
The First Report of Hypogeous Geastrum in Hungary in Mattirolomyces terfezioides-Producing Forests in the Carpathian Basin


 ,
,
Abstract
1. Introduction
2. Materials and Methods
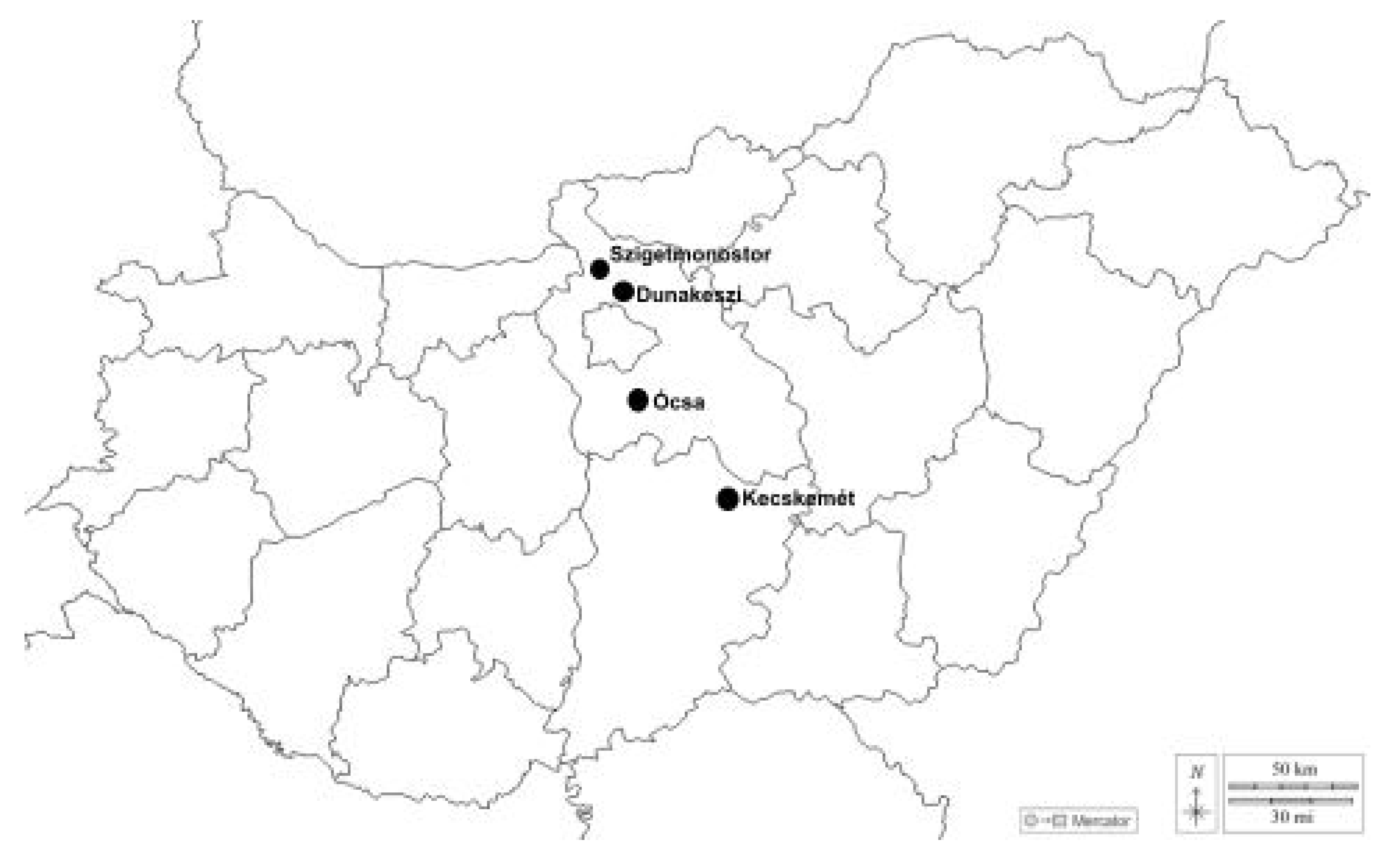
2.1. Study Area
2.2. Fungi Collection, Sampling
2.3. Soil Sampling
2.4. Morphological and Molecular Study
3. Results
3.1. Macromorphological Characteristics
3.2. Micromorphological Characteristics
3.3. Phlyogenetic Analysis
3.4. Soil Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jeppson, M.; Nilsson, R.H.; Larsson, E. European earthstars in Geastraceae (Geastrales, Phallomycetidae)—A systematic approach using morphology and molecular sequence data. Syst. Biodivers. 2013, 11, 437–465. [Google Scholar] [CrossRef]
- Agerer, R. Geastrum fimbriatum Fr. + Fagus sylvatica L. Descr. Ectomyc. 1998, 3, 13–18. [Google Scholar]
- Dominguez de Toledo, L.S.; Castellano, M.A. A revision of the genera Radiigera and Pyrenogaster. Mycologia 1996, 88, 863–884. [Google Scholar] [CrossRef]
- Moreno, G.; Alvarado, P.; Manjón, J.L. Hypogeous Desert Fungi. In Desert Truffles; Springer: Berlin/Heidelberg, Germany, 2014; pp. 3–20. [Google Scholar]
- Zeller, S.M. Representatives of the Mesophelliaceae in North America. Mycologia 1944, 36, 627–637. [Google Scholar] [CrossRef]
- Askew, B.; Miller, O.K., Jr. New evidence of close relationships between Radiigera and Geastrum (Lycoperdales). Can. J. Bot. 1977, 55, 2693–2700. [Google Scholar] [CrossRef]
- Zamora, J.C.; de Diego Calonge, F.; Hosaka, K.; Martín, M.P. Systematics of the genus Geastrum (Fungi: Basidiomycota) revisited. Taxon 2014, 63, 477–497. [Google Scholar] [CrossRef]
- Estrada-Torres, A.; Gaither, T.W.; Miller, D.L.; Lado, C.; Keller, H.W. The myxomycete genus Schenella: Morphological and DNA sequence evidence for synonymy with the gasteromycete genus Pyrenogaster. Mycologia 2005, 97, 139–149. [Google Scholar] [CrossRef]
