
The Pivotal Role of Microscopy in Unravelling the Nature of Microbial Deterioration of Waterlogged Wood: A Review
Abstract
1. Introduction
2. Composition and Structure of Wood
2.1. Composition
2.2. Structure
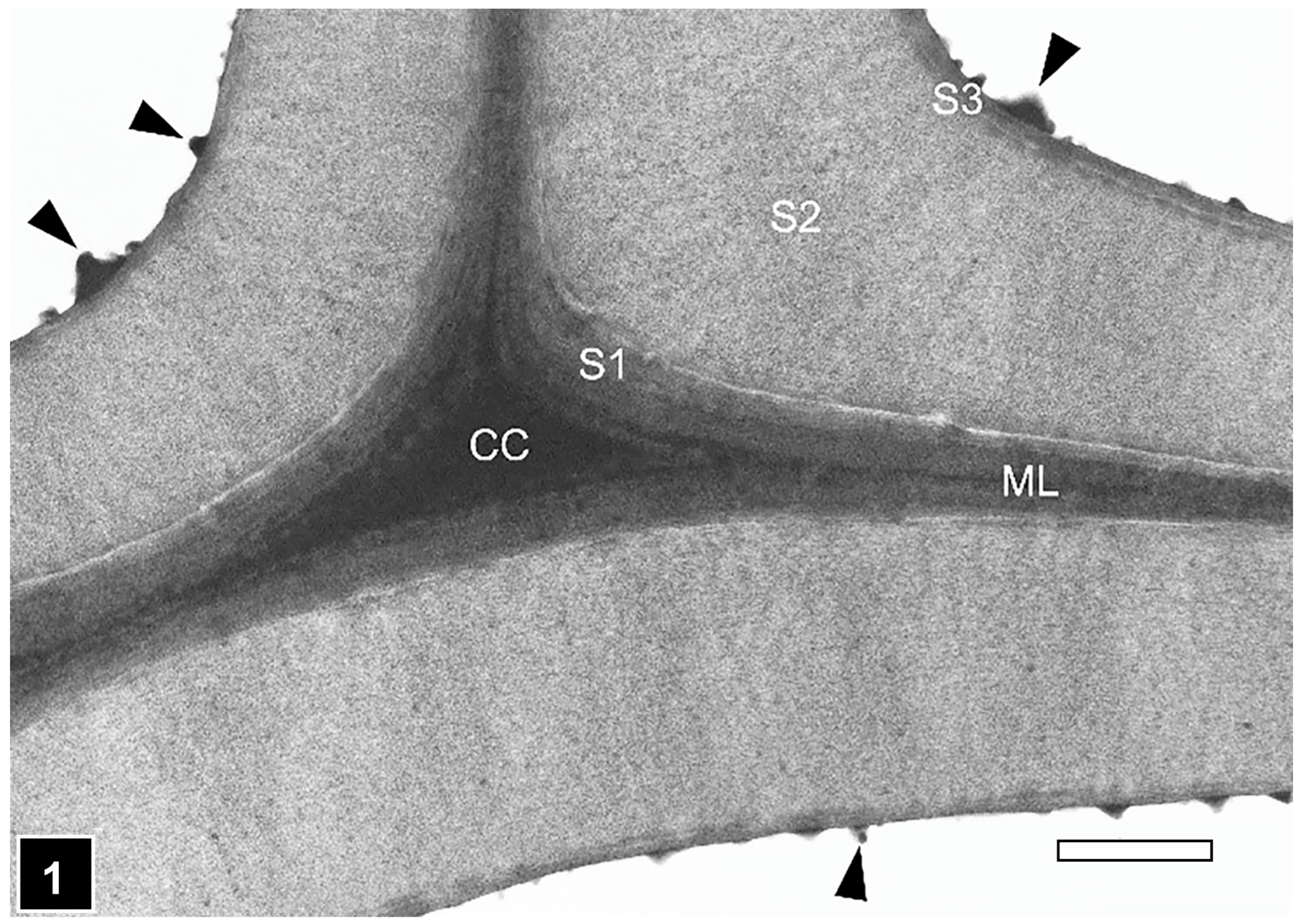
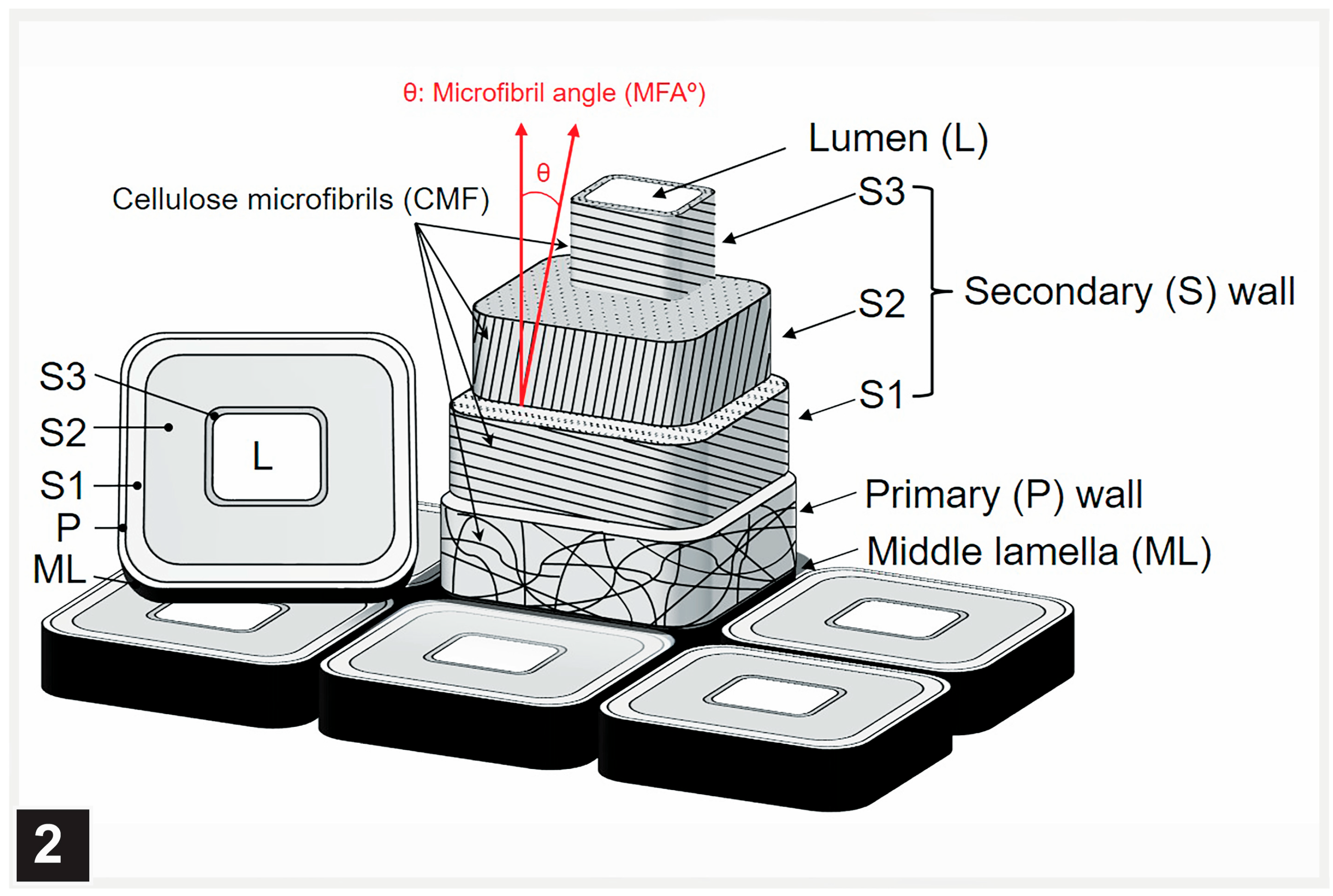
2.3. Cell Wall Ultrastructure
2.3.1. The Typical Three-Layered Secondary Wall
2.3.2. Multilamellar Cell Walls
2.3.3. Compression Wood Cell Walls
2.3.4. Tension Wood Cell Walls
3. Microscopic Techniques
4. Decay Types
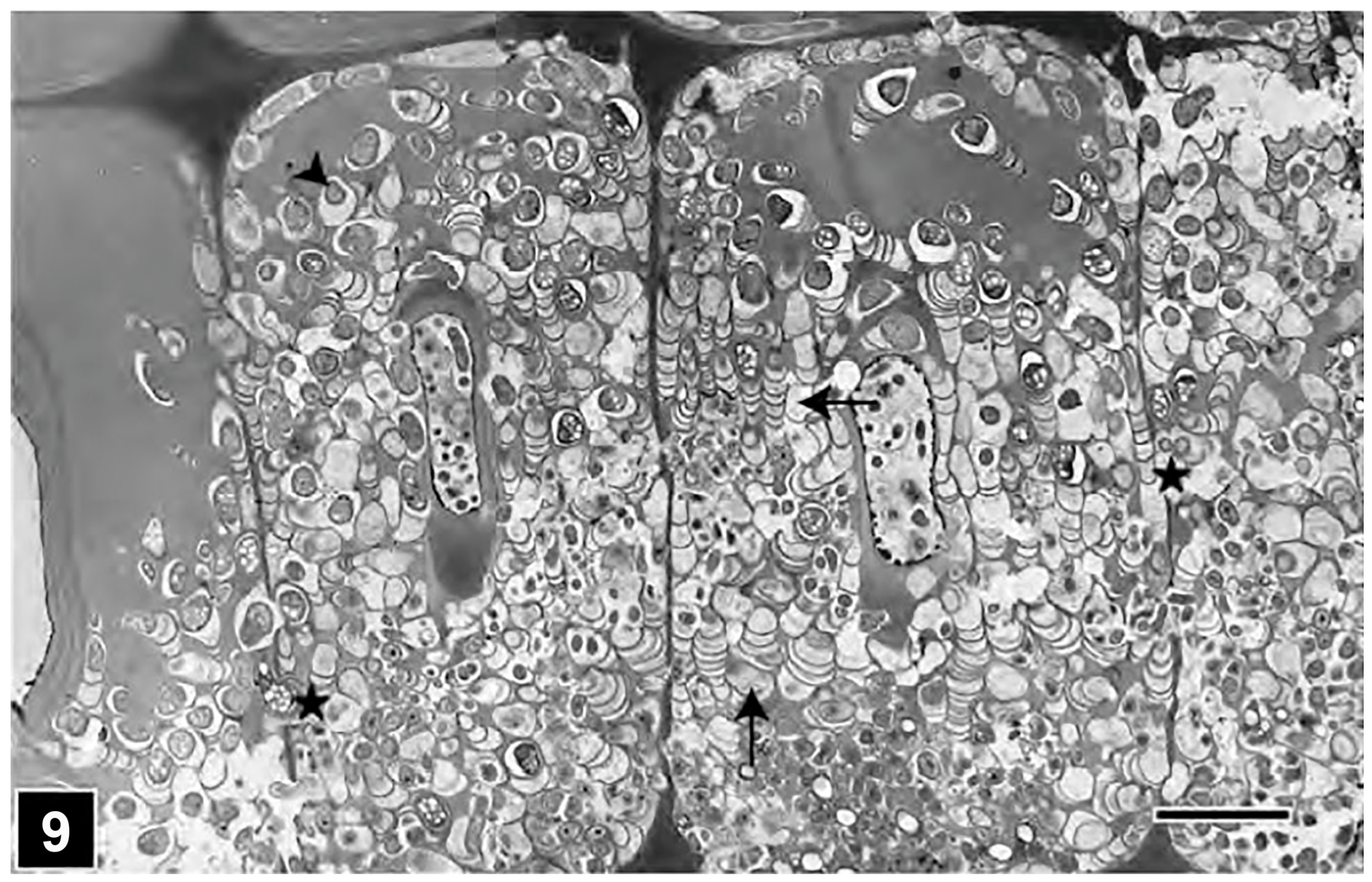
4.1. Fungal Decay
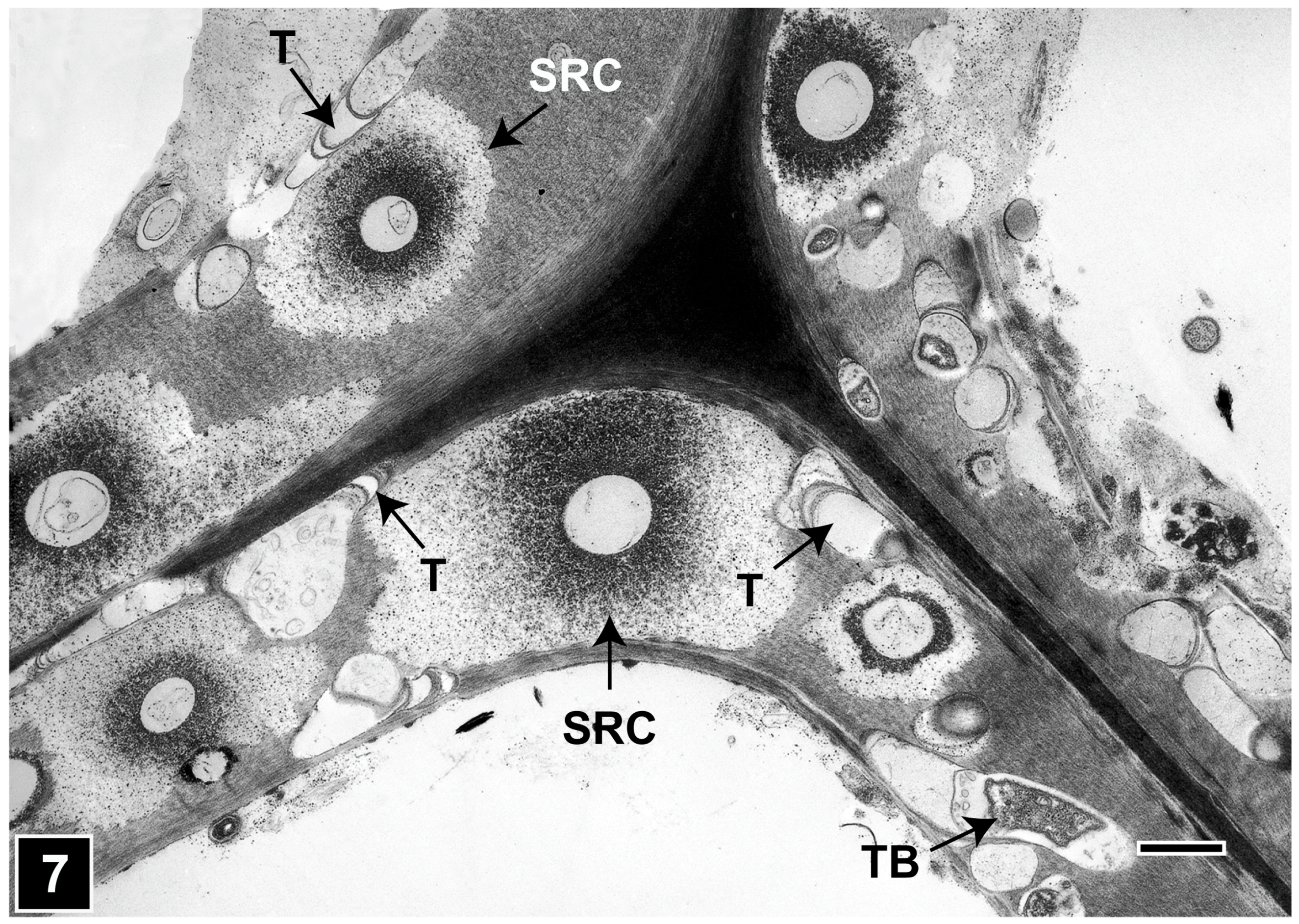
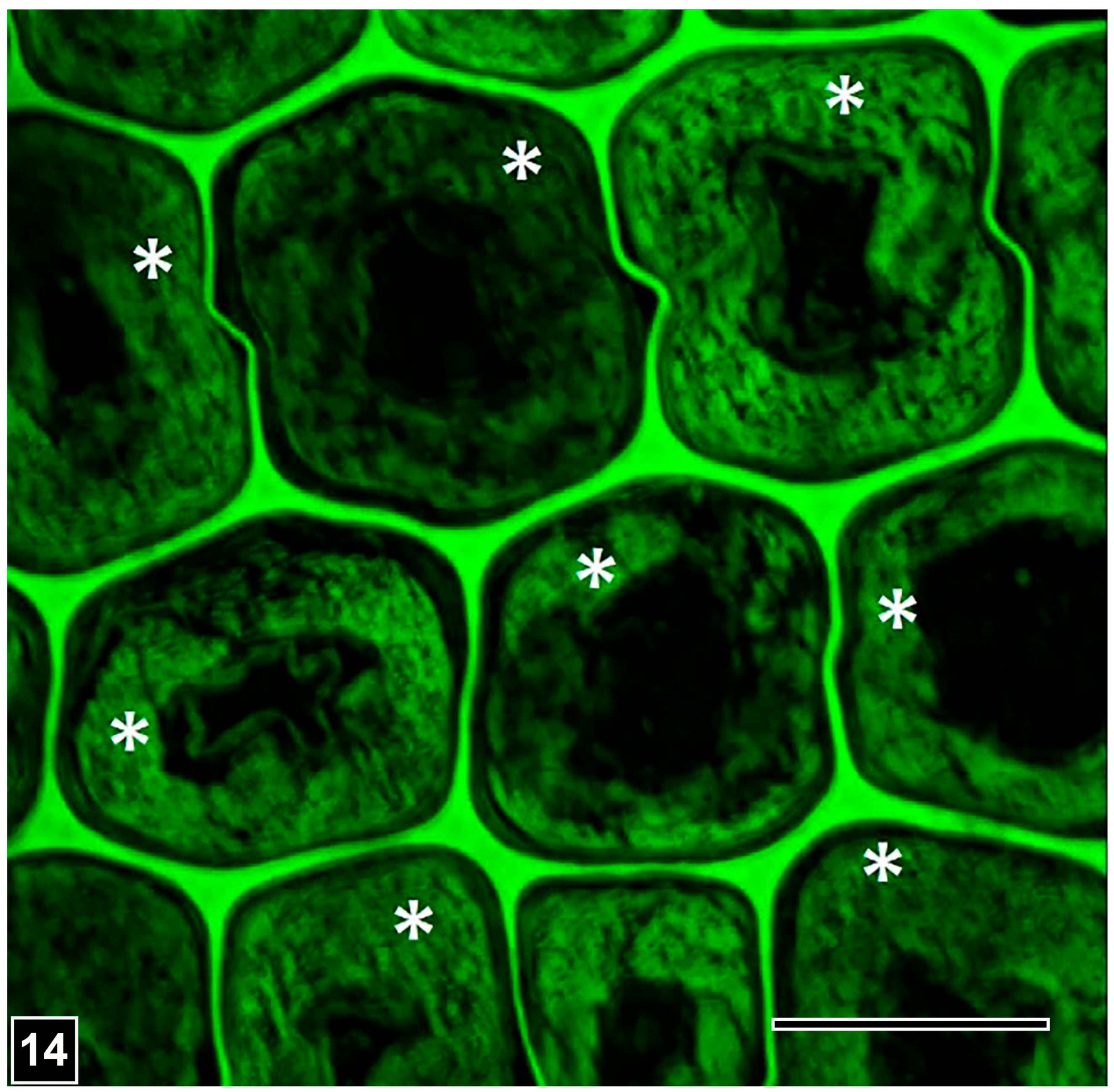
4.1.1. Soft Rot Type I
4.1.2. Soft Rot Type II
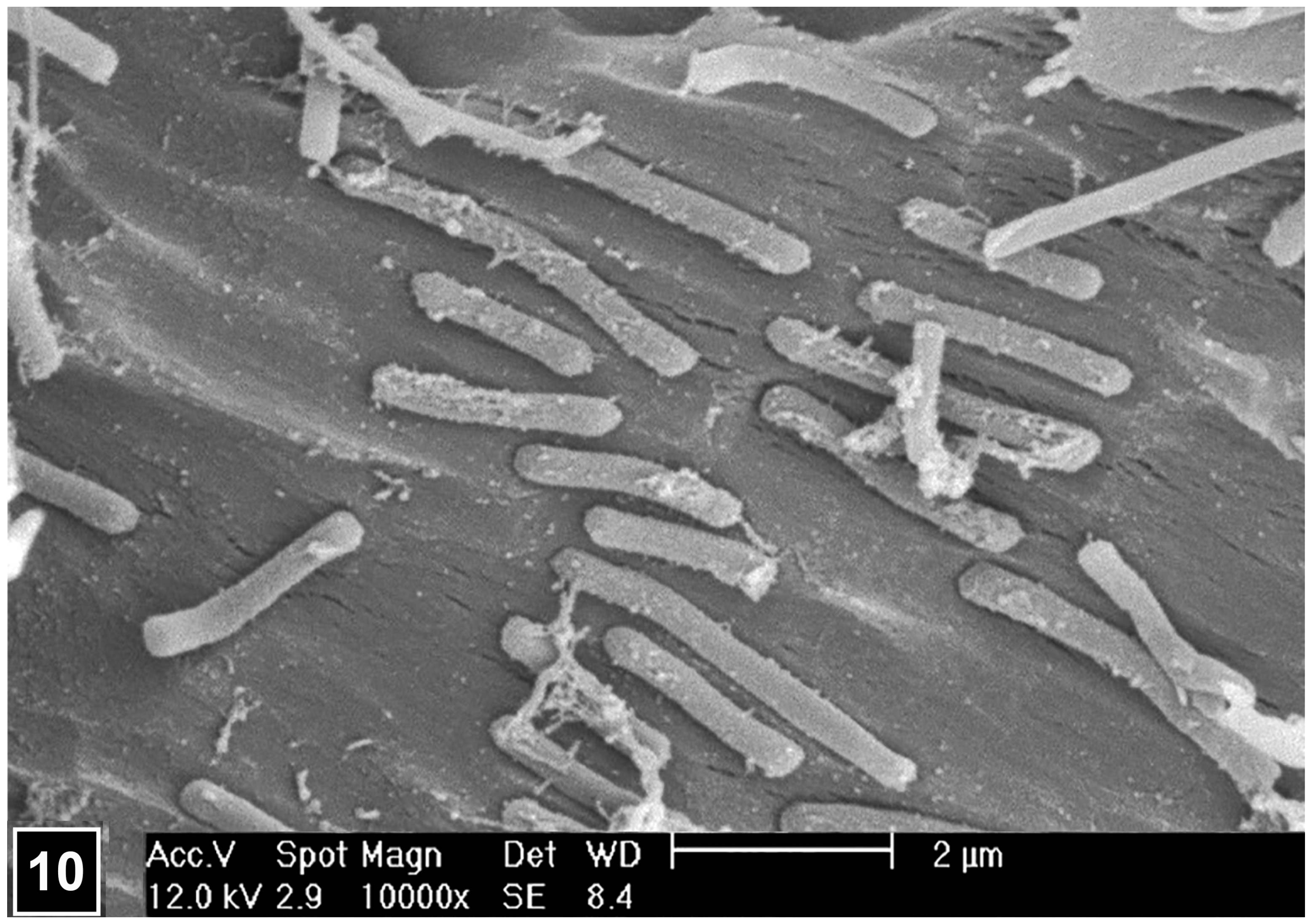
4.2. Microscopic Journey Leading to the Unravelling of Distinctive Bacterial Decay Patterns
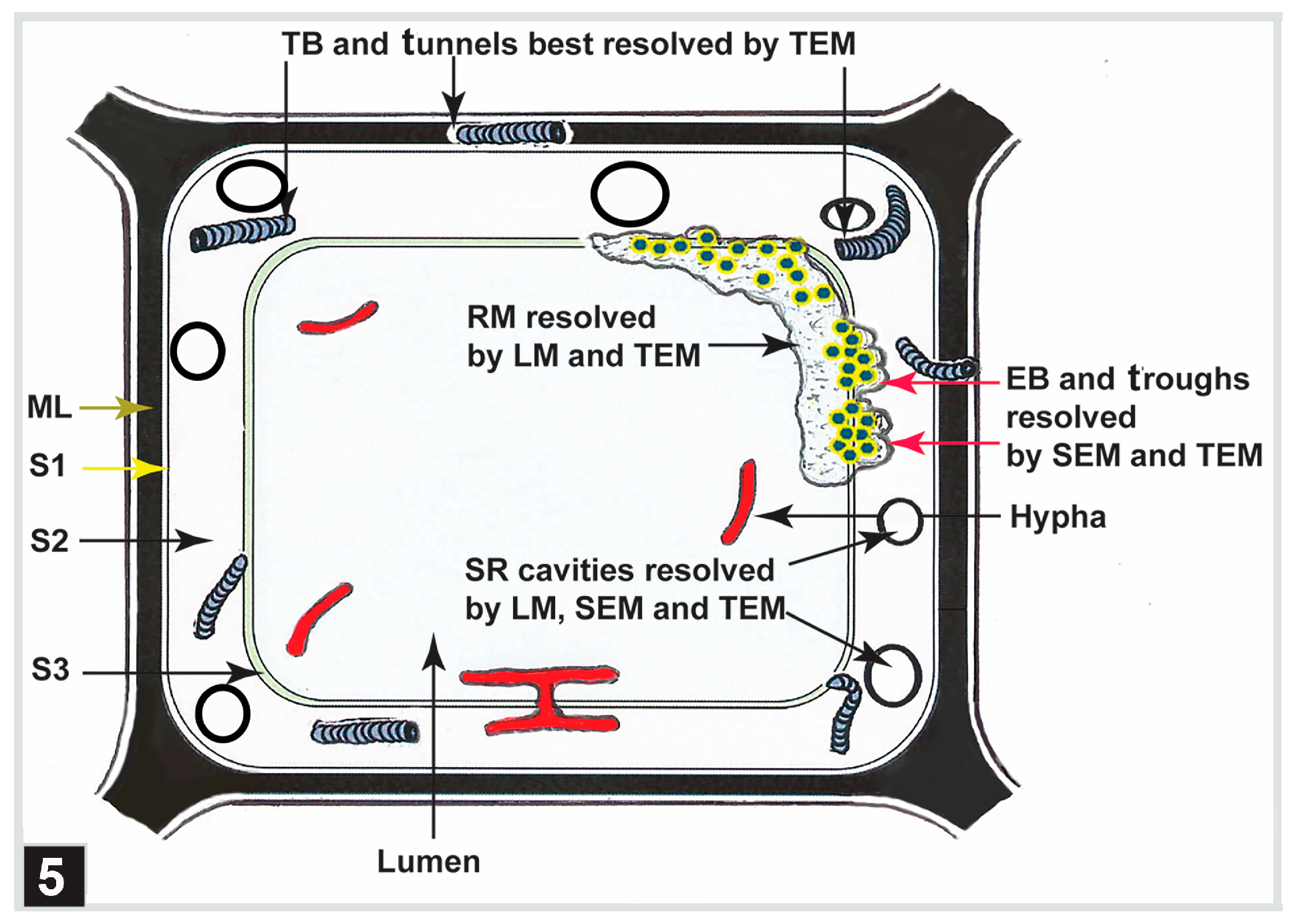
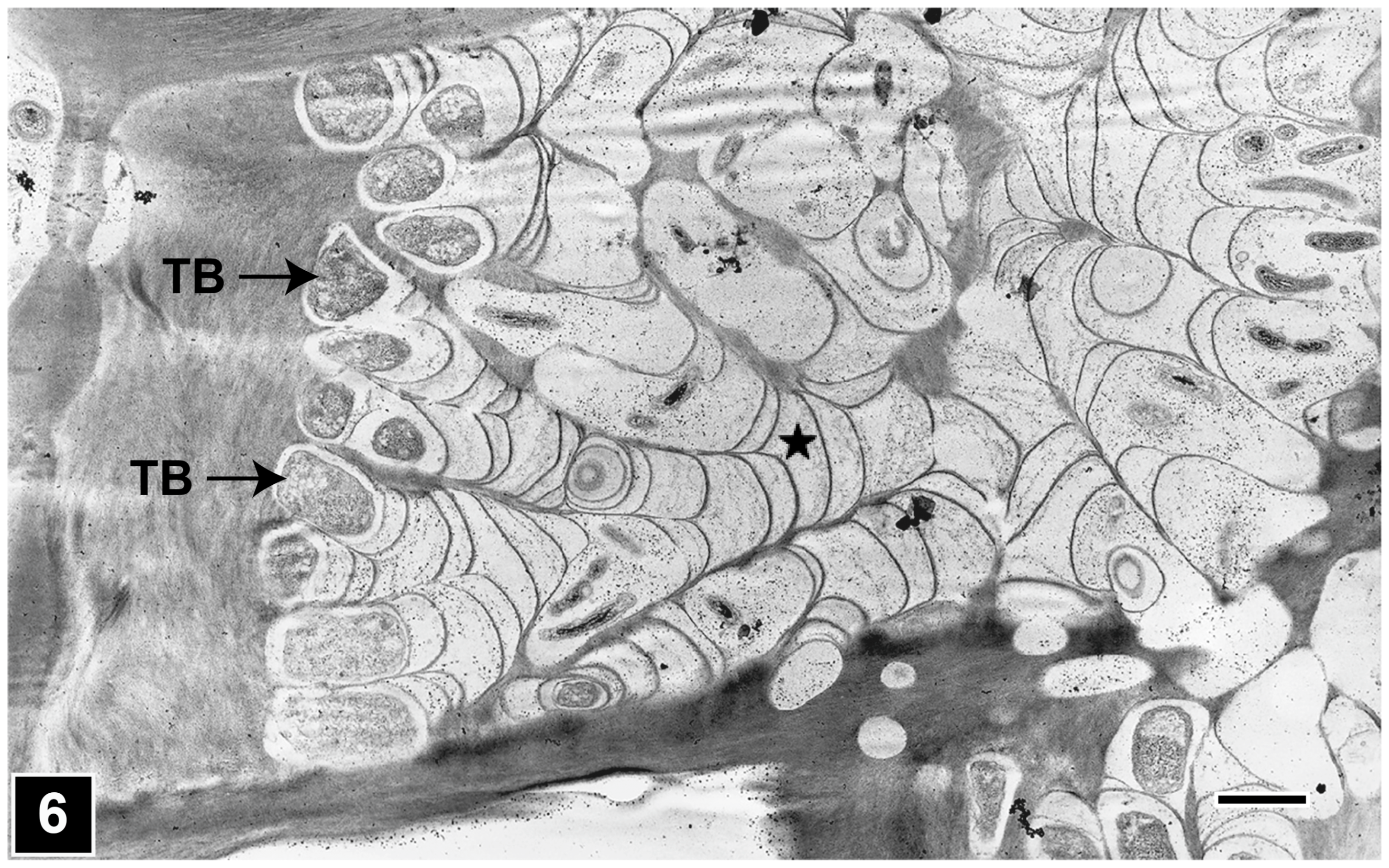
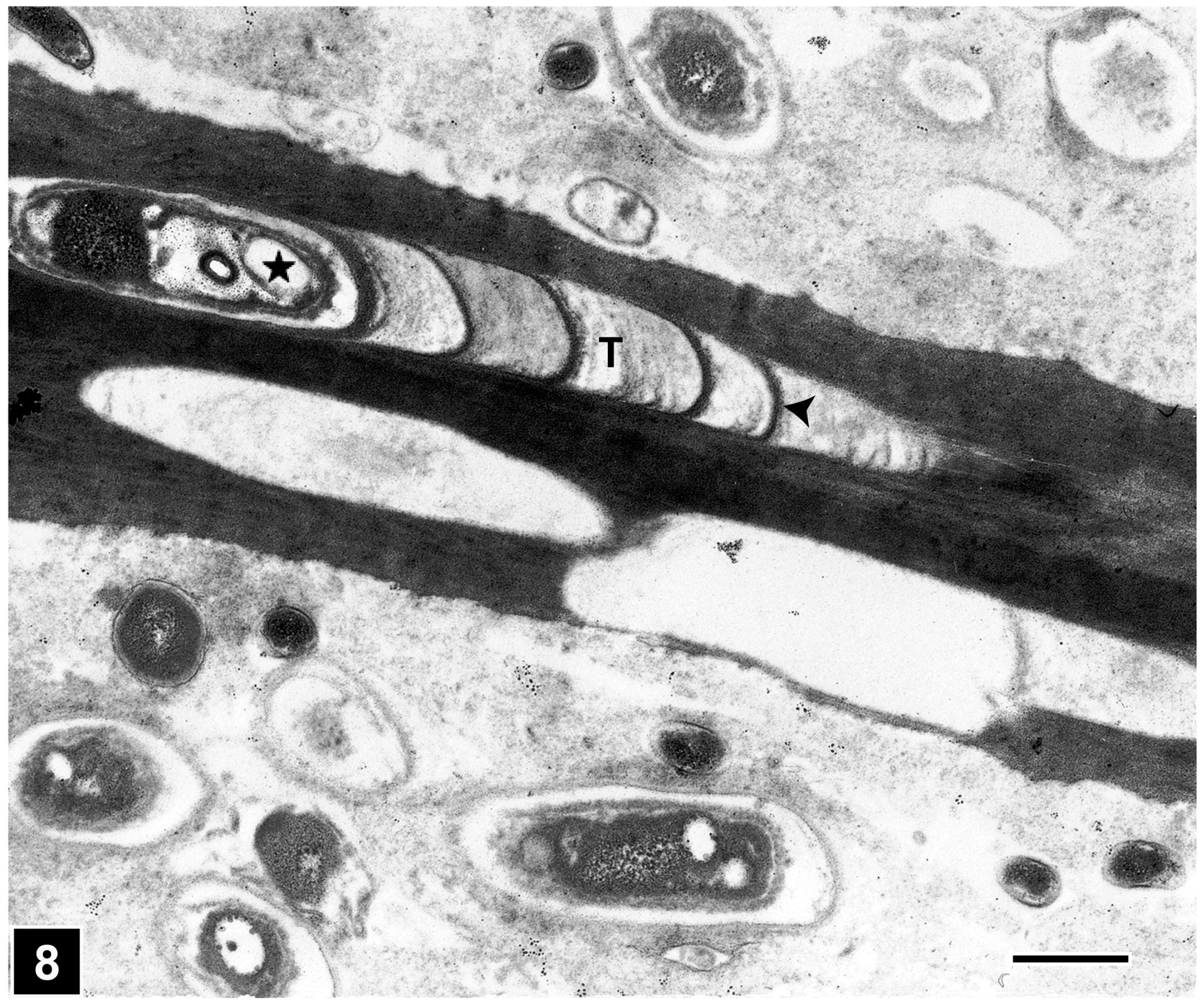
4.2.1. Bacterial Tunnelling
4.2.2. Bacterial Erosion
5. Highly Lignified Cell Wall Regions and Wood Structures Influence Microbial Degradation, as Revealed by Microscopy: Relevance to the Survival of Buried and Waterlogged Archaeological Wooden Objects
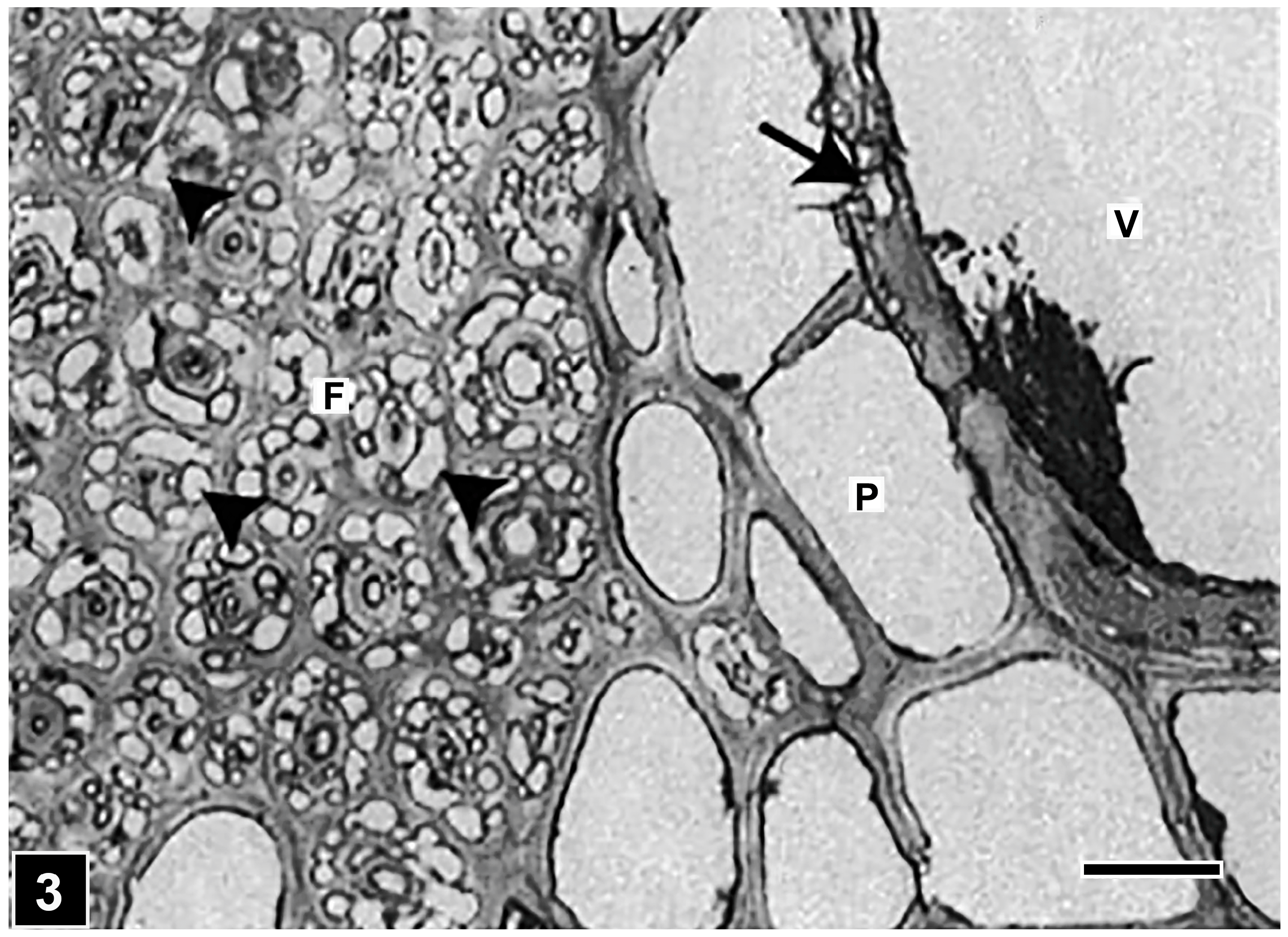
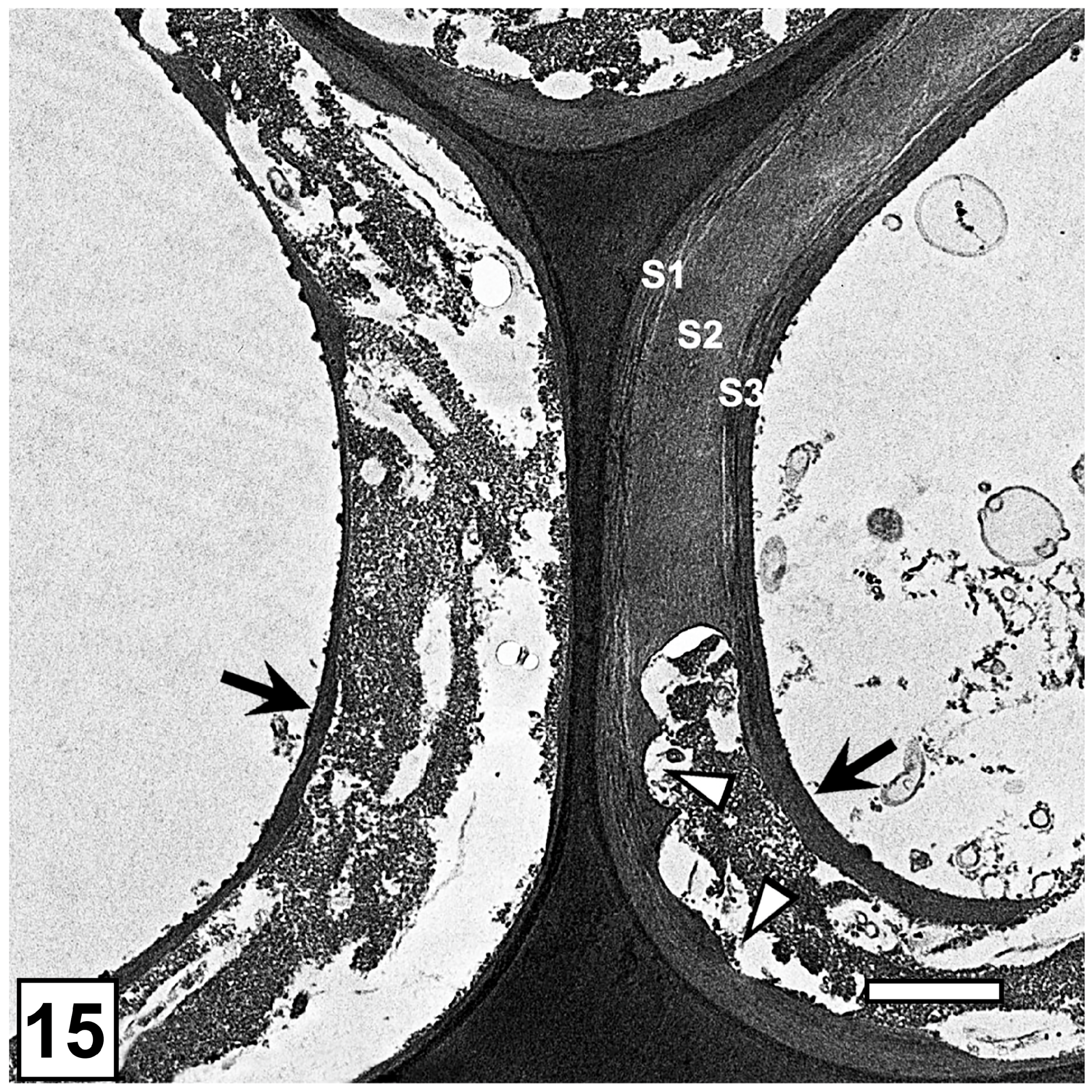
5.1. Middle Lamella and the S3 Layer
5.2. Outer Part of the S2 Layer (S2L) in Compression Wood
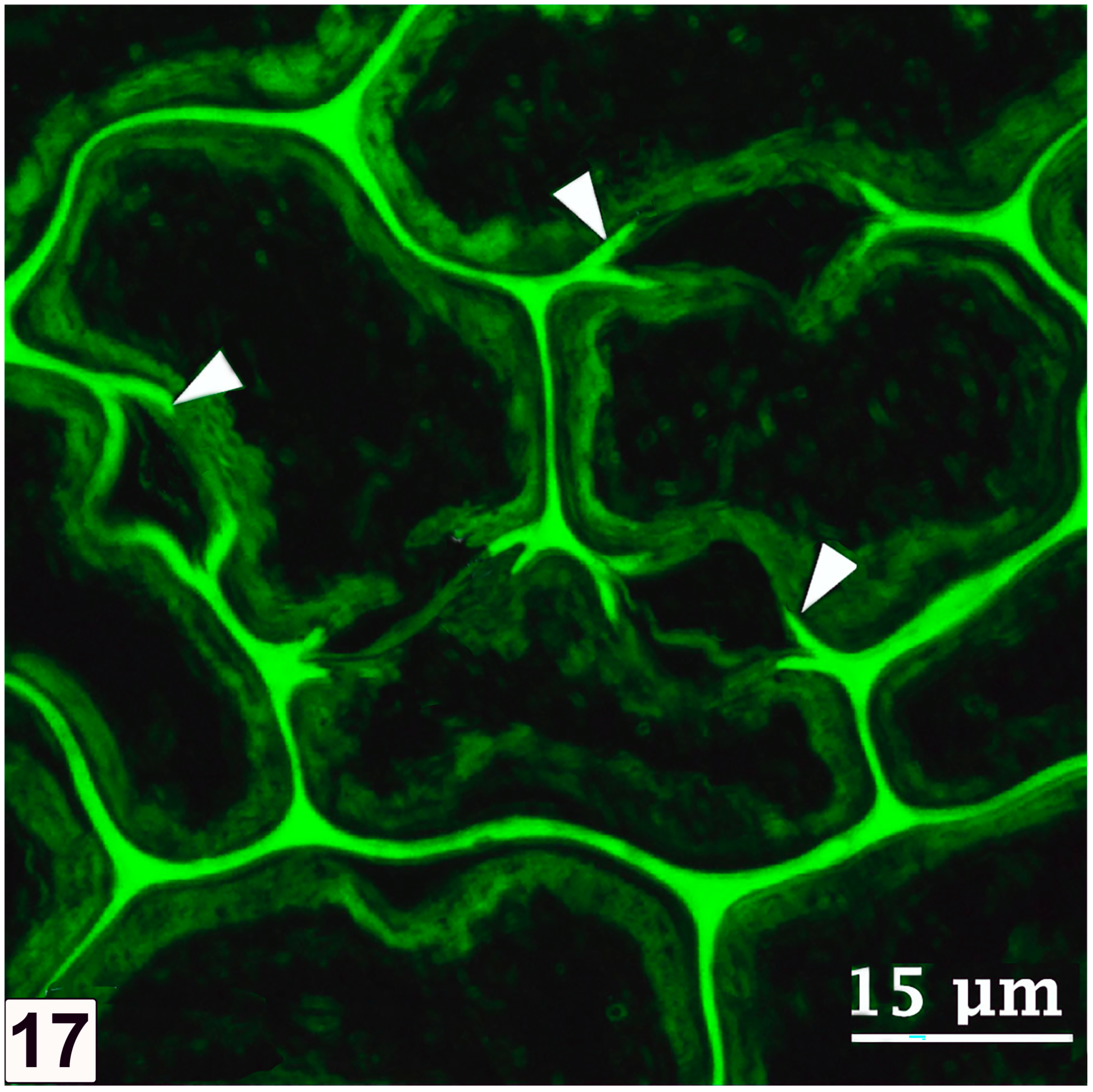
5.3. Initial Pit Borders
5.4. Warts and Vestures
5.5. Highly Lignified Ray Parenchyma
6. Heartwood and Heartwood Extractives Resist Microbial Degradation, as Revealed by Microscopy: Relevance to the Survival of Buried and Waterlogged Archaeological Wooden Objects
7. Concluding Remarks
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thieme, H. Lower Paleolithic throwing spears from Germany. Nature 1997, 385, 807–810. [Google Scholar] [CrossRef] [PubMed]
- Schoch, W.H.; Bigga, G.; Böhner, U.; Richter, P.; Terberger, T. New insights on the wooden weapons from the Paleolithic site of Schöningen. J. Hum. Evol. 2015, 89, 214–225. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, P. Half-million-year-old ‘Lincoln Logs’ may be first wood structure made by ancient humans. Science 2023, 381, 6664. [Google Scholar]
- Pournou, A. Biodegradation of Wooden Cultural Heritage: Origins and Decay Mechanisms in Aquatic and Terrestrial Ecosystems; Springer International Publishing: Berlin, Germany, 2020. [Google Scholar]
- Broda, M.; Hill, C.A.S. Conservation of waterlogged wood—Past, present and future perspectives. Forests 2021, 12, 1193. [Google Scholar] [CrossRef]
- Singh, A.P.; Kim, Y.S.; Chavan, R.R. Advances in understanding microbial deterioration of buried and waterlogged archaeological woods: A review. Forests 2022, 13, 394. [Google Scholar] [CrossRef]
- Klaassen, R.K.W.M. Bacterial decay in wooden foundation piles—Patterns and causes: A study of historical pile foundations in the Netherlands. Int. Biodeter. Biodegrad. 2008, 61, 45–60. [Google Scholar] [CrossRef]
- Elam, J.; Björdal, C.G. A review and case studies of factors affecting the stability of wooden foundation piles in urban environments exposed to construction work. Int. Biodeter. Biodegrad. 2020, 148, 104913. [Google Scholar] [CrossRef]
- Cha, M.Y.; Lee, K.H.; Kim, J.S.; Kim, Y.S. Variations in bacterial decay between cell types and between cell wall regions in waterlogged archaeological wood excavated in the intertidal zone. IAWA J. 2021, 42, 457–474. [Google Scholar] [CrossRef]
- Humar, M.; Balzano, A.; Kržišnik, D.; Lesar, B. Assessment of wooden foundation piles after 125 years of service. Forests 2021, 12, 143. [Google Scholar] [CrossRef]
- Daniel, G. Fungal and bacterial degradation: White rot, brown rot, soft rot and bacteria. In Deterioration and Protection of Sustainable Biomaterials; Schulz, T., Goodell, B., Nicholas, D.D., Eds.; ACS Symposium Series; American Chemical Society: Washington, DC, USA, 2014; pp. 23–56. [Google Scholar]
- Goodell, B.; Winandy, J.E.; Morrell, J.J. Fungal degradation of wood: Emerging data, new insights and changing perceptions. Coatings 2020, 10, 1210. [Google Scholar] [CrossRef]
- Zabel, R.A.; Morrell, J.J. Wood Microbiology: Decay and Its Prevention; Elsevier: San Diego, CA, USA, 2020. [Google Scholar]
- Daniel, G.; Nilsson, T. Developments in the study of soft rot and bacterial decay. In Forest Products Biotechnology; Bruce, A., Palfreyman, J.W., Eds.; Taylor and Francis: London, UK, 1998; pp. 37–62. [Google Scholar]
- Kim, Y.S.; Singh, A.P. Micromorphological characteristics of wood biodegradation in wet environments: A review. IAWA J. 2000, 21, 135–155. [Google Scholar] [CrossRef]
- Björdal, C.G. Microbial degradation of waterlogged archaeological wood. J. Cult. Herit. 2012, 13, S118–S122. [Google Scholar] [CrossRef]
- Singh, A.P. A review of microbial decay found in wooden objects of cultural heritage recovered from buried and waterlogged environments. J. Cult. Herit. 2012, 13, S16–S20. [Google Scholar] [CrossRef]
- Conrad, N.J.; Serangeli, J.; Bigga, G.; Rots, V. A 300,000 year-old-throwing stick from Schöningen, northern Germany, documents the evolution of human hunting. Nat. Ecol. Evol. 2020, 4, 885. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.P.; Butcher, J.A. Bacterial degradation of wood cell walls: A review of degradation patterns. J. Inst. Wood Sci. 1991, 12, 143–157. [Google Scholar]
