Abstract
Plant endosymbionts (endophytes) influence host plant health and express genotype-dependent ecological relationships with plant hosts. A fungal species intended to confer host plant resistance to a forest pathogen was used as inoculum to test for effects of inoculation on disease resistance, microbiomes, and phytochemistry of a threatened pine species planted in a restoration setting. Correlations of inoculation presence/absence, phytochemistry, spatial location of seedlings, maternal seed sources, and fungal endophytic communities in the foliage of six-year-old whitebark pine (Pinus albicaulis) seedlings were assessed five years after an experimental inoculation of seedlings with foliar endophytic fungi cultured from whitebark pine trees at Crater Lake National Park, including Paramyrothecium roridum. We hypothesized that P. roridum would modify host microbiomes in a manner that combats white pine blister rust disease. Our assessment of seedlings in the field five years after inoculation allowed us to consider whether inoculation stimulated long-lasting changes in microbiome communities and whether effects varied by seedling genetic family. Tests for effects of endophyte inoculation on disease resistance were inconclusive due to current low levels of rust infection observed at the field site. Foliar fungal endophyte richness and Shannon diversity varied with maternal seed sources. Isotopic stoichiometry and phytochemistry did not vary with seedling spatial proximity, inoculation treatment, or maternal seed family. However, endophyte community composition varied with both seedling spatial proximity and maternal seed sources. Endophytic communities did not vary with the inoculation treatment, and the hypothesized biocontrol was not detected in inoculated seedlings. We draw three conclusions from this work: (1) fungal microbiomes of whitebark pine seedlings across our study site did not vary with host phytochemical signatures of ecophysiological status, (2) the inoculation of P. albicaulis seedlings with a mixture of fungal endophytes did not lead to persistent systemic changes in seedling foliar microbiomes, and (3) in correspondence with other studies, our data suggest that maternal seed source and spatial patterns influence fungal endophyte community composition.
1. Introduction
Microbes that inhabit plant interiors (endophytes) associate differently with host plants depending on host genotype [1,2,3,4]. Endophyte–plant interactions range from mutualistic to parasitic [5] and often influence important parameters of host plant physiology [6]. For instance, fungal endophytes may improve plant performance under periods of abiotic and biotic stress by altering plant water relations through adjustment of host tissue osmotic potential [6,7], improving host plant heat tolerance [8,9], and influencing host plant disease resistance [10,11,12]. Endophytic microbe–microbe interactions also indirectly influence plant growth and disease susceptibility (see review by Chaudhry et al. [13]). As a result, there is increasing interest in how best to engineer plant–microbe interactions for applications in agronomy [14,15], forestry [16,17], and biological conservation [4,18].
Plant chemical characteristics (phytochemistry), including carbon (C) and nitrogen (N) stable isotope ratios and non-structural carbohydrate concentrations, can vary with plant physiological responses to environmental factors [19,20]. For example, the extent to which plants discriminate against 13C in photosynthesis varies with plant drought stress integrated across the lifespan of foliar tissue [21,22] and has also been shown to vary with fungal endophyte composition [23]. Plant foliar N isotope discrimination varies with differences in N availability as well as plant mycorrhizal colonization [24]. The concentration of plant non-structural carbohydrates, i.e., sugars and starches that serve as carbon reserves and osmolytes [25], may vary with shifts in fungal endophyte communities and functions [26,27]. Endophytic fungi may also alter defensive host phytochemistry and produce their own suite of chemicals that influence or complement host plant defenses against pathogens [10,11,28,29,30,31,32,33].
Plant–microbe associations that aid in plant disease resistance may be engineered through controlled plant microbial inoculation [34,35,36]. Such microbiological controls are often achieved by encouraging antagonistic interspecific interactions among microbes in planta [37,38]. Of particular interest are alterations to mechanisms of ecological succession characterized by physical or chemical interference among endophytes and pathogenic microbes [10,11,28,29,32,39]. Difficulties exist for pre-planting biocontrol inoculations of a long-lived species to confer long-term resistance against a disease. Under this framework, the inoculant must become incorporated into new host tissue as the host develops, restructure the host mycobiome (sensu priority effects) in a manner that confers a biocontrol benefit to the host, or induce systemic resistance in the host. In any case, inoculation-based forest biocontrols would be economically viable only if the beneficial effect of inoculation protects against timber volume loss at least until trees reach stand rotation age, in the context of commercial forestry, and until trees reach reproductive maturity, in the context of forest restoration.
1.1. Whitebark Pine
Whitebark pine (Pinus albicaulis Engelm. (Pinaceae)) is a seral, high-elevation white pine (subgenus Strobus [40]) unique to western North America, where it moderates the rate of snowmelt and provides crucial habitat for alpine flora and a protein source for fauna [41,42]. White pine blister rust disease (WPBR), caused by the non-native fungal pathogen Cronartium ribicola J.C. Fisch. (Basidiomycota Pucciniales), poses a lethal threat to all North American species of five-needle white pines. WPBR causes early mortality in whitebark pines, can reduce or eliminate seed production [43], and has caused extensive mortality in many whitebark pine populations. Mountain pine beetle infestations, climate change, and WPBR together have led to the listing of whitebark pine as an endangered species in Canada, and the recommendation in 2020 for listing as a threatened species under the U.S. Endangered Species Act [44,45].
Enhancing forest resilience through microbiome engineering is increasingly considered an important approach for forest management [46]. In a study of western white pine (P. monticola) seedlings following inoculation with multiple species of fungal endophytes cultured from the foliage of wild P. monticola, endophyte inoculations led to longer survival and reduced WPBR disease severity as compared to the control treatment following experimental infection with WPBR [12]. One year post-infection, only 18% of control-group seedlings survived, whereas 39% of endophyte-treated seedlings survived. Reduced WPBR disease severity may relate to characteristics of host defensive chemistry, which has been found to vary with host plant genotype as well as host foliar microbiomes of whitebark pine [4]. If inoculation with fungal endophytes is capable of providing long-term resistance against WPBR, or can be reapplied to enhance the longevity of disease resistance, inoculations may serve as a useful complement to genetic resistance screening programs already used to help protect P. albicaulis [47]. However, little is known about the longevity of fungal endophyte inoculations under field conditions or their long-term effects on host ecophysiology.
1.2. Hypotheses
For this field trial, an endophyte inoculation solution was developed and applied to P. albicaulis seedlings in the same way as that described in [12], but with endophytes isolated from P. albicaulis at Crater Lake National Park (CLNP), including Paramyrothecium roridum Tode (Ascomycota Hypocreales). Seedlings from seed cones collected within CLNP were inoculated prior to outplanting in a restoration garden located near seed source trees. Using this study system, we investigated the influence of the inoculum on blister rust incidence, fungal endophyte community composition, and whitebark pine phytochemistry. Samples were collected during the fifth growing season post-inoculation. We tested four hypotheses to elucidate the influence of endophyte inoculation on WPBR incidence after multiple seasons post-inoculation, the association of fungal endophyte communities with host phytochemistry, and the post-inoculation associations of endophytic communities with maternal seed sources.
Hypothesis 1:
Fewer endophyte-inoculated seedlings will show signs of WPBR infection as compared to seedlings in the control group.
Hypothesis 2:
Host phytochemistry will vary with fungal endophyte community composition.
Hypothesis 3:
Fungal endophyte inoculation of whitebark pine seedlings will result in a durable change in host phytochemistry and fungal endophyte community diversity and composition compared to a paired control group.
Hypothesis 4:
Fungal endophytic diversity and community composition will vary with maternal seed sources, due to underlying genetic differences in seedlings.
