
Effects of Water Control and Nitrogen Addition on Functional Traits and Rhizosphere Microbial Community Diversity of Haloxylon ammodendron Seedlings

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Experimental Design
2.2.2. Sample Collection and Processing
2.2.3. 16s rDNA Sequencing
2.2.4. Calculation of Rhizosphere Soil Microbial Community Diversity
2.3. Data Analysis
3. Results
3.1. Differences of Aboveground and Underground Functional Traits of H. ammodendron Seedlings
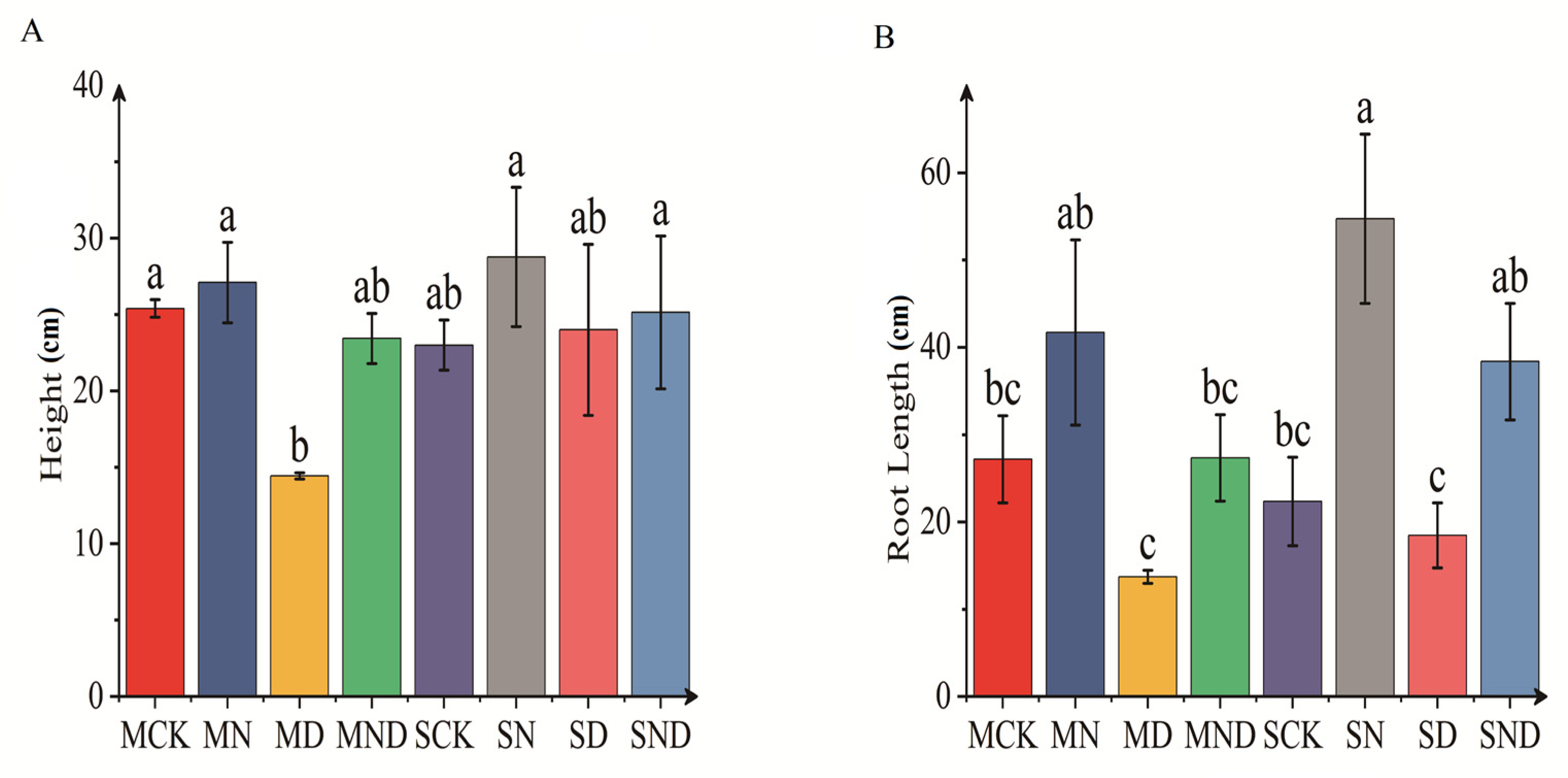
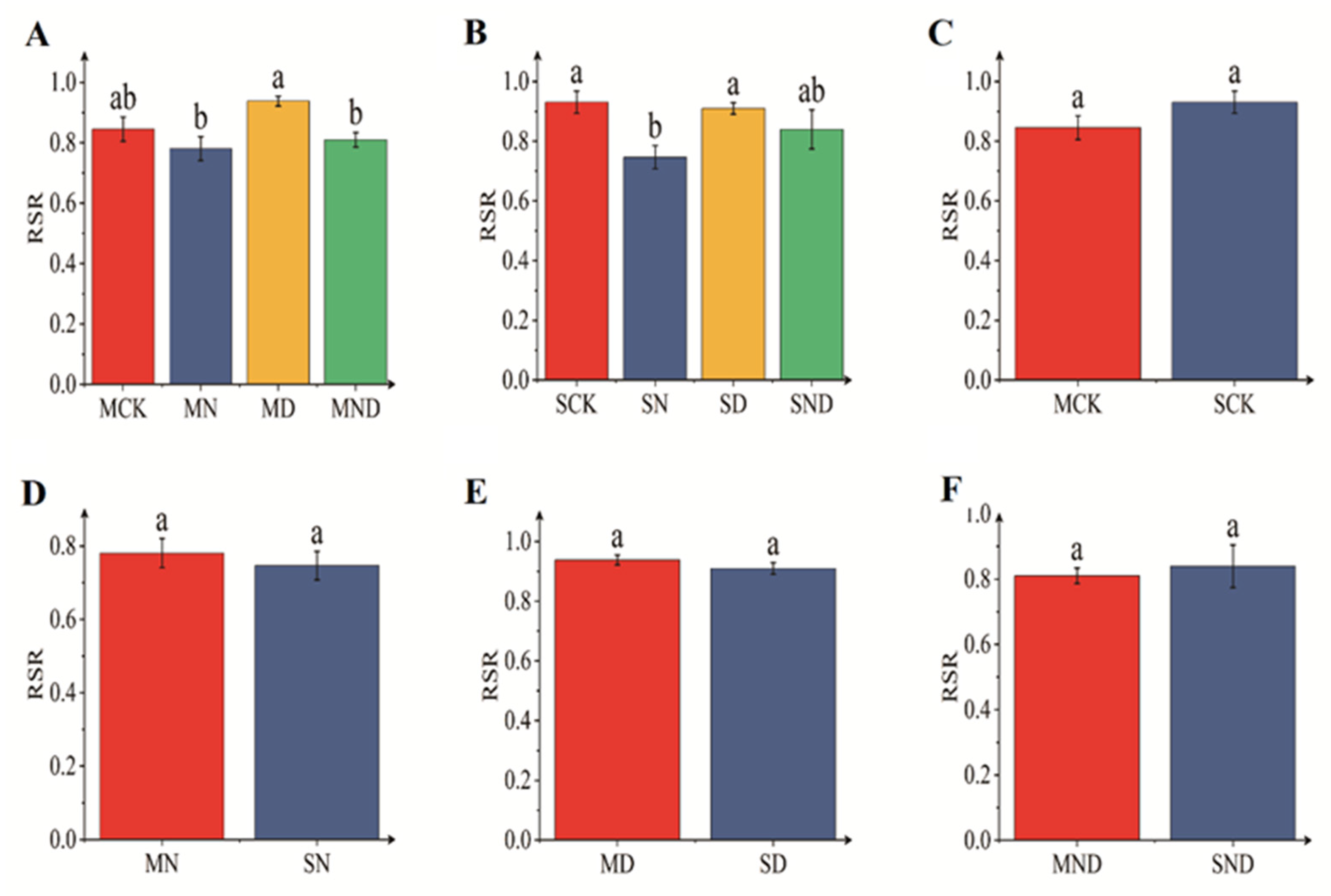
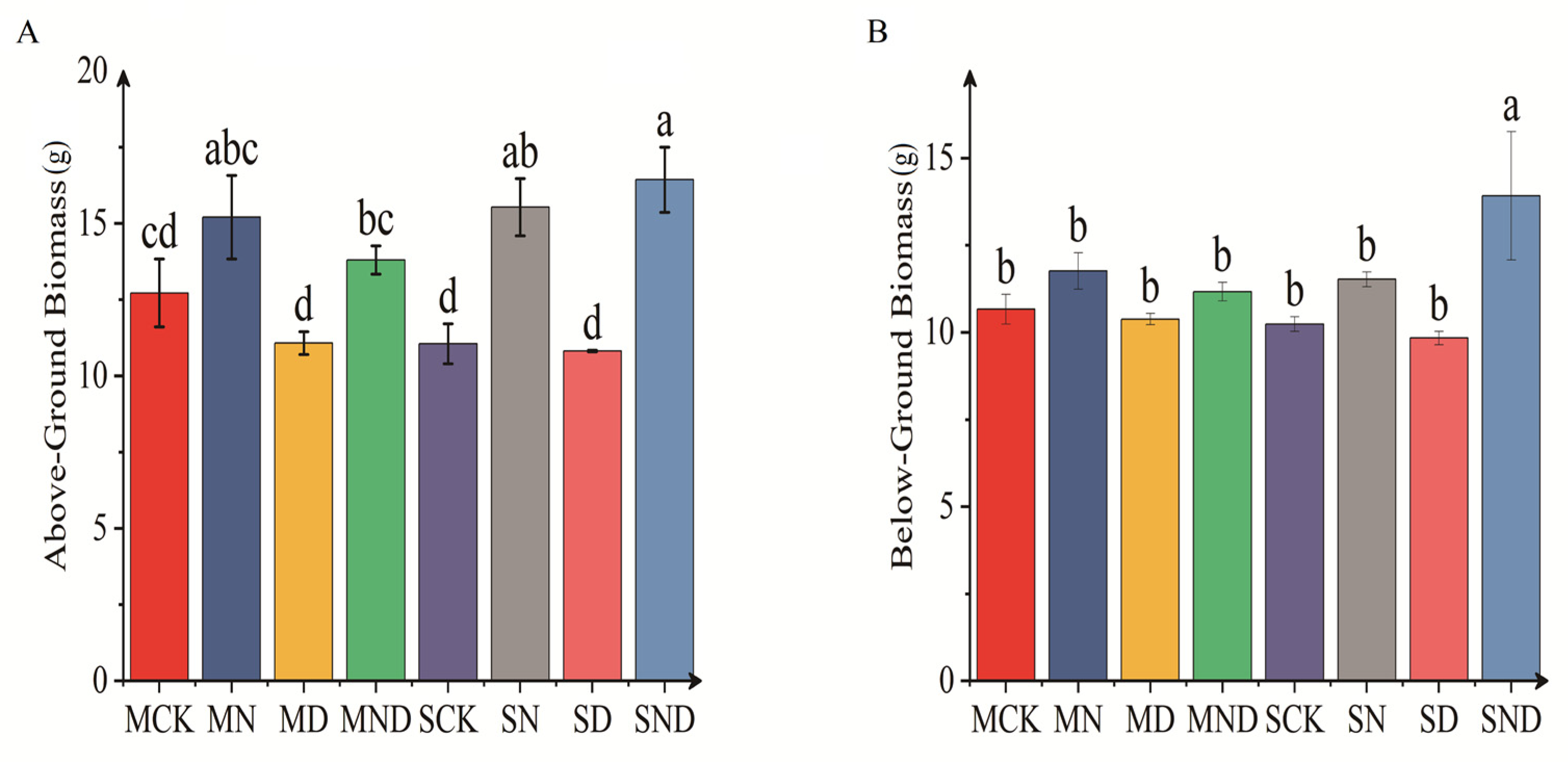
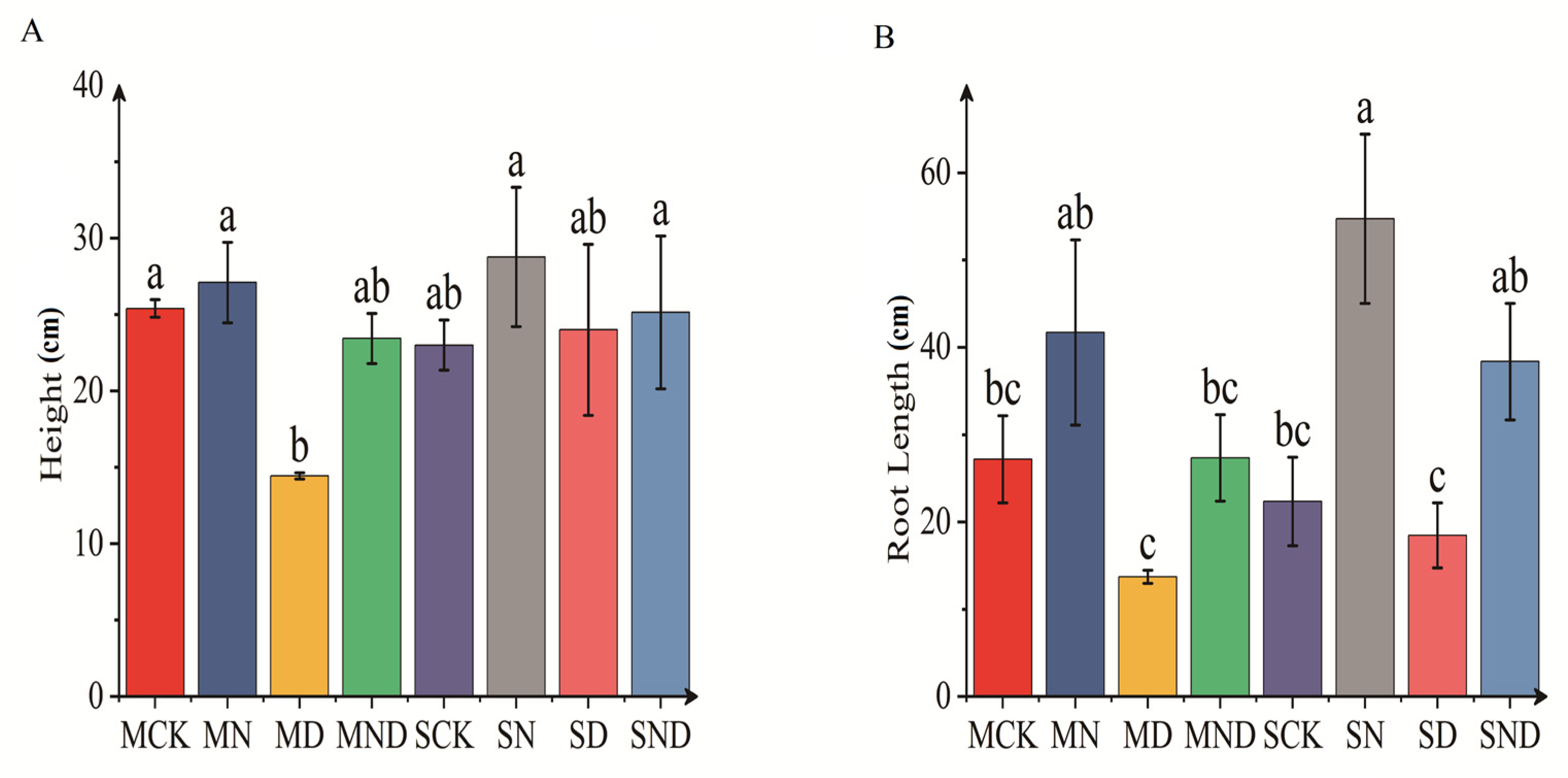
3.1.1. Differences in Aboveground and Underground Morphological Indexes of H. ammodendron Seedlings
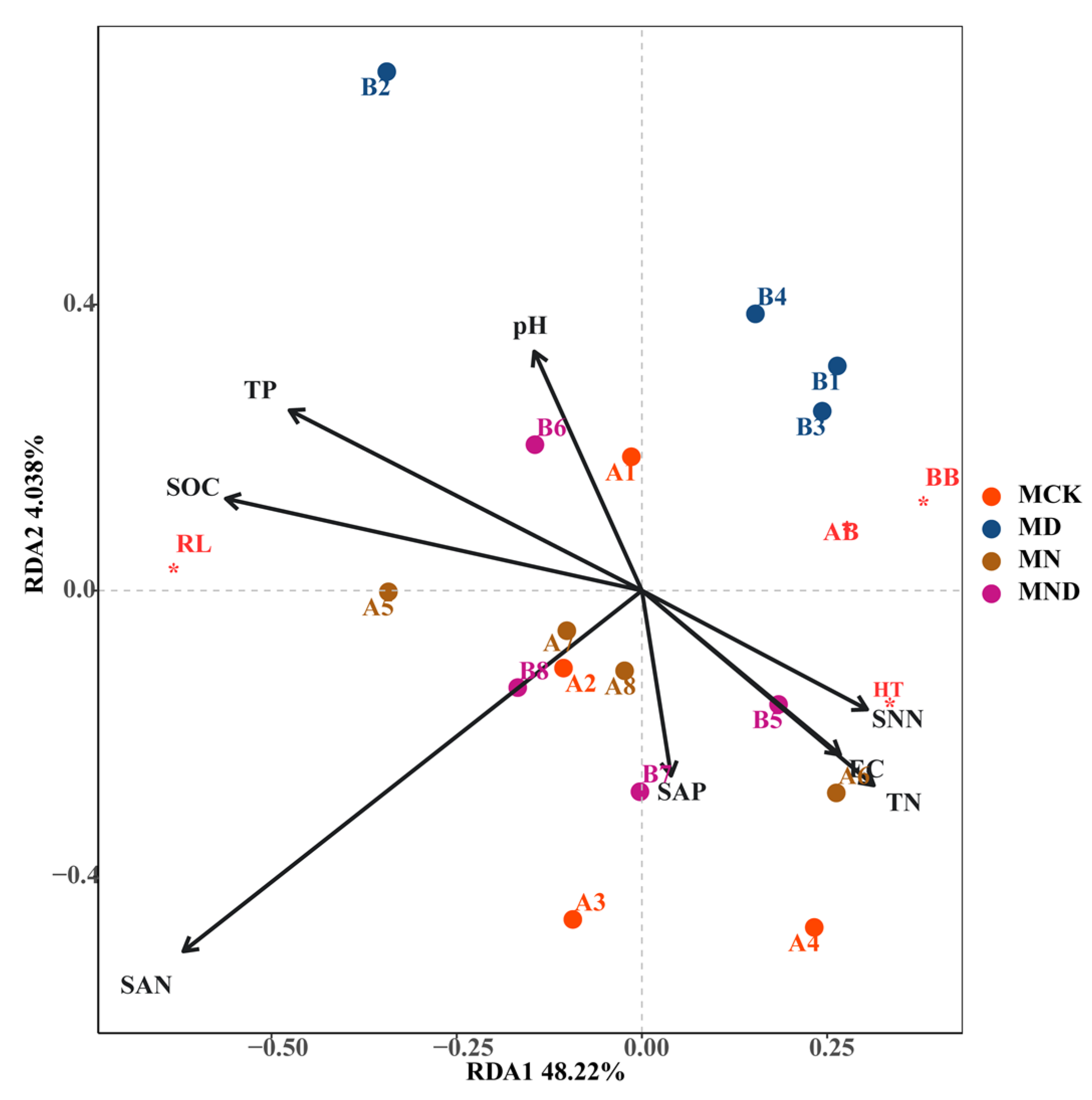
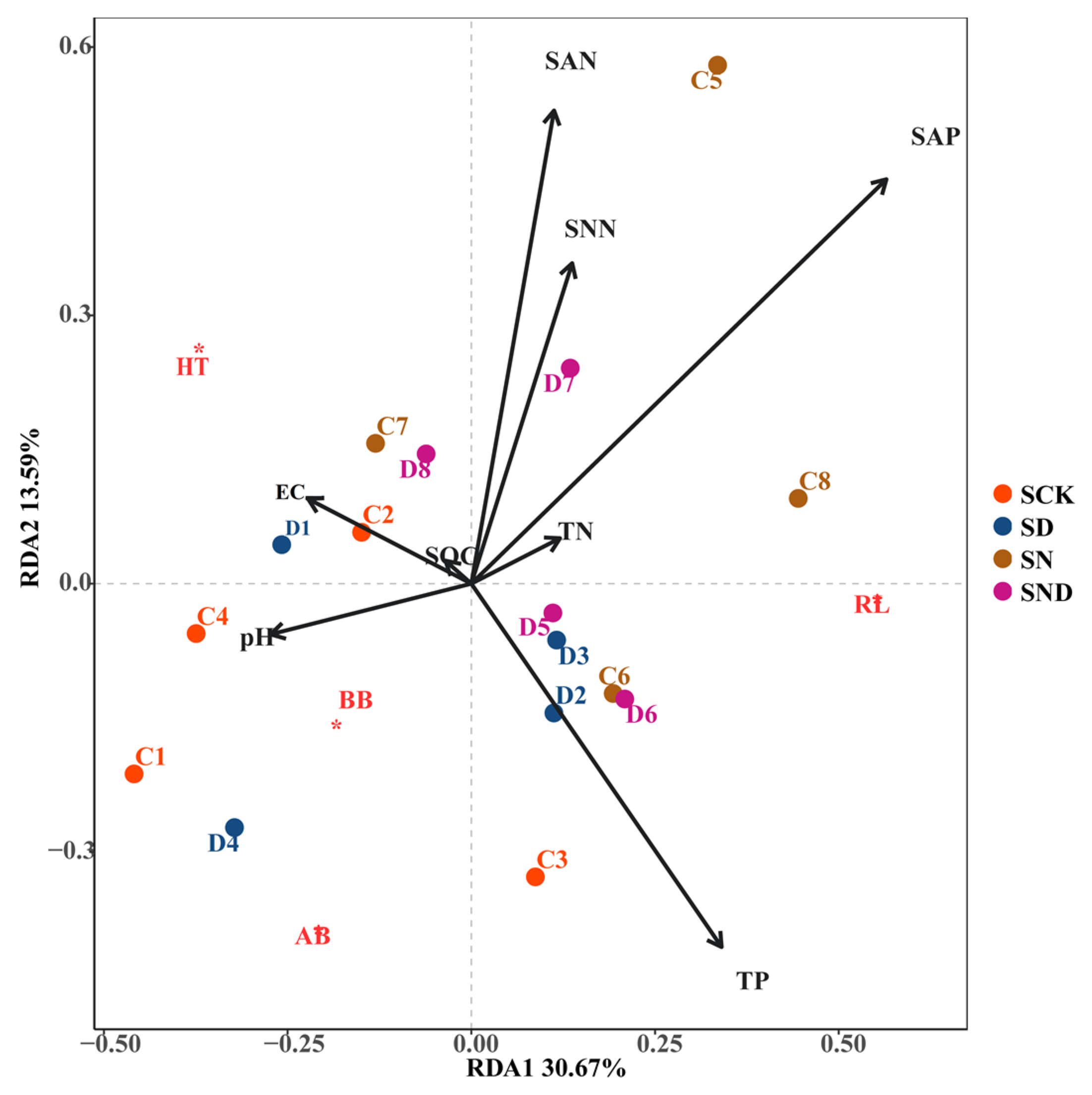
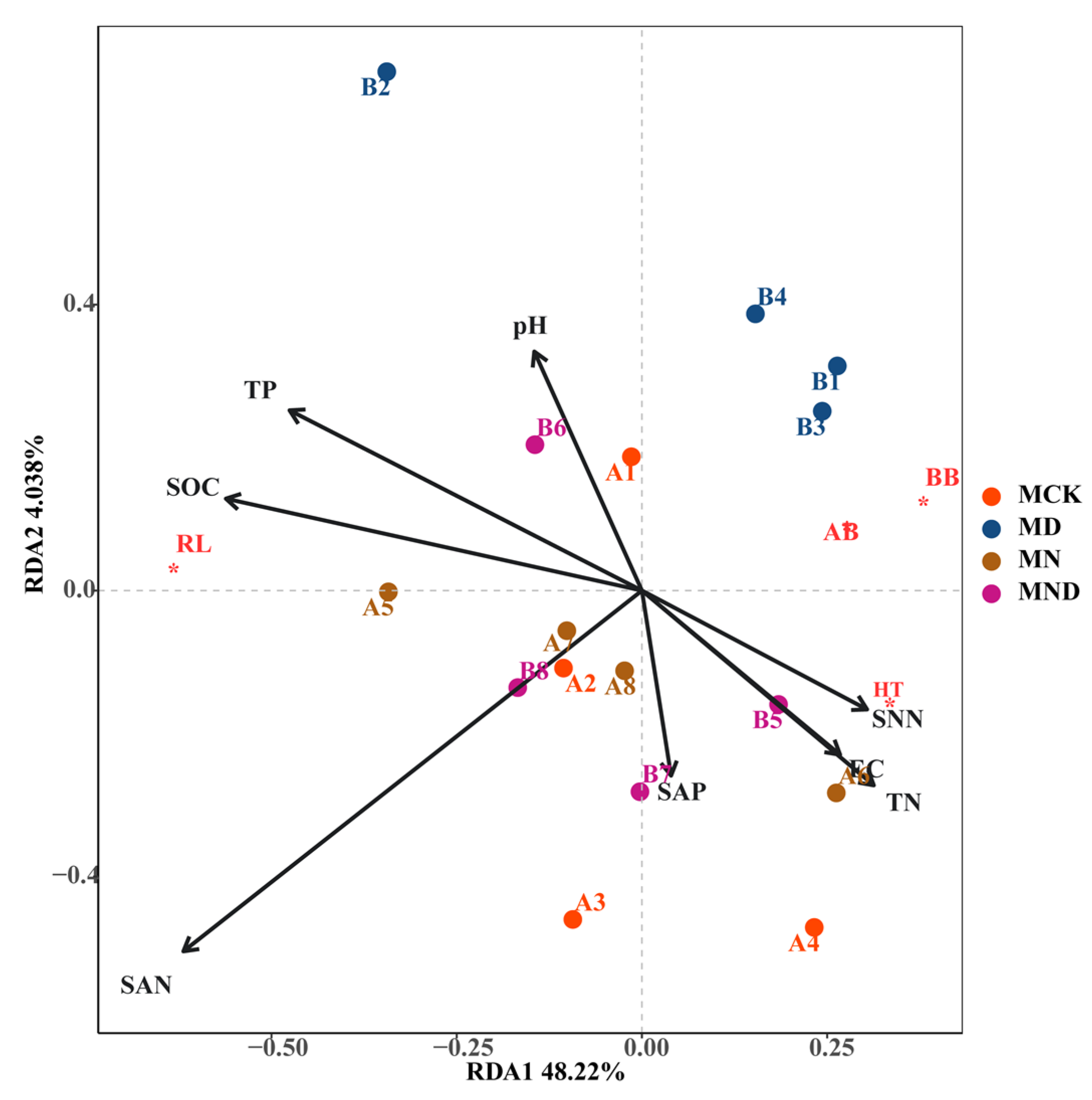
3.1.2. Correlation Analysis between Plant Aboveground and Underground Traits and Environmental Factors
3.2. Diversity of Rhizosphere Bacterial Community in H. ammodendron Seedlings
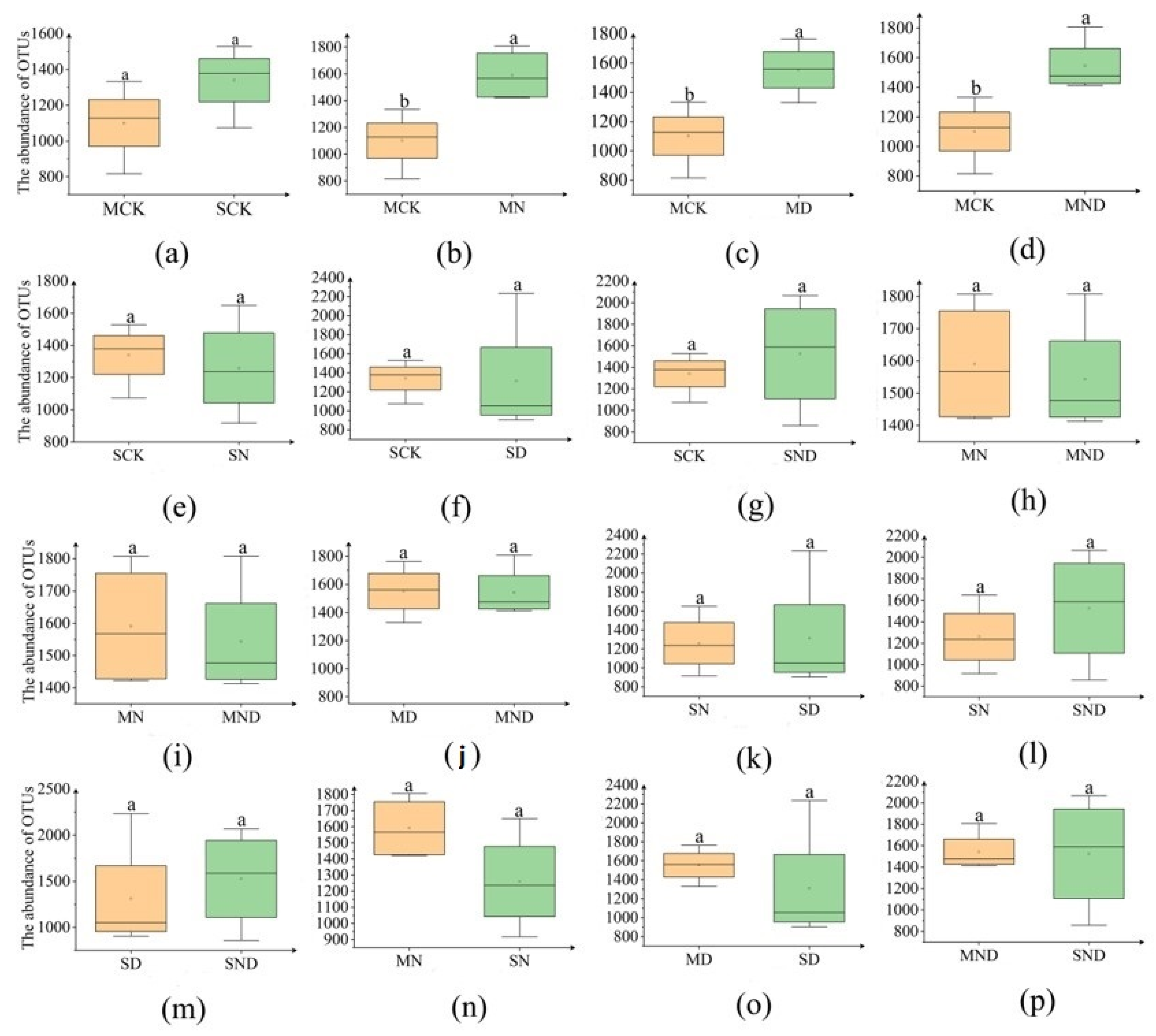
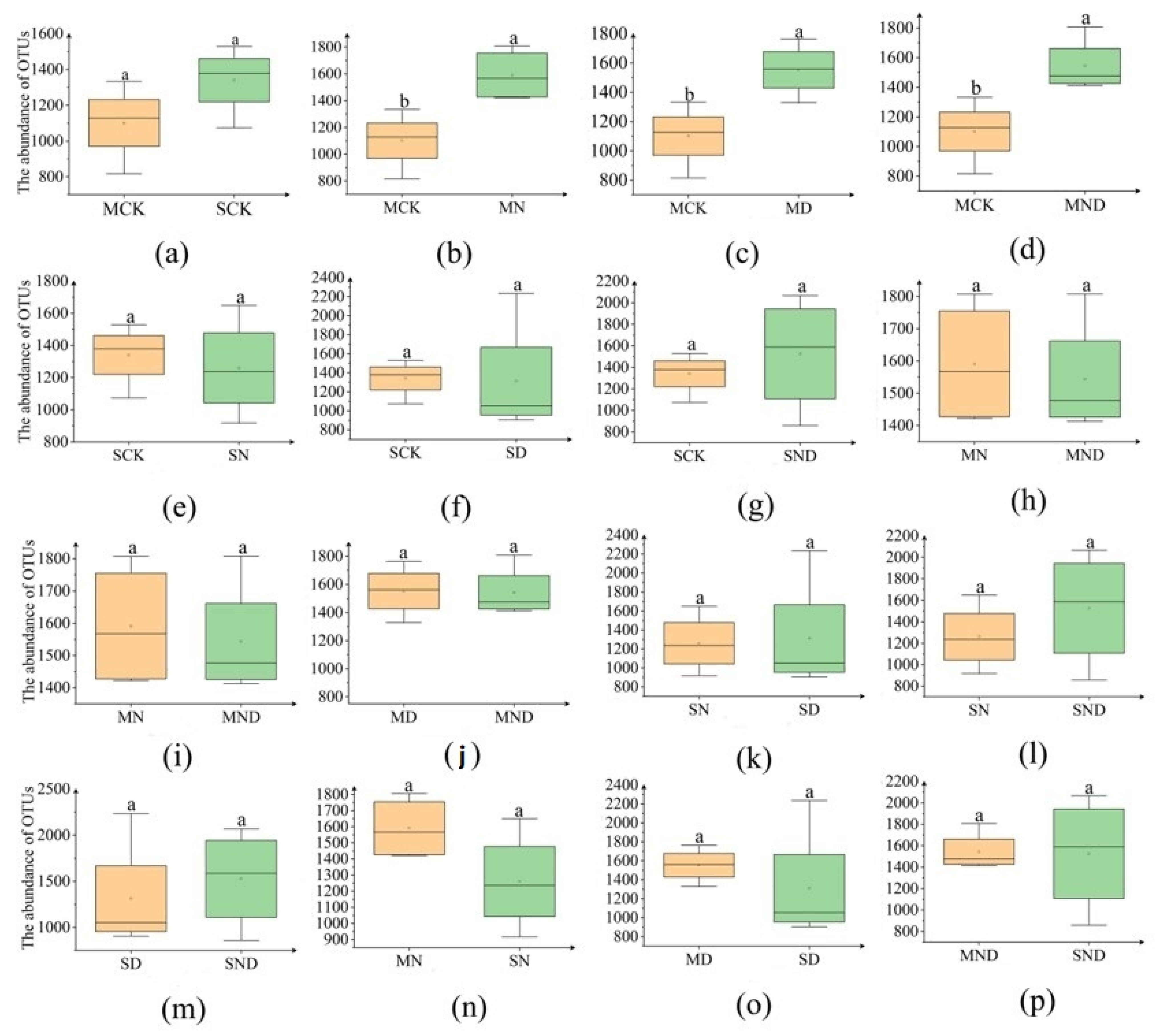
3.2.1. Differences in Rhizosphere Bacterial Community Abundance of H. ammodendron Seedlings
3.2.2. Alpha Diversity of Bacterial Community in H. ammodendron Seedlings
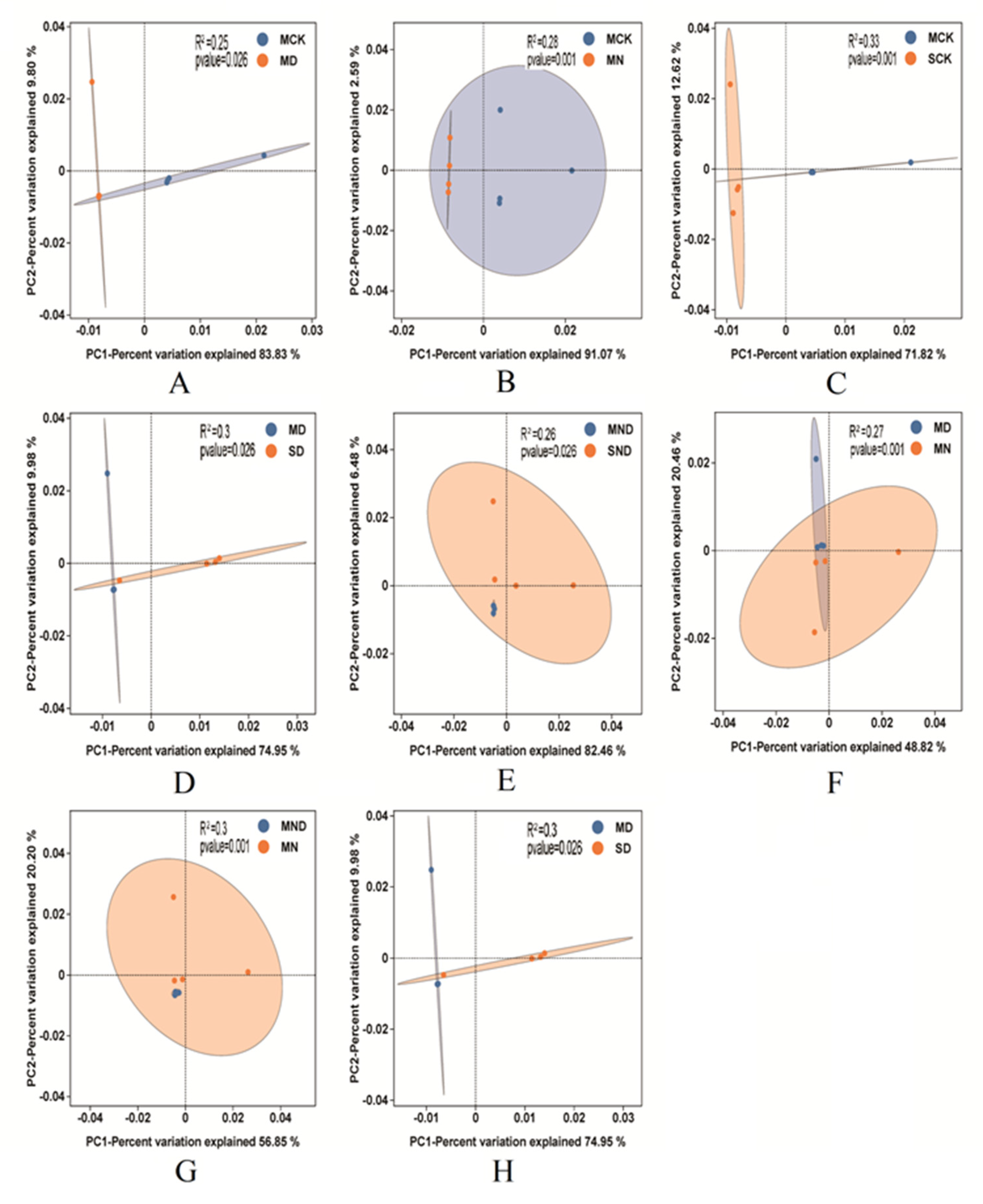
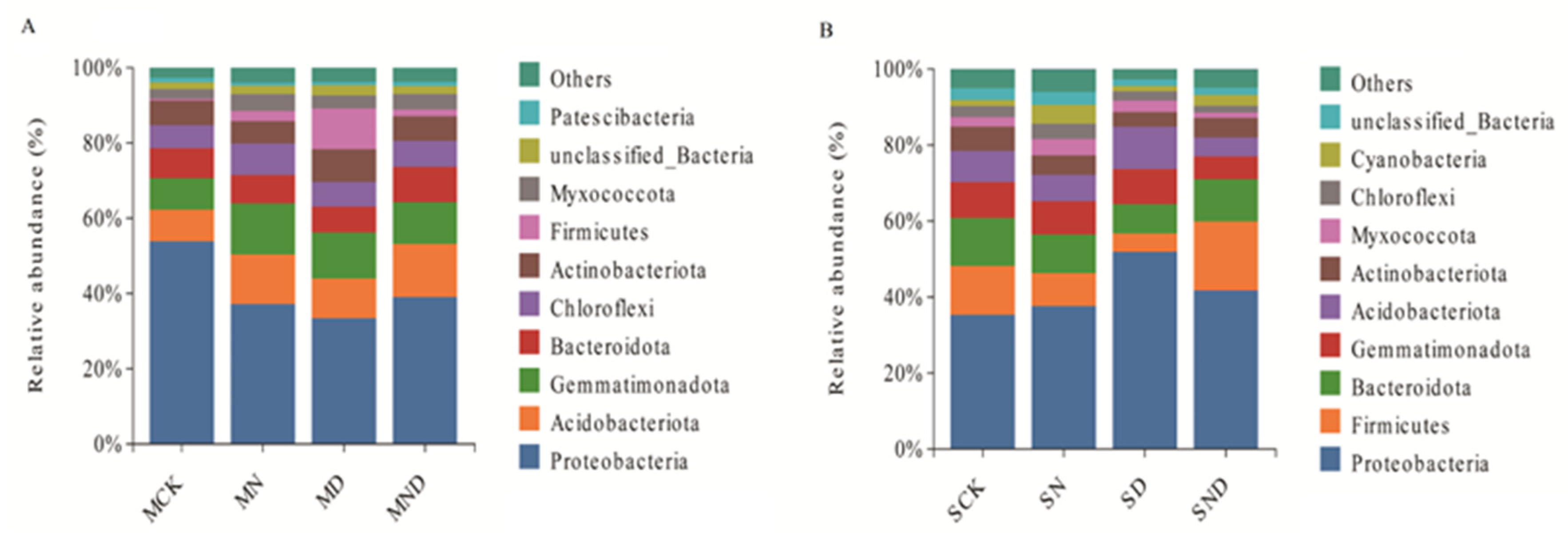
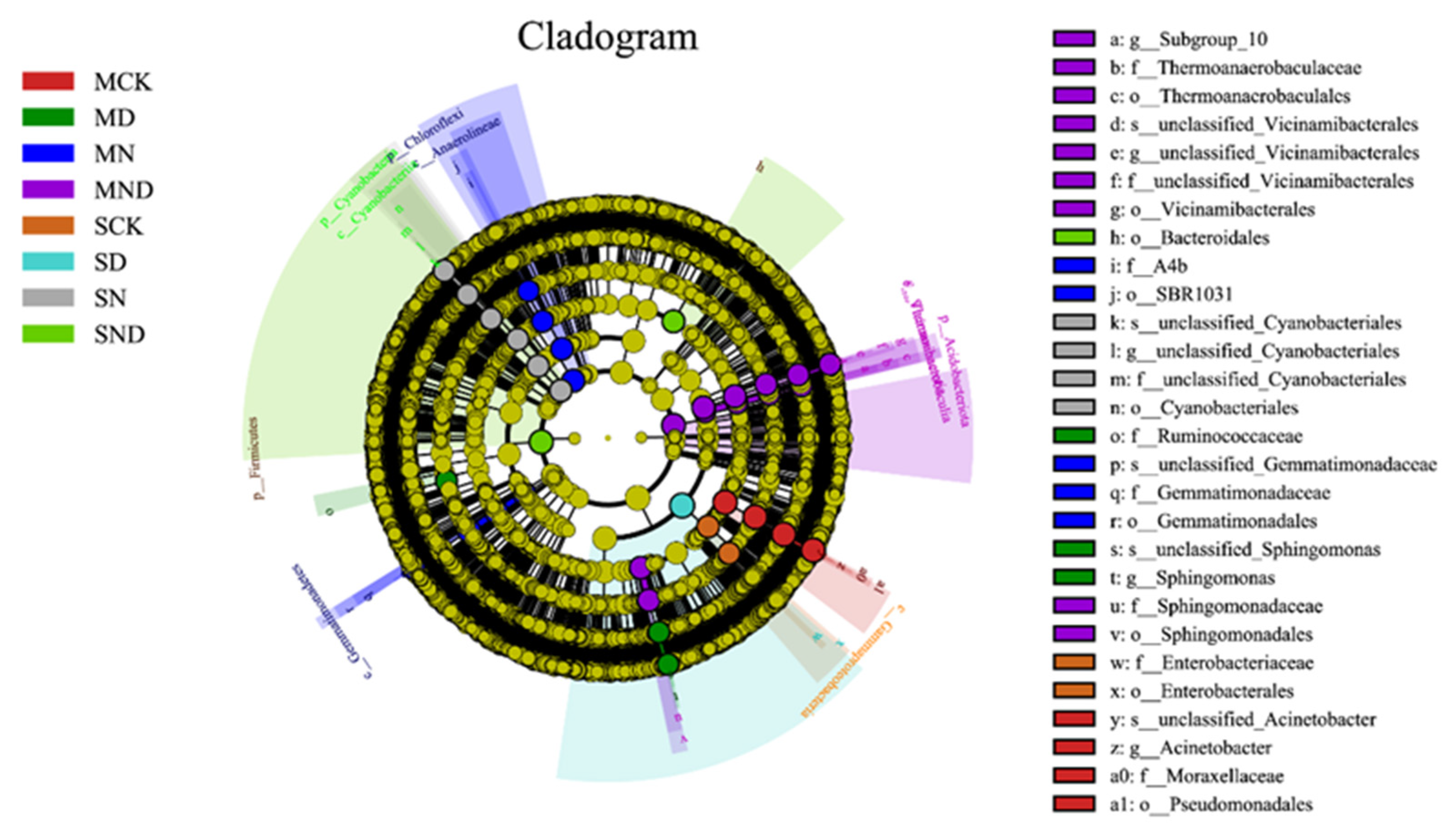
3.2.3. Changes in Soil Bacterial Community Composition under Different Treatments
4. Discussion
4.1. Physiological and Ecological Changes in H. ammodendron Seedlings under Nitrogen and Water Coupling
4.2. Changes in Rhizosphere Microbial Community in H. ammodendron Seedlings