- Daniel, G. Use of electron microscopy for aiding our understanding of wood biodegradation. FEMS Microbiol. Rev. 1994, 13, 199–233. [Google Scholar] [CrossRef]
- Daniel, G. Microscope techniques for understanding wood cell structure and biodegradation. In Secondary Xylem Biology: Origins, Functions, and Applications; Kim, Y.S., Funada, R., Singh, A.P., Eds.; Elsevier: London, UK, 2016; pp. 309–343. [Google Scholar]
- Singh, A.P.; Kim, Y.S.; Singh, T. Bacterial degradation of wood. In Secondary Xylem Biology: Origins, Functions, and Applications; Kim, Y.S., Funada, R., Singh, A.P., Eds.; Elsevier: London, UK, 2016; pp. 169–190. [Google Scholar]
- Schmidt, O.; Liese, W. Occurrence and significance of bacteria in wood. Holzforschung 1994, 48, 271–277. [Google Scholar] [CrossRef]
- Clausen, C.A. Bacterial association with decaying wood: A review. Int. Biodeter. Biodegrad. 1996, 37, 101–107. [Google Scholar] [CrossRef]
- Schmitt, U.; Singh, A.P.; Thieme, H.; Friedrich, P. Electron microscopic characterisation of cell wall degradation of the 400,000-year-old wooden Schöningen spears. Holz Roh Werkst. 2005, 63, 118–122. [Google Scholar] [CrossRef]
- Singh, A.P.; Nilsson, T.; Daniel, G. Bacterial attack of Pinus sylvestris wood under near-anaerobic conditions. J. Inst. Wood Sci. 1990, 11, 237–249. [Google Scholar]
- Kim, Y.S.; Singh, A.P.; Nilsson, T. Bacteria as important degraders in waterlogged archaeological woods. Holzforschung 1996, 50, 389–392. [Google Scholar] [CrossRef]
- Björdal, C.G. Waterlogged Archaeological Wood: Biodeterioration and its Implications for Conservation. Ph.D. Thesis, Swedish University of Agricultural Sciences, Uppsala, Sweden, 2000. [Google Scholar]
- Kim, Y.S.; Singh, A.P. Micromorphological characteristics of waterlogged archaeological woods attacked by marine microorganisms. In Recent Advances in Wood Anatomy; Donaldson, L.A., Singh, A.P., Butterfield, B.G., Whitehouse, L.J., Eds.; New Zealand Forest Research Institute: Rotorua, New Zealand, 1996; pp. 389–399. [Google Scholar]
- Blanchette, R.A. A review of microbial deterioration found in archaeological wood from different environments. Int. Biodeter. Biodegrad. 2000, 46, 189–204. [Google Scholar] [CrossRef]
- Kim, J.S.; Kim, M.; Lim, J.W.; Cha, M.Y.; Lee, K.H.; Yoon, Y.H.; Kim, Y.S. Characterisation of microbial decay and microbial communities in waterlogged archaeological rosewood (Dalbergia species). Forests 2023, 14, 1992. [Google Scholar] [CrossRef]
- Björdal, C.G.; Dayton, P.K. First evidence of microbial wood degradation in the coastal waters of the Antarctic. Sci. Rep. 2020, 10, 12774. [Google Scholar] [CrossRef] [PubMed]
- Giachi, G.; Bettazzi, F.; Chimichi, S.; Staccioli, G. Chemical characterisation of degraded wood in ships discivered in a recent excavation of the Etruscan and Roman harbour of Pisa. J. Cult. Herit. 2003, 4, 75–83. [Google Scholar] [CrossRef]
- Capretti, C.; Macchioni, N.; Pizzo, B.; Galota, G.; Giachi, G.; Giampaola, D. The characterisation of waterlogged archaeological wood. The three Roman ships found in Naples (Italy). Archaeometry 2008, 50, 855–876. [Google Scholar] [CrossRef]
- Macchioni, N.; Pizzo, B.; Capretti, C.; Giachi, G. How an integrated diagnostic approach can help in a correct evaluation of the state of preservation of waterlogged archaeological wooden artefacts. J. Archaeol. Sci. 2012, 39, 3255–3263. [Google Scholar] [CrossRef]
- Macchioni, N.; Capretti, C.; Sozzi, I.; Pizzo, B. Grading the decay of waterlogged archaeological wood according to anatomical characterisation. The case of the Fiave site (N-E Italy). Int. Biodeter. Biodegrad. 2013, 84, 54–64. [Google Scholar] [CrossRef]
- Sjöström, E.; Westermark, U. Chemical composition of wood and pulps: Basic constituents and their distribution. In Analytical Methods in Wood Chemistry, Pulping, and Papermaking; Sjöström, E., Alén, R., Eds.; Springer: Berlin, Germany, 1999; p. 3. [Google Scholar]
- Singh, A.P. Ultrastructural features of compression wood cells in relation to bacterial decay in Pinus radiata. In Recent Advances in Wood Anatomy; Donaldson, L.A., Singh, A.P., Butterfield, B.G., Whitehouse, L.J., Eds.; New Zealand Forest Research Institute: Rotorua, New Zealand, 1996; pp. 400–407. [Google Scholar]
- Singh, A.P. The ultrastructure of the attack of Pinus radiata mild compression wood by erosion and tunnelling bacteria. Can. J. Bot. 1997, 75, 1095–1102. [Google Scholar] [CrossRef]
- Donaldson, L.A.; Singh, A.P. Formation and structure of compression wood. In Cellular Aspects of Wood Formation; Fromm, J., Ed.; Springer: Berlin, Germany, 2013; pp. 225–256. [Google Scholar]
- Kim, Y.S.; Singh, A.P. Micromorphological characteristics of compression wood degradation in waterlogged archaeological pine wood. Holzforschung 1999, 53, 381–385. [Google Scholar] [CrossRef]
- Schmitt, U.; Singh, A.P.; Kim, Y.S. Wood as an ecological niche for microorganisms: Wood formation, structure, and cell wall composition. In Forest Microbiology: Tree Microbiome—Phillosphere, Endosphere and Rhizosphere; Asiegbu, F.O., Kovalchuk, A., Eds.; Elsevier: San Diego, CA, USA, 2021; pp. 17–34. [Google Scholar]