2. Materials and Methods
2.1. Seedling Germination, Inoculation, and Sample Collection
The maternal families examined in this field trial originated from seed collections made in 2005 from five wind-pollinated seed trees occupying the periphery of Crater Lake at Crater Lake National Park, Oregon, USA. Along with macroclimatic characteristics, soil parent materials were similar among maternal trees due to the predominance of pumice and basaltic cobble deposited by the eruption of Mt. Mazama approximately 7700 years ago [48] which formed the cavity now known as Crater Lake. Progeny of the five seed parents showed varying levels of resistance to white pine blister rust in a nursery screening trial (Sniezko, unpublished). Seeds from the maternal trees were germinated in spring 2007 at the U.S. Forest Service Dorena Genetic Resource Center (DGRC; Cottage Grove, OR). Germinated seedlings in 164 cm3 Ray Leach SC 10 Super Cell containers (with cell diameters of 3.8 cm and depths of 21 cm) were grouped by maternal genetic family in three experimental blocks. In October 2008 (after two growing seasons), seedlings were randomly assigned to experimental control (n = 100) and inoculation groups (n = 100; Figure 1). Inoculation group seedlings were inoculated at DGRC with a sterile water suspension of fungal hyphae from endophytes cultured from whitebark pines at Crater Lake NP using methods described in [12]. The mixture contained P. roridum, and P. roridum was the only species identified from the inoculation mixture. Inoculation was conducted by mist-dispersal of the hyphal suspension upon seedling roots and shoots (G. Newcombe, pers. comm.). The P. roridum isolate was collected from healthy whitebark pine needles from Crater Lake NP, as described in [49]. In this study, we focused on the P. roridum component of the inoculation mixture due to its hypothesized potential to act as a deterrent of C. ribicola based on evidence of antagonistic behavior of P. roridum against crop pathogens [49,50,51]. Following inoculation, seedlings were transplanted into larger growth containers for the third and final growing season at DGRC. A total of 192 surviving seedlings from the five maternal families, half inoculated and half control, were planted in fall 2009 on a 5000 m2 plot near the southern rim of CLNP (latitude: 42.90276°, longitude: −122.14541°) in single-family clusters with inoculated and uninoculated individuals in the same cluster (Figure 1C). Randomly selected, paired inoculant/control seedlings were planted adjacent to one another within family groups. In July 2013, three healthy needle fascicles were collected from evenly spaced branches of the lower canopy of each seedling (following [52]). Sampling was limited to 79 individual trees because of high levels of rodent-related seedling mortality (Table 1, Figure 1). Sampled seedlings were visually assessed in the field for WPBR presence. Needle fascicles clipped from each seedling were placed in separate plastic bags and frozen at −20 °C within 12 h of collection following [53]. The spatial extent of the study was small (<300 m2), resulting in mesoscale environmental conditions (substrate, slope, aspect) that were relatively constant among maternal families, and we explicitly controlled for spatial effects in analyses. However, the clustered planting of seedlings from each maternal family (Figure 1) hampered inferences concerning maternal family effects on endophytic communities.
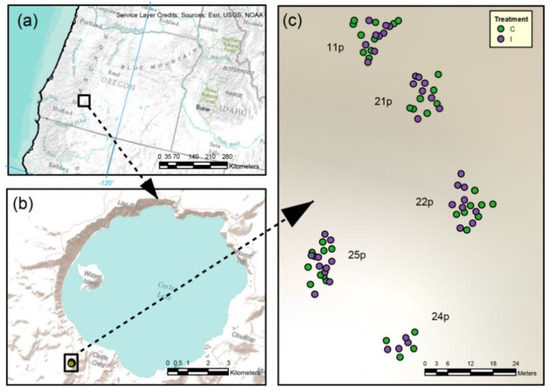
Figure 1.
Maps: (a) Northwest portion of USA, including the state of Oregon; (b) the Crater Lake area; (c) specific area of sample collection, showing locations of whitebark pine seedlings from five seed source trees (11p, 21p, 22p, 24p, 25p) with the original inoculum versus control treatment levels shown by purple (inoculated) and green (control) circles.

Table 1.
Sample sizes for the maternal family (11p, 21p, 22p, 24p, 25p; where “p” refers to parent) and fungal inoculum treatment (C = control, I = inoculum) factor level combinations in the experiment.
2.2. Surface Sterilization and DNA Extraction
Sample fascicles were immersed in sterile deionized water and stirred for 1 min prior to immersion for 1 min in EtOH, 3 min in NaOCl (4% available Cl), 30 s in EtOH, and 30 s in sterile deionized water following [54]. Sterilized samples were air-dried in a sterile flow-hood, and dried needles were then stored at −20 °C under sterile conditions. Following [55,56], negative and positive controls were produced using sterile culturing of pine needles on potato dextrose agar to establish that surface sterilization was adequate but did not preclude fungal growth from inside needle tissue.
Separately for each sample, pine needle tissue was ground to a fine powder using liquid nitrogen in a sterilized mortar and pestle. Genomic DNA was isolated from each sample as follows. Fifty-milligram samples of powderized tissue were added to a separate bead tube from the MoBio PowerPlant Pro kit (Carlsbad, CA, USA), which received 40 μL of the MoBio phenolic separation solution. Two rounds of bead maceration were applied (two minutes per round), punctuated by two minutes on ice. Extracted DNA concentrations were quantified using a NanoDrop 1000 Spectrophotometer (Thermo Scientific). A MoBio extraction tube without needle tissue was included in the DNA extraction protocol as a negative control and treated as a normal sample throughout the protocol through the PCR amplification step.
2.3. Amplification, Sequencing, and Library Analysis
Extracted genomic DNA was amplified following a semi-nested PCR approach. Two sequential combinations of primers specific to the internal transcribed spacer (ITS) region of fungal rDNA were used to isolate the ITS1 locus for next-generation sequencing. The internal primer set was chosen to selectively amplify PCR products contained within the external target region. The external and internal primer pairs both included the ITS1-F forward primer from [57]. We used ITS4 for the external reverse primer and ITS2 as the internal reverse primer [58]. Internal ITS1-F and ITS2 primers were constructed with overhanging adapters prescribed for use with the Illumina MiSeq platform, and an additional guanine for Tm optimization of the ITS1-F primer directly between the primer and adapter. For the internal primers, 5′ ends of ITS1-F and ITS2 primers were ligated to Illumina adapters: 5′-TCGTCGGCAGCGTCAGATGTGTATAAGAGACAG-3′, and 5′-GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAG-3′, respectively.
Touchdown annealing was used in the semi-nested reaction to enhance primer specificity for fungal DNA and to maximize the breadth of taxa that could be amplified [59]. Internal amplicons were generated directly after completion of the external PCR, and semi-nested PCR products were used for all downstream applications directly after internal amplification. The external PCR reaction solution contained 12.5 μL of 2× Dream Taq DNA polymerase (Thermo Scientific), 25 μM ITS1-F primer + adapter, 25 μM ITS4 primer, 10 ng template DNA, and molecular-grade nanopure H2O to 25 μL. The external thermal cycle included an initial denaturation step of 85 s at 95 °C; 10 cycles of 35 s at 95 °C, 55 s at 57.5 °C, and 45 s at 72 °C; 10 cycles of 35 s at 95 °C, 55 s at 57.5 °C, and 120 s at 72 °C; and a final 72 °C elongation step of 10 min.
The internal PCR reaction solution consisted of 5 μ/μL HotStarTaq (Qiagen), 2.5 μM ITS1-F primer + adapter, 2.5 μM ITS2 primer + adapter, 25 mM MgCl2 (Qiagen), 10 mM dNTPs (Qiagen), 2.5 μL of 10× PCR buffer (Qiagen), 10% diluted external PCR product, and nanopure H2O to 25 mL. The internal thermal cycle included an initial hot-start Taq activation step of 15 min at 95 °C followed by an additional 3 min of denaturation at 94 °C; 13 touchdown cycles of 60 s at 94 °C, 60 s starting at 67 °C and decreasing 1 °C for each of the final 12 steps of the touchdown cycle, and 120 s at 72 °C; 12 cycles of 60 s at 94 °C, 60 s at 55 °C, and 120 s at 72 °C; and a final elongation of 10 min at 72 °C.
Cloning of one mixed-template amplicon sample was conducted to confirm the efficacy of the PCR protocol following [53]. Sanger sequencing and MegaBLAST of a cloned product yielded a 100% identity match to the uncultured endophyte Phialocephala sp. (E = 2 × 10109). PCR amplicons were cleaned with the Agencourt AMPure XP-PCR Purification kit (Beckman Coulter) and ligated with a unique sequence barcode indicating host tree using the Nextera XT DNA indexing kit from Illumina. Libraries of 250–300 nt paired-end reads were generated using Illumina MiSeq (V3-600 cycle).
