
Estimation of Tree Species Diversity in Warm Temperate Forests via GEDI and GF-1 Imagery

Abstract
1. Introduction
2. Materials and Methods
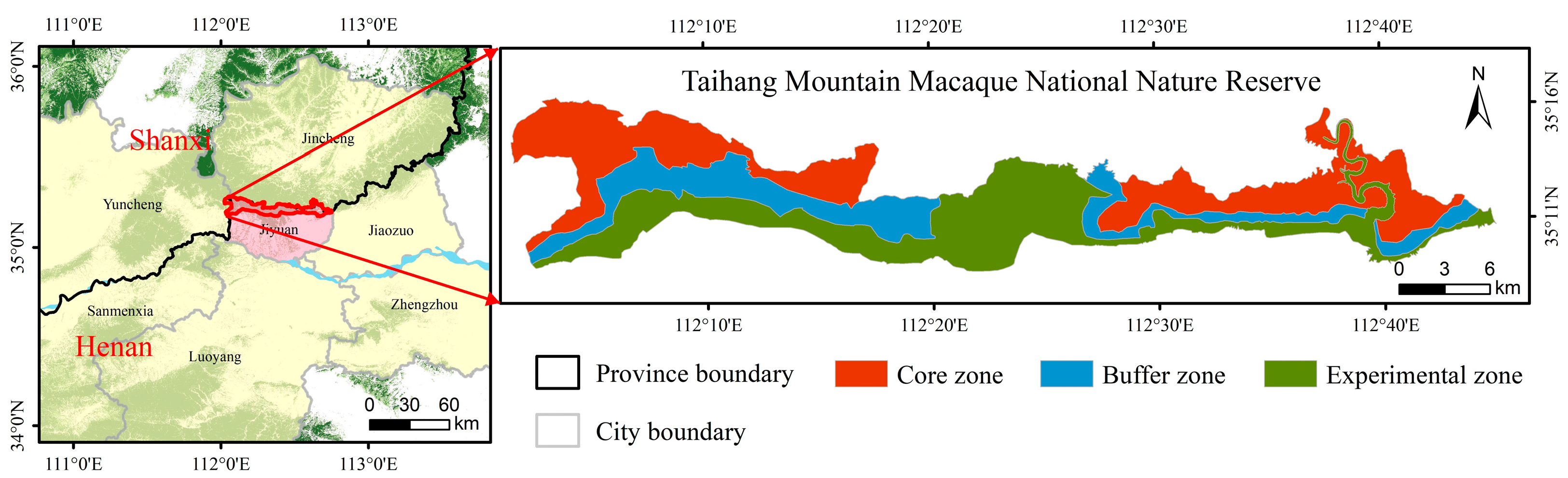
2.1. Study Area
2.2. Field Investigation of Sample Plots
2.3. Tree Species Diversity Indices
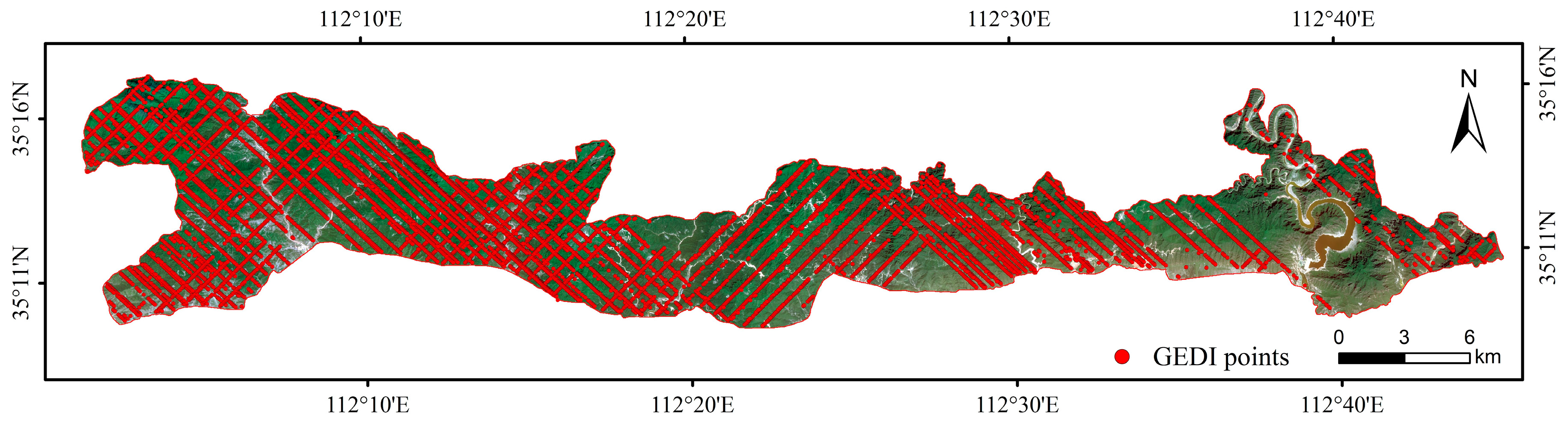
2.4. GEDI Data and Preprocessing
2.5. GF-1 Data and Preprocessing
2.6. Remote Sensing Feature Selection
2.7. Modeling Method
2.7.1. Random Forest
2.7.2. Support Vector Machines
2.7.3. K-Nearest Neighbors
2.7.4. Random Forest Plus Residual Kriging
2.8. Precision Evaluation Indices
3. Results
3.1. RFRK Interpolation
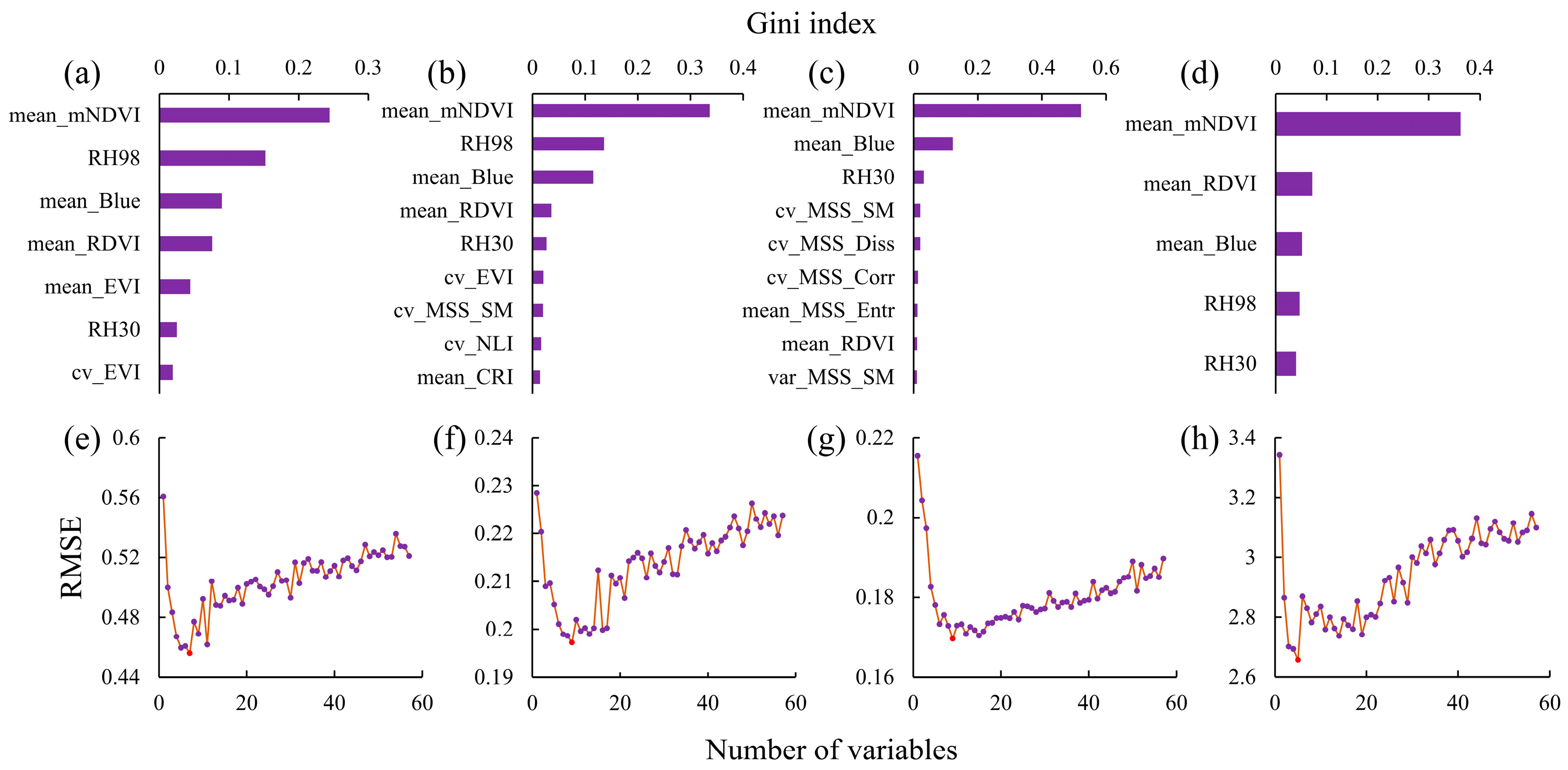
3.2. Feature Selection and Importance Ranking
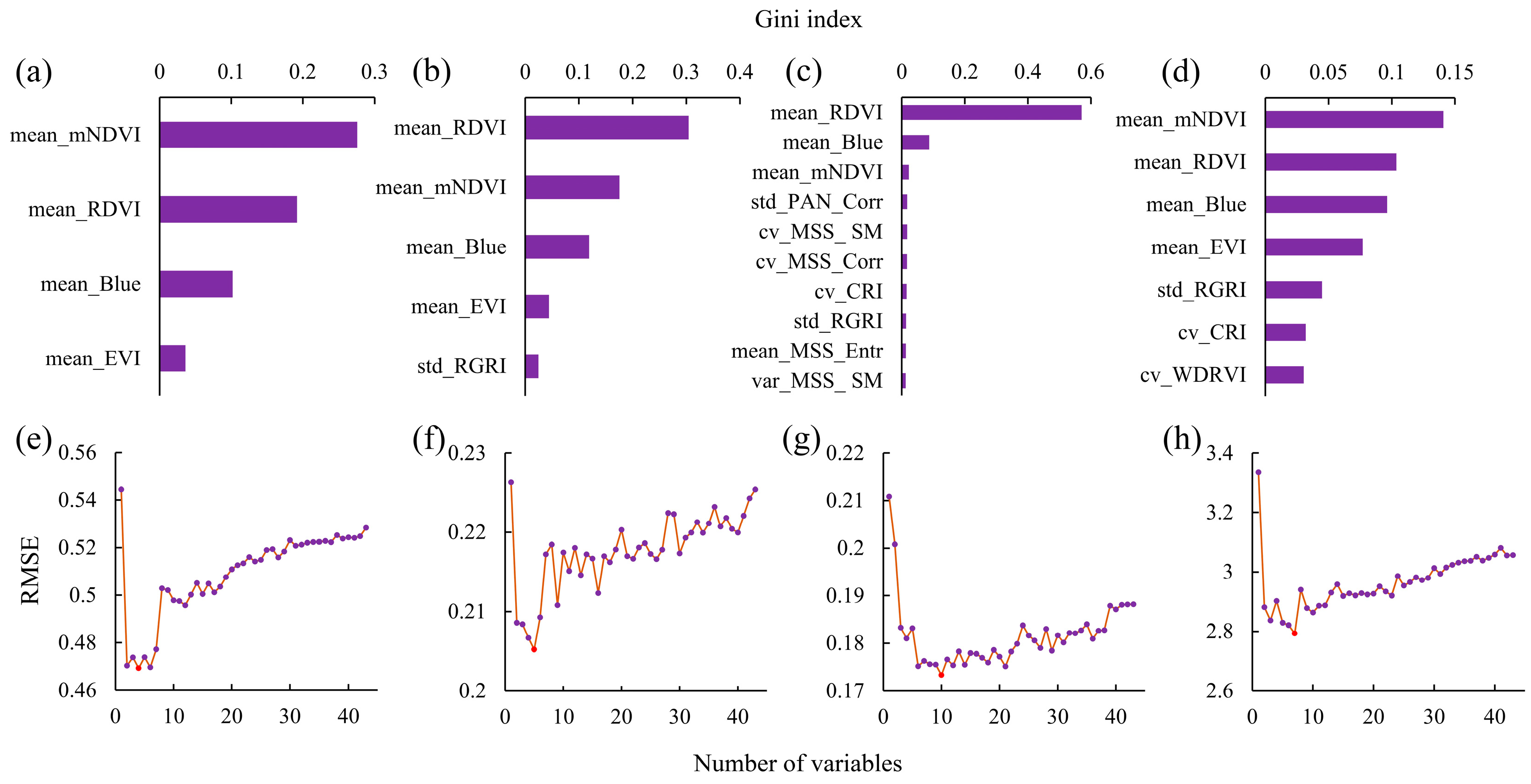
3.2.1. Spectral Feature Screening
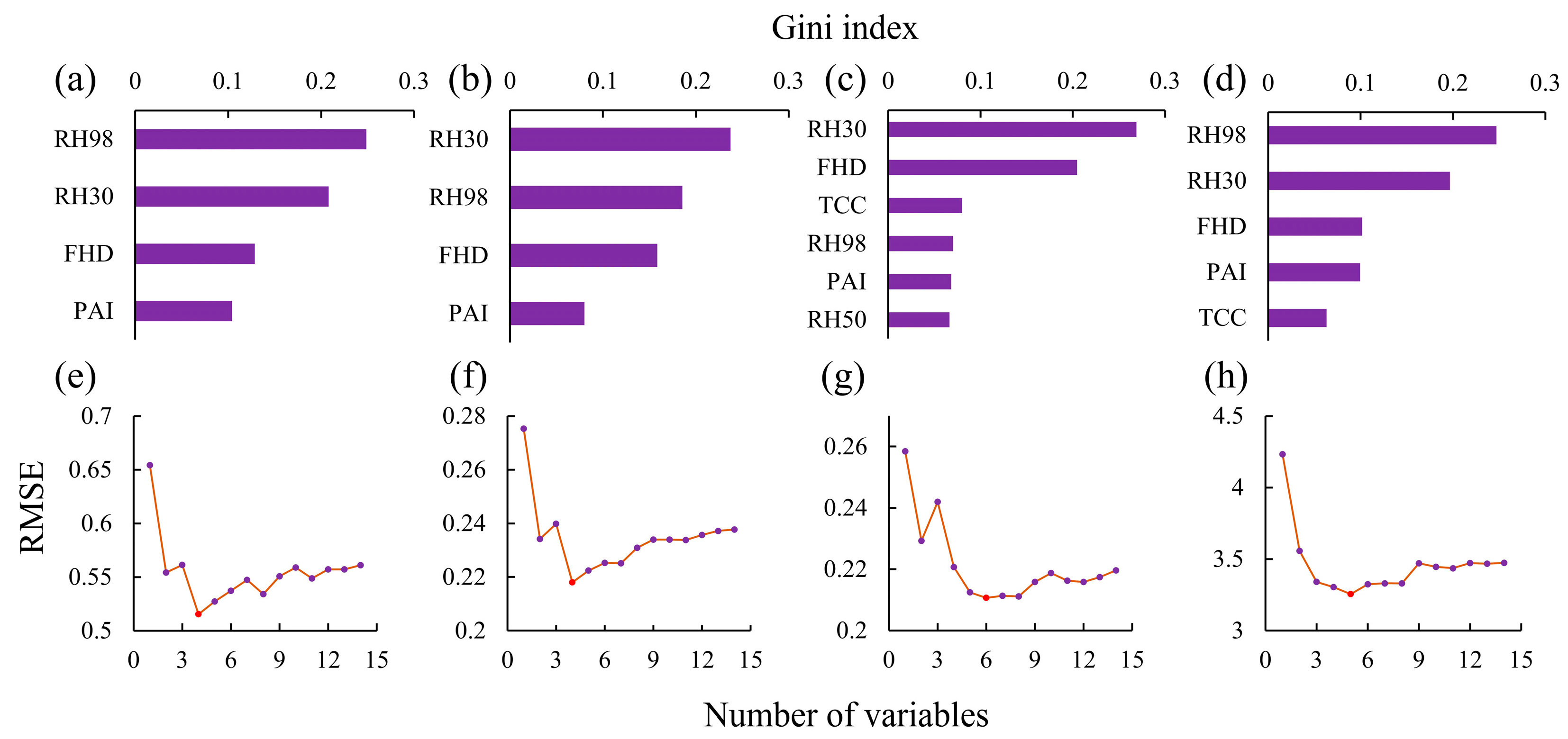
3.2.2. Vertical Structural Feature Screening
3.2.3. Fusion Feature Screening
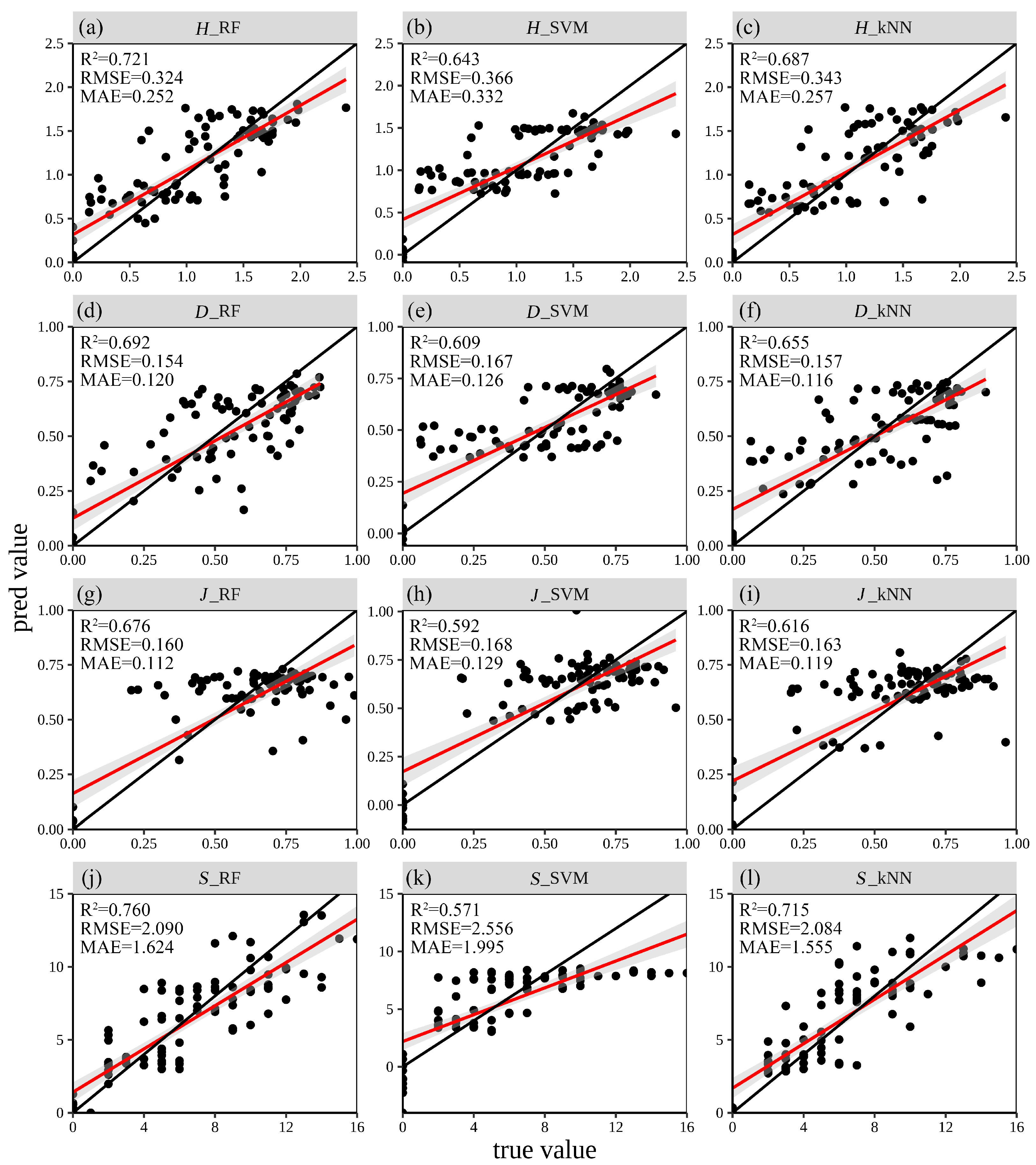
3.3. Estimation Accuracy
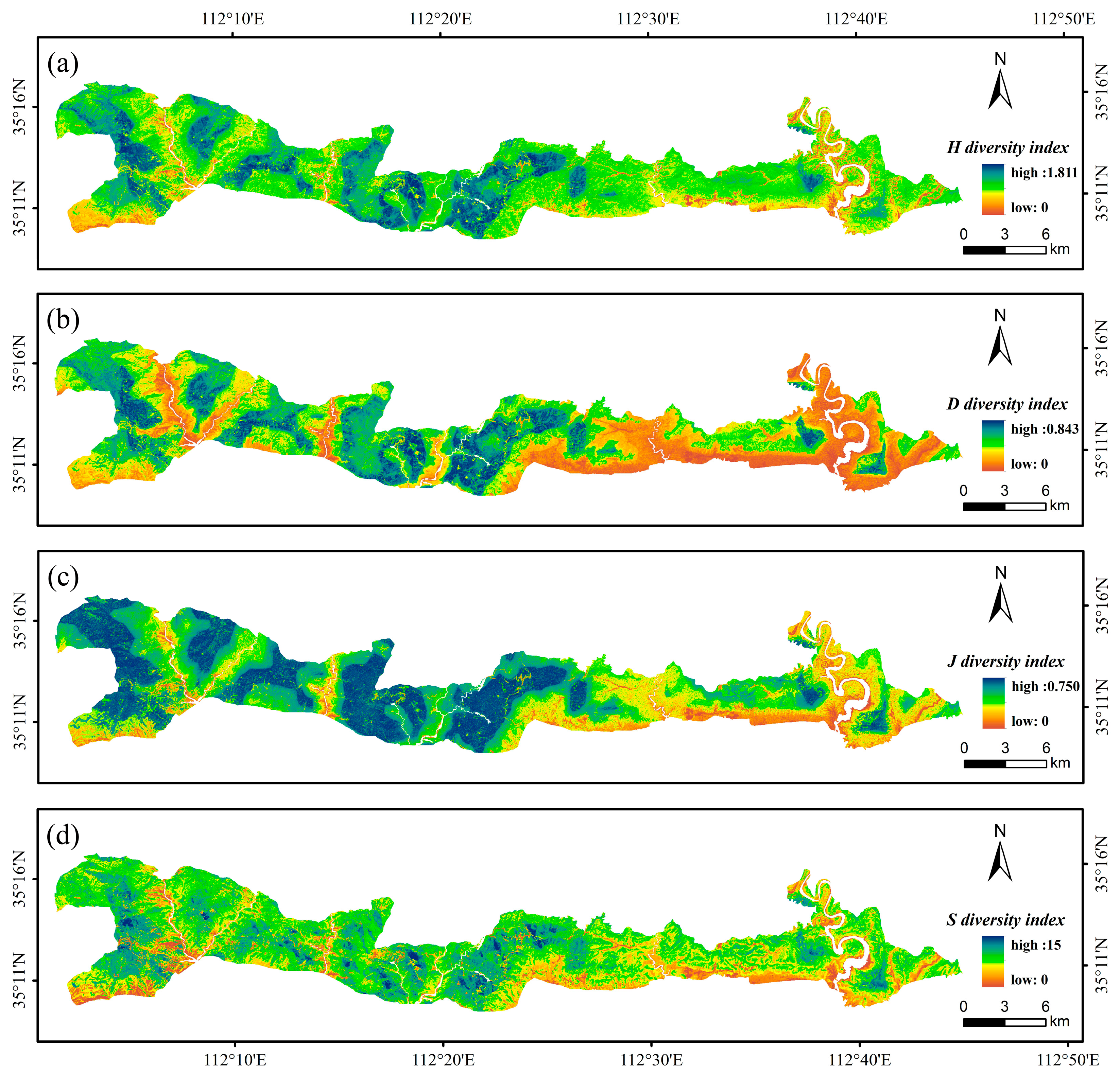
3.4. The Spatial Distribution of Tree Species Diversity
4. Discussion
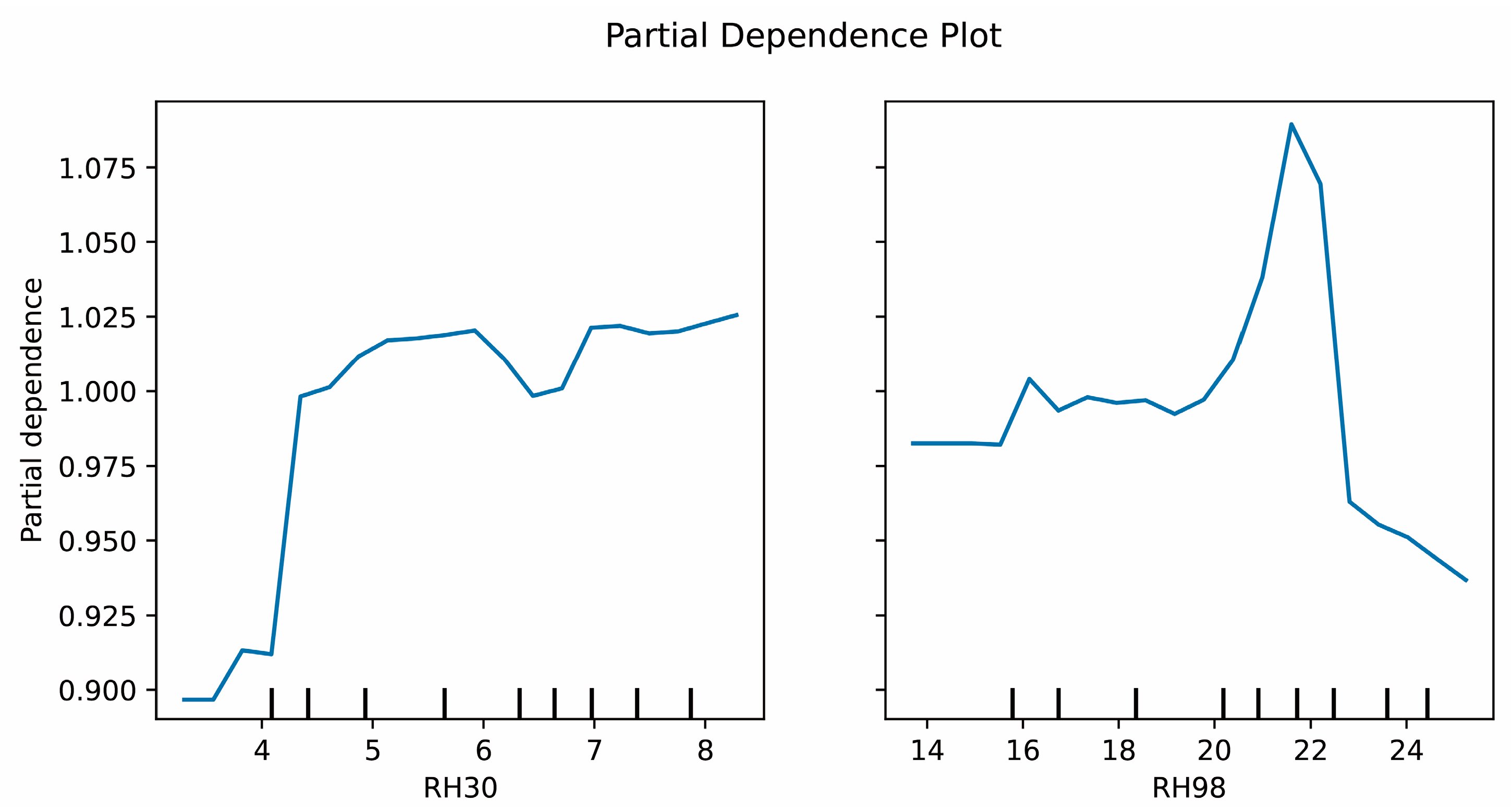
4.1. Estimation of Tree Species Diversity on the Basis of GF-1 and GEDI Data
4.2. Performance Analysis of Three Machine Learning Methods in Tree Species Diversity Modeling
4.3. Spatial Distribution of the Tree Species Diversity Indices
4.4. Limitations and Prospects of This Study
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Barlow, J.; Lennox, G.D.; Ferreira, J.; Berenguer, E.; Lees, A.C.; Nally, R.M.; Thomson, J.R.; Ferraz, S.F.d.B.; Louzada, J.; Oliveira, V.H.F.; et al. Anthropogenic disturbance in tropical forests can double biodiversity loss from deforestation. Nature 2016, 535, 144–147. [Google Scholar] [CrossRef] [PubMed]
- Brockerhoff, E.G.; Barbaro, L.; Castagneyrol, B.; Forrester, D.I.; Gardiner, B.; González-Olabarria, J.R.; Lyver, P.O.B.; Meurisse, N.; Oxbrough, A.; Taki, H.; et al. Forest biodiversity, ecosystem functioning and the provision of ecosystem services. Biodivers. Conserv. 2017, 26, 3005–3035. [Google Scholar] [CrossRef]
- Ćosović, M.; Bugalho, M.N.; Thom, D.; Borges, J.G. Stand Structural Characteristics Are the Most Practical Biodiversity Indicators for Forest Management Planning in Europe. Forests 2020, 11, 343. [Google Scholar] [CrossRef]
- Silaeva, T.; Andreychev, A.; Kiyaykina, O.; Balčiauskas, L. Taxonomic and ecological composition of forest stands inhabited by forest dormouse Dryomys nitedula (Rodentia: Gliridae) in the Middle Volga. Biologia 2021, 76, 1475–1482. [Google Scholar] [CrossRef]
- Immitzer, M.; Atzberger, C.; Koukal, T. Tree Species Classification with Random Forest Using Very High Spatial Resolution 8-Band WorldView-2 Satellite Data. Remote Sens. 2012, 4, 2661–2693. [Google Scholar] [CrossRef]
- Van der Putten, W.H.; Macel, M.; Visser, M.E. Predicting species distribution and abundance responses to climate change: Why it is essential to include biotic interactions across trophic levels. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2025–2034. [Google Scholar] [CrossRef]
- Gu, X.; Jamshidi, S.; Gu, L.; Nadi, S.; Wang, D.; Cammarano, D.; Sun, H. Drought dynamics in California and Mississippi: A wavelet analysis of meteorological to agricultural drought transition. J. Environ. Manag. 2024, 370, 122883. [Google Scholar] [CrossRef]
- Vitasse, Y.; Ursenbacher, S.; Klein, G.; Bohnenstengel, T.; Chittaro, Y.; Delestrade, A.; Monnerat, C.; Rebetez, M.; Rixen, C.; Strebel, N.; et al. Phenological and elevational shifts of plants, animals and fungi under climate change in the European Alps. Biol. Rev. 2021, 96, 1816–1835. [Google Scholar] [CrossRef]
- Dehghanpir, S.; Bazrafshan, O.; Nadi, S.; Jamshidi, S. Assessing the Sustainability of Agricultural Water Use Based on Water Footprints of Wheat and Rice Production. In Sustainability and Water Footprint: Industry-Specific Assessments and Recommendations; Muthu, S.S., Ed.; Springer Nature: Cham, Switzerland, 2024; pp. 57–82. [Google Scholar]
- Muluneh, M.G. Impact of climate change on biodiversity and food security: A global perspective—A review article. Agric. Food Secur. 2021, 10, 36. [Google Scholar] [CrossRef]