- Hillis, W.E. Heartwood and Tree Exudates; Springer: Berlin, Germany, 1987. [Google Scholar]
- Singh, A.P.; Kim, Y.S.; Chavan, R.R. Relationship of wood cell wall ultrastructure to bacterial degradation of wood. IAWA J. 2019, 40, 845–870. [Google Scholar] [CrossRef]
- McFarlane, H.E.; Döring, A.; Persson, S. The cell biology of cellulose synthesis. Ann. Rev. Plant Biol. 2014, 65, 69–94. [Google Scholar] [CrossRef] [PubMed]
- Giddings, T.H.; Brower, D.L.; Staehelin, L.A. Visualisation of particle complexes in the plasma membrane of Micrasterias denticulata associated with the formation of cellulose fibrils in primary and secondary cell walls. J. Cell Biol. 1980, 84, 327–339. [Google Scholar] [CrossRef]
- Mueller, S.C.; Brown, R.M. Evidence for an intramembrane component associated with a cellulose microfibril-synthesising complex in higher plants. J. Cell Biol. 1980, 84, 315–326. [Google Scholar] [CrossRef]
- Ledbetter, M.C.; Porter, K.R. A “microtubule” in plant cell fine structure. J. Cell Biol. 1963, 19, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, L.A. Wood cell wall ultrastructure: The key to understanding wood properties and behaviour. IAWA J. 2019, 40, 645–672. [Google Scholar] [CrossRef]
- Meents, M.J.; Watanabe, Y.; Samuels, A.L. The cell biology of secondary cell wall biosynthesis. Ann. Bot. 2018, 121, 1107–1125. [Google Scholar] [CrossRef] [PubMed]
- McCann, M.C.; Wells, B.; Roberts, K. Direct visualisation of cross-links in the primary cell walls. J. Cell Sci. 1990, 96, 323–334. [Google Scholar] [CrossRef]
- Donaldson, L.A.; Singh, A.P. Bridge-like structures between cellulose microfibrils in radiata pine (Pinus radiata D. Don) kraft pulp and holocellulose. Holzforschung 1998, 52, 449–454. [Google Scholar] [CrossRef]
- Fujino, T.; Sone, Y.; Mitsuishi, Y.; Itoh, T. Characterisation of cross-links between cellulose microfibrils, and their occurrence during elongation growth in pea epicotyl. Plant Cell Physiol. 2000, 41, 486–494. [Google Scholar] [CrossRef]
- Donaldson, L.A. Lignification and lignin topochemistry—An ultrastructural view. Phytochemistry 2001, 57, 859–873. [Google Scholar] [CrossRef]
- Li, M.; Pu, Y.; Ragauskas, A.J. Current understanding of the correlation of lignin structure with biomass recalcitrance. Front. Chem. 2016, 4, 45. [Google Scholar] [CrossRef]
- Zhang, X.; Li, L.; Xu, F. Chemical characteristics of wood cell wall with an emphasis on ultrastructure: A mini-review. Forests 2022, 13, 439. [Google Scholar] [CrossRef]
- Reza, M.; Kontturi, E.; Jääskeläinen, A.-S.; Vuorinen, T.; Ruokolainen, J. Transmission electron microscopy for wood and fibre analysis—A review. BioResources 2015, 10, 6230–6261. [Google Scholar] [CrossRef]
- Donaldson, L.A.; Xu, P. Microfibril orientation across the secondary cell wall of radiata pine tracheids. Trees 2005, 19, 644–653. [Google Scholar] [CrossRef]
- Booker, R.E. The importance of the S3 cell wall layer in collapse prevention and wood hardness. In Proceedings of the 24th Forest Products Research Conference, Melbourne, Australia, 15–18 November 1993; CSIRO Division of Forest Products, Clayton: Victoria, Australia; pp. 1–13. [Google Scholar]
- Booker, R.E.; Sell, J. The nanostructure of the cell wall of softwoods and its functions in a living tree. Holz Roh-Werkst. 1998, 56, 1–8. [Google Scholar] [CrossRef]
- Donaldson, L.A. Microfibril angle: Measurement, variation and relationships—A review. IAWA J. 2008, 29, 345–386. [Google Scholar] [CrossRef]
- Atalla, R.H.; Hackney, J.M.; Uhlin, I.; Thomson, N.S. Hemicelluloses as structural regulators in aggregations on native cellulose. Int. J. Biol. Macromol. 1993, 15, 109–112. [Google Scholar] [CrossRef]
- Berglund, J.; Mikkelsen, D.; Flanagan, B.M.; Dhital, S.; Gaunitz, S.; Henriksson, G.; Lindström, M.E.; Yakubov, G.E.; Gidley, M.J.; Vilaplana, F. Wood hemicelluloses exert distinct biomechanical contributions to cellulose fibrillar network. Nat. Commun. 2020, 11, 4692. [Google Scholar] [CrossRef]
- Martinez-Abad, A.; Berglund, J.; Toriz, G.; Gatenholm, P.; Henriksson, G.; Lindström, M.; Wohlert, J.; Vilaplana, F. Regular motifs in xylan modulate molecular flexibility and interactions with cellulose surfaces. Plant Physiol. 2017, 175, 1579–1592. [Google Scholar] [CrossRef]
- Cosgrove, D.J.; Jarvis, M.C. Comparative structure and biomechanics of plant primary and secondary cell walls. Front. Plant Sci. 2012, 3, 204. [Google Scholar] [CrossRef]
- Burgert, I.; Keplinger, T. Plant micro- and nanomechanics: Experimental techniques for plant cell wall analysis. J. Exp. Bot. 2013, 64, 4635–4649. [Google Scholar] [CrossRef] [PubMed]
- Fromm, J.; Rockell, B.; Lautner, S.; Windeisen, E.; Wanner, G. Lignin distribution in wood cell walls determined by TEM and backscattered SEM techniques. J. Struct. Biol. 2003, 143, 77–84. [Google Scholar] [CrossRef]
- Donaldson, L.A. S3 lignin concentration in radiata pine tracheids. Wood Sci. Technol. 1987, 21, 227–234. [Google Scholar] [CrossRef]
- De Moraes Pereira, M.C.; Junior, C.C. Strength and stiffness of cross laminated timber (CLT) panels produced with Pinus and Eucalyptus: Experimental and analytical comparisons. Rev. Mater. 2019, 24, e12369. [Google Scholar]
