Silicon Uptake and Phytolith Morphology in Dendrocalamus brandisii Seedling Leaf from Different Rearing Methods

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Preparation
2.2. Experimental Methods
2.2.1. Determination of Moisture Content
2.2.2. Determination of Ash and Silicon Content
2.2.3. Phytolith Extraction and Observation
3. Results
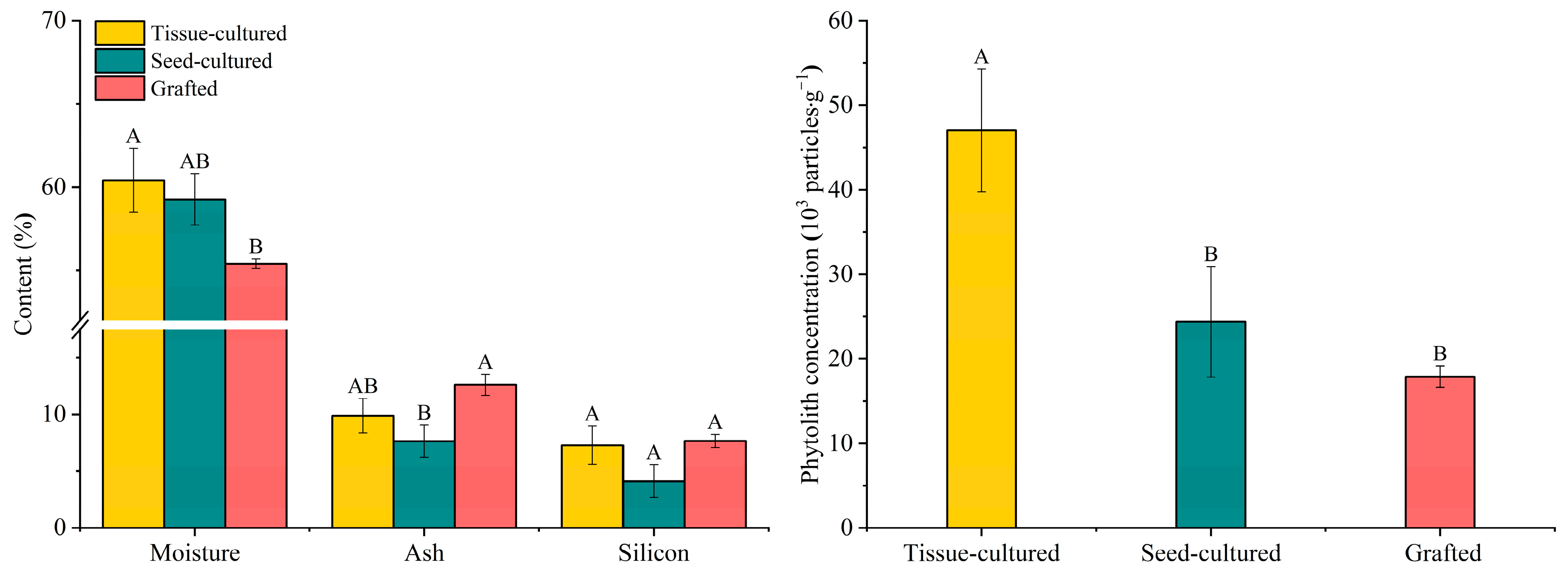
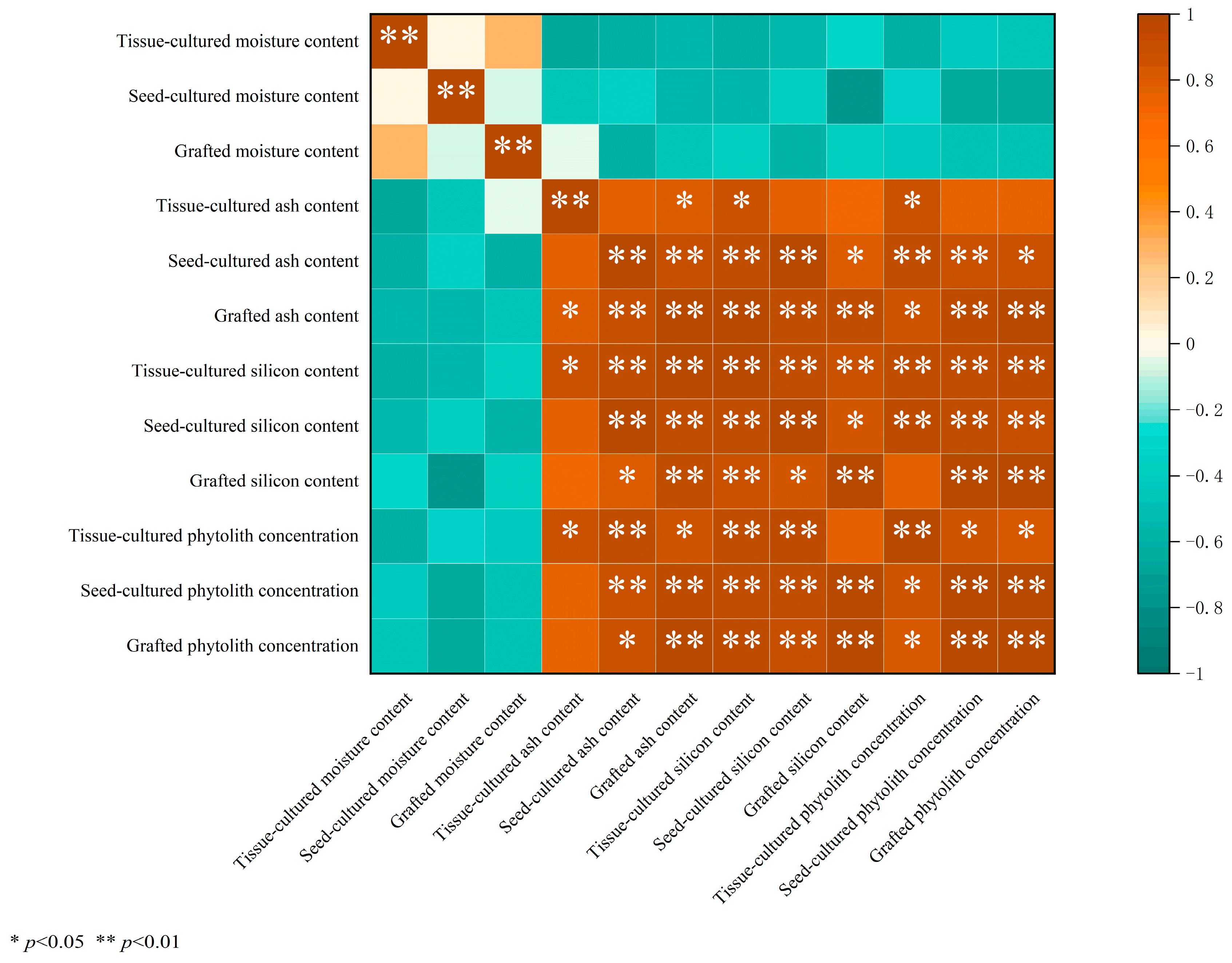
3.1. Variation in Moisture, Ash, Silicon, and Phytolith Concentration
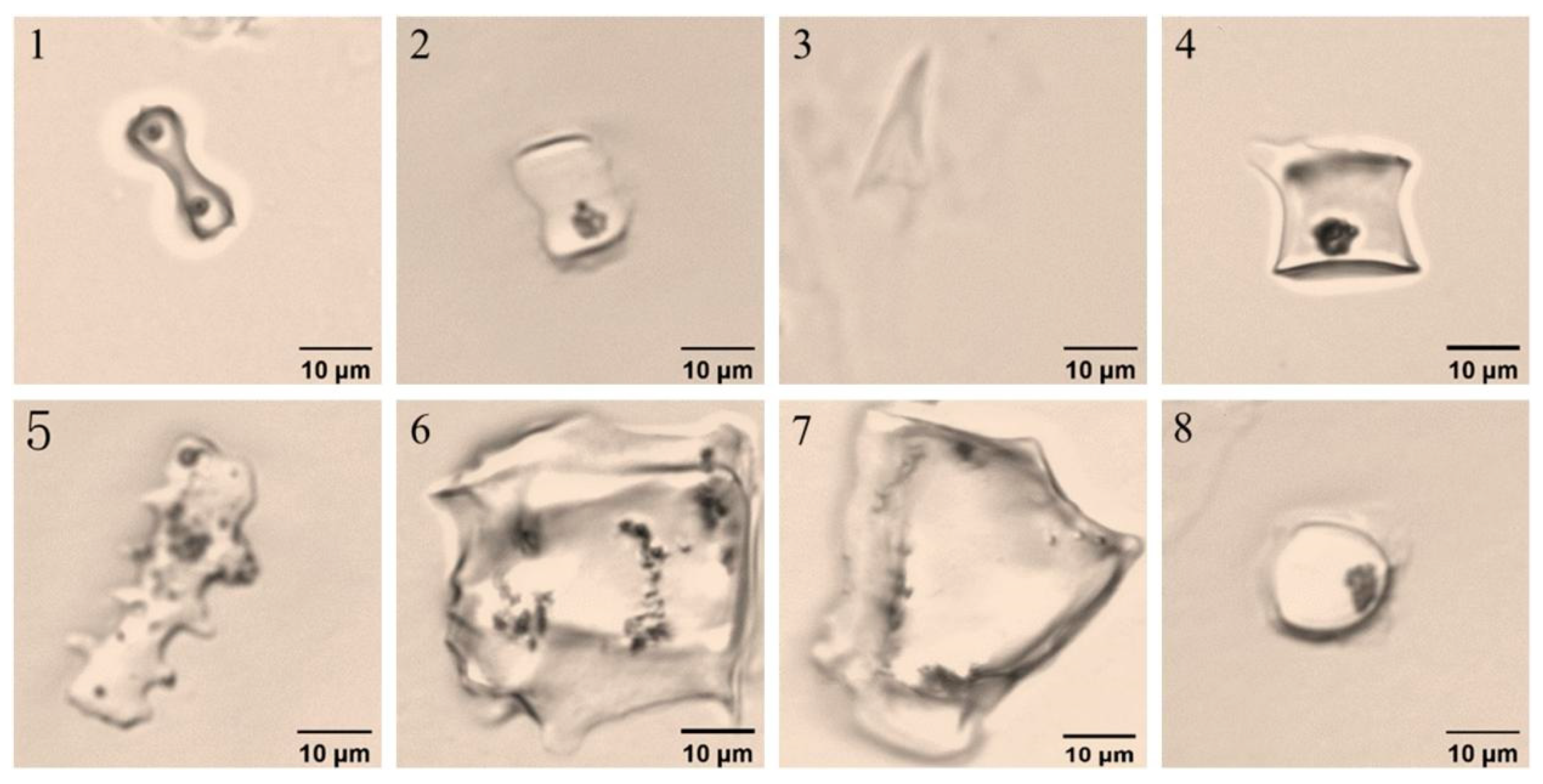
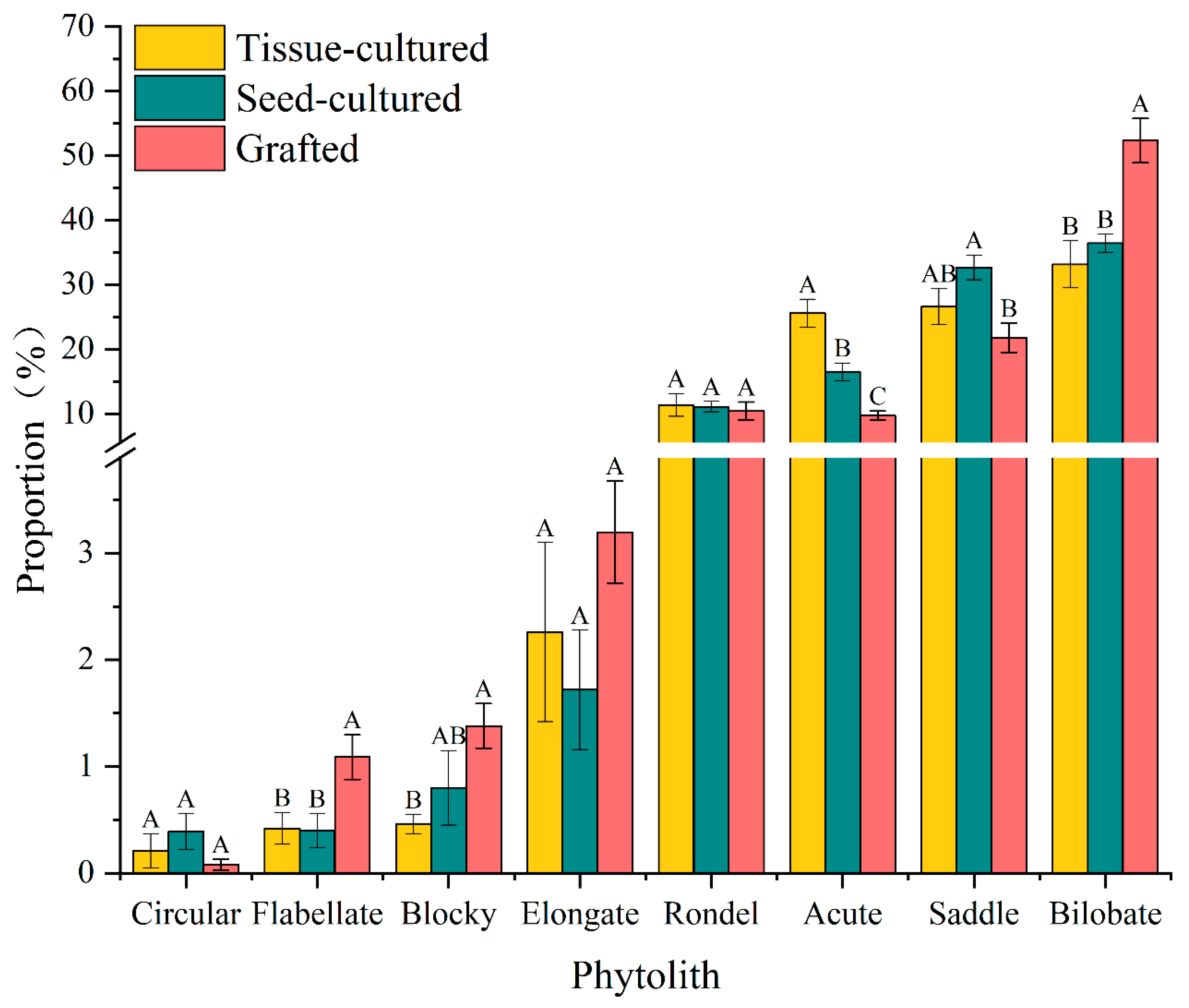
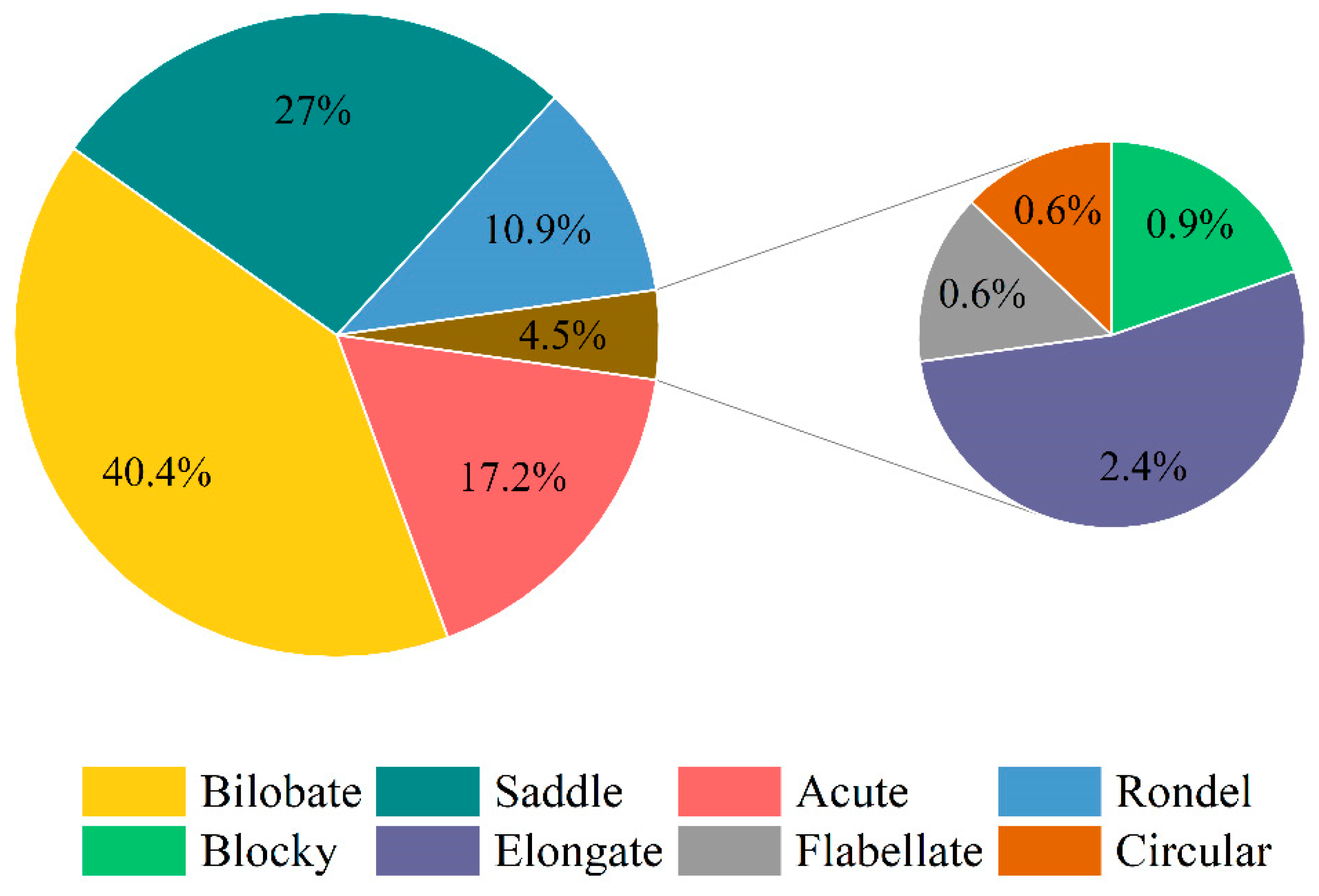
3.2. Variation in Phytolith Morphology
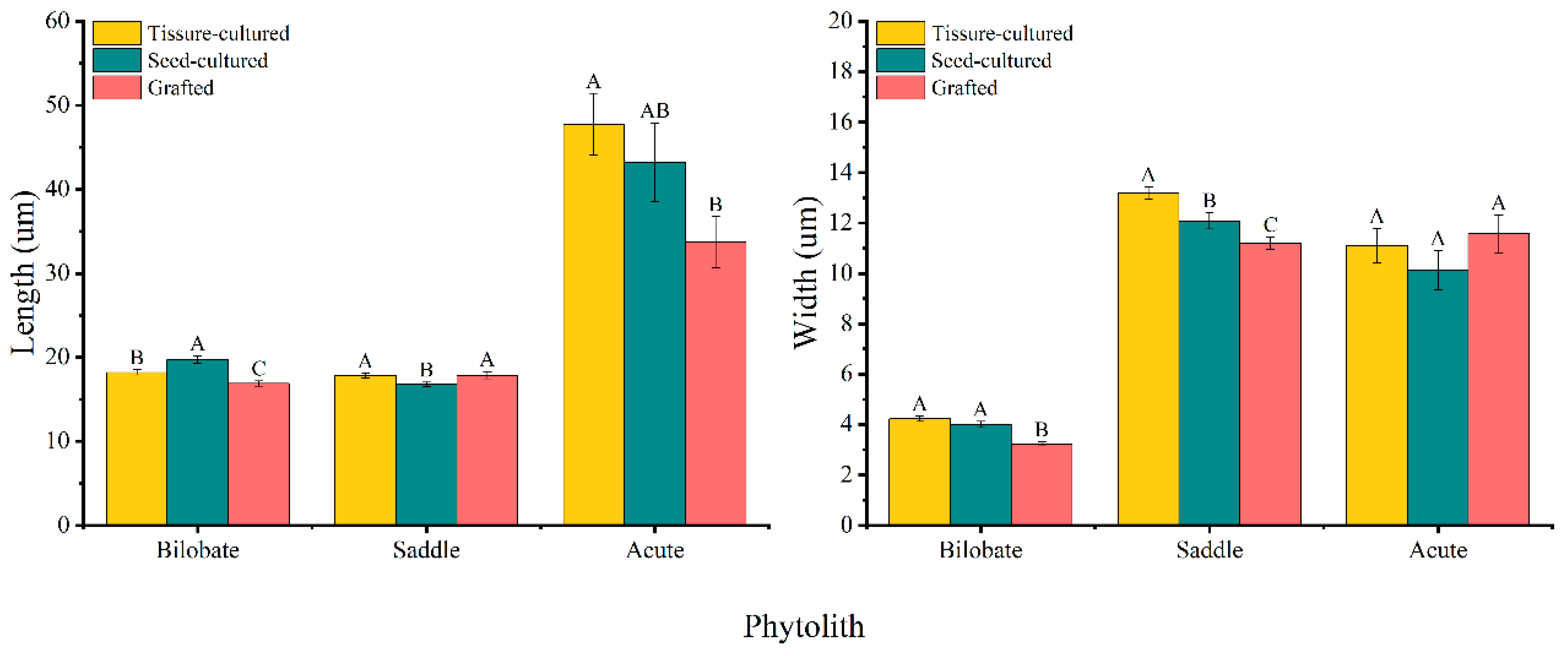
3.3. Variation in Phytolith Size
4. Discussion
4.1. Comparison of Moisture, Ash, Si, and Phytolith Concentration in Different D. brandisii Seedling Leaves
4.2. Morphotypes and Proportions of Phytoliths in Different D. brandisii Seedling Leaves
4.3. Phytolith Sizes in Different D. brandisii Seedling Leaves
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Manivannan, A.; Soundararajan, P.; Jeong, B.R. Silicon: A “Quasi-Essential” element’s role in plant physiology and development. Front. Plant Sci. 2023, 14, 1157185. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, S.; Ohnishi, Y.; Kitagishi, K. Chemical forms, mobility and deposition of silicon in rice plant. Soil. Sci. Plant Nutr. 1962, 8, 15–21. [Google Scholar] [CrossRef]
- Wang, Y.; Lu, H. Phytolith Study and Its Application; China Ocean Press: Beijing, China, 1993. [Google Scholar]
- Twiss, P.C.; Suess, E.; Smith, R.M. Morphological classification of grass phytoliths. Soil. Sci. Soc. Am. J. 1969, 33, 109–114. [Google Scholar] [CrossRef]
- Currie, H.A.; Perry, C.C. Silica in plants: Biological biochemical and chemical studies. Ann. Bot. 2007, 100, 1383–1389. [Google Scholar] [CrossRef]