- Cavender-Bares, J.; Schneider, F.D.; Santos, M.J.; Armstrong, A.; Carnaval, A.; Dahlin, K.M.; Fatoyinbo, L.; Hurtt, G.C.; Schimel, D.; Townsend, P.A.; et al. Integrating remote sensing with ecology and evolution to advance biodiversity conservation. Nat. Ecol. Evol. 2022, 6, 506–519. [Google Scholar] [CrossRef]
- Wang, K.; Franklin, S.E.; Guo, X.; Cattet, M. Remote Sensing of Ecology, Biodiversity and Conservation: A Review from the Perspective of Remote Sensing Specialists. Sensors 2010, 10, 9647–9667. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Gamon, J.A. Remote sensing of terrestrial plant biodiversity. Remote Sens. Environ. 2019, 231, 111218. [Google Scholar] [CrossRef]
- Haq, M.-A. CNN Based Automated Weed Detection System Using UAV Imagery. Comput. Syst. Sci. Eng. 2022, 42, 837–849. [Google Scholar]
- Haq, M.A.; Rahaman, G.; Baral, P.; Ghosh, A. Deep Learning Based Supervised Image Classification Using UAV Images for Forest Areas Classification. J. Indian Soc. Remote Sens. 2021, 49, 601–606. [Google Scholar] [CrossRef]
- Tian, Y.; Huang, H.; Zhou, G.; Zhang, Q.; Xie, X.; Ou, J.; Zhang, Y.; Tao, J.; Lin, J. Mangrove Biodiversity Assessment Using UAV Lidar and Hyperspectral Data in China’s Pinglu Canal Estuary. Remote Sens. 2023, 15, 2622. [Google Scholar] [CrossRef]
- Zhao, Y.; Zeng, Y.; Zheng, Z.; Dong, W.; Zhao, D.; Wu, B.; Zhao, Q. Forest species diversity mapping using airborne LiDAR and hyperspectral data in a subtropical forest in China. Remote Sens. Environ. 2018, 213, 104–114. [Google Scholar] [CrossRef]
- Pangtey, D.; Padalia, H.; Bodh, R.; Rai, I.D.; Nandy, S. Application of remote sensing-based spectral variability hypothesis to improve tree diversity estimation of seasonal tropical forest considering phenological variations. Geocarto Int. 2023, 38, 2178525. [Google Scholar] [CrossRef]
- Mashiane, K.; Ramoelo, A.; Adelabu, S. Prediction of species richness and diversity in sub-alpine grasslands using satellite remote sensing and random forest machine-learning algorithm. Appl. Veg. Sci. 2024, 27, e12778. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, R.; Lin, C.-F.; Zhang, Z.; Zhang, R.; Shang, K.; Zhao, M.; Huang, J.; Wang, X.; Li, Y.; et al. Remote sensing of subtropical tree diversity: The underappreciated roles of the practical definition of forest canopy and phenological variation. For. Ecosyst. 2023, 10, 100122. [Google Scholar] [CrossRef]
- Ma, S.; Chen, G.; Cai, Q.; Ji, C.; Zhu, B.; Tang, Z.; Hu, S.; Fang, J. Mycorrhizal dominance influences tree species richness and richness–biomass relationship in China’s forests. Ecology 2025, 106, e4501. [Google Scholar] [CrossRef]
- Rocchini, D.; Boyd, D.S.; Féret, J.-B.; Foody, G.M.; He, K.S.; Lausch, A.; Nagendra, H.; Wegmann, M.; Pettorelli, N. Satellite remote sensing to monitor species diversity: Potential and pitfalls. Remote Sens. Ecol. Conserv. 2016, 2, 25–36. [Google Scholar] [CrossRef]
- Kamoske, A.G.; Dahlin, K.M.; Read, Q.D.; Record, S.; Stark, S.C.; Serbin, S.P.; Zarnetske, P.L. Towards mapping biodiversity from above: Can fusing lidar and hyperspectral remote sensing predict taxonomic, functional, and phylogenetic tree diversity in temperate forests? Glob. Ecol. Biogeogr. 2022, 31, 1440–1460. [Google Scholar] [CrossRef]
- Ming, L.; Liu, J.; Quan, Y.; Li, M.; Wang, B.; Wei, G. Mapping tree species diversity in a typical natural secondary forest by combining multispectral and LiDAR data. Ecol. Indic. 2024, 159, 111711. [Google Scholar] [CrossRef]
- Dubayah, R.; Armston, J.; Healey, S.P.; Bruening, J.M.; Patterson, P.L.; Kellner, J.R.; Duncanson, L.; Saarela, S.; Ståhl, G.; Yang, Z.; et al. GEDI launches a new era of biomass inference from space. Environ. Res. Lett. 2022, 17, 095001. [Google Scholar] [CrossRef]
- Ren, C.; Jiang, H.; Xi, Y.; Liu, P.; Li, H. Quantifying Temperate Forest Diversity by Integrating GEDI LiDAR and Multi-Temporal Sentinel-2 Imagery. Remote Sens. 2023, 15, 375. [Google Scholar] [CrossRef]
- Potapov, P.; Li, X.; Hernandez-Serna, A.; Tyukavina, A.; Hansen, M.C.; Kommareddy, A.; Pickens, A.; Turubanova, S.; Tang, H.; Silva, C.E.; et al. Mapping global forest canopy height through integration of GEDI and Landsat data. Remote Sens. Environ. 2021, 253, 112165. [Google Scholar] [CrossRef]
