Improved Immune Responses Against Zika Virus After Sequential Dengue and Zika Virus Infection in Humans

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Human Blood Samples
2.3. Viruses and Cell Lines
2.4. HLA Typing
2.5. Serology
2.6. RT-PCR Assays for Detection of DENV and ZIKV
2.7. Viral Sequences
2.8. Peptides
2.9. Ex Vivo IFN-γ ELISPOT Assay
2.10. Immunogenicity and HLA Restrictions Prediction
2.11. Statistics
3. Results
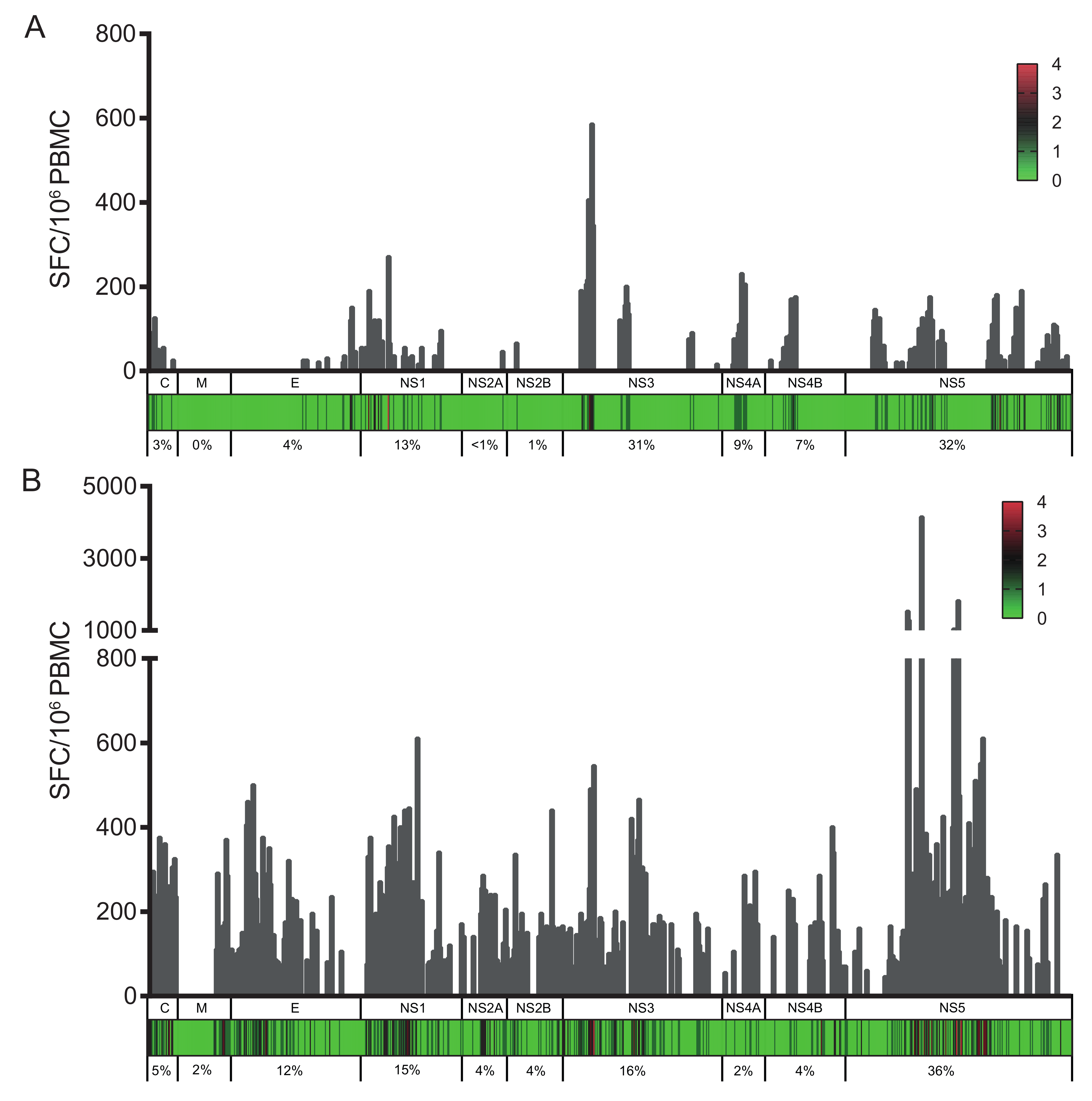
3.1. Identification of Immunodominant Regions of the ZIKV Proteome
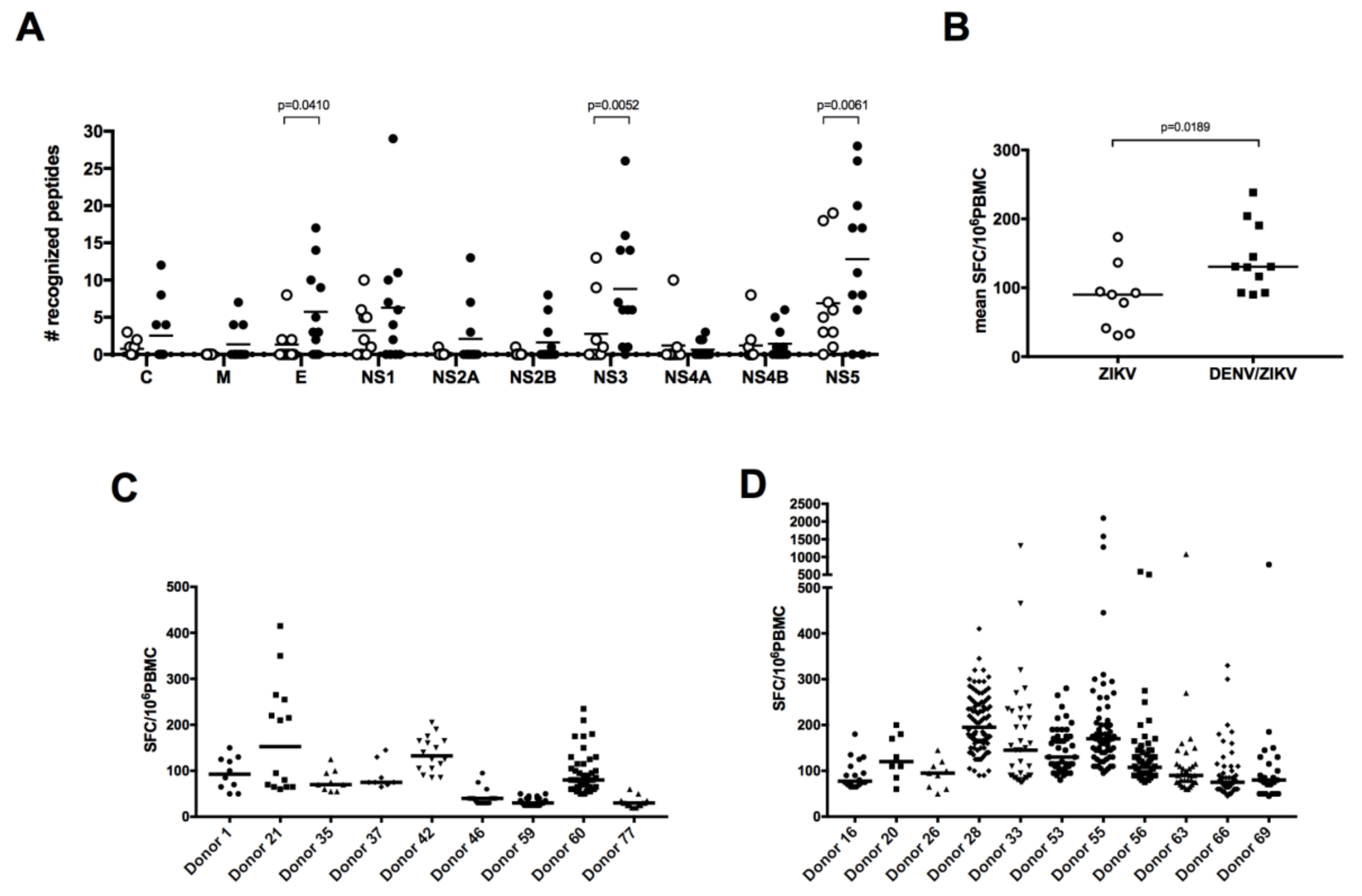
3.2. Broader Responses with a Higher Magnitude in DENV/ZIKV-Immune Donors
3.3. DENV/ZIKV-Cross-Reactive T Cells Mainly Target the NS5 Protein
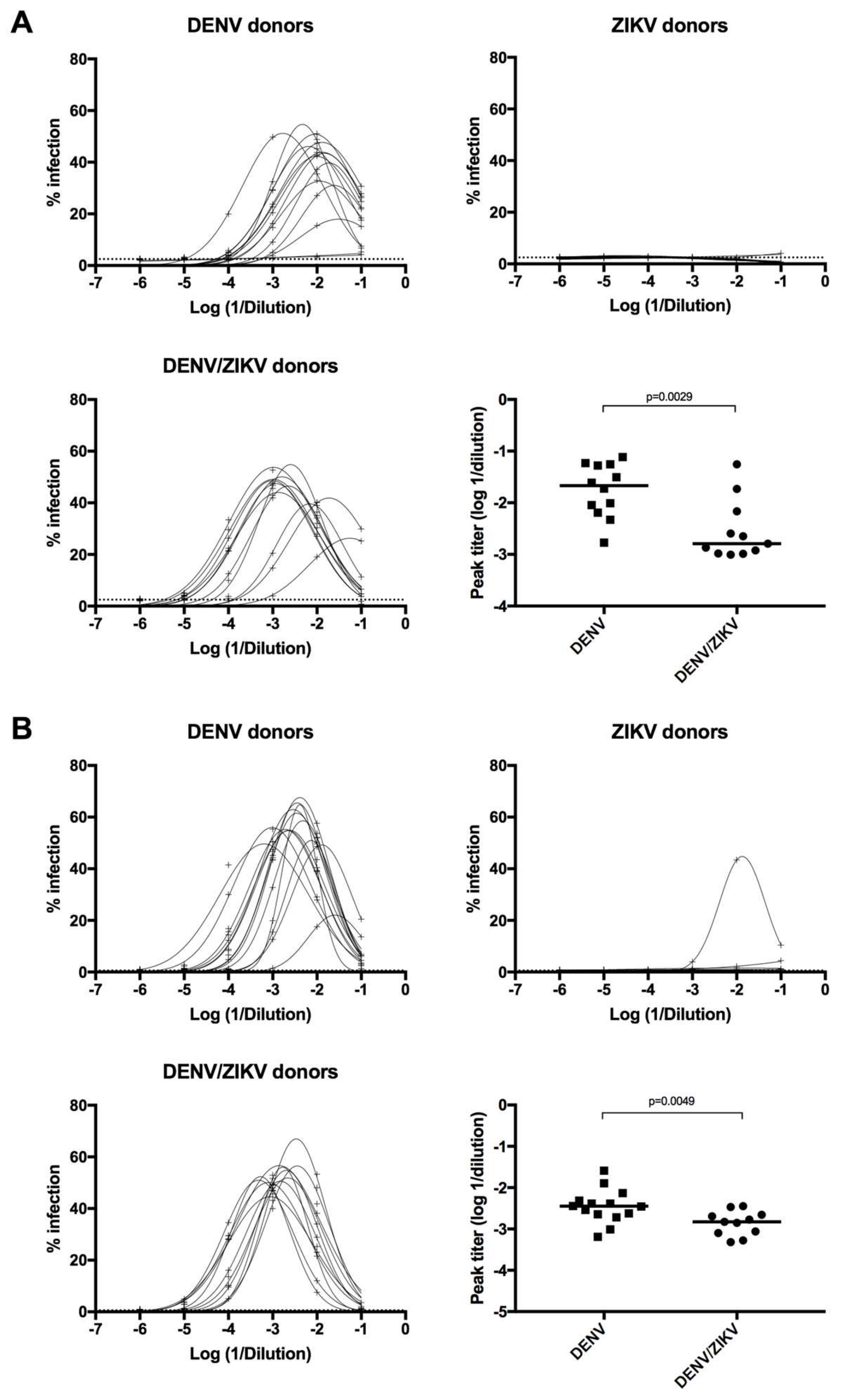
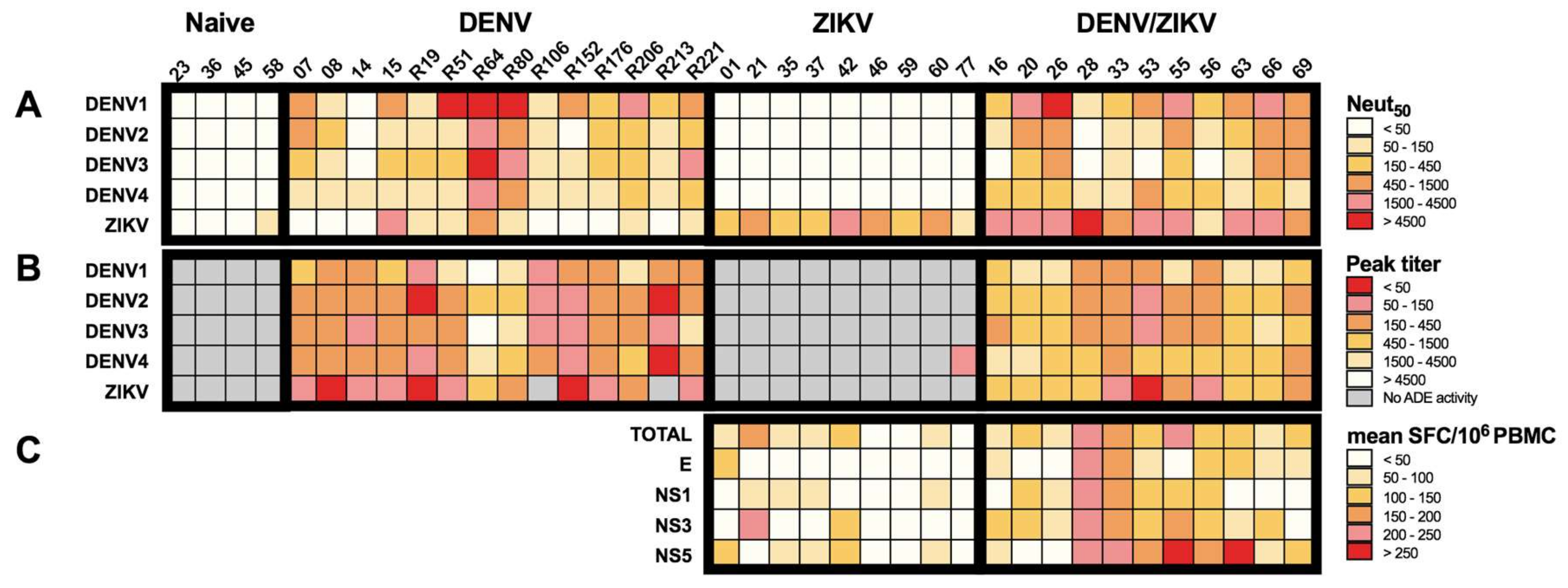
3.4. Increased Neutralizing Antibody Titer against ZIKV in DENV/ZIKV-Immune Donors
3.5. Decreased Enhancing Potential on ZIKV Infection in DENV/ZIKV-Immune Donors
3.6. High Titer of Neutralizing Antibodies with Low Enhancing Activity are Associated with Strong T-Cell Responses
4. Discussion
5. Patents
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kuno, G.; Chang, G.J.; Tsuchiya, K.R.; Karabatsos, N.; Cropp, C.B. Phylogeny of the genus Flavivirus. J. Virol. 1998, 72, 73–83. [Google Scholar] [PubMed]
- Dick, G.W.; Kitchen, S.F.; Haddow, A.J. Zika virus. I. Isolations and serological specificity. Trans. R Soc. Trop. Med. Hyg. 1952, 46, 509–520. [Google Scholar] [CrossRef]
- Duffy, M.R.; Chen, T.H.; Hancock, W.T.; Powers, A.M.; Kool, J.L.; Lanciotti, R.S.; Pretrick, M.; Marfel, M.; Holzbauer, S.; Dubray, C.; et al. Zika virus outbreak on Yap Island, Federated States of Micronesia. N. Engl. J. Med. 2009, 360, 2536–3543. [Google Scholar] [CrossRef] [PubMed]
- Cao-Lormeau, V.M.; Roche, C.; Teissier, A.; Robin, E.; Berry, A.L.; Mallet, H.P.; Sall, A.A.; Musso, D. Zika virus, French polynesia, South pacific, 2013. Emerg. Infect. Dis. 2014, 20, 1085–1086. [Google Scholar] [CrossRef] [PubMed]
- Campos, G.S.; Bandeira, A.C.; Sardi, S.I. Zika Virus Outbreak, Bahia, Brazil. Emerg. Infect. Dis. 2015, 21, 1885–1886. [Google Scholar] [CrossRef] [PubMed]
- Dupont-Rouzeyrol, M.; O’Connor, O.; Calvez, E.; Daures, M.; John, M.; Grangeon, J.P.; Gourinat, A.C. Co-infection with Zika and dengue viruses in 2 patients, New Caledonia, 2014. Emerg. Infect. Dis. 2015, 21, 381–382. [Google Scholar] [CrossRef] [PubMed]
- Zanluca, C.; Melo, V.C.; Mosimann, A.L.; Santos, G.I.; Santos, C.N.; Luz, K. First report of autochthonous transmission of Zika virus in Brazil. Mem. Inst. Oswaldo Cruz. 2015, 110, 569–572. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, O.; Beltran, M.; Nelson, C.A.; Valencia, D.; Tolosa, N.; Farr, S.L.; Padilla, A.V.; Tong, V.T.; Cuevas, E.L.; Espinosa-Bode, A.; et al. Zika virus disease in Colombia-preliminary report. N. Engl. J. Med. 2016. [Google Scholar] [CrossRef] [PubMed]