- Yamanaka, S.; Takeda, H.; Komatsubara, S.; Ito, F.; Usami, H.; Togawa, E.; Yoshino, k. Structures and physiological functions of silica bodies in the epidermis of rice plants. Appl. Phys. Lett. 2009, 95, 123703. [Google Scholar] [CrossRef]
- Massey, F.P.; Hartley, S.E. Physical defences wear you down: Progressive and irreversible impacts of silica on insect herbivores. J. Anim. Ecol. 2009, 78, 281–291. [Google Scholar] [CrossRef]
- Sato, K.; Yamauchi, A.; Ozaki, N.; Ishigure, T.; Oaki, Y.; Imai, H. Optical properties of biosilicas in rice plants. RSC Adv. 2016, 6, 109168–109173. [Google Scholar] [CrossRef]
- Mandlik, R.; Thakral, V.; Raturi, G.; Shinde, S.; Nikolić, M.; Tripathi, D.K.; Sonah, H.; Deshmukh, R. Significance of silicon uptake, transport, and deposition in plants. J. Exp. Bot. 2020, 71, 6703–6718. [Google Scholar] [CrossRef]
- Strömberg, C.A.E.; Di Stilio, V.S.; Song, Z. Functions of phytoliths in vascular plants: An evolutionary perspective. Funct. Ecol. 2016, 30, 1286–1297. [Google Scholar] [CrossRef]
- Bozarth, S.R. Classification of opal phytoliths formed in selected dicotyledons native to the Great Plains. In Phytolith Systematics: Emerging Issues; Springer: Berlin/Heidelberg, Germany, 1992; pp. 193–214. [Google Scholar] [CrossRef]
- Lu, Y.H. Observation of epidermal micromorphology of 23 bamboo leaves. J. Bamboo Res. 1996, 2, 42–44. [Google Scholar]
- Wang, R.H.; Xia, N.H.; Lin, R.S. Epidermal characteristics of the leaves of the genus Bamboo and the genus Peony chophyllum (subfamily Bamboo). J. Trop. Subtrop. Bot. 2002, 10, 22–26. [Google Scholar]
- Zhan, H.; Li, J.; Niu, Z.H.; Li, M.B.; Wang, C.M.; Wang, S.G. Silicon variation and phytolith morphology in different organs of Dendrocalamus brandisii (Munro) Kurz (Bambusoideae). Braz. J. Bot. 2019, 42, 529–541. [Google Scholar] [CrossRef]