- Fareed, N.; Numata, I.; Cochrane, M.A.; Novoa, S.; Tenneson, K.; Melo, A.W.F.d.; da Silva, S.S.; Oliveira, M.V.N.d.; Nicolau, A.; Zutta, B. Aboveground biomass modeling using simulated Global Ecosystem Dynamics Investigation (GEDI) waveform LiDAR and forest inventories in Amazonian rainforests. For. Ecol. Manag. 2025, 578, 122491. [Google Scholar] [CrossRef]
- Guerra-Hernández, J.; Pascual, A. Using GEDI lidar data and airborne laser scanning to assess height growth dynamics in fast-growing species: A showcase in Spain. For. Ecosyst. 2021, 8, 14. [Google Scholar] [CrossRef]
- Wang, C.; Jia, D.; Lei, S.; Numata, I.; Tian, L. Accuracy Assessment and Impact Factor Analysis of GEDI Leaf Area Index Product in Temperate Forest. Remote Sens. 2023, 15, 1535. [Google Scholar] [CrossRef]
- Huang, J.; Xia, T.; Shuai, Y.; Zhu, H. Assessing the Performance of GEDI LiDAR Data for Estimating Terrain in Densely Forested Areas. IEEE Geosci. Remote Sens. Lett. 2023, 20, 6501505. [Google Scholar] [CrossRef]
- Yang, Z.; Shu, Q.; Zhang, L.; Yang, X. Forest Tree Species Diversity Mapping Using ICESat-2/ATLAS with GF-1/PMS Imagery. Forests 2023, 14, 1537. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, X.; Wang, W.; Wei, H.; Wang, J.; Li, Z.; Li, X.; Wu, H.; Hu, Q. Understanding the potentials of early-season crop type mapping by using Landsat-8, Sentinel-1/2, and GF-1/6 data. Comput. Electron. Agric. 2024, 224, 12. [Google Scholar] [CrossRef]
- Guan, Y.; Tian, X.; Zhang, W.; Marino, A.; Huang, J.; Mao, Y.; Zhao, H. Forest Canopy Cover Inversion Exploration Using Multi-Source Optical Data and Combined Methods. Forests 2023, 14, 1527. [Google Scholar] [CrossRef]
- Liang, Y.; Liang, Y.; Tu, X. Identification and spatial pattern analysis of abandoned farmland in Jiangxi Province of China based on GF-1 satellite image and object-oriented technology. Front. Environ. Sci. 2024, 12, 1423868. [Google Scholar] [CrossRef]
- Bai, J.; Ren, C.; Shi, X.; Xiang, H.; Zhang, W.; Jiang, H.; Ren, Y.; Xi, Y.; Wang, Z.; Mao, D. Tree species diversity impacts on ecosystem services of temperate forests. Ecol. Indic. 2024, 167, 112639. [Google Scholar] [CrossRef]
- Donnini, J.; Kross, A.; Alejo, C. Spectral Diversity as a Predictor of Tree Diversity: Exploring Challenges and Opportunities Across Forest Ecosystems. Can. J. Remote Sens. 2024, 50, 2403495. [Google Scholar] [CrossRef]
- Thanh Noi, P.; Kappas, M. Comparison of Random Forest, k-Nearest Neighbor, and Support Vector Machine Classifiers for Land Cover Classification Using Sentinel-2 Imagery. Sensors 2018, 18, 18. [Google Scholar] [CrossRef]
- Vanguri, R.; Laneve, G.; Hościło, A. Mapping forest tree species and its biodiversity using EnMAP hyperspectral data along with Sentinel-2 temporal data: An approach of tree species classification and diversity indices. Ecol. Indic. 2024, 167, 112671. [Google Scholar] [CrossRef]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Cortes, C.; Vapnik, V. Support-vector networks. Mach. Learn. 1995, 20, 273–297. [Google Scholar] [CrossRef]
- Duda, R.O.; Hart, P.E. Pattern Classification and Scene Analysis; Wiley: New York, NY, USA, 1973. [Google Scholar]
- Belgiu, M.; Drăguţ, L. Random forest in remote sensing: A review of applications and future directions. ISPRS J. Photogramm. Remote Sens. 2016, 114, 24–31. [Google Scholar] [CrossRef]
- Cervantes, J.; Garcia-Lamont, F.; Rodríguez-Mazahua, L.; Lopez, A. A comprehensive survey on support vector machine classification: Applications, challenges and trends. Neurocomputing 2020, 408, 189–215. [Google Scholar] [CrossRef]
- Sheykhmousa, M.; Mahdianpari, M.; Ghanbari, H.; Mohammadimanesh, F.; Ghamisi, P.; Homayouni, S. Support Vector Machine Versus Random Forest for Remote Sensing Image Classification: A Meta-Analysis and Systematic Review. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2020, 13, 6308–6325. [Google Scholar] [CrossRef]