- Metsky, H.C.; Matranga, C.B.; Wohl, S.; Schaffner, S.F.; Freije, C.A.; Winnicki, S.M.; West, K.; Qu, J.; Baniecki, M.L.; Gladden-Young, A.; et al. Zika virus evolution and spread in the Americas. Nature 2017, 546, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Oehler, E.; Watrin, L.; Larre, P.; Leparc-Goffart, I.; Lastere, S.; Valour, F.; Baudouin, L.; Mallet, H.; Musso, D.; Ghawche, F. Zika virus infection complicated by Guillain-Barre syndrome--case report, French Polynesia, December 2013. Eurosurveillance 2014, 19, 20720. [Google Scholar] [CrossRef] [PubMed]
- Cao-Lormeau, V.M.; Blake, A.; Mons, S.; Lastere, S.; Roche, C.; Vanhomwegen, J.; Dub, T.; Baudouin, L.; Teissier, A.; Larre, P.; et al. Guillain-Barre Syndrome outbreak associated with Zika virus infection in French Polynesia: A case-control study. Lancet. 2016, 387, 1531–1539. [Google Scholar] [CrossRef]
- Cauchemez, S.; Besnard, M.; Bompard, P.; Dub, T.; Guillemette-Artur, P.; Eyrolle-Guignot, D.; Salje, H.; Van Kerkhove, M.D.; Abadie, V.; Garel, C.; et al. Association between Zika virus and microcephaly in French Polynesia, 2013-15: A retrospective study. Lancet. 2016, 387, 2125–2132. [Google Scholar] [CrossRef]
- Soares de Araujo, J.S.; Regis, C.T.; Gomes, R.G.; Tavares, T.R.; Rocha Dos Santos, C.; Assuncao, P.M.; Nobrega, R.V.; Pinto, D.F.; Bezerra, B.V.; Mattos, S.D. Microcephaly in north-east Brazil: A retrospective study on neonates born between 2012 and 2015. Bull. World Health Organ. 2016, 94, 835–840. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, J.Y.; Du, S.Y.; Shan, C.; Nie, K.X.; Zhang, R.; Li, X.F.; Zhang, R.; Wang, T.; Qin, C.F.; et al. Evolutionary enhancement of Zika virus infectivity in Aedes aegypti mosquitoes. Nature 2017, 545, 482–486. [Google Scholar] [CrossRef] [PubMed]
- Xia, H.; Luo, H.; Shan, C.; Muruato, A.E.; Nunes, B.T.D.; Medeiros, D.B.A.; Zou, J.; Xie, X.; Giraldo, M.I.; Vasconcelos, P.F.C.; et al. An evolutionary NS1 mutation enhances Zika virus evasion of host interferon induction. Nat. Commun. 2018, 9, 414. [Google Scholar] [CrossRef] [PubMed]
- Katzelnick, L.C.; Coloma, J.; Harris, E. Dengue: Knowledge gaps, unmet needs, and research priorities. Lancet Infect. Dis. 2017, 17, e88–e100. [Google Scholar] [CrossRef]
- Dejnirattisai, W.; Supasa, P.; Wongwiwat, W.; Rouvinski, A.; Barba-Spaeth, G.; Duangchinda, T.; Sakuntabhai, A.; Cao-Lormeau, V.M.; Malasit, P.; Rey, F.A.; et al. Dengue virus sero-cross-reactivity drives antibody-dependent enhancement of infection with zika virus. Nat. Immunol. 2016, 17, 1102–1108. [Google Scholar] [CrossRef] [PubMed]
- Stettler, K.; Beltramello, M.; Espinosa, D.A.; Graham, V.; Cassotta, A.; Bianchi, S.; Vanzetta, F.; Minola, A.; Jaconi, S.; Mele, F.; et al. Specificity, cross-reactivity, and function of antibodies elicited by Zika virus infection. Science 2016, 353, 823–826. [Google Scholar] [CrossRef] [PubMed]