Sequenced reads were de-multiplexed at the Molecular Research Core Facility (MRCF) at Idaho State University in Pocatello, Idaho, with the bcl2fastq Conversion Software from Illumina. All further sequence processing was performed in mothur (v.1.35.1) following the mothur MiSeq standard operating procedure (accessed March of 2020) as described in [60]. Briefly, bi-directional reads were assembled into contigs using consensus screening [61]. Reads that failed to join were discarded. Contigs of length < 225 nt, containing ambiguous bases, or containing ≥ 8 homopolymers were discarded. Contig length was constrained from an average of 265 nt to 225 nt. Contigs were pre-clustered using single-linkage hierarchical agglomerative classification, which results in lower OTU-calling error rates than pre-clustering using the more common complete-linkage approach [62], and then screened for chimeras using the de novo UCHIME detection algorithm [63]. Sequence libraries were normalized to 2646 unique sequences per tree; the largest common number of unique sequences detected across all trees. We then aligned contigs against the dynamic UNITE database formatted for mothur (v. 7) and clustered to operational taxonomic units (OTUs) at ≥97% sequence similarity using the Needleman–Wunsch average neighbor clustering algorithm [64,65]. The UNITE database and 97% cutoff for OTU delimitation, common to studies of Basidiomycetes, as well as the PCR protocol described above were chosen to optimize the detection of the Basidiomycete species of particular interest in this study, C. ribicola (the fungal pathogen hypothesized to be controllable with P. roridum). The steps above were used to generate a multivariate endophytic community dataset with 637 fungal operational taxonomic units (OTUs) distinguishing 79 sampled whitebark pine trees. Molecular data from this Targeted Locus Study project have been deposited at DDBJ/EMBL/GenBank under accession No. KBTW00000000. The version described in this paper is the first version, KBTW01000000.
2.4. Phytochemical Analysis
Our assessment of phytochemistry included C and N isotope discrimination and non-structural carbohydrates in healthy-looking seedlings. Concentrations of foliar non-structural carbohydrates (NSCs; glucose, fructose, and sucrose in our study), were measured using a Roche Yellow-Line Glucose/Fructose and Sucrose Assay Kit. Ground pine needle tissue was weighed to 4 mg ± 0.1 mg, and 2 mL of water was added to each sample and heated for 30 min to extract soluble sugars into an aqueous solution following [66]. A 200 µL aliquot of each sample was treated with the enzymes invertase, phosphoisomerase, and glucose hexokinase to convert sucrose, fructose, and glucose to 6-phosphogluconate. Spectrophotometric analysis was then conducted on digested aliquots using a Synergy Microplate Reader (Biotek Instruments, Winooski, VT, USA) set to read absorbance at 340 nm to determine the quantity of NAD converted to NADH, which is directly proportional to soluble sugar concentrations. These determinations were made on a subset of the homogenized pine needle tissue remaining after DNA extraction, drawn from a balanced proportion of experimental/control seedlings and maternal families (n = 30). Absolute mass and relative proportions of NSCs were calculated for glucose, fructose, and sucrose per gram of leaf sample.
Foliar tissue carbon and nitrogen isotope discrimination (denoted δ13C and δ15N, respectively) were measured using isotope ratio mass-spectrometry, on a random subset of ground needle samples from the five maternal families (n = 53). Four milligrams of needle tissue were packed into 4 × 6 mm tins and analyzed at the Center for Archaeology, Materials and Applied Spectroscopy at Idaho State University using a Costech ECS 4010 Elemental Analyzer interfaced to a Delta V advantage isotope ratio mass spectrometer through the ConFlo IV system. Elemental analyses were conducted using evolutionary flash-combustion and chromatographic separation techniques with a furnace temperature of 1000 °C and reduction oven temperature of 650 °C. We measured δ13C as ‰ values relative to the VPDB standard. We report δ15N values as ‰ values relative to atmospheric N2. Four in-house standards (ISU Peptone, Costech Acetanilide, Glycine, and DORM-3), calibrated against international standards (IAEA-N-1, IAEA-N-2, USGS-25, USGS-40, USGS-41, USGS-24, IAEA-600), were used to create a two-point calibration curve to correct raw isotope measures [67,68].
2.5. Statistical Analyses
We used general linear mixed effect models (Ch. 10 of [69]) to consider treatment (inoculum vs. control), maternal family, and interaction effects on endophytic fungal OTU Shannon diversity [70], OTU richness, and phytochemistry (non-structural carbohydrate composition, δ13C, and δ15N). As noted above, carbohydrate analyses were conducted on a stratified random subset of 30 of the 79 sampled trees, whereas δ13C and δ15N values were obtained for a stratified random subset of 50 of the 79 sampled trees. In all linear mixed effect models, inoculum treatment was designated as a fixed effect, and maternal family and the interaction of treatment and family were designated as random effects. To help address spatial clustering of genetic families, linear mixed effect models were spatially explicit. Spherical spatial covariance structures were used [71] based on preliminary analyses of potentially valid spatial autocorrelation structures using the Akaike information criterion (AIC; [72,73]). We performed indicator species analyses [74] to determine whether endophytic species demonstrated a preference for treatment levels, using the R function indval from the package labdsv [75]. Only sequences that could be given a taxonomic assignment with ≥90% confidence were assessed in indicator species analyses [76].
Multivariate approaches were specifically chosen and applied to gauge the main effects of maternal family and inoculum treatment on multivariate responses (i.e., fungal endophyte community composition, isotopic stoichiometry (δ13C, δ15C), non-structural carbohydrate composition (glucose, fructose, sucrose)) and to parse these effects from potentially confounding patterns in the spatial planting of seedling families. We created distance-based Moran eigenvector maps (MEMdbs) from sample spatial coordinates to quantify multivariate patterns in spatial proximity at multiple independent spatial scales. To create MEMdbs, we used the length of the largest edge from a minimum spanning tree of Euclidean distances of spatial coordinates as the truncation threshold for analyses and binary weighting [77] (p. 863). Of the 78 Moran eigenvectors generated, only the first 11 had positive eigenvalues. Following [77], we limited our multivariate spatial considerations to those dimensions. The resulting eigenvectors were vector-fit onto ordinations and included as covariates in variance partitioning analysis and distance-based redundancy analysis.
Variance partitioning analysis (VPA; [78]) calculates variation in a multivariate response, accounted for by two to four explanatory variables and their combined effects. VPA was used to parse out the effects of confounding multivariate covariates, including spatial effects to allow undiluted expression of the variance explained by the main effects of interest.
Variance partitioning analyses are complementary to distance-based redundancy analysis (dbRDA; refs. [79,80]), which can be used to apply formal multivariate null hypothesis tests, while expressly conditioning on covariates. We used this approach to test for the effect of genetic family, the inoculum treatment, spatial proximity (positive MEMdbs), and their interactions on endophyte community composition, isotopic stoichiometry, and non-structural carbohydrate composition in separate dbRDA models. We used Bray–Curtis dissimilarity [81] as the underlying resemblance metric in dbRDAs.
Non-metric multidimensional scaling (NMDS [82]) was used to visualize patterns among the endophytic OTU communities of whitebark pine seedlings. Bray–Curtis dissimilarity was again used as the resemblance metric. NMDS projections were the lowest stress solution obtained from 20 random starting configurations to prevent convergence to local minima. We used vector and factor fitting analyses [83] to test for associations of environmental variables, including MEMdbs, with the simplified community space of the ordination projection.
3. Results
Sampled seedlings did not show signs of WPBR, and thus hypothesis 1 was discarded.
3.1. Sequence Library Summary
Sequence filtering and screening retained 209,034 sequences that were grouped into 637 OTUs, 148 of which could be classified as Linnaean taxa within 123 genera, 75 families, 44 orders, 18 classes, and 4 phyla. As expected, the dominant phylum was Ascomycota, comprising 98.4% of the sequences. Basidiomycota accounted for 1.6% of the sequences. Neither P. roridum nor C. ribicola were identified in samples. See Appendix A for further details concerning sequence library results.
3.2. Host Phytochemistry Did Not Vary with Fungal Endophyte Community Composition
Non-structural carbohydrates were unassociated with either fungal endophyte OTU Shannon diversity (Pearson’s r = 0.16, p = 0.39, n = 30) or community composition, as summarized by a three-dimensional NMDS ordination of samples (n = 30) for which isotope data were obtained (R2 = 0.06, p = 0.75, NMDS stress = 0.129). Significant Pearson correlations were also absent between fungal endophyte OTU diversity and either δ15N or δ13C (r = 0.12, p = 0.41 and r = 0.02, p = 0.87, respectively, n = 53), and associations were not present for either δ15N or δ13C and fungal endophyte OTU community composition, as summarized by a three-dimensional NMDS ordination of samples (n = 50) for which isotope data were obtained (R2 = 0.02, p = 0.84 and r < 0.01, p = 0.98, respectively, NMDS stress = 0.153). See Appendix B for a summary of sample phytochemistry values.