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wang, J.; Duan, B.; Zhang, Y. Effects of Experimental Warming on Growth, Biomass Allocation, and Needle Chemistry of Abies Faxoniana in Even-Aged Monospecific Stands. Plant Ecol. 2012, 213, 47–55. [Google Scholar] [CrossRef]
- Yang, X.; Long, Y.; Sarkar, B.; Li, Y.; Lü, G.; Ali, A.; Yang, J.; Cao, Y.-E. Influence of Soil Microorganisms and Physicochemical Properties on Plant Diversity in an Arid Desert of Western China. J. For. Res. 2021, 32, 2645–2659. [Google Scholar] [CrossRef]
- Mokany, K.; Raison, R.J.; Prokushkin, A.S. Critical Analysis of Root: Shoot Ratios in Terrestrial Biomes. Glob. Chang. Biol. 2005, 12, 84–96. [Google Scholar] [CrossRef]
- Singla-Pareek, S.L. Transcription Factors and Plants Response to Drought Deficit: Current Understanding and Future Directions. Front. Plant Sci. 2016, 7, 1029. [Google Scholar] [CrossRef]
- Xu, L.; Naylor, D.; Dong, Z.; Simmons, T.; Pierroz, G.; Hixson, K.K.; Kim, Y.-M.; Zink, E.M.; Engbrecht, K.M.; Wang, Y.; et al. Drought Delays Development of the Sorghum Root Microbiome and Enriches for Monoderm Bacteria. Proc. Natl. Acad. Sci. USA 2018, 115, E4284–E4293. [Google Scholar] [CrossRef]
- Fang, S.; Liang, X. Response Mechanisms of Plants Under Saline-Alkali Deficit. Front. Plant Sci. 2021, 72, 673–689. [Google Scholar] [CrossRef]
- Fang, Y.; Xiong, L. General Mechanisms of Drought Response and Their Application in Drought Resistance Improvement in Plants. Front. Plant Sci. 2021, 12, 667458. [Google Scholar] [CrossRef]
- Etesami, H. Bacterial Mediated Alleviation of Heavy Metal Deficit and Decreased Accumulation of Metals in Plant Tissues_ Mechanisms and Future Prospects. Ecotoxicol. Environ. Saf. 2018, 147, 175–191. [Google Scholar] [CrossRef]