- Zhang, S.; Li, X.; Zong, M.; Zhu, X.; Wang, R. Efficient kNN Classification With Different Numbers of Nearest Neighbors. IEEE Trans. Neural Netw. Learn. Syst. 2018, 29, 1774–1785. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Fu, T.; Gao, H.; Li, M.; Liu, J. Climatic and Non-Climatic Drivers of Plant Diversity along an Altitudinal Gradient in the Taihang Mountains of Northern China. Diversity 2023, 15, 66. [Google Scholar] [CrossRef]
- Zhao, H.; Wang, Q.-R.; Fan, W.; Song, G.-H. The Relationship between Secondary Forest and Environmental Factors in the Southern Taihang Mountains. Sci. Rep. 2017, 7, 16431. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.-l.; Xu, Q.-h.; Zhao, H.-p.; Liang, W.-d.; Sun, L.-m. Vegetation changes of the Taihang Mountains since the last glacial. Chin. Geogr. Sci. 2000, 10, 261–269. [Google Scholar] [CrossRef]
- Wang, F.; Liu, J.; Deng, W.; Fu, T.; Gao, H.; Qi, F. Assessing impacts of ecological restoration project on water retention function in the Taihang Mountain area, China. Ecohydrology 2024, 17, e2638. [Google Scholar] [CrossRef]
- Lu, J.; Hou, J.; Wang, H.; Qu, W. Current Status of Macaca mulatta in Taihangshan Mountains Area, Jiyuan, Henan, China. Int. J. Primatol. 2007, 28, 1085–1091. [Google Scholar] [CrossRef]
- Luo, Y.; Zhou, M.; Jin, S.; Wang, Q.; Yan, D. Changes in phylogenetic structure and species composition of woody plant communities across an elevational gradient in the southern Taihang Mountains, China. Glob. Ecol. Conserv. 2023, 42, e02412. [Google Scholar] [CrossRef]
- Feng, E.; Zhang, L.; Kong, Y.; Xu, X.; Wang, T.; Wang, C. Distribution Characteristics of Active Soil Substances along Elevation Gradients in the Southern of Taihang Mountain, China. Forests 2023, 14, 370. [Google Scholar] [CrossRef]
- Jin, S.-S.; Zhang, Y.-Y.; Zhou, M.-L.; Dong, X.-M.; Chang, C.-H.; Wang, T.; Yan, D.-F. Interspecific Association and Community Stability of Tree Species in Natural Secondary Forests at Different Altitude Gradients in the Southern Taihang Mountains. Forests 2022, 13, 373. [Google Scholar] [CrossRef]
- Cui, Z.; Shao, Q.; Grueter, C.C.; Wang, Z.; Lu, J.; Raubenheimer, D. Dietary diversity of an ecological and macronutritional generalist primate in a harsh high-latitude habitat, the Taihangshan macaque (Macaca mulatta tcheliensis). Am. J. Primatol. 2019, 81, e22965. [Google Scholar] [CrossRef]
- Condit, R. Research in large, long-term tropical forest plots. Trends Ecol. Evol. 1995, 10, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Li, X.; Zhang, Z.; Yang, J.; Zhao, Y.; Yang, Z.; Hu, Q. Effects of natural vegetative restoration on soil fungal and bacterial communities in bare patches of the southern Taihang Mountains. Ecol. Evol. 2019, 9, 10432–10441. [Google Scholar] [CrossRef] [PubMed]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Simpson, E.H. Measurement of Diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Pielou, E.C. The measurement of diversity in different types of biological collections. J. Theor. Biol. 1966, 13, 131–144. [Google Scholar] [CrossRef]
- Morris, E.K.; Caruso, T.; Buscot, F.; Fischer, M.; Hancock, C.; Maier, T.S.; Meiners, T.; Müller, C.; Obermaier, E.; Prati, D.; et al. Choosing and using diversity indices: Insights for ecological applications from the German Biodiversity Exploratories. Ecol. Evol. 2014, 4, 3514–3524. [Google Scholar] [CrossRef]
- East, A.; Hansen, A.; Jantz, P.; Currey, B.; Roberts, D.W.; Armenteras, D. Validation and Error Minimization of Global Ecosystem Dynamics Investigation (GEDI) Relative Height Metrics in the Amazon. Remote Sens. 2024, 16, 3550. [Google Scholar] [CrossRef]
- Duncanson, L.; Kellner, J.R.; Armston, J.; Dubayah, R.; Minor, D.M.; Hancock, S.; Healey, S.P.; Patterson, P.L.; Saarela, S.; Marselis, S.; et al. Aboveground biomass density models for NASA’s Global Ecosystem Dynamics Investigation (GEDI) lidar mission. Remote Sens. Environ. 2022, 270, 112845. [Google Scholar] [CrossRef]
- Torresani, M.; Rocchini, D.; Sonnenschein, R.; Zebisch, M.; Hauffe, H.C.; Heym, M.; Pretzsch, H.; Tonon, G. Height variation hypothesis: A new approach for estimating forest species diversity with CHM LiDAR data. Ecol. Indic. 2020, 117, 106520. [Google Scholar] [CrossRef]