Endophyte inoculation of whitebark pine seedlings did not influence host phytochemistry or fungal endophyte community diversity and composition.
Inoculation treatment did not influence total non-structural carbohydrates ( = 0.93, p = 0.334), δ15N ( = 0.04, p = 0.837), δ13C ( = 0.21, p = 0.664), fungal endophyte diversity ( = 1.68, p = 0.194), or fungal endophyte community composition (F1.65 = 0.85, p = 0.58; Table 2).
Table 2.
Distance-based redundancy analysis for endophyte composition, isotope stoichiometry, and non-structural carbohydrates for predictor main effects, 999 permutations used. p-values are adjusted for simultaneous inference using Holm’s sequential Bonferroni procedure.
3.3. Fungal Endophyte Community Composition Varied with Seed Source
Shannon OTU diversity ( = 5.19, p = 0.011; Figure 2A) and OTU richness ( = 6.61, p = 0.005; Figure 2B) varied across maternal families. Spatial proximity patterns defined by the first 11 MEMdbs are shown in Figure 3. MEMdb 1, 3, and 6 were particularly relevant to our analyses. Note that in MEMdb 1, loadings of spatial coordinates increased from south to north, but in a counterclockwise pattern, with the lowest weights occurring in the southwest and the highest weights occurring in the northwest (Figure 3). For MEMdb 3, eigenvector loadings generally increased from northwest to southeast, strongly distinguishing family 24p (Figure 3). We assessed the associations of the first 11 MEMdbs, easting, northing, and maternal family with the projection using vector and factor fitting (Table 3).
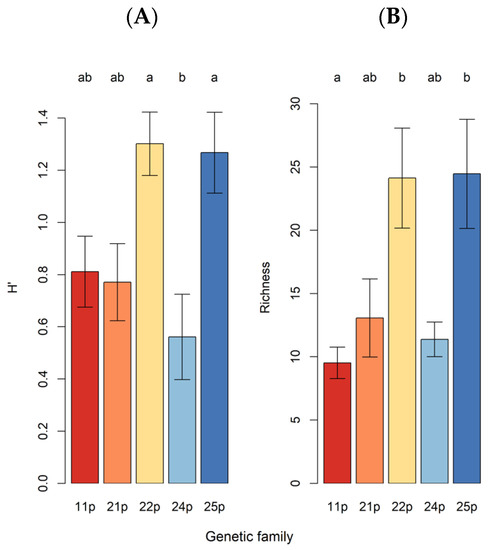
Figure 2.
Shannon diversity (H′; (A)) and OTU richness (Richness; (B)) across maternal families. Bars are means, errors are SEMs. Different alphabetic letters above bars represent gene families with statistically different (α = 0.05) population means after controlling for family-wise type I error using Tukey′s HSD, with pooled variance based on omnibus mixed model estimates for σ2.

Figure 3.
Moran eigenvector map weights projected onto spatial coordinates for seedlings (see Figure 1). Lighter grayscale colors indicate larger positive loadings for seedling coordinates in eigenvectors.
Table 3.
Results of vector (quantitative variables) and factor (categorical) fitting to the NMDS projection (Figure 4), 999 permutations used. Statistically significant results are shown in boldface type.
Based on the stress criterion [82], a three-dimensional NMDS solution (stress = 0.16) was required to adequately describe variation in foliar fungal endophyte OTU communities (Figure 4). Statistically significant (α= 0.05) correlates of the projection are overlaid on the ordination as arrows. Levels for the statistically significant categorical variable maternal family are distinguished with 95% confidence ellipses.
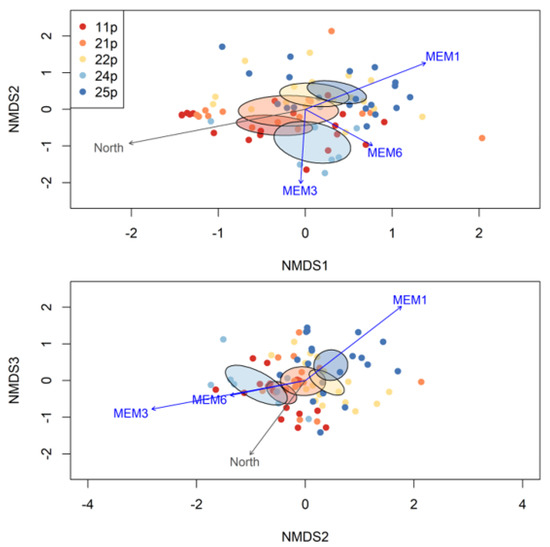
Figure 4.
A 3-dimensional NMDS ordination based on Bray–Curtis dissimilarities (stress = 0.16). Arrows show statistically significant associates of the NMDS projection based on vector fitting analyses. Arrows point in the direction of most rapid increase along projection axes. Arrow length is scaled by the magnitude of R2 statistics (Table 3). Ellipses are 95% confidence ellipses for the true bivariate centroids of maternal family OTU communities within the projection.
Fungal endophyte community composition remained strongly correlated with maternal family after explicitly controlling for spatial proximity (MEMdbs 1-11) in a dbRDA model (F11.62 = 1.61, p = 0.004; Table 3). The cause for this outcome is clear in the variance partitioning analysis shown in Figure 5. Note that although spatial proximity and the interaction of spatial proximity and maternal genetic family explain relatively large proportions of variation in endophyte composition, a non-trivial amount of explained variation can be attributed to genetic family alone (Figure 5a). This trend, however, did not extend to isotopic stoichiometry or carbohydrates. For isotopic stoichiometry (δ15N or δ13C considered jointly), an appreciable amount of variation is explained by the interaction of spatial proximity and genetic family; however, these predictors are almost entirely confounded, and their individual effects are small (Figure 5b, Table 3). For non-structural carbohydrates (glucose, fructose, and sucrose considered jointly), very little variation was explained by the measured predictors (Figure 5c, Table 3).
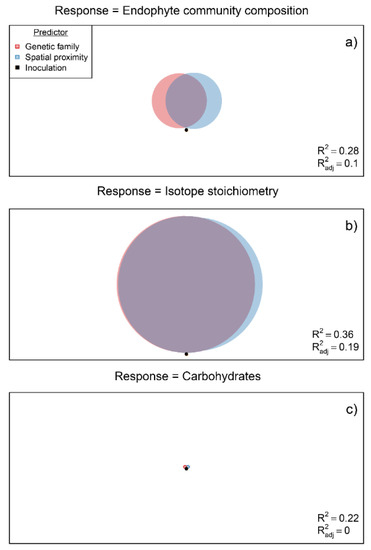
Figure 5.
Variance partitioning analyses for the multivariate responses: (a) endophyte community composition, (b) isotopic stoichiometry, and (c) non-structural carbohydrate composition, with respect to maternal genetic family, inoculation treatment, spatial proximity, and their interaction. Circles, representing predictors, are scaled by their coefficient of partial determination, (i.e., the proportion of explained variation when conditioning on other predictors) in the variance partitioning models. Note that for the treatment was ≈ 0 for all models. The conventional multiple coefficient of determination, (i.e., the proportion variance explained in the model collectively by all the predictors), and the adjusted coefficient of the determination (i.e., adjusted for the spurious effect of additional variables, see the work of Aho (2016)) are shown in the lower right-hand corner of figures.
4. Discussion
The lack of WPBR presence on sampled seedlings led us to discard hypothesis 1, which asserted that fewer endophyte-inoculated seedlings than control seedlings would show signs of WPBR infection. We also rejected research hypotheses 2 and 3, i.e., that host phytochemistry varies with endophytic communities and that the inoculation treatment produced lasting differences in host phytochemistry and endophytic communities. Spatial confounding in the restoration planting trial complicated the interpretation of maternal family results. Nonetheless, given (1) strongly significant results for maternal family effects in univariate mixed effect models and dbRDAs, (2) a lack of variation in mesoclimate across planting locations within the restoration site, (3) the explicit consideration of proximity effects in analyses, and (4) ample prior evidence of correlations between fungal endophyte communities and host genetics [3,4,84,85], we cautiously suggest that our data provide evidence of host genotype-mediated variation in fungal endophyte communities of whitebark pine in their natural habitat, and thus support hypothesis 4.