- Xu, R.; He, H.; Guo, H.; Zhu, F.; Wang, S.; Dai, C.; Zheng, X.; Xie, D.; Li, H.; Wang, C.; et al. Characteristics of silicon and phytolith distribution in bamboo (Ferrocalamus strictus): Variations between different organs and ages. Rev. Palaeobot. Palynol. 2023, 311, 104817. [Google Scholar] [CrossRef]
- Motomura, H.; Fujii, T.; Suzuki, M. Silica deposition in relation to ageing of leaf tissues in Sasa veitchii (Carriere) Rehder (Poaceae: Bambusoideae). Ann. Bot. 2004, 93, 235–248. [Google Scholar] [CrossRef]
- Niu, Z.; He, W.; Wang, C.; Zhang, L.; Zhan, H.; Wang, S. The Changes in Phytolith Morphology in Culms of Dendrocalamus Giganteus. J. Bamboo Res. 2016, 3, 9–14+25. [Google Scholar] [CrossRef]
- Zhu, F.W.; Niu, Z.H.; Li, J.; Yu, L.X.; Wang, S.G.; Wang, C.M.; Zhan, H. Changes in silicon content and phytolith morphology in Dendrocalamus giganteus at different phenological stages. J. Southwest For. Univ. 2022, 42, 71–77. [Google Scholar] [CrossRef]
- Yi, X.X.; Ma, X.; Liu, M.L.; Wang, F.; Zhang, Q.P. Research progress on effect of silicon on plant stress tolerance and its mechanism. Crop Res. 2020, 34, 398–404. [Google Scholar]
- Li, R.; Fan, J.; Carter, J.; Jiang, N.; Gu, Y.S. Monthly variations of phytoliths in the leaves of the bamboo Dendrocalamus ronganensis (Poaceae: Bambusoideae). Rev. Palaeobot. Palynol. 2017, 246, 62–69. [Google Scholar] [CrossRef]
- Li, D.Z.; Wang, Z.P.; Zhu, Z.D.; Xia, N.H.; Jia, L.Z.; Guo, Z.H.; Yang, G.Y.; Stapleton, C.M.A. Bambuseae. In Flora of China; Wu, Z.Y., Raven, P.H., Hong, D.Y., Eds.; Science Press: Beijing, China, 2006. [Google Scholar]
- He, R.; Qiu, J.; Luo, B. Analysis of Ash and Silica Content of Six Bamboo Species. World Bamboo Ratt. 2016, 14, 4. [Google Scholar] [CrossRef]