- Brown, L.A.; Morris, H.; Meier, C.; Knohl, A.; Lanconelli, C.; Gobron, N.; Dash, J.; Danson, F.M. Stage 1 Validation of Plant Area Index From the Global Ecosystem Dynamics Investigation. IEEE Geosci. Remote Sens. Lett. 2023, 20, 2505005. [Google Scholar] [CrossRef]
- Wang, C.; Elmore, A.J.; Numata, I.; Cochrane, M.A.; Shaogang, L.; Huang, J.; Zhao, Y.; Li, Y. Factors affecting relative height and ground elevation estimations of GEDI among forest types across the conterminous USA. GIsci. Remote Sens. 2022, 59, 975–999. [Google Scholar] [CrossRef]
- Diaz-Kloch, N.; Murray, D.L. Harmonizing GEDI and LVIS Data for Accurate and Large-Scale Mapping of Foliage Height Diversity. Can. J. Remote Sens. 2024, 50, 2341762. [Google Scholar] [CrossRef]
- Li, X.; Li, L.; Ni, W.; Mu, X.; Wu, X.; Vaglio Laurin, G.; Vangi, E.; Stereńczak, K.; Chirici, G.; Yu, S.; et al. Validating GEDI tree canopy cover product across forest types using co-registered aerial LiDAR data. ISPRS J. Photogramm. Remote Sens. 2024, 207, 326–337. [Google Scholar] [CrossRef]
- Bruening, J.M.; Fischer, R.; Bohn, F.J.; Armston, J.; Armstrong, A.H.; Knapp, N.; Tang, H.; Huth, A.; Dubayah, R. Challenges to aboveground biomass prediction from waveform lidar. Environ. Res. Lett. 2021, 16, 125013. [Google Scholar] [CrossRef]
- Liu, L.; Pang, Y.; Ren, H.; Li, Z. Predict Tree Species Diversity from GF-2 Satellite Data in a Subtropical Forest of China. Sci. Silvae Sin. 2019, 55, 61–74. [Google Scholar]
- Rouse, J.W.; Haas, R.H.; Schell, J.A.; Deering, D.W. Monitoring vegetation systems in the great plains with ERTS. In Third ERTS Symposium; NASA: Washington, DC, USA, 1973; Volume 1, pp. 309–317. [Google Scholar]
- Gitelson, A.A.; Zur, Y.; Chivkunova, O.B.; Merzlyak, M.N. Assessing Carotenoid Content in Plant Leaves with Reflectance Spectroscopy. Photochem. Photobiol. 2002, 75, 272–281. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Tucker, C.J. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Liu, H.Q.; Huete, A. A feedback based modification of the NDVI to minimize canopy background and atmospheric noise. IEEE Trans. Geosci. Remote Sens. 1995, 33, 457–465. [Google Scholar] [CrossRef]
- Haboudane, D.; Miller, J.R.; Pattey, E.; Zarco-Tejada, P.J.; Strachan, I.B. Hyperspectral vegetation indices and novel algorithms for predicting green LAI of crop canopies: Modeling and validation in the context of precision agriculture. Remote Sens. Environ. 2004, 90, 337–352. [Google Scholar] [CrossRef]
- Huete, A.R. A soil-adjusted vegetation index (SAVI). Remote Sens. Environ. 1988, 25, 295–309. [Google Scholar] [CrossRef]
- Haboudane, D.; Miller, J.R.; Tremblay, N.; Zarco-Tejada, P.J.; Dextraze, L. Integrated narrow-band vegetation indices for prediction of crop chlorophyll content for application to precision agriculture. Remote Sens. Environ. 2002, 81, 416–426. [Google Scholar] [CrossRef]
- Elvidge, C.D.; Chen, Z. Comparison of broad-band and narrow-band red and near-infrared vegetation indices. Remote Sens. Environ. 1995, 54, 38–48. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Kaufman, Y.J.; Merzlyak, M.N. Use of a green channel in remote sensing of global vegetation from EOS-MODIS. Remote Sens. Environ. 1996, 58, 289–298. [Google Scholar] [CrossRef]
- Gitelson, A.A. Wide Dynamic Range Vegetation Index for Remote Quantification of Biophysical Characteristics of Vegetation. J. Plant Physiol. 2004, 161, 165–173. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Peng, Y.; Masek, J.G.; Rundquist, D.C.; Verma, S.; Suyker, A.; Baker, J.M.; Hatfield, J.L.; Meyers, T. Remote estimation of crop gross primary production with Landsat data. Remote Sens. Environ. 2012, 121, 404–414. [Google Scholar] [CrossRef]
- Pinty, B.; Verstraete, M.M. GEMI: A non-linear index to monitor global vegetation from satellites. Vegetatio 1992, 101, 15–20. [Google Scholar] [CrossRef]
- Sripada, R.P.; Heiniger, R.W.; White, J.G.; Meijer, A.D. Aerial Color Infrared Photography for Determining Early In-Season Nitrogen Requirements in Corn. Agron. J. 2006, 98, 968–977. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Merzlyak, M.N. Remote sensing of chlorophyll concentration in higher plant leaves. Adv. Space Res. 1998, 22, 689–692. [Google Scholar] [CrossRef]