Seeds from each maternal family were naturally wind-pollinated, and resulting seedlings are thus a mixture of half- and full-sib progeny. Maternal family explained 24% of the variation in the NMDS summary of fungal endophyte communities across whitebark pine genotypes, but only 12% of the variation ( in VPAs. Presumably, this is because the latter approach controlled for spatial proximity.
Maternal families in our trial were previously shown to vary in their susceptibility to C. ribicola. For instance, previous unpublished data revealed that family 22p displayed greater quantitative resistance to C. ribicola infection than all other families, whereas 25p was the most susceptible of the families included in the present study. Although our dbRDAs found that fungal endophyte communities varied significantly across maternal families after conditioning for spatial effects (Table 3), maternal families 22p and 25p were not significantly different in permutational analysis of variance (PERMANOVA [77]) pairwise comparisons after controlling for both spatial effects and family-wise type I error. Significant pairwise differences between other families are summarized in Table S11 in the Supplementary Materials. The lack of community difference between families 22p and 25p may be due to experimental noise introduced by uncontrolled spatial autocorrelation (e.g., spatial variation in wind-dispersed fungal propagules), a lack of importance of WPBR resistance to fungal endophyte community assembly, unmeasured variation in the heritability of quantitative WPBR resistance, or an artifact of the age of the pines we sampled. With respect to the last effect, a microbiome study of young tomato plants found that microbiomes varied across plant genotypes, but that microbiomes were decreasingly distinct among host genotypes as plants aged [86]. Identifying persistent differences in fungal endophyte communities based on host susceptibility to WPBR could be of value to forest protection efforts, but such a trend was not detected in this study or in previous related research [60]. A previous study reported that differences between endophytic communities of resistant and susceptible whitebark pine seedlings only occurred after inoculation with C. ribicola [4]. That study also showed that OTU richness of susceptible genetic families was lower than resistant genetic families following inoculation and that fungal endophyte communities varied with host defensive phytochemistry (terpenes).
Non-structural carbohydrate levels and C and N isotopic stoichiometry did not vary with maternal family after controlling for spatial autocorrelation (Figure 5, Table 3), although maternal family was a significant predictor of phytochemistry when spatial effects were ignored (p = 0.005, Figure S8, Table S1, Supplementary Materials). This discrepancy suggests that unmeasured clumped spatial microgradients of light intensity, soil chemistry, and proximity to surrounding vegetation within our planting site may have influenced plant chemistry. Further study of this trend should be considered in other restoration sites to understand potential drivers of variation in seedling performance, as the heterogeneity we observed in δ13C and δ15N across measured plants suggests that potentially important differences in drought stress may have existed among plants. Differences in plant drought stress may qualitatively change interactions that occur between hosts and endophytes [6]. However, we found a lack of correlation of δ13C and δ15N to either maternal family or inoculation treatment, and a lack of association between endophytic composition and diversity and variation in either non-structural carbohydrates or isotopic signatures. We conclude that host non-defensive phytochemistry was not a driver of variation in fungal endophyte communities across maternal families observed in this study, and conversely that differences in fungal endophyte communities observed across maternal families did not produce variation in phytochemistry.
Whereas our results were negative with respect to our first three hypotheses and represent only a snapshot of the endophyte communities of whitebark pines, this study is the first to investigate drivers of fungal endophyte community variation in the seedlings of whitebark pine in their natural habitat. We note that the lack of difference between microbiomes of inoculated and non-inoculated seedlings may have simply resulted from short-lived effects of inoculation or unsuccessful inoculation. However, as the endophyte inoculation mixture originated from CRLA whitebark pine needles, it is possible that upon outplanting, both inoculated and non-inoculated seedlings became naturally colonized with fungal consortia similar to that represented in the inoculum mix, which would have precluded identification of microbiome differences attributable to inoculation. Nonetheless, among other contributions of the present work, fungal endophyte and phytochemical results described here provide useful baseline data for subsequent studies of the permanent restoration plot that was sampled for this work. To our knowledge, this study is the first to report host-genetic effects on foliar fungal communities of a high-elevation conifer species of imminent conservation concern in a permanent restoration outplanting.
Given that the seedlings in this field trial did not show signs of WPBR infection, the most parsimonious explanation for our lack of detection of C. ribicola is that it indeed was not present in our samples. Similarly, though our methods were adequate for detecting reads of P. roridum in sequence libraries (pers. comm. G. Newcombe), we did not detect the species in either inoculated or control samples despite the fact that the strain of P. roridum from which the inoculant was developed had been cultured from whitebark pines naturally occurring at CLNP [49]. However, the P. roridum cultures obtained from whitebark pine (described in [49]) were reportedly found in only 2 of the 10 trees sampled in that work, so our lack of detection of P. roridum in our samples may be due to the low abundance of that species at CLNP. Additionally, detection of P. roridum in host plants may have been precluded by a lack of systemic establishment of the endophyte in hosts following inoculation such that the inoculant was not present in the cohort of needles that was sampled. However, reports suggest that the longevity of whitebark pine foliage ranges from five to eight years [87].
5. Conclusions
Three conclusions can be drawn from this work: (1) fungal microbiomes of whitebark pine seedlings across our study site did not vary with host phytochemical signatures of ecophysiological status, (2) the inoculation of P. albicaulis seedlings with a mixture of fungal endophytes did not lead to persistent systemic changes in seedling foliar microbiomes, and (3) in correspondence with other studies, our data suggest that maternal seed source and spatial patterns influence fungal endophyte community composition. Progress can be made on related study systems by attempting to re-isolate P. roridum from inoculated host-plant foliage shortly after inoculation and yearly thereafter using both next-generation and culturing techniques, such as those used by the group that isolated the P. roridum strain at CLNP that was used in the present study [49]. Additionally, it is unknown whether differences exist between fungal endophyte communities of naturally occurring seedlings in the wild versus nursery-grown seedlings before and after outplanting, which could indicate an influence of nursery rearing on the microbial biodiversity of restoration sites. It also remains unknown whether there is a long-term microbiome of pines and other perennial plants that grow new foliage annually, and to what extent it is influenced by the microbiome of previous plant tissue versus the microbiomes of surrounding plants. The potential benefit of microbial inoculation of seedlings to serve ecosystem management and restoration objectives will be clarified through a better understanding of the processes involved in plant microbiome assembly. In turn, elucidation of the ecological rules that govern endophytic fungal community characteristics and dynamics can be achieved through continuing work in this and related study systems.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/f13060824/s1, Supplementary Materials Analyses.
Author Contributions
Seedling outplanting study design: R.A.S.; outplanting execution: US Park Service personnel; microbiome and phytochemical study design and execution: E.R.V.M.; sampling design: E.R.V.M. and K.A.; field and lab work: E.R.V.M.; oversight: K.R. and K.A.; statistical analyses: E.R.V.M. and K.A.; manuscript drafting: E.R.V.M.; critical reviews and input: all co-authors. All authors have read and agreed to the published version of the manuscript.
Funding
This research was generously supported by a Graduate Student Research Grant from The Mazamas, the Idaho State University Department of Biological Sciences Molecular Research Core Facility SEED Grant, and the University Research Council at Idaho State University.
Data Availability Statement
Molecular data have been deposited at DDBJ/EMBL/GenBank under accession No. KBTW00000000. All other data used in this study are openly available in FigShare under: Moler, Ehren; Reinhardt, Keith; Sniezko, Richard A.; Aho, Ken (2021): First report of fungal endophyte communities and non-defensive phytochemistry of biocontrol-inoculated whitebark pine seedlings in a restoration planting. figshare. Dataset. https://doi.org/10.6084/m9.figshare.14071448.v1 (accessed on 8 April 2022).
