
Reconstructing Late-Holocene Paleoenvironments from the World’s Most Inland Rhizophora mangle

 , ,
, ,  , and
, and
Abstract
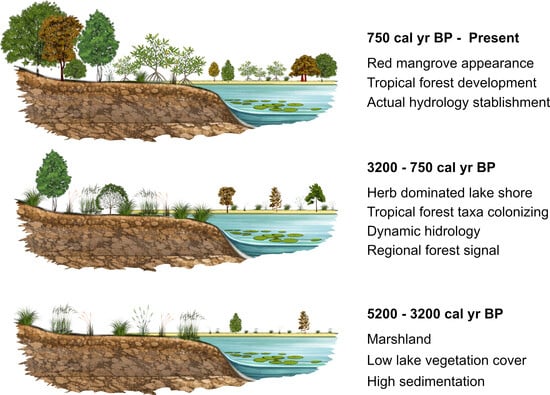
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Laboratory Methods
3. Results
Geochemical Analysis
4. Discussion
4.1. Late-Holocene Dynamics of Rhizophora mangle in Tabasco and the Wider Caribbean Context
4.2. Mangrove Absence and Wetland Conditions Prior to 1250 cal yr BP
4.3. Establishment and Expansion of Rhizophora mangle After 750 cal yr BP
4.4. Persistence and Stability in the Last Millennium
4.5. The Management of the Lower Usumacinta/Candelaria River Basin
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Osland, M.J.; Feher, L.C.; López-Portillo, J.; Day, R.H.; Suman, D.O.; Guzmán Menéndez, J.M.; Rivera-Monroy, V.H. Mangrove Forests in a Rapidly Changing World: Global Change Impacts and Conservation Opportunities along the Gulf of Mexico Coast. Estuar. Coast. Shelf Sci. 2018, 214, 120–140. [Google Scholar] [CrossRef]
- Valderrama-Landeros, L.H.; López-Portillo, J.; Velázquez-Salazar, S.; Alcántara-Maya, J.A.; Troche-Souza, C.; Rodríguez-Zúñiga, M.T.; Vázquez-Balderas, B.; Villeda-Chávez, E.; Cruz-López, M.I.; Ressl, R. Regional Distribution and Change Dynamics of Mangroves in México between 1970/80 and 2015. Wetlands 2020, 40, 1295–1305. [Google Scholar] [CrossRef]
- Velázquez-Salazar, S.; Rodríguez-Zúñiga, M.T.; Alcántara-Maya, J.A.; Villeda-Chávez, E.; Valderrama-Landeros, L.; Troche-Souza, C.; Vázquez-Balderas, B.; Pérez-Espinosa, I.; Cruz-López, M.I.; Ress, I.R.; et al. Manglares de México. Actualización Y Análisis de Los Datos 2020; Comisión Nacional para el Conocimiento y Uso de la Biodiversidad: Mexico City, Mexico, 2021. [Google Scholar]
- Maxwell, T.L.; Spalding, M.D.; Friess, D.A.; Murray, N.J.; Rogers, K.; Rovai, A.S.; Smart, L.S.; Weilguny, L.; Adame, M.F.; Adams, J.B.; et al. Soil Carbon in the World’s Tidal Marshes. Nat. Commun. 2024, 15, 10265. [Google Scholar] [CrossRef]
- Alongi, D.M. Mangrove Forests: Resilience, Protection from Tsunamis, and Responses to Global Climate Change. Estuar. Coast. Shelf Sci. 2008, 76, 1–13. [Google Scholar] [CrossRef]
- Portillo, J.L.; Ezcurra, E. Los Manglares de México: Una Revisión. Madera Bosques 2002, 8, 27–51. [Google Scholar] [CrossRef]
- Rull, V. Rise and Fall of Caribbean Mangroves. Sci. Total Environ. 2023, 885, 163851. [Google Scholar] [CrossRef]
- Aragón-Moreno, A.A.; Islebe, G.A.; Roy, P.D.; Torrescano-Valle, N.; Mueller, A.D. Climate Forcings on Vegetation of the Southeastern Yucatán Peninsula (Mexico) during the Middle to Late Holocene. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2018, 495, 214–226. [Google Scholar] [CrossRef]
- Monacci, N.M.; Meier-Grünhagen, U.; Finney, B.P.; Behling, H.; Wooller, M.J. Paleoecology of Mangroves along the Sibun River, Belize. Quat. Res. 2011, 76, 220–228. [Google Scholar] [CrossRef]
- Goldberg, L.; Lagomasino, D.; Thomas, N.; Fatoyinbo, T. Global Declines in Human-Driven Mangrove Loss. Glob. Change Biol. 2020, 26, 5844–5855. [Google Scholar] [CrossRef] [PubMed]
- Rull, V. Origin and Evolution of Caribbean Mangroves: A Time-Continuum Ecological Approach, 2024th ed.; Ecological Studies: Analysis and Synthesis; Springer International Publishing: Cham, Switzerland, 2024. [Google Scholar]