- Viegas, D.X.; Viegas, T.P.; Ferreira, A.D. Moisture content of fine forest fuels and fire occurrencein central Portugal. Int. J. Wildland Fire 1992, 2, 69–85. [Google Scholar] [CrossRef]
- GB/T2677.3-1993; Fibrous Raw Material—Determination of Ash. Standards Press of China: Qinhuangdao, China, 1993.
- GB/T 7978-1987; Pulps—Determination of Alcohol–Silicon Dioxide. Standards Press of China: Qinhuangdao, China, 1987.
- Pearsall, D.M. Paleoethnobotany: A Handbook of Procedures, 3rd ed.; Routledge: New York, NY, USA, 2015. [Google Scholar] [CrossRef]
- Madella, M.; Alexandre, A.; Ball, T. International Code for Phytolith Nomenclature 1.0. Ann. Bot. 2005, 96, 253–260. [Google Scholar] [CrossRef]
- Neumann, K.; Stromberg, C.; Ball, T.; Albert, R.; Vrydaghs, L.; Cummings, L.S. International Code for Phytolith Nomenclature (ICPN) 2.0. Ann. Bot. 2019, 124, 189–199. [Google Scholar] [CrossRef]
- Zhan, H. Morphological Characteristics of Phytolithic Body of 10 Kinds of Clumped Bamboo and the Effect of Exogenous Silicon on the Cold Tolerance of Bamboo Seedlings. Master’s Thesis, Southwest Forestry University, Kunming, China, 2017. [Google Scholar]
- Matichenkov, V.V.; Bochamikova, E.A. Chapter 13 The relationship between silicon and soil physical and chemical properties. Stud. Plant Sci. 2001, 8, 209–219. [Google Scholar] [CrossRef]
- Gong, J.L.; Zhang, H.C.; Long, H.Y.; Hu, Y.Q.; Dai, Q.G.; Huo, Z.Y.; Xu, K.; Wei, H.Y.; Gao, H. Progress in research of nutrition functions and physiological mechanisms of silicon in rice. J. Plant Physiol. 2012, 48, 1–10. [Google Scholar] [CrossRef]
- Yoshida, S.; Ohnishi, Y.; Kitagishi, K. Histochemistry of silicon in rice plant. Soil. Sci. Plant Nutr. 1962, 8, 36–41. [Google Scholar] [CrossRef]
- Ma, J.F.; Takahashi, E. Interaction between calcium and silicon in water-cultured rice plants. Plant Soil. 1993, 148, 107–113. [Google Scholar] [CrossRef]