- Paul, L.M.; Carlin, E.R.; Jenkins, M.M.; Tan, A.L.; Barcellona, C.M.; Nicholson, C.O.; Michael, S.F.; Isern, S. Dengue virus antibodies enhance Zika virus infection. Clin. Transl. Immunol. 2016, 5, e117. [Google Scholar] [CrossRef] [PubMed]
- Priyamvada, L.; Quicke, K.M.; Hudson, W.H.; Onlamoon, N.; Sewatanon, J.; Edupuganti, S.; Pattanapanyasat, K.; Chokephaibulkit, K.; Mulligan, M.J.; Wilson, P.C.; et al. Human antibody responses after dengue virus infection are highly cross-reactive to Zika virus. Proc. Natl. Acad. Sci. USA 2016, 113, 7852–7857. [Google Scholar] [CrossRef] [PubMed]
- Bardina, S.V.; Bunduc, P.; Tripathi, S.; Duehr, J.; Frere, J.J.; Brown, J.A.; Nachbagauer, R.; Foster, G.A.; Krysztof, D.; Tortorella, D.; et al. Enhancement of Zika virus pathogenesis by preexisting antiflavivirus immunity. Science 2017, 356, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Grifoni, A.; Pham, J.; Sidney, J.; O’Rourke, P.H.; Paul, S.; Peters, B.; Martini, S.R.; de Silva, A.D.; Ricciardi, M.J.; Magnani, D.M.; et al. Prior Dengue virus exposure shapes T cell immunity to Zika virus in humans. J. Virol. 2017, 91, e01469-17. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.; Tang, W.W.; Sheets, N.; Ellison, J.; Sette, A.; Kim, K.; Shresta, S. Identification of Zika virus epitopes reveals immunodominant and protective roles for dengue virus cross-reactive CD8+ T cells. Nat. Microbiol. 2017, 2, 17036. [Google Scholar] [CrossRef] [PubMed]
- Herrera, B.B.; Tsai, W.Y.; Chang, C.A.; Hamel, D.J.; Wang, W.K.; Lu, Y.; Mboup, S.; Kanki, P.J. Sustained specific and cross-reactive T cell responses to Zika and Dengue virus NS3 in West Africa. J. Virol. 2018, 92, e01992-17. [Google Scholar] [CrossRef] [PubMed]
- Weiskopf, D.; Angelo, M.A.; de Azeredo, E.L.; Sidney, J.; Greenbaum, J.A.; Fernando, A.N.; Broadwater, A.; Kolla, R.V.; De Silva, A.D.; de Silva, A.M.; et al. Comprehensive analysis of dengue virus-specific responses supports an HLA-linked protective role for CD8+ T cells. Proc. Natl. Acad. Sci. USA 2013, 110, e2046–e2053. [Google Scholar] [CrossRef] [PubMed]
- Weiskopf, D.; Bangs, D.J.; Sidney, J.; Kolla, R.V.; De Silva, A.D.; de Silva, A.M.; Crotty, S.; Peters, B.; Sette, A. Dengue virus infection elicits highly polarized CX3CR1+ cytotoxic CD4+ T cells associated with protective immunity. Proc. Natl. Acad. Sci. USA 2015, 112, e4256–4263. [Google Scholar] [CrossRef] [PubMed]
- Rivino, L. T cell immunity to dengue virus and implications for vaccine design. Expert Rev. Vaccines 2016, 15, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Alam, A.; Ali, S.; Ahamad, S.; Malik, M.Z.; Ishrat, R. From ZikV genome to vaccine: In silico approach for the epitope-based peptide vaccine against Zika virus envelope glycoprotein. Immunology 2016, 149, 386–399. [Google Scholar] [CrossRef] [PubMed]
- Dar, H.; Zaheer, T.; Rehman, M.T.; Ali, A.; Javed, A.; Khan, G.A.; Babar, M.M.; Waheed, Y. Prediction of promiscuous T-cell epitopes in the Zika virus polyprotein: An in silico approach. Asian Pac. J. Trop. Med. 2016, 9, 844–850. [Google Scholar] [CrossRef] [PubMed]