- Wang, X.; Ren, H.; Zhang, B.; Fei, B.; Burgert, I. Cell wall structure and formation of maturing fibres of Moso bamboo (Phyllostachys pubescens) increase buckling resistance. JR Soc. Interface 2012, 9, 988–996. [Google Scholar] [CrossRef] [PubMed]
- Parthasarathy, M.V.; Klotz, L.H. Palm “wood”. Part II: Ultrastructural aspects of sieve elements, tracheary elements and fibre. Wood Sci. Technol. 1976, 10, 247–277. [Google Scholar] [CrossRef]
- Parameswaran, N.; Liese, W. Ultrastructural aspects of bamboo cells. Cellul. Chem. Technol. 1980, 14, 587–609. [Google Scholar]
- Daniel, G.; Nilsson, T. Polylaminate concentric cell wall layering in fibres of Homalium foetidum and its effect on degradation by microfungi. In Recent Advances in Wood Anatomy; Donaldson, L.A., Singh, A.P., Butterfield, B.G., Whitehouse, L.J., Eds.; New Zealand Forest Research Institute: Rotorua, New Zealand, 1996; pp. 369–372. [Google Scholar]
- Shah, D.U.; Konnerth, J.; Ramage, M.H.; Gusenbauer, C. Mapping thermal conductivity across bamboo cell walls with scanning thermal microscopy. Sci. Rep. 2019, 9, 16667. [Google Scholar] [CrossRef]
- Singh, A.P.; Wong, A.H.H.; Kim, Y.S.; Wi, S.G. Resistance of the S1 layer in kempas heartwood fibres to soft rot decay. IAWA J. 2018, 39, 37–42. [Google Scholar] [CrossRef]
- Singh, A.P.; Wong, A.H.H.; Kim, Y.S.; Wi, S.G. The Relationship of Cell Wall Ultrastructure to Soft Rot Decay in Kempas (Koompassia malaccensis) Heartwood; Document No. IRG/WP/04-10541; International Research Group on Wood Preservation: Stockholm, Sweden, 2004. [Google Scholar]
- Timell, T.E. Compression Wood in Gymnosperms; Springer: Berlin, Germany, 1986. [Google Scholar]
- Donaldson, L.A.; Singh, A.P.; Yoshinaga, A.; Takabe, K. Lignin distribution in mild compression wood of Pinus radiata. Can. J. Bot. 1999, 77, 41–50. [Google Scholar]
- Singh, A.P.; Donaldson, L.A. Ultrastructure of tracheid cell walls in radiata pine (Pinus radiata) mild compression wood. Can. J. Bot. 1999, 77, 32–40. [Google Scholar]
- Yumoto, M.; Ishida, S.; Fukazawa, K. Studies on the formation and structure of gymnosperm wood cells induced by artificial inclination in young trees of Picea glauca. IV. Gradation of the severity of compression wood tracheids. Res. Bull. Coll. Exp. For. Hokkaido Univ. 1983, 42, 409–454. [Google Scholar]
- Singh, A.P.; Kim, Y.S.; Park, B.D.; Chung, G.C.; Wong, A.H.H. Presence of a distinct S3 layer in mild compression wood tracheids of Pinus radiata. Holzforschung 2003, 57, 243–247. [Google Scholar] [CrossRef]
- Altaner, C.; Hapca, A.I.; Knox, J.P.; Jarvis, M.C. Detection of β-1,4-galactan in compression wood of Sitka spruce [Picea sitchensis (Bong.) Carriére] by immunofluorescence. Holzforschung 2007, 61, 311–316. [Google Scholar] [CrossRef]
- Schmitt, U.; Singh, A.P.; Frankenstein, C.; Möller, R. Cell wall modifications in woody stems induced by mechanical stress. NZ J. For. Sci. 2006, 36, 72–86. [Google Scholar]
- Donaldson, L.A.; Knox, J.P. Localisation of cell wall polysaccharides in normal and compression wood of radiata pine: Relationships with lignification and microfibril orientation. Plant Physiol. 2012, 158, 642–653. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Chavan, R.R.; Smith, B.G.; McArdle, B.H.; Harris, P.J. Tracheid cell-wall structures and locations of 1,4-β-D-galactans and 1,3-β-D-glucans in compression woods of radiata pine (Pinus radiata D. Don). BMC Plant Biol. 2016, 16, 194. [Google Scholar] [CrossRef]
- Singh, A.P.; Wakeling, R.N. Bacterial Degradation of Pinus radiata Compression Wood; Document No. IRG/WP/96-10153; International Research Group on Wood Preservation: Stockholm, Sweden, 1996. [Google Scholar]
- Donaldson, L.A.; Singh, A.P. Reaction wood. In Secondary Xylem Biology: Origins, Functions, and Applications; Kim, Y.S., Funada, R., Singh, A.P., Eds.; Elsevier: London, UK, 2016; pp. 93–110. [Google Scholar]
- Foston, M.; Hubbell, C.A.; Samuel, R.; Jung, S.; Fan, H.; Ding, S.-Y.; Zeng, V.; Jawdy, S.; Davis, M.; Sykes, R.; et al. Chemical, ultrastructural and supramolecular analysis of tension wood in Populus tremula × alba as a model substrate for reduced recalcitrance. Energy Environ. Sci. 2011, 4, 4962–4971. [Google Scholar] [CrossRef]
- Okuyama, T.; Yamamoto, H.; Yoshida, M.; Hattori, Y.; Archer, R.R. Growth stresses in tension wood: Role of microfibrils and lignification. Ann. For. Sci. 1994, 51, 291–300. [Google Scholar] [CrossRef]
- Baum, S.; Schwarze, F.W.M.R.; Fink, S. Persistence of the gelatinous layer within altered tension wood fibres of beech degraded by Ustulina deusta. New Phytol. 2000, 147, 347–355. [Google Scholar] [CrossRef]
- Lehringer, C.; Daniel, G.; Schmitt, U. TEM/FE-SEM studies on tension wood fibres of Acer spp., Fagus sylvatica L. and Quercus robur L. Wood Sci. Technol. 2009, 43, 691–702. [Google Scholar] [CrossRef]
- Kim, J.S.; Daniel, G. Distribution of lignin, pectins and hemicelluloses in tension wood fibres of European ash (Fraxinus excelsior). IAWA J. 2019, 40, 741–764. [Google Scholar] [CrossRef]