Acknowledgments
We thank Crater Lake National Park personnel including Jen Hooke and Elena Thomas, George Newcombe at the University of Idaho, and personnel at the Dorena Genetic Resource Center. We thank Vern Winston, Carolyn Webber, Scot Kelchner, and the late Timothy Magnuson for consultation regarding molecular techniques. We thank undergraduate research technician Peter Eckert for assistance with DNA extraction and phytochemical analyses.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Appendix A
Sequence Library Summary (Continued)
Leotiomycetes was the most abundant fungal class, accounting for 77% of sequences classified to or below that taxonomic level. Leotiomycetes contains the order Leotiales, which occurred in the greatest abundance in our sequence library and is typically the most abundant order in gymnosperm foliage [88]. The second most abundant order represented 12.4% of classified sequences and is placed in the holding group Ascomycota incertae sedis, exemplifying the large number of presently unidentified taxa uncovered in this study. The most abundant orders found are known to contain plant-associated, largely saprotrophic fungal taxa, such as the Leotiales and Rhytismatales. However, sequences aligning with ≥80% confidence to the Chytridiales, containing many plant pathogens; Teloschistales, containing mostly lichenized species; and even Diaporthales, which are mostly found on angiosperms, were well-represented. The most abundant sequences classifiable to at least the level of genus belonged to class Leotiomycetes, order Leotiales or Rhytismatales. Hyaloscypha sp. was the most abundant classifiable taxon, accounting for 19% of the total reads. The genus has been observed on decaying snags and, like most endophytes, may be a latent saprotroph [89]. Coccomyces multangularis, which has been reported as a saprophyte on fallen leaves in Japan [90], was the second most abundant species (12.6% of all reads). Lophophacidium dooksii, the causal agent of Dooks needle blight in eastern North America [91], was the third most abundant species at 9.3% of the library (megablast = 91% similarity, 3 × 1061). Gyoerffyella entomobryoides, a saprophyte of woody tissue [92], accounted for 9.1% of the 209,034 total reads.
Appendix B
Table A1.
Average phytochemical values of whitebark pine needles. Numbers are means (ranges given in parentheses).
Table A1.
Average phytochemical values of whitebark pine needles. Numbers are means (ranges given in parentheses).
| δ13C (‰) | δ15N (‰) | Glucose (g/g Sample) | Fructose (g/g Sample) | Sucrose (g/g Sample) |
|---|---|---|---|---|
| −29.028 (−30.8–>−25.7) | −0.786 (−2.9–>−2.2) | 0.0083 (0.0009–0.0501) | 0.0096 (0.0–0.0443) | 0.0106 (0.0–0.0788) |
References
- Smith, K.P.; Goodman, R.M. Host variation for interactions with beneficial plant-associated microbes. Annu. Rev. Phytopathol. 1999, 37, 473–491. [Google Scholar] [CrossRef] [PubMed]
- Sthultz, C.M.; Whitham, T.G.; Kennedy, K.; Deckert, R.; Gehring, C.A. Genetically based susceptibility to herbivory influences the ectomycorrhizal fungal communities of a foundation tree species. New Phytol. 2009, 184, 657–667. [Google Scholar] [CrossRef] [PubMed]
- Lamit, L.J.; Busby, P.E.; Lau, M.K.; Compson, Z.G.; Wojtowicz, T.; Keith, A.R.; Zinkgraf, M.S.; Schweitzer, J.A.; Shuster, S.M.; Gehring, C.A.; et al. Tree genotype mediates covariance among communities from microbes to lichens and arthropods. J. Ecol. 2015, 103, 840–850. [Google Scholar] [CrossRef]
- Bullington, L.S.; Lekberg, Y.; Sniezko, R.; Larkin, B. The influence of genetics, defensive chemistry and the fungal microbiome on disease outcome in whitebark pine trees. Mol. Plant Pathol. 2018, 19, 1847–1858. [Google Scholar] [CrossRef]
- Yuan, Z.; Zhang, C.; Lin, F. Role of Diverse Non-Systemic Fungal Endophytes in Plant Performance and Response to Stress: Progress and Approaches. J. Plant Growth Regul. 2010, 29, 116–126. [Google Scholar] [CrossRef]
- Rodriguez, R.J.; Redman, R.S.; Henson, J.M. The Role of Fungal Symbioses in the Adaptation of Plants to High Stress Environments. Mitig. Adapt. Strateg. Glob. Chang. 2004, 9, 261–272. [Google Scholar] [CrossRef]
- Nagabhyru, P.; Dinkins, R.D.; Wood, C.L.; Bacon, C.W.; Schardl, C.L. Tall fescue endophyte effects on tolerance to water-deficit stress. BMC Plant Biol. 2013, 13, 127. [Google Scholar] [CrossRef]
- Rodriguez, R.J.; White, J.F.; Arnold, A.E.; Redman, R.S. Fungal endophytes: Diversity and functional roles. New Phytol. 2009, 182, 314–330. [Google Scholar] [CrossRef]
- Giauque, H.; Hawkes, C.V. Climate affects symbiotic fungal endophyte diversity and performance. Am. J. Bot. 2013, 100, 1435–1444. [Google Scholar] [CrossRef]
- Arnold, A.E.; Mejía, L.C.; Kyllo, D.; Rojas, E.I.; Maynard, Z.; Robbins, N.; Herre, E.A. Fungal endophytes limit pathogen damage in a tropical tree. Proc. Natl. Acad. Sci. USA 2003, 100, 15649–15654. [Google Scholar] [CrossRef]
- Mueller, G.M.; Bills, G.F.; Foster, M.S. Biodiversity of Fungi: Inventory and Monitoring Methods; Elsevier: Amsterdam, The Netherlands; Academic Press: Cambridge, MA, USA, 2004. [Google Scholar]
- Ganley, R.J.; Sniezko, R.A.; Newcombe, G. Endophyte-mediated resistance against white pine blister rust in Pinus monticola. For. Ecol. Manag. 2008, 255, 2751–2760. [Google Scholar] [CrossRef]
- Chaudhry, V.; Runge, P.; Sengupta, P.; Doehlemann, G.; Parker, J.E.; Kemen, E. Shaping the leaf microbiota: Plant–microbe–microbe interactions. J. Exp. Bot. 2021, 72, 36–56. [Google Scholar] [CrossRef] [PubMed]
- Lata, R.; Chowdhury, S.; Gond, S.K.; White, J.F. Induction of abiotic stress tolerance in plants by endophytic microbes. Lett. Appl. Microbiol. 2018, 66, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Alkahtani, M.D.F.; Fouda, A.; Attia, K.A.; Al-Otaibi, F.; Eid, A.M.; Ewais, E.E.-D.; Hijri, M.; St-Arnaud, M.; Hassan, S.E.-D.; Khan, N.; et al. Isolation and Characterization of Plant Growth Promoting Endophytic Bacteria from Desert Plants and Their Application as Bioinoculants for Sustainable Agriculture. Agronomy 2020, 10, 1325. [Google Scholar] [CrossRef]
- Rabiey, M.; Hailey, L.E.; Roy, S.R.; Grenz, K.; Al-Zadjali, M.A.; Barrett, G.A.; Jackson, R.W. Endophytes vs tree pathogens and pests: Can they be used as biological control agents to improve tree health? Eur. J. Plant Pathol. 2019, 155, 711–729. [Google Scholar] [CrossRef]
- Puri, A.; Padda, K.P.; Chanway, C.P. Can naturally-occurring endophytic nitrogen-fixing bacteria of hybrid white spruce sustain boreal forest tree growth on extremely nutrient-poor soils? Soil Biol. Biochem. 2020, 140, 107642. [Google Scholar] [CrossRef]
- Van Oppen MJ, H.; Oliver, J.K.; Putnam, H.M.; Gates, R.D. Building coral reef resilience through assisted evolution. Proc. Natl. Acad. Sci. USA 2015, 112, 2307–2313. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Craine, J.M.; Elmore, A.J.; Aidar, M.P.M.; Bustamante, M.; Dawson, T.E.; Hobbie, E.A.; Kahmen, A.; Mack, M.C.; McLauchlan, K.K.; Michelsen, A.; et al. Global patterns of foliar nitrogen isotopes and their relationships with climate, mycorrhizal fungi, foliar nutrient concentrations, and nitrogen availability. New Phytol. 2009, 183, 980–992. [Google Scholar] [CrossRef]
- Warren, C.R.; McGrath, J.F.; Adams, M.A. Water availability and carbon isotope discrimination in conifers. Oecologia 2001, 127, 476–486. [Google Scholar] [CrossRef]
- Guerrieri, R.; Belmecheri, S.; Ollinger, S.V.; Asbjornsen, H.; Jennings, K.; Xiao, J.; Stocker, B.D.; Martin, M.; Hollinger, D.Y.; Bracho-Garrillo, R.; et al. Disentangling the role of photosynthesis and stomatal conductance on rising forest water-use efficiency. Proc. Natl. Acad. Sci. USA 2019, 116, 16909–16914. [Google Scholar] [CrossRef] [PubMed]