- Gamon, J.A.; Surfus, J.S. Assessing leaf pigment content and activity with a reflectometer. New Phytol. 1999, 143, 105–117. [Google Scholar] [CrossRef]
- Guyon, I.; Weston, J.; Barnhill, S.; Vapnik, V. Gene Selection for Cancer Classification using Support Vector Machines. Mach. Learn. 2002, 46, 389–422. [Google Scholar] [CrossRef]
- Guo, P.-T.; Li, M.-F.; Luo, W.; Tang, Q.-F.; Liu, Z.-W.; Lin, Z.-M. Digital mapping of soil organic matter for rubber plantation at regional scale: An application of random forest plus residuals kriging approach. Geoderma 2015, 237–238, 49–59. [Google Scholar] [CrossRef]
- Silveira, E.M.O.; Espírito Santo, F.D.; Wulder, M.A.; Acerbi Júnior, F.W.; Carvalho, M.C.; Mello, C.R.; Mello, J.M.; Shimabukuro, Y.E.; Terra, M.C.N.S.; Carvalho, L.M.T.; et al. Pre-stratified modelling plus residuals kriging reduces the uncertainty of aboveground biomass estimation and spatial distribution in heterogeneous savannas and forest environments. For. Ecol. Manag. 2019, 445, 96–109. [Google Scholar] [CrossRef]
- Liu, X.; Su, Y.; Hu, T.; Yang, Q.; Liu, B.; Deng, Y.; Tang, H.; Tang, Z.; Fang, J.; Guo, Q. Neural network guided interpolation for mapping canopy height of China’s forests by integrating GEDI and ICESat-2 data. Remote Sens. Environ. 2022, 269, 112844. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Su, Y.; Guo, Q.; Xue, B.; Hu, T.; Alvarez, O.; Tao, S.; Fang, J. Spatial distribution of forest aboveground biomass in China: Estimation through combination of spaceborne lidar, optical imagery, and forest inventory data. Remote Sens. Environ. 2016, 173, 187–199. [Google Scholar] [CrossRef]
- Palmer, M.W.; Earls, P.G.; Hoagland, B.W.; White, P.S.; Wohlgemuth, T. Quantitative tools for perfecting species lists. Environmetrics 2002, 13, 121–137. [Google Scholar] [CrossRef]
- Sims, D.A.; Gamon, J.A. Relationships between leaf pigment content and spectral reflectance across a wide range of species, leaf structures and developmental stages. Remote Sens. Environ. 2002, 81, 337–354. [Google Scholar] [CrossRef]
- Cazzolla Gatti, R.; Di Paola, A.; Bombelli, A.; Noce, S.; Valentini, R. Exploring the relationship between canopy height and terrestrial plant diversity. Plant Ecolog. 2017, 218, 899–908. [Google Scholar] [CrossRef]
- Marselis, S.M.; Tang, H.; Armston, J.; Abernethy, K.; Alonso, A.; Barbier, N.; Bissiengou, P.; Jeffery, K.; Kenfack, D.; Labrière, N.; et al. Exploring the relation between remotely sensed vertical canopy structure and tree species diversity in Gabon. Environ. Res. Lett. 2019, 14, 094013. [Google Scholar] [CrossRef]
- Torresani, M.; Rocchini, D.; Alberti, A.; Moudrý, V.; Heym, M.; Thouverai, E.; Kacic, P.; Tomelleri, E. LiDAR GEDI derived tree canopy height heterogeneity reveals patterns of biodiversity in forest ecosystems. Ecol. Inf. 2023, 76, 102082. [Google Scholar] [CrossRef]
- Sun, G.; Ranson, K.J.; Guo, Z.; Zhang, Z.; Montesano, P.; Kimes, D. Forest biomass mapping from lidar and radar synergies. Remote Sens. Environ. 2011, 115, 2906–2916. [Google Scholar] [CrossRef]
- Ni-Meister, W.; Lee, S.; Strahler, A.H.; Woodcock, C.E.; Schaaf, C.; Yao, T.; Ranson, K.J.; Sun, G.; Blair, J.B. Assessing general relationships between aboveground biomass and vegetation structure parameters for improved carbon estimate from lidar remote sensing. J. Geophys. Res. Biogeosci. 2010, 115, G00E11. [Google Scholar] [CrossRef]
- Valizadeh, E.; Asadi, H.; Jaafari, A.; Tafazoli, M. Machine learning prediction of tree species diversity using forest structure and environmental factors: A case study from the Hyrcanian forest, Iran. Environ. Monit. Assess. 2023, 195, 1334. [Google Scholar] [CrossRef]
- Zagajewski, B.; Kluczek, M.; Raczko, E.; Njegovec, A.; Dabija, A.; Kycko, M. Comparison of Random Forest, Support Vector Machines, and Neural Networks for Post-Disaster Forest Species Mapping of the Krkonoše/Karkonosze Transboundary Biosphere Reserve. Remote Sens. 2021, 13, 2581. [Google Scholar] [CrossRef]
- Qu, F.; Sun, Y.; Zhou, M.; Liu, L.; Yang, H.; Zhang, J.; Huang, H.; Hong, D. Vegetation Land Segmentation with Multi-Modal and Multi-Temporal Remote Sensing Images: A Temporal Learning Approach and a New Dataset. Remote Sens. 2024, 16, 3. [Google Scholar] [CrossRef]