- Sawada, D.; Kalluri, U.C.; O’Neill, H.; Urban, V.; Langan, P.; Davison, B.; Pingali, S.V. Tension wood structure and morphology conducive for better enzymatic digestion. Biotechnol. Biofuels 2018, 11, 44. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Jebrane, M.; Terziev, N.; Daniel, G. Enzymatic hydrolysis of the gelatinous layer in tension wood of Salix varieties as a measure of accessible cellulose for biofuels. Biotechnol. Biofuels Bioprod. 2021, 14, 141. [Google Scholar] [CrossRef] [PubMed]
- Toumpanaki, E.; Shah, D.U.; Eichhorn, S.J. Beyond what meets the eye: Imaging and imagining wood mechanical-structural properties. Adv. Mater. 2021, 33, 2001613. [Google Scholar] [CrossRef]
- Elliot, A.D. Confocal microscopy: Principles and modern practices. Curr. Protoc. Cytom. 2020, 92, e68. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, L.A. Localising molecules in plant cell walls using fluorescence microscopy. In Methods in Molecular Biology; Pellicciari, C., Biggiogera, M., Malatesta, M., Eds.; Springer Nature: Berlin/Heidelberg, Germany, 2023; Volume 2566, pp. 243–259. [Google Scholar]
- Koch, G.; Kleist, G. Application of UV microspectrophotometry to localise lignin and phenolic extractives in plant cell walls. Holzforschung 2001, 55, 563–567. [Google Scholar] [CrossRef]
- Čufar, K.; Gričar, J.; Zupančič, M.; Koch, G.; Schmitt, U. Anatomy, cell wall structure and topochemistry of waterlogged archaeological wood aged 5200 and 4500 years. IAWA J. 2008, 29, 55–68. [Google Scholar] [CrossRef]
- Rehbein, M.; Koch, G.; Schmitt, U.; Huckfeldt, T. Topochemical and transmission electron microscopic studies of bacterial decay in pine (Pinus sylvestris L.) harbour foundation piles. Micron 2013, 44, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Gierlinger, N.; Schwanninger, M. Chemical imaging of poplar wood cell walls by confocal Raman microscopy. Plant Physiol. 2006, 140, 1246–1254. [Google Scholar] [CrossRef]
- Gierlinger, N.; Keplinger, T.; Harrington, M. Imaging plant cell walls by confocal Raman microscopy. Nat. Protoc. 2012, 7, 1694–1708. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, N.B.; Gierlinger, N.; Thygesen, L.G. Bacterial and abiotic decay in waterlogged archaeological Picea abies (L.) Karst studied by confocal Raman imaging and ATR-FTIR spectroscopy. Holzforschung 2015, 69, 103–112. [Google Scholar] [CrossRef]
- Gierlinger, N. New insights into plant cell walls byvibrational microspectroscopy. Appl. Specrosc. Rev. 2018, 53, 517–551. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.P.; Kim, Y.S.; Wi, S.G.; Lee, K.H.; Kim, I.J. Evidence of the degradation of middle lamella in a waterlogged archaeological wood. Holzforschung 2003, 57, 115–119. [Google Scholar] [CrossRef]
- Xu, P.; Donaldson, L.A.; Gergely, Z.R.; Staehelin, L.A. Dual-axis electron tomography: A new approach for investigating the spatial organisation of wood cellulose microfibrils. Wood Sci. Technol. 2007, 41, 101–116. [Google Scholar] [CrossRef]
- Takabe, K.; Kim, J.S. Rapid freezing and immunocytochemistry provide new information on cell wall formation in woody plants. In Secondary Xylem Biology: Origins, Functions, and Applications; Kim, Y.S., Funada, R., Singh, A.P., Eds.; Elsevier: London, UK, 2016; pp. 345–361. [Google Scholar]
- Gilani, M.S.; Boone, M.N.; Mader, K.; Swarze, F.W.M.R. Synchrotron X-ray micro-tomography imaging and analysis of wood degraded by Physisporinus vitreus and Xylaria longipes. J. Struct. Biol. 2014, 187, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Gelbrich, J.; Mai, C.; Militz, H. Chemical changes in wood degraded by bacteria. Int. Biodeter. Biodegrad. 2008, 61, 24–32. [Google Scholar] [CrossRef]
- Anagnost, S.E. Light microscopic diagnosis of wood decay. IAWA J. 1998, 19, 141–167. [Google Scholar] [CrossRef]
- Anagnost, S.E.; Mark, R.E.; Hanna, R.B. Utilisation of soft rot cavity orientation for the determination of microfibril angle. Part I. Wood Fiber Sci. 2000, 32, 81–87. [Google Scholar]
- Khalili, S.; Nilsson, T.; Daniel, G. The use of soft rot fungi for determining the microfibrillar orientation in the S2 layer of pine tracheids. Holz Roh Werkst. 2001, 58, 439–447. [Google Scholar] [CrossRef]
- Brändström, J.; Daniel, G.; Nilsson, T. Use of soft rot cavities to determine microfibril angles in wood: Advantages, disadvantages and possibilities. Holzforschung 2002, 56, 468–472. [Google Scholar] [CrossRef]
- Nilsson, T. Formation of soft rot cavities in various cellulose fibres by Humicola alopallonella Meyers and Moore; Skogshögskolan: Hässleholm, Sweden, 1974. [Google Scholar]
- Singh, A.P. Initial pit borders in Pinus radiata are resistant to degradation by soft rot fungi and erosion bacteria but not tunnelling bacteria. Holzforschung 1997, 51, 15–18. [Google Scholar]
- Singh, A.P.; Schmitt, U.; Möller, R.; Dawson, B.S.W.; Koch, G. Ray tracheids in Pinus radiata are more highly resistant to soft rot as compared to axial tracheids: Relationship to lignin concentration. Wood Sci. Technol. 2006, 40, 16–25. [Google Scholar] [CrossRef]
- Kim, Y.S.; Singh, A.P. Wood as cultural heritage material and its deterioration by biotic and abiotic agents. In Secondary Xylem Biology: Origins, Functions, and Applications; Kim, Y.S., Funada, R., Singh, A.P., Eds.; Elsevier: London, UK, 2016; pp. 233–257. [Google Scholar]
- Brandström, J. Microfibril angle of the S1 cell wall layer of Norway spruce compression wood tracheids. IAWA J. 2004, 25, 415–423. [Google Scholar] [CrossRef]
