Strategies to Encapsulate the Staphylococcus aureus Bacteriophage phiIPLA-RODI

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Phage Propagation and Enumeration
2.3. Encapsulation Processes
2.4. Characterization of Nanovesicles
2.5. Viability and Encapsulation Efficiency
2.6. Stability of Encapsulated Phages During Storage
2.7. Stability of Encapsulated Phages Under Extreme Conditions
2.8. Statistical Analysis
3. Results
3.1. Bacteriophage phiIPLA-RODI Can Be Successfully Entrapped in Different Nanovesicles
3.2. The Efficiency of phiIPLA-RODI Encapsulation Is Not Greatly Influenced by Component Concentration
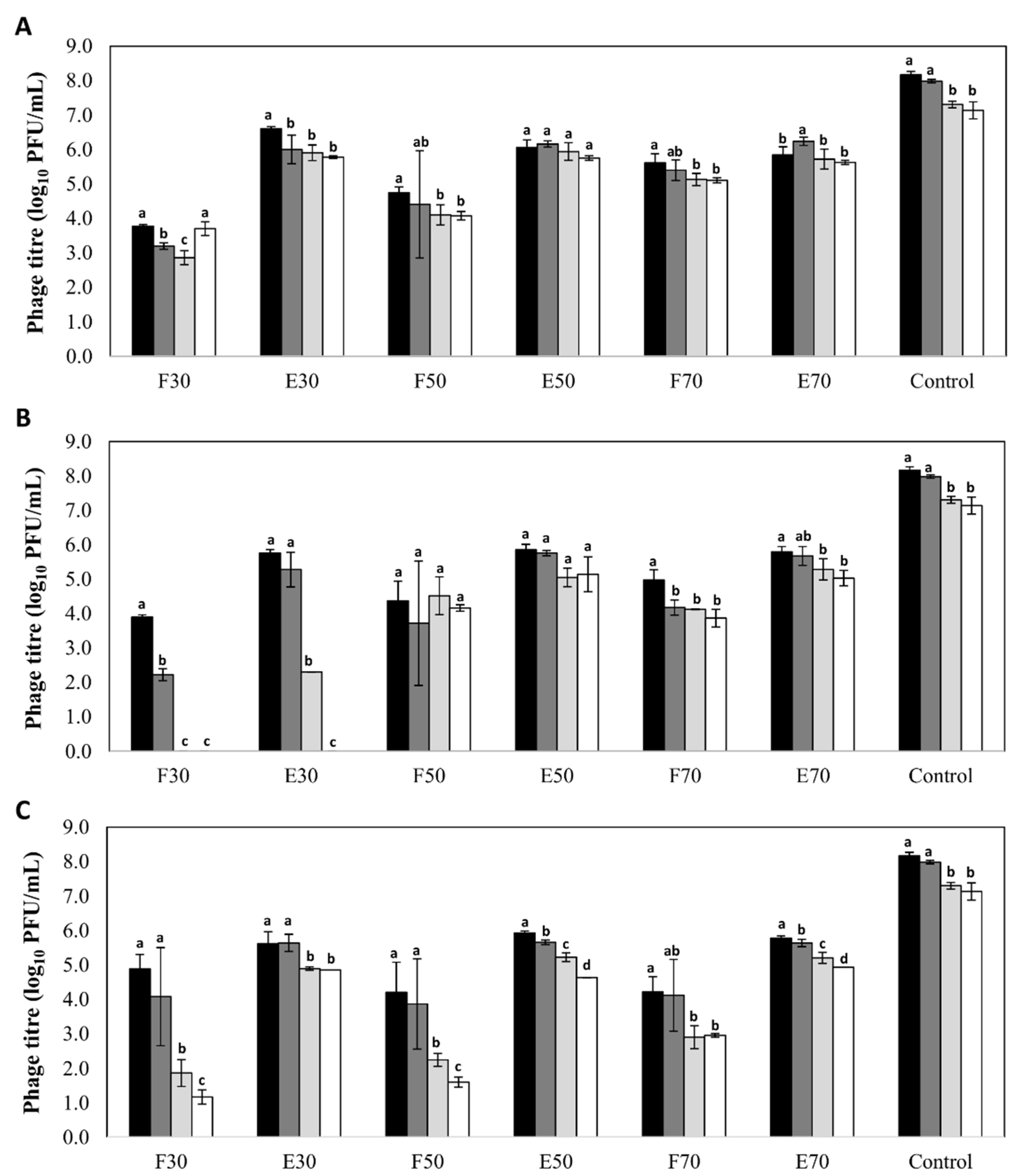
3.3. Stability of Encapsulated Phage Particles during Storage at Low Temperature
3.4. Niosomes Protect Phages from Low pH and High Temperature
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Clokie, M.R.; Millard, A.D.; Letarov, A.V.; Heaphy, S. Phages in nature. Bacteriophage 2011, 1, 31–45. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.; Hawkins, C.H.; Anggård, E.E.; Harper, D.R. A controlled clinical trial of a therapeutic bacteriophage preparation in chronic otitis due to antibiotic-resistant Pseudomonas aeruginosa; a preliminary report of efficacy. Clin. Otolaryngol. 2009, 34, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Saez, A.C.; Zhang, J.; Rostagno, M.H.; Ebner, P.D. Direct feeding of microencapsulated bacteriophages to reduce Salmonella colonization in pigs. Foodborne Pathog. Dis. 2011, 8, 1269–1274. [Google Scholar] [CrossRef] [PubMed]
- Borie, C.; Albala, I.; Sánchez, P.; Sánchez, M.L.; Ramírez, S.; Navarro, C.; Morales, M.A.; Retamales, A.J.; Robeson, J. Bacteriophage treatment reduces Salmonella colonization of infected chickens. Avian Dis. 2008, 52, 64–67. [Google Scholar] [CrossRef] [PubMed]
- Kutter, E.; de Vos, D.; Gvasalia, G.; Alavidze, Z.; Gogokhia, L.; Kuhl, S.; Abedon, S.T. Phage therapy in clinical practice: Treatment of human infections. Curr. Pharm. Biotechnol. 2010, 11, 69–86. [Google Scholar] [CrossRef] [PubMed]
- García, P.; Martínez, B.; Obeso, J.M.; Rodríguez, A. Bacteriophages and their application in food safety. Lett. Appl. Microbiol. 2008, 47, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Bueno, E.; García, P.; Martínez, B.; Rodríguez, A. Phage inactivation of Staphylococcus aureus in fresh and hard-type cheeses. Int. J. Food Microbiol. 2012, 158, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Colom, J.; Cano-Sarabia, M.; Otero, J.; Cortés, P.; Maspoch, D.; Llagostera, M. Liposome-encapsulated bacteriophages for enhanced oral phage therapy against Salmonella spp. Appl. Environ. Microbiol. 2015, 81, 4841–4849. [Google Scholar] [CrossRef] [PubMed]