- Li, W.J.; Wang, J.L.; Jiang, L.M.; Lv, G.H.; Hu, D.; Wu, D.Y.; Yang, X.D. Rhizosphere effect and water constraint jointly determined the roles of microorganism in soil phosphorus cycling in arid desert regions. CATENA 2023, 222, 106809. [Google Scholar] [CrossRef]
- Huang, Z.; Zhang, X.; Zheng, G.; Gutterman, Y. Influence of Light, Temperature, Salinity and Storage on Seed Germination of Haloxylon ammodendron. J. Arid Environ. 2003, 55, 453–464. [Google Scholar] [CrossRef]
- Sheng, Y.A.N.; Zheng, W.H.; Pei, K.Q.; Ma, K.P. Genetic Variation within and Among Populations of a Dominant Desert Tree H. ammodendron (Amaranthaceae) in China. Ann. Bot. 2005, 96, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Kour, D.; Yadav, A.N. Bacterial Mitigation of Drought Deficit in Plants: Current Perspectives and Future Challenges. Curr. Microbiol. 2022, 79, 248. [Google Scholar] [CrossRef] [PubMed]
- Hassan, F.A.S.; Ali, E.F.; Mahfouz, S.A. Comparison between different fertilization sources, irrigation frequency and their combinations on the growth and yield of coriander plant. Aust. J. Basic Appl. Sci. 2012, 6, 600–615. [Google Scholar]
- Boyle, S.A.; Yarwood, R.R.; Bottomley, P.J.; Myrold, D.D. Bacterial and Fungal Contributions to Soil Nitrogen Cycling under Douglas FIr and Red Alder at Two Sites in Oregon. Soil Biol. 2008, 40, 443–450. [Google Scholar] [CrossRef]
- He, K.N.; Wang, H.; Wang, W.L.; Zhang, T. Salinity Effects on Germination and Plant Growth of H. ammodendron at Qaidam Basin. J. Phys. Conf. Ser. 2020, 1578, 012234. [Google Scholar] [CrossRef]
- Vos, M.; Wolf, A.B.; Jennings, S.J.; Kowalchuk, G.A. Micro-Scale Determinants of Bacterial Diversity in Soil. FEMS Microbiol. Rev. 2013, 37, 936–954. [Google Scholar] [CrossRef]