- Hale, M.D.; Eaton, R.A. The ultrastructure of soft rot fungi. Cavity-forming hyphae in wood cell walls. Mycologia 1985, 77, 594–605. [Google Scholar] [CrossRef]
- Anagnost, S.E.; Worrall, J.; Wang, C. Diffuse cavity formation in soft rot of pine. Wood Sci. Technol. 1994, 28, 199–208. [Google Scholar] [CrossRef]
- Blanchette, R.A.; Held, B.W.; Jurgens, J.A.; McNew, D.L.; Harrington, T.C.; Duncan, S.M.; Farrell, R.L. Wood-destroying soft rot fungi in the historic expedition huts of Antarctica. Appl. Environ. Microbiol. 2004, 70, 1328–1335. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.P.; Dawson, B.S.W.; Rickard, C.; Bond, J.; Singh, A. Light, confocal and scanning electron microscopy of wood-adhesive interface. Microsc. Anal. 2008, 22, 5–8. [Google Scholar]
- Daniel, G.; Nilsson, T. Interactions between soft rot fungi and CCA preservatives in Betula verrucosa. J. Inst. Wood Sci. 1989, 11, 162–171. [Google Scholar]
- Nilsson, T.; Daniel, G.; Kirk, T.K.; Obst, J.R. Chemistry and microscopy of wood decay by some higher ascomycetes. Holzforschung 1989, 43, 11–18. [Google Scholar] [CrossRef]
- Luna, M.L.; Murace, M.A.; Keil, G.D.; Otano, M.E. Patterns of decay caused by Pycnoporus sanguineus and Ganoderma lucidum (Aphyllophorales) in poplar woods. IAWA J. 2004, 25, 425–433. [Google Scholar] [CrossRef]
- Daniel, G. Fungal degradation of wood cell walls. In Secondary Xylem Biology: Origins, Functions, and Applications; Kim, Y.S., Funada, R., Singh, A.P., Eds.; Elsevier: London, UK, 2016; pp. 131–167. [Google Scholar]
- Björdal, C.G.; Daniel, G.; Nilsson, T. Depth of burial, an important factor in controlling bacterial decay of waterlogged archaeological poles. Int. Biodeter. Biodegrad. 2000, 45, 15–26. [Google Scholar] [CrossRef]
- Blanchette, R.A.; Nilsson, T.; Daniel, G.; Abad, A. Biological degradation of wood. In Archaeological Wood; Rowell, R.M., Barbour, R.J., Eds.; American Chemical Society: Washington DC, USA, 1990; pp. 141–174. [Google Scholar]
- Kim, Y.S.; Singh, A.P. Ultrastructural aspects of bacterial attacks on a submerged ancient wood. Mok. Gakk. 1994, 40, 554–562. [Google Scholar]
- Björdal, C.G.; Nilsson, T.; Daniel, G. Microbial decay of waterlogged archaeological wood found in Sweden. Int. Biodeter. Biodegrad. 1999, 43, 63–73. [Google Scholar] [CrossRef]
- Nilsson, T.; Daniel, G. Tunnelling Bacteria; Document No. IRG/WP 1186; International Research Group on Wood Preservation: Stockholm, Sweden, 1983. [Google Scholar]
- Singh, A.P.; Nilsson, T.; Daniel, G. Attack of pulped fibres by erosion bacteria. Mater. Org. 1987, 22, 257–269. [Google Scholar]
- Singh, A.P.; Butcher, J.A. Degradation of CCA-treated Pinus radiata posts by erosion bacteria. J. Inst. Wood Sci. 1985, 10, 140–144. [Google Scholar]
- Daniel, G.; Nilsson, T. Ultrastructural Observations on Wood Degrading Erosion Bacteria; Document No. IRG/WP 1283; International Research Group on Wood Preservation: Stockholm, Sweden, 1986. [Google Scholar]
- Daniel, G.; Nilsson, T.; Singh, A.P. Degradation of lignocellulosics by unique tunnel-forming bacteria. Can. J. Microbiol. 1987, 33, 943–948. [Google Scholar] [CrossRef]
- Nilsson, T.; Singh, A.P. Tunnelling bacteria and tunnelling of wood cell walls. In McGraw-Hill Encyclopedia of Science and Technology; McGraw-Hill: New York, NY, USA, 2014; pp. 395–399. [Google Scholar]
- Singh, A.P. Certain aspects of bacterial degradation of Pinus radiata wood. IAWA Bull. N.S. 1989, 10, 405–415. [Google Scholar] [CrossRef]
- Pedersen, N.B.; Lucejko, J.J.; Modugno, F.; Björdal, C.G. Correlation between bacterial decay and chemical changes in waterlogged archaeological wood analysed by light microscopy and Py-GC/MS. Holzforschung 2021, 75, 635–645. [Google Scholar] [CrossRef]
- Daniel, G.; Singh, A.P.; Nilsson, T. Ultrastructural and immunocytochemical studies on the window and bordered pit membranes of Pinus sylvestris L. In Recent Advances in Wood Anatomy; Donaldson, L.A., Singh, A.P., Butterfield, B.G., Whitehouse, L.J., Eds.; New Zealand Forest Research Institute: Rotorua, New Zealand, 1996; pp. 373–383. [Google Scholar]
- Hafrén, J.; Daniel, G.; Westermark, U. The distribution of acidic and esterified pectin in cambium, developing xylem and mature xylem of Pinus sylvestris. IAWA J. 2000, 21, 157–168. [Google Scholar] [CrossRef]
- Nagashima, Y.; Fukuda, R.; Sato, S.; Moroboshi, N.; Haraguchi, T. Coexistence of microfungal and bacterial degradation in a single wood cell wall. Mok. Gakk. 1990, 36, 480–486. [Google Scholar]
- Santhakumaran, L.N.; Singh, A.P. Destruction of Two Tropical Timbers by Marine Borers and Microorganisms in Goa Waters (India); Document No. IRG/WP 4176-92; International Research Group on Wood Preservation: Stockholm, Sweden, 1990. [Google Scholar]
- Singh, A.P.; Wakeling, R.N. Presence of widespread bacterial attacks in preservative-treated cooling tower timbers. NZ J. For. Sci. 1997, 27, 79–85. [Google Scholar]
- Singh, A.P.; Nilsson, T.; Daniel, G. Ultrastructure of the attack of the wood of two high lignin tropical hardwood species, Homalium foetidum and Alstonia scholaris, by tunnelling bacteria. J. Inst. Wood Sci. 1987, 11, 26–42. [Google Scholar]