- Sheng, H.; Knecht, H.J.; Kudva, I.T.; Hovde, C.J. Application of bacteriophages to control intestinal Escherichia coli O157:H7 levels in ruminants. Appl. Environ. Microbiol. 2006, 72, 5359–5366. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, D.; Rodríguez-Rubio, L.; Martínez, B.; Rodríguez, A.; García, P. Bacteriophages as Weapons against Bacterial Biofilms in the Food Industry. Front. Microbiol. 2016, 7, 825. [Google Scholar] [CrossRef] [PubMed]
- Hammerl, J.A.; Jäckel, C.; Alter, T.; Janzcyk, P.; Stingl, K.; Knüver, M.T.; Hertwig, S. Reduction of Campylobacter jejuni in broiler chicken by successive application of group II and group III phages. PLoS ONE 2014, 9, e114785. [Google Scholar] [CrossRef] [PubMed]
- Soni, K.A.; Nannapaneni, R. Removal of Listeria monocytogenes biofilms with bacteriophage P100. J. Food Protect. 2010, 73, 1519–31524. [Google Scholar] [CrossRef]
- Anal, A.K.; Singh, H. Recent advances in microencapsulation of probiotics for industrial applications and targeted delivery. Trends Food Sci. Technol. 2007, 18, 240–251. [Google Scholar] [CrossRef]
- Dias, M.I.; Ferreira, I.C.; Barreiro, M.F. Microencapsulation of bioactives for food applications. Food Funct. 2015, 6, 1035–1052. [Google Scholar] [CrossRef] [PubMed]
- Pando, D.; Beltrán, M.; Gerone, I.; Matos, M.; Pazos, C. Resveratrol entrapped niosomes as yoghurt additive. Food Chem. 2015, 170, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.L.; Paris, H.L.; Beals, J.W.; Binns, S.E.; Giordano, G.R.; Scalzo, R.L.; Bell, C. Liposomal-encapsulated ascorbic acid: Influence on vitamin C bioavailability and capacity to protect against ischemia–reperfusion injury. Nutr. Metab. Insights 2016, 9, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, G.; Matos, M.; Barrero, P.; Pando, D.; Iglesias, O.; Pazos, C. Iron-entrapped niosomes and their potential application for yogurt fortification. LWT Food Sci. Technol. 2016, 74, 550–556. [Google Scholar] [CrossRef]