- Treseder, K.K. Nitrogen Additions and Microbial Biomass: A Meta-analysis of Ecosystem Studies. Ecol. Lett. 2008, 11, 1111–1120. [Google Scholar] [CrossRef]
- Gutknecht, J.M.; Field, C.B.; Balser, T.C. Microbial Communities and Their Responses to Simulated Global Change Fluctuate Greatly Over Multiple Years. Glob. Chang. Biol. 2012, 18, 2256–2269. [Google Scholar] [CrossRef]
- Edwards, J.; Johnson, C.; Santos-Medellín, C.; Lurie, E.; Podishetty, N.K.; Bhatnagar, S.; Eisen, J.A.; Sundaresan, V. Structure, Variation, and Assembly of the Root-Associated Microbiomes of Rice. Proc. Natl. Acad. Sci. USA 2015, 112, E911–E920. [Google Scholar] [CrossRef]
- Turner, B.L.; Driessen, J.P.; Haygarth, P.M.; Mckelvie, I.D. Potential Contribution of Lysed Bacterial Cells to Phosphorus Solubilisation in Two Rewetted Australian Pasture Soils. Soil Biol. Biochem. 2003, 35, 187–189. [Google Scholar] [CrossRef]
- Etzold, S.; Ferretti, M.; Reinds, G.J.; Solberg, S.; Gessler, A.; Waldner, P.; Schaub, M.; Simpson, D.; Benham, S.; Hansen, K.; et al. Nitrogen Deposition Is the Most Important Environmental Driver of Growth of Pure, Even-Aged and Managed European Forests. For. Ecol. Manag. 2020, 458, 117762. [Google Scholar] [CrossRef]
- Valliere, J.M.; Allen, E.B. Interactive Effects of Nitrogen Deposition and Drought-Deficit on Plant-Soil Feedbacks of Artemisia Californica Seedlings. Plant Soil. 2016, 403, 277–290. [Google Scholar] [CrossRef]
- Li, Y.; Hu, W.; Zou, J.; He, J.; Wang, Y.; Chen, B.; Meng, Y.; Wang, S.; Zhou, Z. Effects of Soil Drought on Cottonseed Kernel Carbohydrate Metabolism and Kernel Biomass Accumulation. Plant Physiol. Biochem. 2023, 195, 170–181. [Google Scholar] [CrossRef]
