 2019, 11, 38. https://doi.org/10.3390/v11010038
Tóth R, Mészáros I, Hüser D, Forró B, Marton S, Olasz F, Bányai K, Heilbronn R, Zádori Z. Methylation Status of the Adeno-Associated Virus Type 2 (AAV2). Viruses. 2019; 11(1):38. https://doi.org/10.3390/v11010038
Chicago/Turabian StyleTóth, Renáta, István Mészáros, Daniela Hüser, Barbara Forró, Szilvia Marton, Ferenc Olasz, Krisztián Bányai, Regine Heilbronn, and Zoltán Zádori. 2019. "Methylation Status of the Adeno-Associated Virus Type 2 (AAV2)" Viruses 11, no. 1: 38. https://doi.org/10.3390/v11010038
APA StyleTóth, R., Mészáros, I., Hüser, D., Forró, B., Marton, S., Olasz, F., Bányai, K., Heilbronn, R., & Zádori, Z. (2019). Methylation Status of the Adeno-Associated Virus Type 2 (AAV2). Viruses, 11(1), 38. https://doi.org/10.3390/v11010038