- Parmentier, J.; Thewes, B.; Gropp, F.; Fricker, G. Oral peptide delivery by tetraether lipid liposomes. Int. J. Pharm. 2011, 415, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Schoina, V.; Terpou, A.; Angelika-Ioanna, G.; Koutinas, A.; Kanellaki, M.; Bosnea, L. Use of Pistacia terebinthus resin as immobilization support for Lactobacillus casei cells and application in selected dairy products. J. Food Sci. Technol. 2015, 52, 5700–5708. [Google Scholar] [CrossRef] [PubMed]
- D’Orazio, G.; Di Gennaro, P.; Boccarusso, M.; Presti, I.; Bizzaro, G.; Giardina, S.; Michelotti, A.; Labra, M.; La Ferla, B. Microencapsulation of new probiotic formulations for gastrointestinal delivery: In vitro study to assess viability and biological properties. Appl. Microbiol. Biotechnol. 2015, 99, 9779–9789. [Google Scholar] [CrossRef] [PubMed]
- Rodklongtan, A.; La-Ongkham, O.; Nitisinprasert, S.; Chitprasert, P. Enhancement of Lactobacillus reuteri KUB-AC5 survival in broiler gastrointestinal tract by microencapsulation with alginate-chitosan semi-interpenetrating polymer networks. J. Appl. Microbiol. 2014, 117, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Malheiros, P.S.; Sant’Anna, V.; Utpott, M.; Brandelli, A. Antilisterial activity and stability of nanovesicle-encapsulated antimicrobial peptide P34 in milk. Food Control 2012, 23, 42–47. [Google Scholar] [CrossRef]
- Were, L.M.; Bruce, B.; Davidson, P.M.; Weiss, J. Encapsulation of nisin and lysozyme in liposomes enhances efficacy against Listeria monocytogenes. J. Food Prot. 2004, 67, 922–927. [Google Scholar] [CrossRef] [PubMed]
- Choińska-Pulit, A.; Mituła, P.; Śliwka, P.; Łaba, W.; Kurzępa-Skaradzińska, A. Bacteriophage encapsulation: Trends and potential applications. Trends Food Sci. Technol. 2015, 45, 212–221. [Google Scholar] [CrossRef]
- Malik, D.J.; Sokolov, I.J.; Vinner, G.K.; Mancuso, F.; Cinquerrui, S.; Vladisavljevic, G.T.; Clokie, M.R.J.; Garton, N.J.; Stapley, A.G.F.; Kirpichnikova, A. Formulation, stabilisation and encapsulation of bacteriophage for phage therapy. Adv. Colloid Interface Sci. 2017, 249, 100–133. [Google Scholar] [CrossRef] [PubMed]
- Lowy, F.D. Staphylococcus aureus infections. N. Engl. J. Med. 1998, 339, 520–532. [Google Scholar] [CrossRef] [PubMed]
- Le Loir, Y.; Baron, F.; Gautier, M. Staphylococcus aureus and food poisoning. Genet. Mol. Res. 2003, 2, 63–76. [Google Scholar] [PubMed]
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2015. EFSA J. 2016, 14, 4634. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). National Outbreak Reporting System (NORS); Centers for Disease Control and Prevention (CDC): Atlanta, GA, USA, 2017.