- Li, W.; Jin, C.; Guan, D.; Wang, Q.; Wang, A.; Yuan, F.; Wu, J. The Effects of Simulated Nitrogen Deposition on Plant Root Traits: A Meta-Analysis. Soil Biol. Biochem. 2015, 82, 112–118. [Google Scholar] [CrossRef]
- Zhang, X.; Lei, L.; Lai, J.; Zhao, H.; Song, W. Effects of Drought Deficit and Water Recovery on Physiological Responses and Gene Expression in Maize Seedlings. BMC Plant Biol. 2018, 18, 68. [Google Scholar] [CrossRef]
- Ravelo-Ortega, G.; Raya-González, J.; López-Bucio, J. Compounds from Rhizosphere Microbes That Promote Plant Growth. Curr. Opin. Plant Biol. 2023, 73, 102336. [Google Scholar] [CrossRef]
- Li, W.; Lei, X.; Zhang, R.; Cao, Q.; Yang, H.; Zhang, N.; Liu, S.; Wang, Y. Shifts in Rhizosphere Microbial Communities in Oplopanax Elatus Nakai Are Related to Soil Chemical Properties under Different Growth Conditions. Sci. Rep. 2022, 12, 11485. [Google Scholar] [CrossRef]
- Bach, E.M.; Williams, R.J.; Hargreaves, S.K.; Yang, F.; Hofmockel, K.S. Greatest Soil Microbial Diversity Found in Micro-Habitats. Soil Biol. Biochem. 2018, 118, 217–226. [Google Scholar] [CrossRef]
- Fuchslueger, L.; Bahn, M.; Fritz, K.; Hasibeder, R.; Richter, A. Experimental Drought Reduces the Transfer of Recently Fixed Plant Carbon to Soil Microbes and Alters the Bacterial Community Composition in a Mountain Meadow. New Phytol. 2014, 201, 916–927. [Google Scholar] [CrossRef]
- Kaurin, A.; Mihelič, R.; Kastelec, D.; Grčman, H.; Bru, D.; Philippot, L.; Suhadolc, M. Resilience of Bacteria, Archaea, Fungi and N-Cycling Microbial Guilds under Plough and Conservation Tillage, to Agricultural Drought. Soil Biol. Biochem. 2018, 120, 233–245. [Google Scholar] [CrossRef]