- Björdal, C.G.; Elam, J. Bacterial degradation of nine wooden foundation piles from Gothenburg historic city centre and correlation to wood quality, environment, and the time in service. Int. Biodeter. Biodegrad. 2021, 164, 105288. [Google Scholar] [CrossRef]
- Holt, D.M.; Jones, E.B.G. Bacterial degradation of lignified wood cell walls in anaerobic aquatic habitats. Appl. Environ. Microbiol. 1983, 46, 722–727. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, U.P. Raman imaging to investigate ultrastructure and composition of plant cell walls: Distribution of lignin and cellulose in black spruce wood (Picea mariana). Planta 2006, 224, 1141–1153. [Google Scholar] [CrossRef] [PubMed]
- Fergus, B.J.; Procter, A.R.; Scott, J.A.N.; Goring, D.A.I. The distribution of lignin in spruce wood as determined by ultraviolet microscopy. Wood Sci. Technol. 1969, 3, 117–138. [Google Scholar] [CrossRef]
- Xu, F.; Zhong, X.C.; Sun, R.C.; Lu, Q. Anatomy, ultrastructure and lignin distribution in cell wall of Caragana korshinskii. Ind. Crops Prod. 2006, 24, 186–193. [Google Scholar] [CrossRef]
- Sant’anna, C.; Costa, L.T.; Abud, Y.; Biancatto, L.; Miguens, F.C.; De Souza, W. Sugarcane cell wall structure and lignin distribution investigated by confocal and electron microscopy. Microsc. Res. Tech. 2013, 76, 829–834. [Google Scholar] [CrossRef]
- Donaldson, L.A.; Vaidya, A. Visualising recalcitrance by colocalisation of cellulase, lignin and cellulose in pretreated pine biomass using fluorescence microscopy. Sci. Rep. 2017, 7, 44386. [Google Scholar] [CrossRef] [PubMed]
- Maceda, A.; Terrazas, T. Fluorescence microscopy methods for the analysis and characterisation of lignin. Polymers 2022, 14, 961. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, L.A. Within- and between-tree variation in the tracheid cell wall of Pinus radiata. NZ J. For. Sci. 1985, 15, 361–369. [Google Scholar]
- Singh, A.P.; Nilsson, T.; Daniel, G. Variable resistance of Pinus sylvestris wood components to attack by wood degrading bacteria. In Recent Advances in Wood Anatomy; Donaldson, L.A., Singh, A.P., Butterfield, B.G., Whitehouse, L.J., Eds.; New Zealand Forest Research Institute: Rotorua, New Zealand, 1996; pp. 408–416. [Google Scholar]
- Murmanis, L.; Sachs, I.B. Seasonal development of secondary xylem in Pinus strobus L. Wood Sci. Technol. 1969, 3, 177–193. [Google Scholar] [CrossRef]
- Liese, W. The warty layer. In Cellular Ultrastructure of Woody Plants; Côté, W.A., Ed.; Syracuse University Press: New York, NY, USA, 1965. [Google Scholar]
- Ohtani, J.; Meylan, B.A.; Butterfield, B.G. Occurrence of warts in the vessel elements and fibres of New Zealand woods. NZ J. Bot. 1983, 21, 359–372. [Google Scholar] [CrossRef]
- Kuo, M.; Manwiller, F.G. Morphological and chemical characteristics of the warty layer in red pine (Pinus resinosa AIT.). Wood Fiber Sci. 1986, 18, 239–247. [Google Scholar]
- Singh, A.P.; Nilsson, T.; Daniel, G. Alstonia scholaris vestures are resistant to degradation by tunnelling bacteria. IAWA J. 1993, 14, 119–126. [Google Scholar] [CrossRef]
- Kim, J.S.; Awano, T.; Yoshinaga, A.; Takabe, K. Ultrastructure of the innermost surface of differentiating normal and compression wood tracheids as revealed by field emission scanning electron microscopy. Planta 2012, 235, 1209–1219. [Google Scholar] [CrossRef]
- Hepler, P.K.; Fosket, D.E.; Newcomb, E.H. Lignification during secondary wall formation in Coleus: An electron microscopic study. Am. J. Bot. 1970, 57, 85–96. [Google Scholar] [CrossRef]
- Maurer, A.; Fengel, D.A. A process for improving the quality and lignin staining of ultrathin sections from wood tissue. Holzforschung 1990, 44, 453–460. [Google Scholar] [CrossRef]
- Donaldson, L.A. Lignin distribution during latewood formation in Pinus radiata D. Don. IAWA Bull. N.S. 1992, 13, 381–387. [Google Scholar] [CrossRef]
- Bland, D.E.; Foster, R.C.; Logan, A.F. The mechanism of permanganate and osmium tetroxide fixation and the distribution of the lignin in the cell wall of Pinus radiata. Holzforschung 1971, 25, 137–143. [Google Scholar] [CrossRef]
- Kanabayashi, T.; Miyafuji, H. Topochemical and morphological characterisation of wood cell wall treated with the ionic liquid, 1-ethylpyridium bromide. Planta 2015, 242, 509–518. [Google Scholar] [CrossRef]
- Kim, J.S.; Awano, T.; Yoshinaga, A.; Takabe, K. Distribution of hemicelluloses in warts and warty layer in normal and compression wood tracheids of Cryptomeria japonica. J. Korean Wood Sci. Technol. 2011, 39, 420–428. [Google Scholar] [CrossRef]
- Jansen, S.; Smets, E.; Baas, P. Vestures in woody plants: A review. IAWA J. 1998, 19, 347–382. [Google Scholar] [CrossRef]
- Jansen, S.; Baas, P.; Gasson, P.; Smets, E. Vestured pits: Do they promote safer water transport? Int. J. Plant Sci. 2003, 164, 405–413. [Google Scholar] [CrossRef]
- Bailey, I.W. The cambium and its derivative tissues. VIII. Structure, distribution, and diagnostic significance of vestured pits in dicotyledons. J. Arnold Arbor. 1933, 14, 259–273. [Google Scholar] [CrossRef]
- Wardrop, A.B.; Ingle, H.D.; Davies, G.W. Nature of vestured pits in angiosperms. Nature 1963, 197, 202–203. [Google Scholar] [CrossRef]
- Parameswaran, N.; Liese, W. Vestured pits in vessels and tracheids of Gnetum. IAWA Bull. 1974, 4, 3–7. [Google Scholar]