- Usman Mirza, M.; Rafique, S.; Ali, A.; Munir, M.; Ikram, N.; Manan, A.; Salo-Ahen, O.M.; Idrees, M. Towards peptide vaccines against Zika virus: Immunoinformatics combined with molecular dynamics simulations to predict antigenic epitopes of Zika viral proteins. Sci. Rep. 2016, 6, 37313. [Google Scholar] [CrossRef] [PubMed]
- Elong Ngono, A.; Vizcarra, E.A.; Tang, W.W.; Sheets, N.; Joo, Y.; Kim, K.; Gorman, M.J.; Diamond, M.S.; Shresta, S. Mapping and role of the CD8+ T cell response during primary Zika virus infection in mice. Cell Host Microbe 2017, 21, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Li, S.; Zhang, Y.; Han, X.; Jia, B.; Liu, H.; Liu, D.; Tan, S.; Wang, Q.; Bi, Y.; et al. CD8+ T cell immune response in immunocompetent mice during Zika virus infection. J. Virol. 2017, 91, e00900-17. [Google Scholar] [CrossRef] [PubMed]
- Pardy, R.D.; Rajah, M.M.; Condotta, S.A.; Taylor, N.G.; Sagan, S.M.; Richer, M.J. Analysis of the T cell response to Zika Virus and identification of a novel CD8+ T cell epitope in immunocompetent mice. PLoS Pathog. 2017, 13, e1006184. [Google Scholar] [CrossRef] [PubMed]
- Winkler, C.W.; Myers, L.M.; Woods, T.A.; Messer, R.J.; Carmody, A.B.; McNally, K.L.; Scott, D.P.; Hasenkrug, K.J.; Best, S.M.; Peterson, K.E. Adaptive immune responses to Zika virus are important for controlling virus infection and preventing infection in brain and testes. J. Immunol. 2017, 198, 3526–3535. [Google Scholar] [CrossRef] [PubMed]
- Aubry, M.; Teissier, A.; Huart, M.; Merceron, S.; Vanhomwegen, J.; Roche, C.; Vial, A.L.; Teururai, S.; Sicard, S.; Paulous, S.; et al. Zika virus seroprevalence, French Polynesia, 2014–2015. Emerg. Infect. Dis. 2017, 23, 669–672. [Google Scholar] [CrossRef] [PubMed]
- De Alwis, R.; Williams, K.L.; Schmid, M.A.; Lai, C.Y.; Patel, B.; Smith, S.A.; Crowe, J.E.; Wang, W.K.; Harris, E.; de Silva, A.M. Dengue viruses are enhanced by distinct populations of serotype cross-reactive antibodies in human immune sera. PLoS Pathog. 2014, 10, e1004386. [Google Scholar] [CrossRef] [PubMed]
- De Alwis, R.; de Silva, A.M. Measuring antibody neutralization of dengue virus (DENV) using a flow cytometry-based technique. Methods Mol. Biol. 2014, 1138, 27–39. [Google Scholar] [PubMed]
- Calvo, E.P.; Sanchez-Quete, F.; Duran, S.; Sandoval, I.; Castellanos, J.E. Easy and inexpensive molecular detection of dengue, chikungunya and zika viruses in febrile patients. Acta Trop. 2016, 163, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Andreatta, M.; Karosiene, E.; Rasmussen, M.; Stryhn, A.; Buus, S.; Nielsen, M. Accurate pan–specific prediction of peptide-MHC class II binding affinity with improved binding core identification. Immunogenetics 2015, 67, 641–650. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, M.; Andreatta, M. NetMHCpan-3.0; improved prediction of binding to MHC class I molecules integrating information from multiple receptor and peptide length datasets. Genome Med. 2016, 8, 33. [Google Scholar] [CrossRef] [PubMed]