- Mejía, L.C.; Herre, E.A.; Sparks, J.P.; Winter, K.; García, M.N.; Van Bael, S.A.; Stitt, J.; Shi, Z.; Zhang, Y.; Guiltinan, M.J.; et al. Pervasive effects of a dominant foliar endophytic fungus on host genetic and phenotypic expression in a tropical tree. Front. Microbiol. 2014, 5, 479. [Google Scholar] [CrossRef] [PubMed]
- Craine, J.M.; Brookshire, E.N.J.; Cramer, M.D.; Hasselquist, N.J.; Koba, K.; Marin-Spiotta, E.; Wang, L. Ecological interpretations of nitrogen isotope ratios of terrestrial plants and soils. Plant Soil 2015, 396, 1–26. [Google Scholar] [CrossRef]
- Hartmann, H.; Trumbore, S. Understanding the roles of nonstructural carbohydrates in forest trees–From what we can measure to what we want to know. New Phytol. 2016, 211, 386–403. [Google Scholar] [CrossRef]
- Odokonyero, K.; Cardoso Arango, J.A.; de la Cruz Jimenez Serna, J.; Rao, I.M.; Acuña, T.B. Influence of fungal endophyte on plant water status, non-structural carbohydrate content and biomass partitioning in Brachiaria grasses grown under drought stress. In Proceedings of the Building Productive, Diverse and Sustainable Landscapes, 17th Australian Agronomy Conference, Conference Proceedings, Hobart, Australia, 20–24 September 2015; pp. 593–595. [Google Scholar]
- Wiley, E.; Rogers, B.J.; Hodgkinson, R.; Landhäusser, S.M. Nonstructural carbohydrate dynamics of lodgepole pine dying from mountain pine beetle attack. New Phytol. 2016, 209, 550–562. [Google Scholar] [CrossRef]
- Wicker, E.F. Natural Control of White Pine Blister Rust by Tuberculina maxima. Phytopathology 1981, 71, 997–1000. [Google Scholar] [CrossRef]
- Isaac, S. Fungal-Plant Interactions; Springer Science & Business Media: Berlin/Heidelberg, Germany, 1991. [Google Scholar]
- Dickie, I.A.; Fukami, T.; Wilkie, J.P.; Allen, R.B.; Buchanan, P.K. Do assembly history effects attenuate from species to ecosystem properties? A field test with wood-inhabiting fungi. Ecol. Lett. 2012, 15, 133–141. [Google Scholar] [CrossRef]
- Estrada, C.; Wcislo, W.T.; Bael SA, V. Symbiotic fungi alter plant chemistry that discourages leaf-cutting ants. New Phytol. 2013, 198, 241–251. [Google Scholar] [CrossRef]
- Hiscox, J.; Savoury, M.; Muller, C.T.; Lindahl, B.D.; Rogers, H.J.; Boddy, L. Priority effects during fungal community establishment in beech wood. ISME J. 2015, 9, 2246–2260. [Google Scholar] [CrossRef]
- Daba, G.M. Myrothecium as Promising Model for Biotechnological Applications, Potentials and Challenges. BJSTR 2019, 16, 12126–12131. [Google Scholar] [CrossRef]
- Mueller, U.G.; Sachs, J.L. Engineering microbiomes to improve plant and animal health. Trends Microbiol. 2015, 23, 606–617. [Google Scholar] [CrossRef]
- Nogales, A.; Nobre, T.; Valadas, V.; Ragonezi, C.; Döring, M.; Polidoros, A.; Arnholdt-Schmitt, B. Can functional hologenomics aid tackling current challenges in plant breeding? Brief. Funct. Genom. 2016, 15, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Peiffer, J.A.; Spor, A.; Koren, O.; Jin, Z.; Tringe, S.G.; Dangl, J.L.; Buckler, E.S.; Ley, R.E. Diversity and heritability of the maize rhizosphere microbiome under field conditions. Proc. Natl. Acad. Sci. USA 2013, 110, 6548–6553. [Google Scholar] [CrossRef] [PubMed]
- Pandey, R.R.; Arora, D.K.; Dubey, R.C. Antagonistic interactions between fungal pathogens and phylloplane fungi of guava. Mycopathologia 1993, 124, 31–39. [Google Scholar] [CrossRef]
- Rubini, M.R.; Silva-Ribeiro, R.T.; Pomella, A.W.; Maki, C.S.; Araújo, W.L.; Dos Santos, D.R.; Azevedo, J.L. Diversity of endophytic fungal community of cacao (Theobroma cacao L.) and biological control of Crinipellis perniciosa, causal agent of Witches’ Broom Disease. Int. J. Biol. Sci. 2005, 1, 24–33. [Google Scholar] [CrossRef]
- Frankland, J.C. Fungal succession—Unravelling the unpredictable. Mycol. Res. 1998, 102, 1–15. [Google Scholar] [CrossRef]
- Geils, B.W.; Hummer, K.E.; Hunt, R.S. White pines, Ribes, and blister rust: A review and synthesis. For. Pathol. 2010, 40, 147–185. [Google Scholar] [CrossRef]
- Tomback, D.F.; Arno, S.F.; Keane, R.E. Whitebark Pine Communities: Ecology And Restoration; Island Press: Washington, DC, USA, 2001. [Google Scholar]
- Murray, M.P.; Rasmussen, M.C. Non-native blister rust disease on whitebark pine at Crater Lake National Park. Northwest Sci. 2003, 77, 87–91. [Google Scholar]
- Tomback, D.F.; Achuff, P. Blister rust and western forest biodiversity: Ecology, values and outlook for white pines. For. Pathol. 2010, 40, 186–225. [Google Scholar] [CrossRef]
- Smith, C.M.; Shepherd, B.; Gillies, C.; Stuart-Smith, J. Changes in blister rust infection and mortality in whitebark pine over time. Can. J. For. Res. 2013, 43, 90–96. [Google Scholar] [CrossRef]
- FWS. Endangered and Threatened Wildlife and Plants; Threatened Species Status for Pinus albicaulis (Whitebark Pine) with Section 4(d) Rule. Federal Register. 2020. Available online: https://www.federalregister.gov/documents/2020/12/02/2020-25331/endangered-and-threatened-wildlife-and-plants-threatened-species-status-for-pinus-albicaulis (accessed on 12 December 2020).
- Dominguez-Nunez, J.A.; Albanesi, A.S. Ectomycorrhizal fungi as biofertilizers in forestry. Biostimulants Plant Sci. 2019. [Google Scholar] [CrossRef]
- Sniezko, R.A.; Kegley, A.; Danchok, R.; Long, S. Blister rust resistance in whitebark pine (Pinus albicaulis)-early results following artificial inoculation of seedlings from Oregon, Washington, Idaho, Montana, California, and British Columbia seed sources. In Proceedings of the IUFRO Joint Conference: Genetics of Five-Needle Pines, Rusts of Forest Trees, and Strobusphere, 15–20 June 2014; Schoettle, A.W., Sniezko, R.A., Kliejunas, J.T., Eds.; Proc. RMRS-P-76; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2018; Volume 76, pp. 129–135. [Google Scholar]
- Trappe, M.J.; Cromack, K.; Caldwell, B.A.; Griffiths, R.P.; Trappe, J.M. Diversity of Mat-Forming Fungi in Relation to Soil Properties, Disturbance, and Forest Ecotype at Crater Lake National Park, Oregon, USA. Diversity 2012, 4, 196–223. [Google Scholar] [CrossRef]
- Worapong, J.; Sun, J.; Newcombe, G. First report of Myrothecium roridum from a gymnosperm. N. Am. Fungi 2009, 4, 1–6. [Google Scholar] [CrossRef][Green Version]
- Ponnappa, K.M. On the pathogenicity of Myrothecium roridum-Eichhornia crassipes isolate. Hyacinth Control J. 1970, 8, 18–20. [Google Scholar]
- Gees, R. Evaluation of a Strain of Myrothecium roridum as a Potential Biocontrol Agent against Phytophthora cinnamomi. Phytopathology 1989, 79, 1079. [Google Scholar] [CrossRef]
- Ganley, R.J.; Newcombe, G. Fungal endophytes in seeds and needles of Pinus monticola. Mycol. Res. 2006, 110, 318–327. [Google Scholar] [CrossRef]
- Lindahl, B.D.; Nilsson, R.H.; Tedersoo, L.; Abarenkov, K.; Carlsen, T.; Kjøller, R.; Kõljalg, U.; Pennanen, T.; Rosendahl, S.; Stenlid, J.; et al. Fungal community analysis by high-throughput sequencing of amplified markers–A user’s guide. New Phytol. 2013, 199, 288–299. [Google Scholar] [CrossRef]
- Deng, J.X.; Paul, N.C.; Li, M.J.; Seo, E.Y.; Sung, G.H.; Yu, S.H. Molecular Characterization and Morphology of Two Endophytic Peyronellaea Species from Pinus koraiensis in Korea. Mycobiology 2011, 39, 266–271. [Google Scholar] [CrossRef]
- Arnold, A.E.; Lutzoni, F. Diversity and Host Range of Foliar Fungal Endophytes: Are Tropical Leaves Biodiversity Hotspots? Ecology 2007, 88, 541–549. [Google Scholar] [CrossRef]
- Oono, R.; Lefèvre, E.; Simha, A.; Lutzoni, F. A comparison of the community diversity of foliar fungal endophytes between seedling and adult loblolly pines (Pinus taeda). Fungal Biol. 2015, 119, 917–928. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. Available online: https://www.sciencedirect.com/book/9780123721808/pcr-protocols (accessed on 20 March 2020).