- Quadraccia, L. Studies on Italian gasteromycetes. I. Two new species of Arachnion and Radiigera (Basidiomycotina, Lycoperdales) from Rome and its environs. Mycotaxon 1996, 58, 331–341. [Google Scholar]
- Montecchi, A.; Sarasini, M. Funghi Ipogei D’Europa; A.M.B. Fondazione Centro Studi Micologici: Vicenza, Italy, 2000. [Google Scholar]
- Borhidi, A. Magyarország Növénytársulásai; Akadémiai Kiadó: Budapest, Hungary, 2003. [Google Scholar]
- Csorbainé, A.G.; Bratek, Z.; Merényi, Z.; Illyès, Z.; Dimény, J. Choiromyces meandriformis and Mattirolomyces terfezioides: Peculiar truffles with new perspectives. In Proceedings of the XVII Convegno Nazionale di Micologia, Pavia, Italy, 10–12 November 2008; pp. 10–12. [Google Scholar]
- Bratek, Z.; Jakucs, E.; Bóka, K.; Szedlay, G. Mycorrhizae between black locust (Robinia pseudoacacia) and Terfezia terfezioides. Mycorrhiza 1996, 6, 271–274. [Google Scholar] [CrossRef]
- Merényi, Z.; Illyés, Z.; Völcz, G.; Bratek, Z. A database and its application for the development of truffle cultivation methods. Osterr. Z. Für Pilzkd. 2010, 19, 239–244. [Google Scholar]
- Vierheilig, H.; Coughlan, A.P.; Wyss, U.R.S.; Piché, Y. Ink and vinegar, a simple staining technique for arbuscular-mycorrhizal fungi. Appl. Environ. Microbiol. 1998, 64, 5004–5007. [Google Scholar] [CrossRef] [PubMed]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Katoh, K.; Toh, H. Recent developments in the MAFFT multiple sequence alignment program. Brief. Bioinform. 2008, 9, 286–298. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Glez-Peña, D.; Gómez-Blanco, D.; Reboiro-Jato, M.; Fdez-Riverola, F.; Posada, D. ALTER: Program-oriented format conversion of DNA and protein alignments. Nucleic Acids Res. 2010, 38, W14–W18. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Posada, D. jModelTest: Phylogenetic model averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree. Tree Figure Drawing Tool. 2009. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 25 May 2025).