- Singh, A.P. On the occurrence of anamalous tubular structures in the vestured pits of petiolar xylem in Eucalyptus delegatensis. IAWA Bull. N.S. 1983, 4, 239–243. [Google Scholar] [CrossRef]
- Watanabe, Y.; Sano, Y.; Asada, T.; Funada, R. Histochemical study of the chemical composition of vestured pits in two species of Eucalyptus. IAWA J. 2006, 27, 33–43. [Google Scholar] [CrossRef]
- Scurfield, G.; Silva, S.R. The vestured pits of Eucalyptus regnans F. Muell.: A study using scanning electron microscopy. Bot. J. Lin. 1970, 63, 313–320. [Google Scholar] [CrossRef]
- Ohtani, J.; Ishida, S. Study on the pit of wood cells using scanning electron microscopy. Vestured pits in Japanese dicotyledonous woods. Res. Bull. Coll. Exp. For. Hokkaido Univ. 1976, 33, 407–436. [Google Scholar]
- Wu, J.; Ohtani, J.; Fukazawa, K. SEM observations on vestured pits in some Yunnan hardwoods. Res. Bull. Coll. Exp. For. Hokkaido Univ. 1988, 45, 789–806. [Google Scholar]
- Donaldson, L.A.; Singh, A.P. Ultrastructure of Terminalia wood from an ancient Polynesian canoe. IAWA Bull. N.S. 1990, 11, 195–202. [Google Scholar] [CrossRef]
- Singh, A.P.; Kim, Y.S.; Wi, S.G. Inhomogeneity in the composition of vesture walls in an archaeological wood. IAWA J. 2002, 23, 77–82. [Google Scholar] [CrossRef]
- Taylor, A.M.; Gartner, B.L.; Morrell, J.J. Heartwood formation and natural durability—A review. Wood Fiber Sci. 2002, 34, 587–611. [Google Scholar]
- Magel, E.A. Biochemistry and physiology of heartwood formation. In Cell and Molecular Biology of Wood Formation; Savidge, R., Barnett, J., Napier, R., Eds.; Bios Scientific Publishers Ltd.: Oxford, UK, 2000; pp. 363–376. [Google Scholar]
- Singh, T.; Singh, A.P. A review on natural products as wood protectant. Wood Sci. Technol. 2012, 46, 851–870. [Google Scholar] [CrossRef]
- Nilsson, T.; Singh, A.P.; Daniel, G. Ultrastructure of the attack of Eusideroxylon zwageri wood by tunnelling bacteria. Holzforschung 1992, 46, 361–367. [Google Scholar] [CrossRef]
- Kleist, G.; Schmitt, U. Evidence of accessory compounds in vessel walls of Sapelli heartwood (Entandrophragma cylindrica). Holz Roh Werkst. 1999, 57, 93–95. [Google Scholar] [CrossRef]
- Kim, Y.S.; Singh, A.P.; Wong, A.H.H.; Eom, T.-J.; Lee, K.H. Micromorphological and chemical characteristics of cengal (Neobalanocarpus heimii) heartwood decayed by soft rot fungi. J. Korean Wood Sci. Technol. 2006, 34, 68–77. [Google Scholar]
- Ajlan, N.; Jusoh, I.; Assim, Z. Role of extractives in the durability of Neobalanocarpus heimii and Shorea falcifera heartwoods. ASM Sc. J. 2019, 12, 1–10. [Google Scholar]
- Chen, J.; Liu, S.; Yin, L.; Cao, H.; Xi, G.; Zhang, Z.; Liu, J.; Luo, R.; Han, L.; Yin, Y.; et al. Non-destructive preservation state estimation of waterlogged archaeological wooden artefacts. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2023, 285, 121840. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Feng, X.; Liu, X. Advances in historical wood consolidation and conservation materials. BioResources 2023, 18, 6680–6723. [Google Scholar] [CrossRef]
- Walsh-Korb, Z.; Stelzner, I.; dos Santos Gabriel, J.; Eggert, G.; Alvérous, L. Morphological study of bio-based polymers in the consolidation of waterlogged wooden objects. Materials 2022, 15, 681. [Google Scholar] [CrossRef]
- Broda, M.; Jakes, J.E.; Li, L.; Antipova, O.A. Archaeological wood conservation with selected organosilicon compounds studied by XFM and nanoindentation. Wood Sci. Technol. 2023, 57, 1277–1298. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Composition (%) | Softwood (SW) | Hardwood (HW) | ||
|---|---|---|---|---|
| Normal Wood | Compression Wood | Normal Wood | Tension Wood | |
| Cellulose | 37–43 | 29–31 | 39–45 | 50–65 |
| Galactoglucomannan (SW) Glucomannan (HW) | 15–20 | 9–12 | 2–5 | 2–4 |
| Arabinoglucuronoxylan (SW) Glucuronoxylan (HW) | 5–10 | 6–8 | 15–30 | 16–23 |
| Galactan | 9–11 | 0–10 | ||
| Lignin | 25–33 | 37–40 | 20–25 | 16–20 |
| Extractives | 2–5 | 2–5 | 2–4 | 2–4 |
| Microscopy | Conventional LM | Conventional SEM | Conventional TEM | CLSM |
|---|---|---|---|---|
| Features |
|
|
|
|
| Common sample preparation methods |
|
|
|
|
| Staining or coating |
|
|
|
|
| Advanced applications |
|
|
|
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, A.P.; Kim, J.S.; Möller, R.; Chavan, R.R.; Kim, Y.S. The Pivotal Role of Microscopy in Unravelling the Nature of Microbial Deterioration of Waterlogged Wood: A Review. Forests 2024, 15, 889. https://doi.org/10.3390/f15050889
Singh AP, Kim JS, Möller R, Chavan RR, Kim YS. The Pivotal Role of Microscopy in Unravelling the Nature of Microbial Deterioration of Waterlogged Wood: A Review. Forests. 2024; 15(5):889. https://doi.org/10.3390/f15050889
Chicago/Turabian StyleSingh, Adya P., Jong Sik Kim, Ralf Möller, Ramesh R. Chavan, and Yoon Soo Kim. 2024. "The Pivotal Role of Microscopy in Unravelling the Nature of Microbial Deterioration of Waterlogged Wood: A Review" Forests 15, no. 5: 889. https://doi.org/10.3390/f15050889
APA StyleSingh, A. P., Kim, J. S., Möller, R., Chavan, R. R., & Kim, Y. S. (2024). The Pivotal Role of Microscopy in Unravelling the Nature of Microbial Deterioration of Waterlogged Wood: A Review. Forests, 15(5), 889. https://doi.org/10.3390/f15050889