- Rull, V. Responses of Caribbean Mangroves to Quaternary Climatic, Eustatic, and Anthropogenic Drivers of Ecological Change: A Review. Plants 2022, 11, 3502. [Google Scholar] [CrossRef]
- Aragón-Moreno, A.A.; Islebe, G.A.; Torrescano-Valle, N.; Hernández-Arana, H.A.; Carrillo-Bastos, A.; Friedel, R. Mangrove Dynamics Driven by Late Holocene Drought at Laguna Bacalar, Mexico. Quat. Environ. Hum. 2024, 2, 100023. [Google Scholar] [CrossRef]
- Aburto-Oropeza, O.; Burelo-Ramos, C.M.; Ezcurra, E.; Ezcurra, P.; Henriquez, C.L.; Vanderplank, S.E.; Zapata, F. Relict Inland Mangrove Ecosystem Reveals Last Interglacial Sea Levels. Proc. Natl. Acad. Sci. USA 2021, 118, e2024518118. [Google Scholar] [CrossRef]
- Peralta-Carreta, C.; Medrano-Pérez, O.R.; Alcudia-Aguilar, A.; Pham, Q.B.; Megia-Vera, H.J. Spatial Distribution of Relict Inland Mangrove (Rhizophora mangle L.) in the San Pedro River Basin: A Transboundary Analysis between Mexico and Guatemala. Estuaries Coast. 2025, 48, 59. [Google Scholar] [CrossRef]
- Hellmuth, N.; Jerez, S.D.; López Dobrusin, E.; Morales Rodríguez, N.; Mendoza, V. (Eds.) Mangle Rojo (Rhizophora mangle) en las Riberas del Río San Pedro; FLAAR Mesoamérica: Guatemala City, Guatemala, 2024. [Google Scholar]
- CONANP. Programa de Manejo Área de Protección de Flora y Fauna Cañon del Usumacinta; Secretaría de Medio Ambiente, Recursos Naturales & Pesca: Mexico City, Mexico, 2015. [Google Scholar]
- CONANP. Estudio Previo Justificativo Para el Establecimiento del Área Natural Protegida Reserva de la Biósfera Wanha’ Tabasco; CONANP: Mexico City, Mexico, 2023. [Google Scholar]
- Faegri, K.; Iversen, J. Textbook of Pollen Analysis, 4th ed.; Faegri, K., Kaland, P.E., Krzywinski, K., Eds.; John Wiley: Hoboken, NJ, USA, 1989. [Google Scholar]
- Palacios-Chávez, R.; Ludlow-Wiechers, B.; Villanueva, R. Flora Palinológica de La Reserva de La Biosfera de Sian Ka’an, Quintana Roo, México; Centro de Investigaciones de Quintana Roo: Chetumal, Mexico, 1991. [Google Scholar]
- Grimm, E.C. Tilia, version 1.7.16; Illinois State Museum, Research and Collection Center: Springfield, IL, USA, 2011. [Google Scholar]
- Navarro, E.; Daesslé, L.W.; Camacho-Ibar, V.F.; Ortiz-Hernández, M.C.; Gutiérrez-Galindo, E.A. La geoquímica de Fe, Ti y Al como indicadora de la sedimentación volcanoclástica en la laguna costera de San Quintín, Baja California, México. Cienc. Mar. 2006, 32, 205–217. [Google Scholar] [CrossRef]
- Urrego, L.E.; Correa-Metrio, A.; González, C.; Castaño, A.R.; Yokoyama, Y. Contrasting Responses of Two Caribbean Mangroves to Sea-Level Rise in the Guajira Peninsula (Colombian Caribbean). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2013, 370, 92–102. [Google Scholar] [CrossRef]
- Bocanegra-Ramírez, D.M.; Li, H.-C.; Vázquez, G.D.; Israde-Alcántara, I.; Bischoff, J.L. Holocene Climate Change and Sea Level Oscillations in the Pacific Coast of Mexico. Quat. Int. 2019, 528, 100–108. [Google Scholar] [CrossRef]
- Espíndola, J.M.; Macías, J.L.; Tilling, R.I.; Sheridan, M.F. Volcanic History of El Chichón Volcano (Chiapas, Mexico) during the Holocene, and Its Impact on Human Activity. Bull. Volcanol. 2000, 62, 90–104. [Google Scholar] [CrossRef]
- Macías, J.L.; Arce, J.L.; Mora, J.C.; Espíndola, J.M.; Saucedo, R.; Manetti, P. A 550-year-old Plinian Eruption at El Chichón Volcano, Chiapas, Mexico: Explosive Volcanism Linked to Reheating of the Magma Reservoir. J. Geophys. Res. 2003, 1–18. [Google Scholar] [CrossRef]