- Simmons, C.P.; Dong, T.; Chau, N.V.; Dung, N.T.; Chau, T.N.; Thao le, T.T.; Dung, N.T.; Hien, T.T.; Rowland-Jones, S.; Farrar, J. Early T-cell responses to dengue virus epitopes in Vietnamese adults with secondary dengue virus infections. J. Virol. 2005, 79, 5665–5675. [Google Scholar] [CrossRef] [PubMed]
- Duangchinda, T.; Dejnirattisai, W.; Vasanawathana, S.; Limpitikul, W.; Tangthawornchaikul, N.; Malasit, P.; Mongkolsapaya, J.; Screaton, G. Immunodominant T-cell responses to dengue virus NS3 are associated with DHF. Proc. Natl. Acad. Sci. USA 2010, 107, 16922–16927. [Google Scholar] [CrossRef] [PubMed]
- Rivino, L.; Kumaran, E.A.; Jovanovic, V.; Nadua, K.; Teo, E.W.; Pang, S.W.; Teo, G.H.; Gan, V.C.; Lye, D.C.; Leo, Y.S.; et al. Differential targeting of viral components by CD4+ versus CD8+ T lymphocytes in dengue virus infection. J. Virol. 2013, 87, 2693–2706. [Google Scholar] [CrossRef] [PubMed]
- Weiskopf, D.; Angelo, M.A.; Bangs, D.J.; Sidney, J.; Paul, S.; Peters, B.; de Silva, A.D.; Lindow, J.C.; Diehl, S.A.; Whitehead, S.; et al. The Human CD8+ T Cell Responses Induced by a Live Attenuated Tetravalent Dengue Vaccine Are Directed against Highly Conserved Epitopes. J. Virol. 2015, 89, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Dikhit, M.R.; Ansari, M.Y.; Vijaymahantesh; Kalyani; Mansuri, R.; Sahoo, B.R.; Dehury, B.; Amit, A.; Topno, R.K.; Sahoo, G.C.; et al. Computational prediction and analysis of potential antigenic CTL epitopes in Zika virus: A first step towards vaccine development. Infect. Genet. Evol. 2016, 45, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Imrie, A.; Meeks, J.; Gurary, A.; Sukhbataar, M.; Kitsutani, P.; Effler, P.; Zhao, Z. Differential functional avidity of dengue virus-specific T-cell clones for variant peptides representing heterologous and previously encountered serotypes. J. Virol. 2007, 81, 10081–10091. [Google Scholar] [CrossRef] [PubMed]
- Tai, W.; He, L.; Wang, Y.; Sun, S.; Zhao, G.; Luo, C.; Li, P.; Zhao, H.; Fremont, D.H.; Li, F.; et al. Critical neutralizing fragment of Zika virus EDIII elicits cross-neutralization and protection against divergent Zika viruses. Emerg. Microbes Infect. 2018, 7, 7. [Google Scholar] [CrossRef] [PubMed]
- Collins, M.H.; McGowan, E.; Jadi, R.; Young, E.; Lopez, C.A.; Baric, R.S.; Lazear, H.M.; de Silva, A.M. Lack of durable cross-neutralizing antibodies against Zika virus from Dengue virus infection. Emerg. Infect. Dis. 2017, 23, 773–781. [Google Scholar] [CrossRef] [PubMed]
- Katzelnick, L.C.; Gresh, L.; Halloran, M.E.; Mercado, J.C.; Kuan, G.; Gordon, A.; Balmaseda, A.; Harris, E. Antibody-dependent enhancement of severe dengue disease in humans. Science. 2017, 358, 929–932. [Google Scholar] [CrossRef] [PubMed]