- Agarie, S.; Uchida, H.; Agata, W.; Kubota, F.; Kaufman, P.B. Effects of silicon on transpiration and leaf conductance in rice plants (Oryza sativa L.). Plant Prod. Sci. 1998, 1, 89–95. [Google Scholar] [CrossRef]
- Zhang, G.Q. Effect of Silicon on the Growth and Some Physiological Characteristics of Ginger. Master’s Thesis, Shandong Agricultural University, Taian, China, 2008. [Google Scholar] [CrossRef]
- Rastogi, A.; Yadav, S.; Hussain, S.; Kataria, S.; Hajihashemi, S.; Kumari, P.; Yang, X.; Brestic, M. Does silicon really matter for the photosynthetic machinery in plants…? Plant Physiol. Biochem. 2021, 169, 40–48. [Google Scholar] [CrossRef]
- Song, Z.; Liu, H.; Li, B.; Yang, X.M. The production of phytolith-occluded carbon in China’s forests: Implications to biogeochemical carbon sequestration. Glob. Chang. Biol. 2013, 19, 2907–2915. [Google Scholar] [CrossRef]
- Liu, J.X.; Huang, Z.T.; Jiang, P.K.; Huang, C.P.; Feng, S.F.; Chen, C.; Yin, S. Effects of parent rock and bamboo age on silicon and phytolith-occluded carbon in the leaves of Moso bamboo. Chin. J. Appl. Ecol. 2017, 28, 2917–2922. [Google Scholar] [CrossRef]
- Wang, G.; Zhou, B.Z.; Li, X.J.; Kong, W.J.; Weng, C.H.; Hu, X.L. Carbon storage and vertical spatial distribution of Dendrocalamopsis vario-striata ecosystem. J. Trop. Subtrop. Bot. 2012, 20, 72–77. [Google Scholar] [CrossRef]
- Piperno, D.R.; Pearsall, D.M. The silica bodies of tropical American grasses: Morphology, taxonomy, and implications for grass systematics and fossil phytolith identification. Smithson. Contrib. Bot. 1998, 85, 1–46. [Google Scholar] [CrossRef]
- Li, Q.; Xu, D.K.; Lu, H.Y. Morphology of phytolith in Bambusoideae (Poaceae) and its ecological significance. Quat. Sci. 2005, 25, 777–784. [Google Scholar] [CrossRef]