- Schmidt, P.-A.; Bálint, M.; Greshake, B.; Bandow, C.; Römbke, J.; Schmitt, I. Illumina metabarcoding of a soil fungal community. Soil Biol. Biochem. 2013, 65, 128–132. [Google Scholar] [CrossRef]
- Moler, E.R.; Aho, K. Whitebark pine foliar fungal endophyte communities in the southern Cascade Range, USA: Host mycobiomes and white pine blister rust. Fungal Ecol. 2018, 33, 104–114. [Google Scholar] [CrossRef]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a Dual-Index Sequencing Strategy and Curation Pipeline for Analyzing Amplicon Sequence Data on the MiSeq Illumina Sequencing Platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [PubMed]
- Huse, S.M.; Welch, D.M.; Morrison, H.G.; Sogin, M.L. Ironing out the wrinkles in the rare biosphere through improved OTU clustering. Environ. Microbiol. 2010, 12, 1889–1898. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- Kunin, V.; Engelbrektson, A.; Ochman, H.; Hugenholtz, P. Wrinkles in the rare biosphere: Pyrosequencing errors can lead to artificial inflation of diversity estimates. Environ. Microbiol. 2010, 12, 118–123. [Google Scholar] [CrossRef]
- Smith, D.P.; Peay, K.G. Sequence Depth, Not PCR Replication, Improves Ecological Inference from Next Generation DNA Sequencing. PLoS ONE 2014, 9, e90234. [Google Scholar]
- Reinhardt, K.; Castanha, C.; Germino, M.J.; Kueppers, L.M.; Pereira, J. Ecophysiological variation in two provenances of Pinus flexilis seedlings across an elevation gradient from forest to alpine. Tree Physiol. 2011, 31, 615–625. [Google Scholar] [CrossRef]
- Amy-Sagers, C.; Reinhardt, K.; Larson, D.M. Ecotoxicological assessments show sucralose and fluoxetine affect the aquatic plant, Lemna minor. Aquat. Toxicol. 2017, 185, 76–85. [Google Scholar] [CrossRef]
- Misarti, N.; Gier, E.; Finney, B.; Barnes, K.; McCarthy, M. Compound-specific amino acid δ15N values in archaeological shell: Assessing diagenetic integrity and potential for isotopic baseline reconstruction. Rapid Commun. Mass Spectrom. 2017, 31, 1881–1891. [Google Scholar] [CrossRef] [PubMed]
- Aho, K.A. Foundational and Applied Statistics for Biologists Using R; CRC Press: Boca Raton, FL, USA, 2013. [Google Scholar]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D. Mixed-Effects Models in S and S-PLUS; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- Akaike, H.; Petrov, B.N.; Csaki, F. Second International Symposium on Information Theory; Akadémiai Kiadó: Budapest, Hungary, 1973. [Google Scholar]
- Aho, K.; Derryberry, D.; Peterson, T. Model selection for ecologists: The worldviews of AIC and BIC. Ecology 2014, 95, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Roberts, D.W. Labdsv: Ordination and multivariate analysis for ecology. R Package Version 2007, 1, 68. [Google Scholar]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian Classifier for Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy. AEM 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L.F. Numerical Ecology; Elsevier: Amsterdam, The Netherlands, 2012; Volume 24. [Google Scholar]
- Borcard, D.; Legendre, P.; Drapeau, P. Partialling out the Spatial Component of Ecological Variation. Ecology 1992, 73, 1045–1055. [Google Scholar] [CrossRef]
- Legendre, P.; Anderson, M.J. Distance-Based Redundancy Analysis: Testing Multispecies Responses in Multifactorial Ecological Experiments. Ecol. Monogr. 1999, 69, 1–24. [Google Scholar] [CrossRef]
- Peres-Neto, P.R.; Legendre, P.; Dray, S.; Borcard, D. Variation partitioning of species data matrices: Estimation and comparison of fractions. Ecology 2006, 87, 2614–2625. [Google Scholar] [CrossRef]
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest communities of southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Kruskal, J.B. Multidimensional scaling by optimizing goodness of fit to a nonmetric hypothesis. Psychometrika 1964, 29, 1–27. [Google Scholar] [CrossRef]
- Oksanen, J.; Simpson, G.L.; Guillaume Blanchet, F.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Vegan: Community Ecology Package. R Package Version 2.5-6. 2019. Available online: http://cran.rproject.org/package=vegan (accessed on 20 March 2020).
- Bálint, M.; Bartha, L.; O’Hara, R.B.; Olson, M.S.; Otte, J.; Pfenninger, M.; Robertson, A.L.; Tiffin, P.; Schmitt, I. Relocation, high-latitude warming and host genetic identity shape the foliar fungal microbiome of poplars. Mol. Ecol. 2015, 24, 235–248. [Google Scholar] [CrossRef] [PubMed]
- Gehring, C.A.; Sthultz, C.M.; Flores-Rentería, L.; Whipple, A.V.; Whitham, T.G. Tree genetics defines fungal partner communities that may confer drought tolerance. Proc. Natl. Acad. Sci. USA 2017, 114, 11169–11174. [Google Scholar] [CrossRef]
- Morella, N.M.; Weng, F.C.-H.; Joubert, P.M.; Metcalf, C.J.E.; Lindow, S.; Koskella, B. Successive passaging of a plant-associated microbiome reveals robust habitat and host genotype-dependent selection. Proc. Natl. Acad. Sci. USA 2020, 117, 1148–1159. [Google Scholar] [CrossRef] [PubMed]
- Schoettle, A.W.; Fahey, T.J.; Shoettle, A.W. Foliage and Fine Root Longevity of Pines. Ecol. Bull. 1994, 43, 136–153. [Google Scholar]
- Sieber, T.N. Endophytic fungi in forest trees: Are they mutualists? Fungal Biol. Rev. 2007, 21, 75–89. [Google Scholar] [CrossRef]
- Käärik, A. Succession of microorganisms during wood decay. In Biological Transformation of Wood by Microorganisms; Springer: Berlin/Heidelberg, Germany, 1975; pp. 39–51. [Google Scholar]
- Yingren, L.; Zengzhi, L.; Zaoshi, X.; Jingru, W.; Shengming, Y. Studies on the genus Coccomyces from China IV. Jun Wu Xi Tong Mycosystema 2001, 20, 1–7. [Google Scholar]
- Mclaughlin, J.A.; Lu, P.; Greifenhagen, S.; Wilson, R. New hosts and differential susceptibility of five-needle pine species to Dooks needle blight (Lophophacidium dooksii). Can. J. Plant Pathol. 2012, 34, 536–544. [Google Scholar] [CrossRef]
- Mäkelä, K. The fungi of wintered branches of outdoor roses in Finland. Ann. Agric. Fenn. 1986, 25, 187–197. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).