- Lang, N.; Kalischek, N.; Armston, J.; Schindler, K.; Dubayah, R.; Wegner, J.D. Global canopy height regression and uncertainty estimation from GEDI LIDAR waveforms with deep ensembles. Remote Sens. Environ. 2022, 268, 112760. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scientific Name | Leaf Type | DBH | Tree Height | ||||
|---|---|---|---|---|---|---|---|
| Min | Max | Mean | Min | Max | Mean | ||
| Quercus variabilis | broad | 5 | 74 | 15.75 | 2 | 33 | 10.6 |
| Quercus aliena | broad | 5 | 84 | 19.85 | 2 | 33 | 10.1 |
| Pinus tabuliformis | needle | 5 | 66 | 16.29 | 2 | 35.4 | 6.7 |
| Carpinus turczaninovii | broad | 5 | 79 | 8.90 | 2 | 26 | 5.8 |
| Quercus baronii | broad | 5 | 33 | 8.89 | 2.5 | 16 | 7.7 |
| Quercus aliena var. acuteserrata | broad | 5 | 31.4 | 12.16 | 2.2 | 15.5 | 8.5 |
| Ulmus pumila | broad | 5.2 | 79.6 | 15.16 | 3.5 | 23 | 12.1 |
| Rhus potaninii | broad | 5 | 26.7 | 12.13 | 2.1 | 17.5 | 6.2 |
| Cotinus coggygria var. cinereus | broad | 5 | 45 | 13.45 | 2.5 | 15.6 | 8.8 |
| Castanea mollissima | broad | 5 | 28 | 7.80 | 2.2 | 10.9 | 4.8 |
| Malus spectabilis | broad | 5.1 | 26.7 | 10.61 | 2.3 | 21.2 | 9.2 |
| Carya cathayensis | broad | 5 | 36 | 13.75 | 3 | 16 | 9.1 |
| Robinia pseudoacacia | broad | 5 | 25.2 | 10.40 | 2.4 | 9 | 5.2 |
| Acer davidii | broad | 5 | 37.6 | 12.85 | 2.5 | 27.2 | 8.7 |
| Toxicodendron vernicifluum | broad | 5 | 36.8 | 11.55 | 2 | 14.9 | 6.7 |
| Pistacia chinensis | broad | 5.2 | 41 | 18.12 | 3.7 | 20 | 11.8 |
| Tilia chinensis | broad | 5.1 | 28.3 | 10.66 | 2.1 | 18 | 7.4 |
| Populus × canadensis | broad | 5.1 | 24.5 | 10.41 | 3.1 | 10.2 | 6.1 |
| Acer pictum subsp. mono | broad | 5 | 31.5 | 11.64 | 2.5 | 15.3 | 7.9 |
| Diospyros lotus | broad | 5 | 23.3 | 10.68 | 2 | 22.5 | 8.8 |
| Swida macrophylla | broad | 5 | 23.3 | 10.68 | 2 | 22.5 | 8.8 |
| Deciduous Broadleaf Forest | Evergreen Broadleaf Forest | Needleleaf-Broadleaf Mixed Forest | Needleleaf Forest | Waters | Building Land | Plow Land | |
|---|---|---|---|---|---|---|---|
| Number of sample plots | 165 | 44 | 30 | 25 | 18 | 13 | 5 |
| Diversity Index | Min | Max | Mean | Median | Standard Deviation | Coefficient of Variation | Kurtosis | Skewness |
|---|---|---|---|---|---|---|---|---|
| H | 0.00 | 2.40 | 1.01 | 1.06 | 0.64 | 63.22 | −1.07 | −0.12 |
| D | 0.00 | 0.89 | 0.47 | 0.54 | 0.28 | 58.16 | −1.06 | −0.51 |
| J | 0.00 | 0.99 | 0.55 | 0.64 | 0.27 | 49.78 | −0.31 | −0.92 |
| S | 0.00 | 16.00 | 6.01 | 6.00 | 4.07 | 67.71 | −0.65 | 0.35 |
| Data | Variables | Description |
|---|---|---|
| L2A | RH10, RH20, RH30, RH40, RH50, RH60, RH70, RH80, RH90, RH98 | Based on a 10% increment in the relative height index, RH100 exhibited significant noise and was replaced by RH98 |
| L2B | PAI | Plant area index |
| FHD | Foliage height diversity index | |
| TCC | Total canopy cover | |
| L4A | AGBD | Aboveground biomass density (Mg/ha) |
| Parameter | Condition | Description |
|---|---|---|
| quality_flag | 1 | Indicates that the waveform meets specific criteria based on energy, sensitivity, amplitude, and real-time surface tracking quality and can be expressed as a valid waveform. |
| rx_assess_flag | 0 | Flags indicating various error conditions possible in rxwaveform. |
| degrade_flag | 0 | The state degradation sign is ‘1’, indicating that the state of the indicated direction or positioning information decreases, which affects the accuracy of the data. |
| sensitivity | ≥0.90 | Considering the signal-to-noise ratio of the waveform, the maximum canopy coverage that can be penetrated is indicated. |
| ǀelev_lowestmode—SRTMǀ | ≤50 | Because GEDI is susceptible to clouds in data acquisition, the removal of elev_lowestmode and the SRTM of GEDI footprints have increased differences in spots. |
| Type | Waveband | Wavelength Coverage | Spatial Resolution |
|---|---|---|---|
| Panchromatic | PAN | 0.45–0.90 | 2 m |
| Multispectral | Blue | 0.45–0.52 | 8 m |
| Green | 0.52–0.59 | 8 m | |
| Red | 0.63–0.69 | 8 m | |
| NIR | 0.77–0.89 | 8 m |
| Vegetation Indices | Expression | References |