- Priyamvada, L.; Hudson, W.; Ahmed, R.; Wrammert, J. Humoral cross-reactivity between Zika and dengue viruses: Implications for protection and pathology. Emerg. Microbes Infect. 2017, 6, e33. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.C.; Martin, E.; Shimada, M.; Godfrey, S.B.; Fricke, J.; Locastro, S.; Lai, N.Y.; Liebesny, P.; Carlson, J.M.; Brumme, C.J.; et al. Aminopeptidase substrate preference affects HIV epitope presentation and predicts immune escape patterns in HIV-infected individuals. J. Immunol. 2012, 188, 5924–5934. [Google Scholar] [CrossRef] [PubMed]
- Ricciardi, M.J.; Magnani, D.M.; Grifoni, A.; Kwon, Y.C.; Gutman, M.J.; Grubaugh, N.D.; Gangavarapu, K.; Sharkey, M.; Silveira, C.G.T.; et al. Ontogeny of the B- and T-cell response in a primary Zika virus infection of a dengue-naive individual during the 2016 outbreak in Miami, FL. PLoS Negl. Trop. Dis. 2017, 11, e0006000. [Google Scholar] [CrossRef] [PubMed]
- Swanstrom, J.A.; Plante, J.A.; Plante, K.S.; Young, E.F.; McGowan, E.; Gallichotte, E.N.; Widman, D.G.; Heise, M.T.; de Silva, A.M.; Baric, R.S. Dengue virus envelope dimer epitope monoclonal antibodies isolated from dengue patients are protective against Zika virus. MBio 2016, 7, e01123-16. [Google Scholar] [CrossRef] [PubMed]
- Morens, D.M.; Halstead, S.B. Measurement of antibody-dependent infection enhancement of four dengue virus serotypes by monoclonal and polyclonal antibodies. J. Gen. Virol. 1990, 71, 2909–29014. [Google Scholar] [CrossRef] [PubMed]
- Guy, B.; Chanthavanich, P.; Gimenez, S.; Sirivichayakul, C.; Sabchareon, A.; Begue, S.; Yoksan, S.; Luxemburger, C.; Lang, J. Evaluation by flow cytometry of antibody-dependent enhancement (ADE) of dengue infection by sera from Thai children immunized with a live-attenuated tetravalent dengue vaccine. Vaccine 2004, 22, 3563–3574. [Google Scholar] [CrossRef] [PubMed]
- Barba-Spaeth, G.; Dejnirattisai, W.; Rouvinski, A.; Vaney, M.C.; Medits, I.; Sharma, A.; Simon-Loriere, E.; Sakuntabhai, A.; Cao-Lormeau, V.M.; Haouz, A.; et al. Structural basis of potent Zika-dengue virus antibody cross-neutralization. Nature 2016, 536, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Robbiani, D.F.; Bozzacco, L.; Keeffe, J.R.; Khouri, R.; Olsen, P.C.; Gazumyan, A.; Schaefer-Babajew, D.; Avila-Rios, S.; Nogueira, L.; Patel, R.; et al. Recurrent potent human neutralizing antibodies to Zika virus in Brazil and Mexico. Cell 2017, 169, 597–609. [Google Scholar] [CrossRef] [PubMed]
- Rogers, T.F.; Goodwin, E.C.; Briney, B.; Sok, D.; Beutler, N.; Strubel, A.; Nedellec, R.; Le, K.; Brown, M.E.; Burton, D.R.; et al. Zika virus activates de novo and cross-reactive memory B cell responses in dengue-experienced donors. Sci. Immunol. 2017, 2, 6809. [Google Scholar] [CrossRef] [PubMed]
- McCracken, M.K.; Gromowski, G.D.; Friberg, H.L.; Lin, X.; Abbink, P.; De La Barrera, R.; Eckles, K.H.; Garver, L.S.; Boyd, M.; Jetton, D.; et al. Impact of prior flavivirus immunity on Zika virus infection in rhesus macaques. PLoS Pathog. 2017, 13, e1006487. [Google Scholar] [CrossRef] [PubMed]