- Nooren, C.; Hoek, W.; Tebbens, L.; Pozzo, A. Tephrochronological Evidence for the Late Holocene Eruption History of El Chichón Volcano, Mexico. Geofis. Int. 2009, 48, 97–112. [Google Scholar] [CrossRef]
- Tilling, R.I.; Rubin, M.; Sigurdsson, H.; Carey, S.; Duffield, W.A.; Rose, W.I. Holocene Eruptive Activity of El Chichón Volcano, Chiapas, Mexico. Science 1984, 224, 747–749. [Google Scholar] [CrossRef] [PubMed]
- Monacci, N.M.; Meier-Grünhagen, U.; Finney, B.P.; Behling, H.; Wooller, M.J. Mangrove Ecosystem Changes during the Holocene at Spanish Lookout Cay, Belize. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2009, 280, 37–46. [Google Scholar] [CrossRef]
- Jones, M.C.; Wingard, G.L.; Stackhouse, B.; Keller, K.; Willard, D.; Marot, M.; Landacre, B.; E Bernhardt, C. Rapid Inundation of Southern Florida Coastline despite Low Relative Sea-Level Rise Rates during the Late-Holocene. Nat. Commun. 2019, 10, 3231. [Google Scholar] [CrossRef]
- Yao, Q.; Liu, K.-B.; Platt, W.J.; Rivera-Monroy, V.H. Palynological Reconstruction of Environmental Changes in Coastal Wetlands of the Florida Everglades since the Mid-Holocene. Quat. Res. 2015, 83, 449–458. [Google Scholar] [CrossRef]
- Medina-Elizalde, M.; Rohling, E.J. Collapse of Classic Maya Civilization Related to Modest Reduction in Precipitation. Science 2012, 335, 956–959. [Google Scholar] [CrossRef]
- Medina-Elizalde, M.; Burns, S.J.; Polanco-Martínez, J.M.; Beach, T.; Lases-Hernández, F.; Shen, C.-C.; Wang, H.-C. High-Resolution Speleothem Record of Precipitation from the Yucatan Peninsula Spanning the Maya Preclassic Period. Glob. Planet. Change 2016, 138, 93–102. [Google Scholar] [CrossRef]
- Lozano-García, M.d.S.; Caballero, M.; Ortega, B.; Rodriguez, A.; Sosa, S. Tracing the Effects of the Little Ice Age in the Tropical Lowlands of Eastern Mesoamerica. Proc. Natl. Acad. Sci. USA 2007, 104, 16200–16203. [Google Scholar] [CrossRef] [PubMed]
- Islebe, G.A.; Carrillo-Bastos, A.; Aragón-Moreno, A.A.; Valdez-Hernández, M.; Torrescano-Valle, N.; Cabanillas-Terán, N. Holocene Paleoecology and Paleoclimatology of South and South-Eastern Mexico: A Palynological and Geospatial Approach. In The Holocene and Anthropocene Environmental History of Mexico: A Paleoecological Approach on Mesoamerica; Torrescano-Valle, N., Islebe, G.A., Roy, P., Eds.; Springer: Cham, Switzerland, 2019; pp. 195–208. [Google Scholar]
- Wooller, M.J.; Behling, H.; Smallwood, B.J.; Fogel, M. Mangrove Ecosystem Dynamics and Elemental Cycling at Twin Cays, Belize, during the Holocene. J. Quat. Sci. 2004, 19, 703–711. [Google Scholar] [CrossRef]
- Liendo, R.; Solleiro-Rebolledo, E.; Solis-Castillo, B.; Sedov, S.; Ortiz-Pérez, A. 7 Population Dynamics and Its Relation to Ancient Landscapes in the Northwestern Maya Lowlands: Evaluating Resilience and Vulnerability: Evaluating Resilience and Vulnerability. Archeol. Pap. Am. Anthropol. Assoc. 2014, 24, 84–100. [Google Scholar]
- Muñoz-Salinas, E.; Cook, D.; Castillo, M.; Beach, T.; Luzzadder-Beach, S. Four Millennia of Geomorphic Change and Human Settlement in the Lower Usumacinta–Grijalva River Basin, Mexico. Prog. Phys. Geogr. 2023, 47, 227–248. [Google Scholar] [CrossRef]
- Matheny, R.; Gurr, D. Variation in Prehistoric Agricultural Systems of the New World. Annu. Rev. Anthropol. 1983, 12, 79–103. [Google Scholar] [CrossRef]