- Gu, Y.S.; Liu, H.Y.; Wang, H.L.; Li, R.C.; Yu, J.X. Phytoliths as a method of identification for three genera of woody bamboos (Bambusoideae) in tropical southwest China. J. Archaeol. Sci. 2016, 68, 46–53. [Google Scholar] [CrossRef]
- Tao, X.Y. Study on the Taxonomic Significance of Bamboo Leaf Phytolithic Morphology and Micro-Element Composition. Master’s Thesis, Guilin University of Technology, Guilin, China, 2021. [Google Scholar] [CrossRef]
- Zhang, X.; Gélin, U.; Spicer, R.A.; Wu, F.; Farnsworth, A.; Chen, P.; Del, R.C.; Li, S.; Liu, J.; Jian, H.; et al. Rapid Eocene diversification of spiny plants in subtropical woodlands of central Tibet. Nat. Commun. 2022, 13, 3787. [Google Scholar] [CrossRef] [PubMed]
- Tao, X.Y.; Wen, M.D.; Li, R.C.; Vachula, R.S.; Pang, L.N.; Li, C.; Yang, K.Q.; Jiang, N. Phytolith sizes and assemblages differentiate genera and ecotypes of woody bamboos in subtropical Southwest China. Rev. Palaeobot. Palynol. 2020, 272, 104129. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Seedling Type | Tissue-Cultured | Seed-Cultured | Grafted | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Leaf | Young | Mature | T | p | Young | Mature | T | p | Young | Mature | T | p |
| Moisture (%) | 62.87 ± 2.97 a | 57.96 ± 1.90 A | 1.391 | 0.237 | 60.98 ± 0.76 ab | 57.56 ± 2.89 A | 1.145 | 0.316 | 55.71 ± 0.29 b | 55.12 ± 0.55 A | 1.08 | 2.011 |
| Ash (%) | 6.91 ± 0.58 b | 12.87 ± 1.54 AB | −3.635 | 0.022 | 4.56 ± 0.35 c | 10.73 ± 0.69 B | −7.943 | 0.001 | 10.61 ± 0.46 a | 14.61 ± 0.39 A | −6.694 | 0.003 |
| Si (%) | 3.57 ± 0.56 b | 10.99 ± 0.40 A | −10.731 | 0 | 0.95 ± 0.041 c | 7.31 ± 0.57 B | −11.186 | 0.008 | 6.49 ± 0.03 a | 8.82 ± 0.56 B | −4.128 | 0.053 |
| Phytolith concentration (103 particles.g−1) | 31.13 ± 0.39 a | 62.93 ± 3.41 A | −9.274 | 0.001 | 10.32 ± 0.16 c | 38.43 ± 3.99 B | −7.04 | 0.002 | 15.20 ± 0.39 b | 20.56 ± 0.86 C | 5.689 | 0.005 |