- Kurosu, M.; Siricilla, S.; Mitachi, K. Advances in MRSA drug discovery: Where are we and where do we need to be? Expert Opin. Drug Discov. 2013, 8, 1095–1116. [Google Scholar] [CrossRef] [PubMed]
- Rhoads, D.D.; Wolcott, R.D.; Kuskowski, M.A.; Wolcott, B.M.; Ward, L.S.; Sulakvelidze, A. Bacteriophage therapy of venous leg ulcers in humans: Results of a phase I safety trial. J. Wound Care 2009, 18, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Fischetti, V.A. Using phage lytic enzymes to control pathogenic bacteria. BMC Oral Health 2006, 6, S16. [Google Scholar] [CrossRef] [PubMed]
- El Haddad, L.; Roy, J.P.; Khalil, G.E.; St-Gelais, D.; Champagne, C.P.; Labrie, S.; Moineau, S. Efficacy of two Staphylococcus aureus phage cocktails in cheese production. Int. J. Food Microbiol. 2016, 217, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.; Xin, Y.; Zhang, C.; Ouyang, X.; Kong, J. The potential of the endolysin Lysdb from Lactobacillus delbrueckii phage for combating Staphylococcus aureus during cheese manufacture from raw milk. Appl. Microbiol. Biotechnol. 2016, 100, 3545–3554. [Google Scholar] [CrossRef] [PubMed]
- García, P.; Madera, C.; Martínez, B.; Rodríguez, A. Biocontrol of Staphylococcus aureus in curd manufacturing processes using bacteriophages. Int. Dairy J. 2007, 17, 1232–1239. [Google Scholar] [CrossRef]
- Obeso, J.M.; García, P.; Martínez, B.; Arroyo-López, F.N.; Garrido-Fernández, A.; Rodríguez, A. Use of logistic regression for prediction of the fate of Staphylococcus aureus in pasteurized milk in the presence of two lytic phages. Appl. Environ. Microbiol. 2010, 76, 6038–6046. [Google Scholar] [CrossRef] [PubMed]
- García, P.; Madera, C.; Martinez, B.; Rodríguez, A.; Suarez, J.E. Prevalence of bacteriophages infecting Staphylococcus aureus in dairy samples and their potential as biocontrol agents. J. Dairy Sci. 2009, 92, 3019–3026. [Google Scholar] [CrossRef] [PubMed]
- Tabla, R.; Martinez, B.; Rebollo, J.E.; Gonzalez, J.; Ramirez, M.R.; Roa, I.; Rodriguez, A.; Garcia, P. Bacteriophage performance against Staphylococcus aureus in milk is improved by high hydrostatic pressure treatments. Int. J. Food Microbiol. 2012, 156, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, D.; Vandenheuvel, D.; Martínez, B.; Rodríguez, A.; Lavigne, R.; García, P. Two Phages, phiIPLA-RODI and phiIPLA-C1C, lyse mono- and dual-species staphylococcal biofilms. Appl. Environ. Microbiol. 2015, 81, 3336–3348. [Google Scholar] [CrossRef] [PubMed]
- Marianecci, C.; Di Marzio, L.; Rinaldi, F.; Celia, C.; Paolino, D.; Alhaique, F.; Esposito, S.; Carafa, M. Niosomes from 80s to present: The state of the art. Adv. Colloid Interface Sci. 2014, 205, 187–206. [Google Scholar] [CrossRef] [PubMed]
- Bangham, A.D.; Standish, M.M.; Watkins, J.C. Diffusion of univalent ions across the lamellae of swollen phospholipids. J. Mol. Biol. 1965, 13, 238–252. [Google Scholar] [CrossRef]
- Coffey, B.; Mills, S.; Coffey, A.; McAuliffe, O.; Ross, R.P. Phage and their lysins as biocontrol agents for food safety applications. Annu. Rev. Food Sci. Technol. 2010, 1, 449–468. [Google Scholar] [CrossRef] [PubMed]