- Van, D.L.T.A.; Lilleskov, E.A.; Pregitzer, K.S.; Miller, R.M. Simulated Nitrogen Deposition Causes a Decline of Intra- and Extraradical Abundance of Arbuscular Mycorrhizal Fungi and Changes in Microbial Community Structure in Northern Hardwood Forests. Ecosystems 2010, 13, 683–695. [Google Scholar] [CrossRef]
- Singh, P.; Singh, R.K.; Zhou, Y.; Wang, J.; Jiang, Y.; Shen, N.; Wang, Y.; Yang, L.; Jiang, M. Unlocking the Strength of Plant Growth Promoting Pseudomonas in Improving Crop Productivity in Normal and Challenging Environments: A Review. J. Plant Interact. 2022, 17, 220–238. [Google Scholar] [CrossRef]
- Fitzpatrick, C.R.; Copeland, J.; Wang, P.W.; Guttman, D.S.; Kotanen, P.M.; Johnson, M.T.J. Assembly and Ecological Function of the Root Microbiome across Angiosperm Plant Species. Proc. Natl. Acad. Sci. USA 2018, 115, E1157–E1165. [Google Scholar] [CrossRef] [PubMed]
- Hillebrand, H. Meta-analysis on Pulse Disturbances Reveals Differences in Functional and Compositional Recovery across Ecosystems. Ecol. Lett. 2020, 23, 575–585. [Google Scholar] [CrossRef]
- Zhou, Z.; Wang, C.; Zheng, M.; Jiang, L.; Luo, Y. Patterns and Mechanisms of Responses by Soil Microbial Communities to Nitrogen Addition. Soil Biol. Biochem. 2017, 115, 433–441. [Google Scholar] [CrossRef]
- Zhang, T.; Chen, H.Y.H.; Ruan, H. Global Negative Effects of Nitrogen Deposition on Soil Microbes. ISME J. 2018, 12, 1817–1825. [Google Scholar] [CrossRef]
- Arnett, A.E.; Louda, S.M. Re-Test of Rhinocyllus Conicus Host Specificity, and the Prediction of Ecological Risk in Biological Control. Biol. Conserv. 2002, 106, 251–257. [Google Scholar] [CrossRef]