| Seedlings | Leaf | Bilobate | Saddle | Acute | Rondel | Elongate | Blocky | Flabellate | Circular |
|---|---|---|---|---|---|---|---|---|---|
| Tissue-cultured | Young | 25.56 ± 0.99 | 32.44 ± 2.26 | 25.51 ± 2.85 | 15.23 ± 0.10 | 0.39 ± 0.07 | 0.40 ± 0.09 | 0.16 ± 0.08 | 0.32 ± 0.32 |
| Mature | 40.72 ± 2.88 | 20.76 ± 0.90 | 25.62 ± 3.91 | 7.48 ± 0.61 | 4.12 ± 0.25 | 0.52 ± 0.17 | 0.68 ± 0.20 | 0.09 ± 0.09 | |
| T | −4.979 | 4.799 | −0.024 | 12.604 | −14.197 | −0.642 | −2.475 | 0.525 | |
| p | 0.008 | 0.009 | 0.982 | 0.000 | 0.000 | 0.556 | 0.069 | 0.000 | |
| Seed-cultured | Young | 34.73 ± 2.44 | 36.04 ± 2.47 | 15.70 ± 1.90 | 12.50 ± 1.05 | 0.64 ± 0.36 | 0.08 ± 0.08 | 0.08 ± 0.08 | 0.24 ± 0.14 |
| Mature | 38.14 ± 1.03 | 29.33 ± 0.99 | 17.25 ± 2.36 | 9.71 ± 0.37 | 2.79 ± 0.53 | 1.52 ± 0.29 | 0.71 ± 0.12 | 0.54 ± 0.33 | |
| T | −1.289 | 2.519 | −0.510 | 2.495 | −3.356 | −4.847 | −4.516 | −0.866 | |
| p | 0.267 | 0.065 | 0.637 | 0.067 | 0.028 | 0.008 | 0.011 | 0.436 | |
| Grafted | Young | 45.44 ± 2.63 | 25.80 ± 2.77 | 9.27 ± 1.09 | 12.97 ± 1.44 | 3.36 ± 1.00 | 1.71 ± 0.27 | 1.37 ± 0.34 | 0.08 ± 0.08 |
| Mature | 59.27 ± 2.01 | 17.73 ± 1.29 | 10.14 ± 0.99 | 7.89 ± 1.05 | 3.04 ± 0.39 | 1.04 ± 0.17 | 0.80 ± 0.16 | 0.08 ± 0.08 | |
| T | −4.179 | 2.640 | −0.589 | 2.850 | 0.301 | 2.109 | 1.516 | −0.016 | |
| p | 0.014 | 0.058 | 0.587 | 0.046 | 0.778 | 0.103 | 0.204 | 0.988 |
| Seedlings | Tissue-Cultured | Seed-Cultured | Grafted | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Leaf | Young | Mature | T | p | Young | Mature | T | p | Young | Mature | T | p | |
| Bilobate | Length (um) | 17.01 ± 0.33 b | 19.36 ± 0.44 A | −4.231 | 0 | 20.12 ± 0.60 a | 19.37 ± 0.69 A | 0.834 | 0.408 | 17.17 ± 0.48 b | 16.57 ± 0.38 B | 0.969 | 0.337 |
| Width (um) | 3.98 ± 0.14 a | 4.47 ± 0.16 A | −2.34 | 0.23 | 4.03 ± 0.19 a | 3.97 ± 0.16 B | 0.265 | 0.792 | 3.25 ± 0.10 b | 3.20 ± 0.13 C | 0.262 | 0.795 | |