- Pantoja, P.; Perez-Guzman, E.X.; Rodriguez, I.V.; White, L.J.; Gonzalez, O.; Serrano, C.; Giavedoni, L.; Hodara, V.; Cruz, L.; Arana, T.; et al. Zika virus pathogenesis in rhesus macaques is unaffected by pre-existing immunity to dengue virus. Nat. Commun. 2017, 8, 15674. [Google Scholar] [CrossRef] [PubMed]
- Dan, J.M.; Lindestam Arlehamn, C.S.; Weiskopf, D.; da Silva Antunes, R.; Havenar-Daughton, C.; Reiss, S.M.; Brigger, M.; Bothwell, M.; Sette, A.; Crotty, S. A Cytokine-Independent Approach To Identify Antigen-Specific Human Germinal Center T Follicular Helper Cells and Rare Antigen-Specific CD4+ T Cells in Blood. J. Immunol. 2016, 197, 983–993. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Nb of Samples | ZIKV Status 1,2 | DENV Status 1,2 | Age 3 (Year) | Sex (% Male) |
|---|---|---|---|---|---|
| A | 14 | Negative | Positive | 17 (84–1) | 36 |
| B | 9 | Positive | Negative | 30 (184–1) | 56 |
| C | 5 | Positive | Positive | 27 (213–5) | 20 |
| D | 4 | Acute | Positive | 33 (205–4) | 25 |
| E | 2 | Acute | Acute | 27 (252–8) | 50 |
| Peptide 1 | Sequence | SFC/Million PBMC 2 | HLA | Score 3 |
|---|---|---|---|---|
| C869–4 | KDLAAMLRI | 65 | B*55:01 | 1.4 |
| B*40:02 | 2.0 | |||
| C899–7 | AAMLRIINA | 75 | A*02:01 | 4.5 |
| NS1637–7 | MENIMWRSVEGELNA | 310 | DRB1*04:05 | 50 |
| NS1687–6 | WRSVEGELN | 206 | B*40:02 | 48 |
| B*18:01 | 48 | |||
| NS11671–75 | VWLKVREDY | 75 | A*29:02 | 1.3 |
| NS12802–88 | CPGTKVHVE | 170 | B*35:01 | 8.5 |
| B*35:31 | 6.5 | |||
| NS32192–33 | TVILAPTRVVAAEME | 165 | DRB1*08:02 | 1.5 |
| NS33113–25 | AAIFMTATPPGTRDA | 470 | DRB1*04:01 | 4.0 |
| NS33133–21 | IFMTATPPG | 30 | A*24:02 | 5.0 |
| NS33143–22 | FMTATPPGT | 85 | A*02:17 | 5.5 |
| NS4B1121–20 | AIILLVAHY | 88 | A*29:02 | 0.6 |
| A*11:01 | 3.5 | |||
| NS4B1151–23 | LLVAHYMYL | 68 | A*02:05 | 0.3 |
| NS4B1161–24 | LVAHYMYLI | 35 | A*69:01 | 0.15 |
| A*02:05 | 0.2 | |||
| NS52983–06 | NHPYRTWAY 4 | 358 | B*35:01 | 3.0 |
| NS52993–07 | HPYRTWAYH 4 | 308 | B*35:01 | 0.4 |
| NS53023–10 | RTWAYHGSY | 205 | A*01:01 | 0.5 |
| NS53483–56 | TPYGQQRVF 4 | 1681 | B*35:31 | 0.7 |
| B*35:01 | 0.3 | |||
| NS54254–33 | EAVNDPRFW | 465 | B*44:03 | 5.0 |
| B*15:17 | 7.0 | |||
| B*35:01 | 5.0 | |||
| NS54614–75 | KKQGEFGKAKGSRAI | 405 | DRB1*07:01 | 32 |
| NS54734–87 | RAIWYMWLGARFLEF | 505 | DRB1*07:01 | 16 |
| NS56096–17 | YALNTFTNL | 42 | B*35:43 | 0.4 |
| B*35:31 | 0.25 |
| Peptide 1 | Sequence | ZIKV | DENV/ZIKV | Identity (%) | |||||
|---|---|---|---|---|---|---|---|---|---|
| Donors | SFC/million PBMC 2 | Donors | SFC/million PBMC 2 | DENV Serotypes | |||||
| 1 | 2 | 3 | 4 | ||||||
| C49-63 | AILAFLRFTAIKPSL | 60 | 60 | 28,63 | 365 | 60 | 53 | 60 | 40 |
| E67-81 | DMASDSRCPTQGEAY | 33 | 465 | 67 | 53 | 67 | 53 | ||
| E87-101 | DTQYVCKRTLVDRGW | 56 | 505 | 67 | 54 | 73 | 67 | ||
| NS119-33 | VFVYNDVEAWRDRYK | 21,46,60 | 195 | 28,56 | 380 | 47 | 33 | 47 | 40 |