- Moreira, X.; Abdala-Roberts, L. Specificity and Context-Dependency of Plant–Plant Communication in Response to Insect Herbivory. Curr. Opin. Insect Sci. 2019, 32, 15–21. [Google Scholar] [CrossRef]
- Dowie, N.J.; Grubisha, L.C.; Trowbridge, S.M.; Klooster, M.R.; Miller, S.L. Variability of Ecological and Autotrophic Host Specificity in a Mycoheterotrophic System: Pterospora andromedea and Associated Fungal and Conifer Hosts. Fungal Ecol. 2016, 20, 97–107. [Google Scholar] [CrossRef]
- Zhou, Z.; Zheng, M.; Xia, J.; Wang, C. Nitrogen Addition Promotes Soil Microbial Beta Diversity, and the Stochastic Assembly. Sci. Total Environ. 2022, 806, 150569. [Google Scholar] [CrossRef]
- Cheng, J. Bacteroides Utilization for Dietary Polysaccharides and Their Beneficial Effects on Gut Health. Food Sci. Hum. Wellness 2022, 11, 1101–1110. [Google Scholar] [CrossRef]
- Hu, Y.; Chen, M.; Yang, Z.; Cong, M.; Zhu, X.; Jia, H. Soil Microbial Community Response to Nitrogen Application on a Swamp Meadow in the Arid Region of Central Asia. Front. Microbiol. 2022, 12, 797306. [Google Scholar] [CrossRef] [PubMed]
- Genderjahn, S.; Mashal, A.; Kai, M.; Fabian, H.; Dirk, W. Desiccation- and Saline-Tolerant Bacteria and Archaea in Kalahari Pan Sediments. Front. Microbiol. 2018, 9, 2082. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.B.; Peng, L.Z.; Li, Y.M.; Li, Y. Competitive interactions between two desert shrub seedings towards variation in soil nitrogen and phosphorus content. Arid. Land Geog. Raphy. 2018, 41, 83–91. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample ID | ACE Mean ± Se | Chao1 Mean ± Se | Simpson Mean ± Se | Shannon Mean ± Se |
|---|---|---|---|---|
| MCK | 1360.69 ± 97 a | 1361.36 ± 97 a | 0.992 ± 0.002 a | 8.893 ± 0.193 abc |