| Saddle | Length (um) | 17.25 ± 0.32 a | 18.35 ± 0.40 A | −2.145 | 0.036 | 17.39 ± 0.40 a | 16.12 ± 0.37 B | 2.338 | 0.023 | 16.69 ± 0.51 a | 19.05 ± 0.62 A | −2.952 | 0.005 |
| Width (um) | 12.50 ± 0.26 a | 13.85 ± 0.39 A | −2.883 | 0.006 | 13.16 ± 0.46 a | 11.00 ± 0.34 B | 3.745 | 0 | 11.36 ± 0.31 b | 11.02 ± 0.37 B | 0.709 | 0.481 | |
| Acute | Length (um) | 51.60 ± 5.41 a | 43.89 ± 4.96 A | 1.051 | 0.298 | 31.36 ± 3.82 b | 55.04 ± 8.08 A | −2.648 | 0.011 | 40.12 ± 5.73 ab | 27.25 ± 1.54 B | 2.167 | 0.037 |
| Width (um) | 10.44 ± 1.05 ab | 11.75 ± 0.87 A | −0.965 | 0.339 | 9.53 ± 0.80 b | 10.71 ± 1.34 B | −0.759 | 0.451 | 12.63 ± 0.88 a | 10.50 ± 1.19 B | 1.435 | 0.157 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dai, C.; Xu, R.; Yu, L.; Zhu, F.; Li, M.; Li, J.; Wang, S.; Wang, C.; Zhan, H. Silicon Uptake and Phytolith Morphology in Dendrocalamus brandisii Seedling Leaf from Different Rearing Methods. Forests 2023, 14, 1877. https://doi.org/10.3390/f14091877
Dai C, Xu R, Yu L, Zhu F, Li M, Li J, Wang S, Wang C, Zhan H. Silicon Uptake and Phytolith Morphology in Dendrocalamus brandisii Seedling Leaf from Different Rearing Methods. Forests. 2023; 14(9):1877. https://doi.org/10.3390/f14091877
Chicago/Turabian StyleDai, Cefeng, Rui Xu, Lixia Yu, Fangwei Zhu, Maobiao Li, Juan Li, Shuguang Wang, Changming Wang, and Hui Zhan. 2023. "Silicon Uptake and Phytolith Morphology in Dendrocalamus brandisii Seedling Leaf from Different Rearing Methods" Forests 14, no. 9: 1877. https://doi.org/10.3390/f14091877
APA StyleDai, C., Xu, R., Yu, L., Zhu, F., Li, M., Li, J., Wang, S., Wang, C., & Zhan, H. (2023). Silicon Uptake and Phytolith Morphology in Dendrocalamus brandisii Seedling Leaf from Different Rearing Methods. Forests, 14(9), 1877. https://doi.org/10.3390/f14091877