- Cseh, P.; Merényi, Z.; Bóna, L.; Varga, T.; Bóka, K.; Nagy, I.; Kaounas, V.; Vidal, J.M.; Paz, A.; Bratek, Z. Taxonomic characterisation of the Regianum clade (genus Tuber) and the trait evolution of spore size among true truffles. Mycol. Progress. 2024, 23, 11. [Google Scholar] [CrossRef]
- Paz, A.; Lavoise, C.; Chautrand, P.; Slavova, M.; Daniëls, P.P.; Rojo, C. Geastrum nadalii sp. nov. In: Crous, P.W.; et al. Fungal Planet description sheets: 1614–1696. Fungal Syst. Evol. 2024, 13, 241–242. [Google Scholar]
- Sousa, J.O.; Morais, L.A.; Nascimento, Y.M.; Baseia, I.G. Updates on the geographic distribution of three Geastrum species from Brazilian semi-arid region. Mycosphere 2014, 5, 467–474. [Google Scholar] [CrossRef]
- Čejka, T.; Trnka, M.; Büntgen, U. Sustainable cultivation of the white truffle (Tuber magnatum) requires ecological understanding. Mycorrhiza 2023, 33, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Belfiori, B.; D’Angelo, V.; Riccioni, C.; Leonardi, M.; Paolocci, F.; Pacioni, G.; Rubini, A. Genetic structure and phylogeography of Tuber magnatum populations. Diversity 2020, 12, 44. [Google Scholar] [CrossRef]
- Bratek, Z.; Gogan, A.; Halasz, K.; Bagi, I.; Erdei, V.; Bujaku, G. The northernmost habitats of Tuber magnatum known from Hungary. In Proceedings of the First Hypogean Mushroom Conference, Rabat, Morocco, 6–8 April 2004; Kabar, L., Ed.; Rabat Net: Rabat, Morocco, 2004; pp. 28–30. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lab Herbarium Code | Geographical Microregion | Locality | Date of Collection | Number of Collected Basidiomata | GenBank Voucher | GenBank Accession Number |
|---|---|---|---|---|---|---|
| ZB4964 | Kiskunság | Near Kecskemét | 14 October 2013 | 1 | no | no |
| ZB5788 | Pest Plain | Ócsa | 16 October 2019 | 5 | no | no |
| ZB5906 | Pest Plain | Dunakeszi | 21 October 2021 | 2 | Rad1 | PQ784779 |
| ZB5972 | Szentendre Island | Szigetmonostor | 2 October 2022 | 9 | Rad2 | PQ784780 |
| ZB5978 | Pest Plain | Ócsa | 9 November 2022 | 12 | Rad3 | PQ784781 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heller, Á.; Péter, B.; Cseh, P.; Habtemariam, A.A.; Nagy, I.; Bratek, Z. The First Report of Hypogeous Geastrum in Hungary in Mattirolomyces terfezioides-Producing Forests in the Carpathian Basin. Forests 2025, 16, 1136. https://doi.org/10.3390/f16071136
Heller Á, Péter B, Cseh P, Habtemariam AA, Nagy I, Bratek Z. The First Report of Hypogeous Geastrum in Hungary in Mattirolomyces terfezioides-Producing Forests in the Carpathian Basin. Forests. 2025; 16(7):1136. https://doi.org/10.3390/f16071136
Chicago/Turabian StyleHeller, Ádám, Balázs Péter, Péter Cseh, Akale Assamere Habtemariam, István Nagy, and Zoltán Bratek. 2025. "The First Report of Hypogeous Geastrum in Hungary in Mattirolomyces terfezioides-Producing Forests in the Carpathian Basin" Forests 16, no. 7: 1136. https://doi.org/10.3390/f16071136
APA StyleHeller, Á., Péter, B., Cseh, P., Habtemariam, A. A., Nagy, I., & Bratek, Z. (2025). The First Report of Hypogeous Geastrum in Hungary in Mattirolomyces terfezioides-Producing Forests in the Carpathian Basin. Forests, 16(7), 1136. https://doi.org/10.3390/f16071136