| NS155-69 | CGISSVSRMENIMWR | 35,46 | 125 | 56 | 275 | 67 | 66 | 60 | 60 |
| NS191-105 | GSVKNPMWRGPQRLP | 21,35,46,60 | 275 | 28 | 165 | 13 | 33 | 20 | 33 |
| NS1107-121 | PVNELPHGWKAWGKS | 28,53 | 430 | 40 | 47 | 47 | 50 | ||
| NS1147-161 | HRAWNSFLVEDHGFG | 46 | 40 | 33,53 | 445 | 67 | 73 | 67 | 76 |
| NS1163-177 | FHTSVWLKVREDYSL | 46 | 35 | 20,28,55 | 450 | 47 | 46 | 53 | 47 |
| NS1195-209 | HSDLGYWIESEKNDT | 28,33 | 615 | 80 | 73 | 66 | 73 | ||
| NS2B117-131 | AAGAWYVYVKTGKRS | 55 | 445 | 33 | 33 | 27 | 27 | ||
| NS3131-145 | PAGTSGSPILDKCGR | 21,42 | 405 | 26,55,63 | 495 | 53 | 61 | 53 | 54 |
| NS3143-157 | CGRVIGLYGNGVVIK | 21 | 350 | 20,55,63,66 | 550 | 60 | 67 | 72 | 80 |
| NS3311-325 | AAIFMTATPPGTRDA | 28,33 | 470 | 80 | 80 | 93 | 80 | ||
| NS513-27 | KARLNQMSALEFYSY | 55,69 | 405 | 53 | 47 | 53 | 40 | ||
| NS5293-307 | WFFDENHPYRTWAYH | 55,69 | 1620 | 67 | 67 | 60 | 67 | ||
| NS5297-311 | ENHPYRTWAYHGSYE | 55,69 | 1330 | 80 | 80 | 73 | 80 | ||
| NS5325-339 | VVRLLSKPWDVVTGV | 28,55,66 | 495 | 73 | 80 | 73 | 67 | ||
| NS5345-359 | TDTTPYGQQRVFKEK | 33,55,69 | 4195 | 93 | 93 | 93 | 93 | ||
| NS5373-387 | QVMSMVSSWLWKELG | 60 | 130 | 55,66,69 | 340 | 40 | 53 | 47 | 47 |
| NS5461-475 | KKQGEFGKAKGSRAI | 28,53 | 405 | 93 | 93 | 93 | 87 | ||
| NS5465-479 | EFGKAKGSRAIWYMW | 28,53,55,56 | 1085 | 100 | 100 | 100 | 93 | ||
| NS5473-487 | RAIWYMWLGARFLEF | 28,55 | 505 | 100 | 100 | 93 | 100 | ||
| NS5481-495 | GARFLEFEALGFLNE | 28,53,56,63 | 1870 | 93 | 100 | 93 | 100 | ||
| NS5546-560 | RFDLENEALITNQME | 28,53,66 | 515 | 60 | 47 | 53 | 60 | ||
| NS5573-586 | TYQNKVVKVLRPAEK | 28,53,56 | 615 | 73 | 67 | 73 | 80 | ||
| NS5849-863 | CGSLIGHRPRTTWAE | 60 | 90 | 33,55 | 340 | 67 | 67 | 67 | 67 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delgado, F.G.; Torres, K.I.; Castellanos, J.E.; Romero-Sánchez, C.; Simon-Lorière, E.; Sakuntabhai, A.; Roth, C. Improved Immune Responses Against Zika Virus After Sequential Dengue and Zika Virus Infection in Humans. Viruses 2018, 10, 480. https://doi.org/10.3390/v10090480
Delgado FG, Torres KI, Castellanos JE, Romero-Sánchez C, Simon-Lorière E, Sakuntabhai A, Roth C. Improved Immune Responses Against Zika Virus After Sequential Dengue and Zika Virus Infection in Humans. Viruses. 2018; 10(9):480. https://doi.org/10.3390/v10090480
Chicago/Turabian StyleDelgado, Félix G., Karina I. Torres, Jaime E. Castellanos, Consuelo Romero-Sánchez, Etienne Simon-Lorière, Anavaj Sakuntabhai, and Claude Roth. 2018. "Improved Immune Responses Against Zika Virus After Sequential Dengue and Zika Virus Infection in Humans" Viruses 10, no. 9: 480. https://doi.org/10.3390/v10090480
APA StyleDelgado, F. G., Torres, K. I., Castellanos, J. E., Romero-Sánchez, C., Simon-Lorière, E., Sakuntabhai, A., & Roth, C. (2018). Improved Immune Responses Against Zika Virus After Sequential Dengue and Zika Virus Infection in Humans. Viruses, 10(9), 480. https://doi.org/10.3390/v10090480