|---|---|---|
| Normalized difference vegetation index (NDVI) | [71] | |
| Carotenoid reflectance index (CRI) | [72] | |
| Enhanced vegetation index (EVI) | [73] | |
| Differential vegetation index (DVI) | [74] | |
| Nonlinear vegetation index (NIL) | [71] | |
| Modified normalized vegetation index (mNDVI) | [75] | |
| Renormalized vegetation index (RDVI) | [76] | |
| Soil-adjusted vegetation index (SAVI) | [77] | |
| Optimized soil-adjusted vegetation index (OSAVI) | [78] | |
| Ratio vegetation index (RVI) | [79] | |
| Green chlorophyll vegetation index (GCVI) | [80] | |
| Wide dynamic range vegetation index (WDRVI) | [81] | |
| Green wide dynamic range vegetation index (GWDRVI) | [82] | |
| Global environmental monitoring index (GEMI) | [83] | |
| Green difference vegetation index (GDVI) | [84] | |
| Green normalized difference vegetation index (GNDVI) | [85] | |
| Green ratio vegetation index (GRVI) | [84] | |
| Red green ratio index (RGRI) | [86] |
| GLMC | |
|---|---|
| Model | Parameters |
|---|---|
| RF | n_estimators = 50, 100, 150, 200, 300, 500, 1000 |
| max_feature = sqrt, log2, none | |
| max_depth = 1, 3, 5, 10, 20, 50, 100, 200, none | |
| SVM | C = 0.1, 0.5, 1, 3, 5, 10 |
| epsilon = 0.01, 0.1, 1 | |
| kernel = linear, poly, rbf, sigmoid, precomputed | |
| kNN | N_neighbors = 1, 2, 3, 5, 7, 9, 11, 15, 20, 25, 30 |
| Features | Data Source | Resolution | Variables |
|---|---|---|---|
| Terrain features | SRTM DEM | 30 m | DEM, slope, and aspect |
| Climate features | WorldClim version 2.1 | 1 km | Annual mean temperature (tm), temperature seasonality (ts), annual mean precipitation (pm), and precipitation seasonality (ps) |
| NDVI | Sentinel-2 | 10 m | NDVI maximum value composite for 2020 |
| Variable | R2 | RMSE | MAE |
|---|---|---|---|
| RH10 | 0.351 | 2.181 | 1.556 |
| RH20 | 0.396 | 2.685 | 1.971 |
| RH30 | 0.429 | 3.039 | 2.235 |
| RH40 | 0.460 | 3.285 | 2.433 |
| RH50 | 0.481 | 3.526 | 2.609 |
| RH60 | 0.481 | 3.749 | 2.801 |
| RH70 | 0.476 | 3.989 | 2.961 |
| RH80 | 0.499 | 4.122 | 3.006 |
| RH90 | 0.519 | 4.375 | 3.158 |
| RH98 | 0.503 | 4.928 | 3.582 |
| PAI | 0.407 | 1.117 | 0.866 |
| FHD | 0.547 | 0.224 | 0.163 |
| TCC | 0.451 | 0.180 | 0.136 |
| AGBD | 0.469 | 53.307 | 37.647 |
| Combined Variable | H | D | J | S | Average | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| R2 | RMSE | R2 | RMSE | R2 | RMSE | R2 | RMSE | R2 | RMSE | ||
| RF | GF-1 | 0.555 | 0.412 | 0.553 | 0.175 | 0.633 | 0.160 | 0.472 | 2.911 | 0.553 | 0.915 |
| GEDI | 0.616 | 0.380 | 0.597 | 0.170 | 0.650 | 0.156 | 0.679 | 2.309 | 0.636 | 0.754 | |
| GF-1 and GEDI | 0.662 | 0.382 | 0.679 | 0.157 | 0.650 | 0.155 | 0.708 | 2.304 | 0.675 | 0.750 | |
| SVM | GF-1 | 0.342 | 0.501 | 0.393 | 0.204 | 0.505 | 0.196 | 0.323 | 3.296 | 0.391 | 1.049 |
| GEDI | 0.573 | 0.401 | 0.547 | 0.192 | 0.386 | 0.420 | 0.506 | 2.741 | 0.503 | 0.939 | |
| GF-1 and GEDI | 0.446 | 0.456 | 0.578 | 0.180 | 0.518 | 0.195 | 0.623 | 2.618 | 0.541 | 0.862 | |
| kNN | GF-1 | 0.389 | 0.483 | 0.377 | 0.206 | 0.555 | 0.177 | 0.427 | 3.031 | 0.437 | 0.974 |
| GEDI | 0.620 | 0.378 | 0.522 | 0.185 | 0.513 | 0.184 | 0.636 | 2.352 | 0.573 | 0.775 | |
| GF-1 and GEDI | 0.551 | 0.410 | 0.608 | 0.173 | 0.594 | 0.176 | 0.625 | 2.613 | 0.595 | 0.843 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Yang, L.; Sun, J.; Zhu, Q.; Wang, T.; Zhao, H. Estimation of Tree Species Diversity in Warm Temperate Forests via GEDI and GF-1 Imagery. Forests 2025, 16, 570. https://doi.org/10.3390/f16040570
Zhang L, Yang L, Sun J, Zhu Q, Wang T, Zhao H. Estimation of Tree Species Diversity in Warm Temperate Forests via GEDI and GF-1 Imagery. Forests. 2025; 16(4):570. https://doi.org/10.3390/f16040570
Chicago/Turabian StyleZhang, Lei, Liu Yang, Jinhua Sun, Qimeng Zhu, Ting Wang, and Hui Zhao. 2025. "Estimation of Tree Species Diversity in Warm Temperate Forests via GEDI and GF-1 Imagery" Forests 16, no. 4: 570. https://doi.org/10.3390/f16040570
APA StyleZhang, L., Yang, L., Sun, J., Zhu, Q., Wang, T., & Zhao, H. (2025). Estimation of Tree Species Diversity in Warm Temperate Forests via GEDI and GF-1 Imagery. Forests, 16(4), 570. https://doi.org/10.3390/f16040570