- Singla, S.; Harjai, K.; Raza, K.; Wadhwa, S.; Katare, O.P.; Chhibber, S. Phospholipid vesicles encapsulated bacteriophage: A novel approach to enhance phage biodistribution. J. Virol. Methods 2016, 236, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Nallamothu, R.; Wood, G.C.; Kiani, M.F.; Moore, B.M.; Horton, F.P.; Thoma, L.A. A targeted liposome delivery system for combretastatin A4: Formulation optimization through drug loading and in vitro release studies. PDA J. Pharm. Sci. Technol. 2006, 60, 144–155. [Google Scholar] [PubMed]
- Leung, S.S.; Morales, S.; Britton, W.; Kutter, E.; Chan, H.K. Microfluidic-assisted bacteriophage encapsulation into liposomes. Int. J. Pharm. 2018, 545, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, H.; Wang, Q.; Lim, L.T.; Balamurugan, S. Encapsulation of Listeria Phage A511 by Alginate to Improve Its Thermal Stability. Methods Mol. Biol. 2018, 1681, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Singla, S.; Harjai, K.; Katare, O.P.; Chhibber, S. Encapsulation of bacteriophage in liposome accentuates its entry in to macrophage and shields it from neutralizing antibodies. PLoS ONE 2016, 11, e0153777. [Google Scholar] [CrossRef] [PubMed]
- Drummond, D.C.; Zignani, M.; Leroux, J.C. Current status of pH-sensitive liposomes in drug delivery. Prog. Lipid Res. 2000, 39, 409–460. [Google Scholar] [CrossRef]
- Kumar, G.P.; Rajeshwarrao, P. Nonionic surfactant vesicular systems for effective drug delivery—An overview. Acta Pharm. Sin. B 2001, 1, 208–219. [Google Scholar] [CrossRef]
- Picon, A.; Gaya, P.; Medina, M.; Nunez, M. Proteinases encapsulated in stimulated release liposomes for cheese ripening. Biotechnol. Lett. 1997, 19, 345–348. [Google Scholar] [CrossRef]
- Kheadr, E.E.; Vuillemard, J.C.; El Deeb, S.A. Accelerated Cheddar cheese ripening with encapsulated proteinases. Int. J. Food Sci. Technol. 2000, 35, 483–495. [Google Scholar] [CrossRef]
- Laloy, E.; Vuillemard, J.C.; Dufour, P.; Simard, R. Release of enzymes from liposomes during cheese ripening. J. Control. Release 1998, 54, 213–222. [Google Scholar] [CrossRef]
- Shukla, S.; Haldorai, Y.; Hwang, S.K.; Bajpai, V.K.; Huh, Y.S.; Han, Y.K. Current Demands for Food-Approved Liposome Nanoparticles in Food and Safety Sector. Front. Microbiol. 2017, 8, 2398. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Vesicles | Composition | Homogenization (rpm)/Duration (min) | Z-Average (µm) | ζ-Potential (mV) | Viability Loss (Log Units) | |
|---|---|---|---|---|---|---|
| Components | Concentration * | |||||
| Niosome | Pronanosome Nio-N™ | 30 mg/mL, | 8000/5 | 0.83 ± 0.11 | −34.3 ± 1.0 | 0.5 ± 0.1 |
| 50 mg/mL, or | 0.85 ± 0.12 | −33.4 ± 0.2 | 1.0 ± 0.2 | |||
| 70 mg/mL | 0.80 ± 0.07 | −35.6 ± 1.3 | 1.1 ± 0.2 | |||
| Liposome | Pronanosome Lipo-N™ | 30 mg/mL, | 5000/5 | 1.51 ± 0.17 | −14.1 ± 1.0 | 1.3 ± 0.1 |
| 50 mg/mL, or | 1.60 ± 0.17 | −14.1 ± 0.5 | 1.2 ± 0.2 | |||
| 70 mg/mL | 1.89 ± 0.03 | −13.5 ± 0.1 | 1.2 ± 0.2 | |||
| Transfersome | Phospholipon 90G and Span 60 (1:1) | 30 mg/mL, | 8000/5 | 0.51 ± 0.07 | −30.3 ± 1.2 | 1.4 ± 0.3 |
| 50 mg/mL, or | 0.55 ± 0.03 | −30.8 ± 2.0 | 1.2 ± 0.0 | |||