- Puleston, D.E. Terracing, Raised Fields, and Tree Cropping in the Maya Lowlands: A New Perspective on the Geography of Power. In Prehispanic Maya Agriculture; Harrison, P.D., Turner, B.L., Eds.; University of New Mexico Press: Albuquerque, NM, USA, 1978; pp. 225–246. [Google Scholar]
- Siemens, A.; Graham, J.A.; Hebda, R.; Heimo, M. “DAMS” on the Candelaria. Anc. Mesoam. 2002, 13, 115–123. [Google Scholar] [CrossRef]
- Gunn, J.D.; Folan, W.J.; Torrescano-Valle, N.; Faust, B.B.; Geovannini-Acuña, H.Z.; Siemens, A.H. From Calakmul to the Sea: The Historical Ecology of a Classic Maya City That Controlled the Candelaria/champoton Watersheds. In The Holocene and Anthropocene Environmental History of Mexico; Springer International Publishing: Cham, Switzerland, 2019; pp. 209–248. [Google Scholar]
- Palka, J. Ancestral Maya Domesticated Waterscapes, Ecological Aquaculture, and Integrated Subsistence. Anc. Mesoam. 2023, 35, 208–236. [Google Scholar] [CrossRef]
- Woodfill, B.K.S.; Andrieu, C. Tikal’s Early Classic Domination of the Great Western Trade Route: Ceramic, Lithic, and Iconographic Evidence. Anc. Mesoam. 2012, 23, 189–209. [Google Scholar] [CrossRef]
- Freidel, D.; Escobedo, H.; Lee, D.; Guenter, S.; Meléndez, J.C. El Perú-Waka’ Y La Ruta Terrestre de La Dinastía Kan Hacia El Altiplano. In XX Simposio de Investigaciones Arqueológicas en Guatemala; Laporte, J.P., Arroyo, B., Mejía, H., Eds.; Museo Nacional de Arqueología y Etnología: Guatemala City, Guatemala, 2006; pp. 59–76. [Google Scholar]
- Krauss, K.W.; Ball, M.C. On the Halophytic Nature of Mangroves. Trees 2013, 27, 7–11. [Google Scholar] [CrossRef]
- Flowers, T.J.; Colmer, T.D. Salinity Tolerance in Halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ICA ID | Depth (cm) | Conventional Age (yr BP) | Material |
|---|---|---|---|
| 14C-6641 | 43 | 290 ± 30 BP | organic sediment |
| 14C-6642 | 67 | 840 ± 30 BP | organic sediment |
| 14C-6643 | 81 | 1580 ± 30 BP | organic sediment |
| 14C-6644 | 113 | 6810 ± 30 BP | seed |
| 14C-6645 | 169 | 4380 ± 40 BP | organic sediment |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Share and Cite
Islebe, G.A.; Burelo-Ramos, C.M.; Aragón-Moreno, A.A.; Torrescano-Valle, N.; Hernández-Arana, H.A.; Ascencio-Rivera, J.M. Reconstructing Late-Holocene Paleoenvironments from the World’s Most Inland Rhizophora mangle. Forests 2026, 17, 303. https://doi.org/10.3390/f17030303
Islebe GA, Burelo-Ramos CM, Aragón-Moreno AA, Torrescano-Valle N, Hernández-Arana HA, Ascencio-Rivera JM. Reconstructing Late-Holocene Paleoenvironments from the World’s Most Inland Rhizophora mangle. Forests. 2026; 17(3):303. https://doi.org/10.3390/f17030303
Chicago/Turabian StyleIslebe, Gerald Alexander, Carlos M. Burelo-Ramos, Alejandro Antonio Aragón-Moreno, Nuria Torrescano-Valle, Héctor Abuid Hernández-Arana, and Jesús Manuel Ascencio-Rivera. 2026. "Reconstructing Late-Holocene Paleoenvironments from the World’s Most Inland Rhizophora mangle" Forests 17, no. 3: 303. https://doi.org/10.3390/f17030303
APA StyleIslebe, G. A., Burelo-Ramos, C. M., Aragón-Moreno, A. A., Torrescano-Valle, N., Hernández-Arana, H. A., & Ascencio-Rivera, J. M. (2026). Reconstructing Late-Holocene Paleoenvironments from the World’s Most Inland Rhizophora mangle. Forests, 17(3), 303. https://doi.org/10.3390/f17030303