| MN | 1276.92 ± 152 a | 1280.19 ± 153 a | 0.989 ± 0.005 a | 8.649 ± 0.211 abc |
| MD | 1334.75 ± 317 a | 1338.82 ± 321 a | 0.975 ± 0.008 a | 8.264 ± 0.662 c |
| MND | 1571.13 ± 278 a | 1565.63 ± 276 a | 0.974 ± 0.02 a | 8.412 ± 0.708 bc |
| SCK | 1118.13 ± 111 a | 1119.48 ± 112 a | 0.971 ± 0.015 a | 8.234 ± 0.468 c |
| SN | 1636.32 ± 105 a | 1635.75 ± 106 a | 0.998 ± 0.001 a | 9.737 ± 0.07 a |
| SD | 1585.50 ± 91 a | 1593.35 ± 94 a | 0.996 ± 0.002 a | 9.542 ± 0.266 ab |
| SND | 1572.96 ± 94 a | 1576.39 ± 96 a | 0.998 ± 0.001 a | 9.688 ± 0.093 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, M.; Jiang, L.; Wu, D.; Li, W.; Yang, H.; He, X. Effects of Water Control and Nitrogen Addition on Functional Traits and Rhizosphere Microbial Community Diversity of Haloxylon ammodendron Seedlings. Forests 2023, 14, 1879. https://doi.org/10.3390/f14091879
Zhu M, Jiang L, Wu D, Li W, Yang H, He X. Effects of Water Control and Nitrogen Addition on Functional Traits and Rhizosphere Microbial Community Diversity of Haloxylon ammodendron Seedlings. Forests. 2023; 14(9):1879. https://doi.org/10.3390/f14091879
Chicago/Turabian StyleZhu, Menghao, Lamei Jiang, Deyan Wu, Wenjing Li, Huifang Yang, and Xuemin He. 2023. "Effects of Water Control and Nitrogen Addition on Functional Traits and Rhizosphere Microbial Community Diversity of Haloxylon ammodendron Seedlings" Forests 14, no. 9: 1879. https://doi.org/10.3390/f14091879
APA StyleZhu, M., Jiang, L., Wu, D., Li, W., Yang, H., & He, X. (2023). Effects of Water Control and Nitrogen Addition on Functional Traits and Rhizosphere Microbial Community Diversity of Haloxylon ammodendron Seedlings. Forests, 14(9), 1879. https://doi.org/10.3390/f14091879