| 70 mg/mL | 0.58 ± 0.06 | −28.6 ± 0.2 | 1.3 ± 0.1 | |||
| Concentration of Components (mg/mL) | Encapsulation Efficiency (% PFU/mL) | ||
|---|---|---|---|
| Niosomes | Liposomes | Transfersomes | |
| 30 | 99.8.0 ± 0.03 | 98.6 ± 0.47 | 76.9 ± 21.49 |
| 50 | 94.5 ± 3.29 | 95.2 ± 4.30 | 95.6 ± 4.96 |
| 70 | 62.3 ± 14.35 *,# | 85.5 ± 9.04 | 96.6 ± 2.89 |
| Nanovesicles | Component (mg/mL) | Phage phiIPLA-RODI | Initial Titer log10 (PFU/mL) | pH 4.5 60 min | Tª 60 °C 90 min | NaCl 4.5 M 60 min |
|---|---|---|---|---|---|---|
| Niosomes | 30 | F | 3.73 ± 0.20 | − | 4.74 ± 0.08 * | |
| E | 5.78 ± 0.03 | 3.78 ± 0.1 * | 3.54 ± 0.50 * | 4.52 ± 0.13 * | ||
| T | 5.78 ± 0.03 | 3.78 ± 0.1 * | 3.54 ± 0.50 * | 4.95 ± 0.02 * | ||
| 50 | F | 4.09 ± 0.12 | − | N/A | 5.32 ± 0.18 * | |
| E | 5.76 ± 0.07 | 4.00 ± 0.19 * | N/A | 4.50 ± 0.10 * | ||
| T | 5.77 ± 0.06 | 4.00 ± 0.19 * | N/A | 5.38 ± 0.15 * | ||
| 70 | F | 5.11 ± 0.07 | − | N/A | 5.79 ± 0.17 * | |
| E | 5.63 ± 0.06 | 3.85 ± 0.16 * | N/A | 4.44 ± 0.11 * | ||
| T | 5.74 ± 0.04 | 3.85 ± 0.16 * | N/A | 5.81 ± 0.16 | ||
| Liposomes | 30 | F | 3.53 ± 0.15 | − | − | 5.03 ± 0.32 * |
| E | 5.80 ± 0.85 | − | − | 4.85 ± 0.22 | ||
| T | 5.80 ± 0.84 | − | − | 5.25 ± 0.25 | ||
| 50 | F | 4.16 ± 0.09 | − | N/A | 5.05 ± 0.12 * | |
| E | 5.14 ± 0.51 | 3.46 ± 0.28 * | − | 4.03 ± 0.37 * | ||
| T | 5.18 ± 0.22 | 3.46 ± 0.28 * | N/A | 5.19 ± 0.14 | ||
| 70 | F | 3.87 ± 0.26 | − | N/A | 4.71 ± 0.68 | |
| E | 5.03 ± 0.22 | 2.30 ± 0.24 * | N/A | 4.48 ± 0.54 | ||
| T | 5.06 ± 0.18 | 2.30 ± 0.24 * | N/A | 4.91 ± 0.62 | ||
| Transfersomes | 30 | F | − | − | − | 4.93 ± 0.53 * |
| E | 4.88 ± 0.21 | − | − | 4.43 ± 0.34 | ||
| T | 4.88 ± 0.21 | − | − | 5.01 ± 0.48 | ||
| 50 | F | − | − | N/A | 5.01 ± 0.43 * | |
| E | 4.63 ± 0.15 | − | N/A | 4.18 ± 0.22 * | ||
| T | 4.63 ± 0.15 | − | N/A | 4.75 ± 0.52 | ||
| 70 | F | 2.95 ± 0.02 | − | N/A | 5.05 ± 0.19 * | |
| E | 4.93 ± 0.05 | − | N/A | 4.60 ± 0.05 * | ||
| T | 4.94 ± 0.10 | − | N/A | 5.18 ± 0.17 | ||
| Control phage in SM buffer | N/A | N/A | 8.52 ± 0.10 | − | 4.75 ± 0.22 * | 8.08 ± 0.48 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Menéndez, E.; Fernández, L.; Gutiérrez, D.; Pando, D.; Martínez, B.; Rodríguez, A.; García, P. Strategies to Encapsulate the Staphylococcus aureus Bacteriophage phiIPLA-RODI. Viruses 2018, 10, 495. https://doi.org/10.3390/v10090495
González-Menéndez E, Fernández L, Gutiérrez D, Pando D, Martínez B, Rodríguez A, García P. Strategies to Encapsulate the Staphylococcus aureus Bacteriophage phiIPLA-RODI. Viruses. 2018; 10(9):495. https://doi.org/10.3390/v10090495
Chicago/Turabian StyleGonzález-Menéndez, Eva, Lucía Fernández, Diana Gutiérrez, Daniel Pando, Beatriz Martínez, Ana Rodríguez, and Pilar García. 2018. "Strategies to Encapsulate the Staphylococcus aureus Bacteriophage phiIPLA-RODI" Viruses 10, no. 9: 495. https://doi.org/10.3390/v10090495
APA StyleGonzález-Menéndez, E., Fernández, L., Gutiérrez, D., Pando, D., Martínez, B., Rodríguez, A., & García, P. (2018). Strategies to Encapsulate the Staphylococcus aureus Bacteriophage phiIPLA-RODI. Viruses, 10(9), 495. https://doi.org/10.3390/v10090495